Infectious bursitis virus (IBDV) causes an acute and highly contagious disease in young chickens, which is responsible for significant losses in the poultry industry worldwide. VP4 cDNA was produced and amplified by optimizing a reverse transcription coupled to polymerase chain reaction (RT-PCR), providing a clear and sensitive assay. Anti-peptide antibodies were raised against a selected peptide from VP4 and used to probe homogenates of infected bursae for the native protein to assess their potential for immunological detection.
Double basic amino acid cleavage sites were suggested for the putative protease and infected samples were tested for activity against the fluorogenic peptide Cbz-Arg-Arg-AMC.
Viral characteristics .1 Morphology
- IBDV proteins
- Pathology and lesions
VPX (also known as pre-VP2, pVP2, or VP2a) undergoes a second, independent and as yet undefined proteolytic processing step to produce VP2 (−40 kDa) ( Kibenge et al., 1997 ). These two residues are conserved in the VP4 sequences of IPNV, all strains of IBDV and DXV (Lejal et al., 2000). This protein is associated with the differentiation phase of B cells characterized by the retention of surface IgM (Ogawa et al., 1998).
Destruction of immature B-cells in the bursa causes immunosuppression, which is more severe in younger birds (Faragher et al., 1972).
Disease diagnosis
- Clinical and histological diagnosis
- Vaccination
ELISAs using a recombinant VP2 protein as the sole antigen may be better correlated with protection (Jackwood et al., 1999). Different batteries of mAbs allow a tentative identification of the American variants (Snyder et al., 1988) and vvIBDVs (Eterradossi et al., 1997). However, they remain vulnerable to highly pathogenic strains that cause high mortality at later stages (van den Berg et al., 1991).
VP5 also appears to play a role in the release of virion progeny (Lombardo et al., 2000).
Proteases
- Protease classification and catalytic mechanisms
- Protease significance in viral infection
This results in the formation of another tetrahedral transition state and subsequent cleavage of the acyl bond. In this case, the attacking nucleophile is the active site sulfur atom Cys25 (papain numbering). Therefore, the glutamic acid acts as a 'shuttle', analogous to one of the catalytic groups in the aspartic acids and the histidine in the serine and cysteine plugs. e).
As described in the previous section, the study of the putative protease of IBDV holds much potential.

General materials and methods
Molecular Biology
Assaying for protein
- Reagents
- Effect of SDS
Protein concentration of unknown samples was calculated using an equation generated by linear regression analysis of a standard curve. The porosity of the gel depends on the relative amount of cross-linking agent relative to the total acrylamide (%C) and on the total mass of acrylamide in the gel volume (%T). The pH in the electrode compartments and the stacking gel is chosen so that the proteins of interest have an intermediate mobility between fast and slow counterions.
When a focused group of proteins reaches the separation gel, the change in pH causes an increase in the mobility of the tracer ion. The smaller pore structure of the separating gel now imposes a seeding effect on the migrating proteins and the separation achieved depends on the charge and size of the protein molecules as well as the concentration of the separating gel. The inclusion of a preincubation step with sodium thiosulfate (Blum et al., 1987) significantly reduces background staining by preventing precipitation of insoluble silver complexes on the gel surface.
Gels are then incubated in appropriate assay buffers, facilitating digestion of the incorporated substrate by immobilized proteases. The molecular mass of protease can be estimated, but not with particular accuracy, as incorporation of the protein substrate increases the apparent molecular mass and does so in a heterogeneous manner. Gels were then incubated in assay buffer (ON, 37°C) to allow digestion of the incorporated substrate and then stained with amido black for 1 hour.
The concentration of PEG in the clear filtrate was then increased to 12% (rn/v) by the addition of 8.5% (rn/v) solid PEG dissolved by gentle stirring. To ensure gentle resuspension, the antibody pellet was removed from the walls of the centrifuge tube with a glass rod and allowed to dissolve slowly in sodium phosphate buffer (1/6 of the original yolk volume).

Isolation of rabbit antibodies from serum
- Reagents
- Procedure
Determination of antibody concentration
When one immunoreactant is immobilized on a solid phase to facilitate separation from free antigen or antibody, EIA is known as ELISA (Engvall and Perlmann, 1971). ELISA assays can be structured to be competitive or non-competitive and in both systems either antigen or antibody can be immobilized on a solid phase. In this study, non-competitive immobilized antigen ELISAs were used to evaluate the progress of anti-peptide antibody production during immunization procedures and to detect the presence of VP4 protein in crude bursa fractions.
Briefly, the well surfaces of polystyrene microtiter plates are coated with antigen and the uncoated surfaces are blocked with an unrelated protein before being exposed to a solution of antigen-specific primary antibody. Secondary antibodies, which usually develop against the IgG type of the primary antibody, detect the primary antibody in the next step. Conjugated enzymes have been chosen so that they can be readily assayed and will typically catalyze the formation of a soluble colored product from a colorless substrate that can be quantified spectrophotometrically.
A variety of soluble chromogenic substrates are available for HRPO, one of the most sensitive being 3,3',5,5'-tetramethylbenzidine (TMB), which was used in this study. PBS-BSA and PBS-casein are both blocking agents and have been used for certain applications when necessary. Nunc Immuno Maxisorp F96 multi-well plates were coated with antigen (150 µl, ON at 4°C) and all uncoated areas were blocked with blocking agent (200 µl, 1 hour at 3°C).
Plates were washed with PBS-Tween®. 3x) before adding appropriate dilutions of primary antibodies in blocking agent (100 ul, 2 hours at 37°C). Plates were washed with PBS-Tween®(3x) before addition of HRPO-linked secondary antibodies (diluted with blocking agent, 120 µl, 1 hour at 37°C) and again with wash.
Western blotting onto nitrocellulose membranes
- Reagents
- Procedure
The protein mixtures were separated by Tris-Tricine SDS-PAGE (Section 3.4.5) and the gels were rinsed with blotting buffer. On one side of the insert, a sandwich was assembled as follows: one piece of Scotchbrite'" foam, three pieces of Whatman No. Care was taken not to trap air bubbles between the layers, especially between the nitrocellulose and the gel.
The cartridge was sealed and inserted into the blotting apparatus (pre-filled with blotting buffer) with gel at the cathode and nitrocellulose at the anode. The apparatus was maintained at 8°C by a circulator in the refrigerator, and the buffer was stirred by a magnetic stirrer to maintain uniform conductivity and temperature during transfer. After protein electrotransfer, nitrocellulose was transiently stained with Ponceau S (30 sec) and rinsed with dH2O to reveal the positions of M standards.
These were marked with a pencil and the stain was completely decolorized by dropwise addition of 500 mM NaOH to the dH2O. Unoccupied protein binding sites were blocked by incubating the nitrocellulose in blocking reagent Clh at RT). The blot was washed with TBS (3 × 5 min) and incubated with an appropriate dilution of primary antibody in blocking reagent (2 h at room temperature).
The blot was washed again with TBS (3 x 5 min) and incubated with secondary antibody suitably diluted with blocking reagent Clh at room temperature. TBS (3 x 5 ml), substrate was added and the color forming reaction was allowed to continue until dark bands were visible against a light colored background.
Introduction
- Reagents
- Procedure
- Reagents
- Inhibition of activity against Cbz-Arg-Arg-AMC
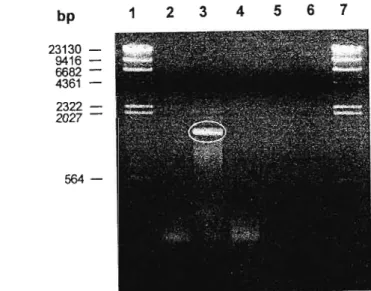
When appropriate, aliquots of the nucleic acid fractions were loaded onto agarose gels (section 2.6.6) to assess purity and concentration. These regions appear to be highly conserved among the virus strains found around the world. The last consideration affects the number of foreign amino acids added to the N-terminus of the recombinant protein.
This is in contrast to the uniform orange fluorescence of the samples stained with EtBr. Similarly, it was not possible to confirm the digestion of the two ends of the RT-PCR product. This suggests that some of the RT-PCR product remained blunted despite the double digestion.
Thus, anti-peptide antibodies share some of the advantages of both anti-protein antibodies and monoclonal antibodies. The A280 eluate was monitored and the activated carrier was collected as the first peak. Only some of these mimic native conformations in the corresponding restricted region of the target protein.
The exclusive presence of the -27 kDa band in uninfected samples (Figure 5-8B) is not easily explained. Interestingly, none of the bands detected in this fraction are actually visible in the corresponding silver stain, Figure 5-7. Proteolytic cleavage of the fluorogenic peptide was assayed in IBOV-infected and uninfected bursal homogenate fractions A to E (Section 5.2), as described in Section 5.4.2.
Inhibition of proteolytic cleavage of the fluorogenic peptide was tested in IBOV-infected and uninfected fractions of bursal homogenates A to E (Section 5.2) as described in Section 5.4.3.
General Discussion
- Significance and implications of results
- RT -rca of IBDV VP4
Restriction analysis of the PCR-amplified VP4 cDNA ultimately showed that the KZN strain appeared to be unique and that VP4 remained unchanged from its inception in 1989 to 1997. RFLPs showed that the VP4 of the KZN strain was most similar to the vv IBDV from the United Kingdom, UK661, both of which are closest in lineage to the vaccine strain PBG98. It should be emphasized again that the possibilities suggested by the RFLP analysis await confirmation and further clarification by sequencing the PCR product.
This involved analysis of the VP4 amino acid sequence and selection of a suitable antigenic peptide (Section 5.3.1). Obviously, it is difficult to assess the quality of anti-VP4 peptide antibodies within such crude samples without testing them with purified VP4. It is also possible that the sequence may reveal the reasons why cloning of the amplified VP4 fragment has proved difficult.
These results call into question the utility of the synthetic Cbz-Arg-Arg-AMC substrate in the current study and emphasize the need for purification of VP4, which can subsequently be tested against a battery of peptide substrates. It is likely that the virulence will be associated with changes in the coat proteins, as has been the case in other parts of the world. This study represents an encouraging initial investigation of the KZN strain of IBDV, which should be pursued further for the sake of locally relevant diagnostics, vaccines and control of a commercially important pathogen.
The characterization and molecular cloning of the double-stranded RNA genome of an Australian strain of infectious bursal disease virus. The infectious bursal agent and pathology of the disease. 1969) Characterization of the infectious bursal agent.