RESPONS KARAKTER FISIOLOGI
DAN PRODUKSI LATEKS
BEBERAPA KLON TANAMAN KARET
TERHADAP STIRlULASI ETILEN
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
y a
A u u
jadjkan aku
sebagai orang
yang selalu bersyukur
. . .
untuk yang tercinta Susi isterku,yang selalu membangun
smnguiku;
yang tersayang amkumkku, Pam, Annis, lpu/& M n ; yang senantiasa
ABSTRAK
RESPONS KAMKTER FISIOLOGI DAN PRODUKSI LATEKS
BEBERAPA KLON TANAMAN
KARET
TERHADAP STIMULASI ETILEN
smmmmn
Pembimbing: Sudirman Yahya, Achmad Surkati Abidin, Alex Hartana, Bambang Sapta Purwoko, dan Siswanto
Penelitian bertujuan untuk mengetahui variasi musiman karakter fisiologi tanaman karet akibat pemberian etepon, dan hubungan antar karakter fisiologi dengan produksi lateks. Penelitian dilaksanakan di Bogor, Jawa Barat (250 m dpl) mulai September 1997 hingga Desember 1998. Percobaan dalam dua faktor perlakuan, yaitu delapan klon karet (AVROS 2037, BPM 24, PR 261, RRIM 712, GT 1, BPM 1, PR 228, dan LCB 1320) dan tiga sistem eksploitasi (& = '/z S d/2, El = '/z
S d/3.ET2.5%, E2 = '/z S d/3.ETS.O%). Peubah yang diarnati adalah produksi lateks,
indeks penyumbatan, kadar karet kering, %-KAS, pH lateks, kadar sukrosa, kadar fosfat anorganik, kadar tiol, aktivitas SOD, produksi etilen, kadar ACC, aktivitas
ACC-oksidase; dan protein dalam fraksi serum-C dan lutoid Hasil penelitian
menunjukkan bahwa perlakuan stimulasi meningkatkan produksi lateks pada empat klon (PR 261, BPM 24, PR 228, dan AVROS 2037). Produksi lateks yang tinggi
ditandai oleh indeks penyumbatan
dan
kadar karet kering yang rendah, serta kadarfosfat anorganik, produksi etilen, dan kejadian KAS yang tinggi. Klon LCB 1320 dan RRLM 712 lebih peka terhadap gangguan KAS. Dari semua klon yang diuji, pengaruh stimulan terhadap penurunan kadar sukrosa hanya nyata pada AVROS 2037; sedangkan kadar ti01 tinggi hanya pada PR 261 sebagai klon yang paling responsif etepon. Kadar ti01 dan aktivitas SOD diduga dapat menghambat mekanisme penuaan dan kejadian KAS. Variasi musiman mempengaruhi semua peubah yang diamati, terutama pada sukrosa, tiol, dan fosfat anorganik. Pada siklus setahun terdapat pola peningkatan produksi lateks, kadar karet kering, dan fosfat anorganik; sedangkan indeks penyumbatan dan pH lateks cenderung menurun. Perlakuan etepon pada klon
AVROS 2037 dan BPM 24 meningkatkan produksi etilen secara langsung, atau
melalui metabolisme etilen dengan ditandai oleh peningkatan kadar ACC. Puncak produksi etilen terjadi 1 hari SAE (setelah aplikasi etepon), sedangkan puncak produksi lateks baru terjadi pada 8 hari SAE. Hasil elektroforesis (SDS-PAGE) mendapatkan protein responsif etilen 27 kDa pada serum-C yang diduga berhubungan dengan influks air dalarn sel pembuluh lateks; serta protein-protein 30,35,37,40, 56, 70, dan 88 kDa pada fraksi lutoid yang diduga berhubungan dengan mekanisme koagulasi lateks. Dengan elektroforesis-2D dari fiaksi ini diperoleh protein 35 dan 70 kDa dengan pHi 5.8.
ABSTRACT
RESPONSE OF PHYSIOLOGICAL CHARACTERS
AND
LATEX PRODUCTION TO
ETHYLENE
STIMULATIONON SEVERAL RUBBER TREE CLONES
SUMARMADJI
Advisors: Sudirman Yahya, Achmad Surkati Abidin, Alex Hartana, Bambang Sapta Purwoko, and Siswanto
The objectives of the study were to determine the seasonal variation of physiological characters of hevea by ethephon application, and the relationship among physiological characters influencing latex production. The experiment was carried out in Bogor, West Java (250 m above sea level) from September 1997 to December 1998. It was designed with two factors of treatment, which were eight hevea clones (AVROS 2037, BPM 24, PR 261, RRIM 712, GT 1, BPM 1, PR 228, and LCB 1320) and three exploitation systems (& = "A S d12, El = "A S d/3.ET2.5%,
ES = % S d/3.ET5.0%). Observed variables were latex production, plugging index,
dry rubber content, %-TPD, latex pH, contents of sucrose, thiol and inorganic phosphate, ethylene release, ACC content, activity of ACC-oxidase and SOD; and proteins in C-serum and lutoid fraction. The results showed that the stimulant increased the latex production in four clones (PR 26 1, BPM 24, PR 228, and AVROS 2037). The high latex production possessed low plugging index and dry rubber content, and high inorganic phosphate content, ethylene release, and incidence of TPD. Potentially, LCB 1320 and RRIM 712 were the most susceptible clones to incidence of TPD. In all clones tested, the significant effect of stimulant to sucrose content was found only in AVROS 2037. Otherwise, the highest thiol content was found in PR 261 as the most ethephon responsive clone. The results also suggested that thiol and SOD activity could inhibit the senescence process and incidence of
TPD. Seasonal variation had influenced to all observed variables, mostly to sucrose, thiol, and inorganic phosphate. When the latex production, dry rubber content and inorganic phosphate content increased, on contrary the plugging index and latex pH decreased. Ethephon treatments in AVROS 2037 and BPM 24 induced the production of ethylene directly, and also endogenously, by increasing ACC content. Ethephon treatment could induce maximal production of ethylene at one day after the application, whereas that of latex occurred at the eighth day. By electrophoresis
(SDS-PAGE), an ethylene responsive protein of 27 kDa was found in C-serum. This
protein probably has relationship with water influx in laticiferous cells. Otherwise, proteins 30,35,37,40, 56,70, and 88 kDa was found in the lutoid, which may have relationship with mechanism of latex coagulation. By 2D-electrophoresis, proteins of 35 and 70 kDa with pHi 5.8 were found in this fraction.
RESPONS KARAKTER FISIOLOGI
DAN PRODUKSI LATEKS
BEBERAPA KLON TANAMAN KARET
TERHADAP STIMULASI ETILEN
Disertasi sebagai salah satu syarat untuk memperoleh gelar
Doktor
pada
Program Pascasarjana Institut Pertanian Bogor
PROGRAM PASCASARJANA
a
INSTITUT PERTANIAN BOGOR
Judul Disertasi
:RESPONS KARAKTER FISIOLOGI DAN
PRODUKSI LATEKS BEBERAPA KLON
TANAMAN KARET
TERHADAP
STIMULASI ETILEN
N a m a
SUMARMADJI
N I M
95526
Program
Studi
:AGRONOMI
Komisi Pembimbing
.
-
I-
&
Dr. Ir. Sudirman Yahya, MSc
Anggota
Anggota h g g o t a
Ketua Direktur
Program Studi Agronomi Program Pascasarjana
JY&=
Dr. Ir. Sudirman Yahya, MSc afrida Manuwoto
Penulis dilahirkan di Rembang, Jawa Tengah tanggal 20 Mei 1959 sebagai anak ke tujuh dari delapan bersaudara hasil perkawinan bapak M. Roestarnadji dan
ibu Kartidjah. Kedua orang tua kini telah almarhum.
Pada tahun 1983 penulis lulus dari Fakultas Pertanian Institut Pertanian Bogor
dan menyandang gelar Sarjana Pertanian. Pada tahun 1989 penulis menyelesaikan
program Magister di Fakultas Pascasa jana, Institut Pertanian Bogor dan menyandang
gelar Magister Sains. Kemudian mulai tahun 1995 penulis berkesempatan mengikuti
pendidikan S3 (Doktor) di Program Pascasarjana, Institut Pertanian Bogor.
Sejak tahun 1984 hingga sekarang, penulis bergabung dengan Pusat Penelitian
Karet di Sungei Putih, Sumatera Utara; sebagai staf peneliti dalam bidang agronomi,
eksploitasi dan usahatani tanaman karet. Penulis telah menikah pada tahun 1986 dengan Susiana, dan sekarang telah dikaruniai dua orang anak laki-laki dan dua orang
anak perempuan; masing-masing Arief Pambudi (12 tahun), Annis Dwilestari (9
UCAPAN TERIMA KASIH
Puji syukur penulis panjatkan ke hadirat Allah SWT atas limpahan rahrnat,
hidayah dan karunia-Nya, sehingga penulis dapat menyusun laporan penelitian untuk
menyelesaikan program S3 dalam bentuk disertasi ini.
Kepada Komisi Pembimbing, penulis menyampaikan terima kasih dan
penghargaan yang talc terhingga khususnya kepada Dr. Ir. Sudirman Yahya, MSc
selaku ketua; Prof. Dr. Ir. H. Achmad Surkati Abidin, 'Dr. Ir. Alex Hartana, MSc, Dr.
Ir. Bambang S. Purwoko, MSc dan Dr. Siswanto, DEA masing-masing selaku
anggota, atas birnbingan dan pengarahan selama penyusunan rencana dan
pelaksanaan penelitian serta penulisan disertasi.
Ucapan terima kasih penulis sampaikan kepada Pimpinan Asosiasi Penelitian
Perkebunan Indonesia (AP21), Pusat Penelitian Karet dan Pimpinan Bagian Proyek
Penelitian Karet Sungei Putih atas kesempatan bagi penulis mengikuti pendidkan S3,
bantuan beasiswa dan dana penelitian selama pendidikan.
Pada kesempatan ini penulis juga menyampaikan terima kasih kepada:
1. Pimpinan beserta staf Institut Pertanian Bogor (IPB) dan Program Pascasarjana
IPB yang telah memberikan kesernpatan dalam pendidikan S3.
2. Pimpinan beserta staf Unit Penelitian Bioteknologi Perkebunan Bogor (UPBP)
yang telah memberikan fasilitas laboratorium dan kebun percobaan, khususnya
teknis laboratorium; serta Sdr. Abad dan Sdr. Mica yang membantu pelaksanaan
dan pengurnpulan data lapangan.
3. Kepala Balai Penelitian Teknologi Karet (BPTK) Bogor beserta staaaryawan
atas bantuan dan kemudahan administratif berhubungan dengan Pusat Penelitian
Karet selama penulis berdomisili di Bogor.
4. Kepala Laboratoriurn Biokimia & Enzimatik, Balitbio Tanaman Pangan beserta
karyawan khususnya Sdr. Eman Sulaiman dan Sdr. Danuwarsa yang membantu
analisis metabolisme etilen.
5. Semua pihak yang telah membantu penulis dalam pendidikan S3 di IPB.
Khusus kepada isteri dan anak-anak tercinta, yang dengan sabar dan penuh
pengertian, senatiasa memberi dorongan moril, membangun semangat dan
menumbuhkan inspirasi, maka dengan ini penulis menyarnpaikan terima kasih yang
tulus dan mendalam.
Akhirnya dengan disertai do'a dan harapan, kiranya hasil penelitian ini
DAFTAR SINGKATAN ACC ACC-oksidase AOS AP2I AVROS BO-2 Ba BB BM BN BPM CEPA CPO dl2, dl3 2D 2,4-D DAL DMAPP dpl DTNB
Eo
E 1
E2 EDTA EFE ERE EREBPS ET FA W s Gcm GACC G-SH GT HEVER IEF
1 -arninocyclopropane-1 -carboxylic acid, prekursor etilen
enzim oksidase terhadap ACC untuk menjadi etilen
active oxygen species, molekul oksigen toksik, senyawa radikal asosiasi penelitian perkebunan Indonesia
algemene veriniging rubber planters oostkust Sumatera kulit perawan bidang sadap kedua
bark application, aplikasi stimulan pada kulit bidang sadap brown bast, KAS, TPD
berat molekul
bark necrosis, gangguan bidang sadap karet yang bersifat patogenik balai penelitian Medan
2-chloroethyl phosphonic acid, etepon, Ethel@ crude palm oil, minyak sawit kasar
frekuensi sadap 2 hari sekali, 3 hari sekali 2 dimensi pada gel elektoforesis
2,4-dichlorophenoxyacetic acid (suatu auksin sintetik) daerah aliran lateks
dimetil alil pirofosfat di atas permukaan laut dithiobis-nitrobenzoat
perlakuan sistem eksploitasi pada percobaan: !4 S d/2
perlakuan sistem eksploitasi pada percobaan: ?4 S d/3.ET2.5%
perlakuan sistem eksploitasi pada percobaan: !4 S d/3.ET5.0%
etilen diamin tetra acetic acid
ethylene forming enzyme, enzim pembentuk etilen
ethylene responsive element, sekuens DNA berkorespondensi etilen protein pengikat sekuens DNA berkorespondensi etilen
sistem eksploitasi dengan aplikasi etepon fosfat anorganik, inorganic phosphate
gram (satuan massa), atau gravitasi (satuan dalam sentrifugasi) groove application, aplikasi stimulan pada alur sadap
satuan produksi lateks gram/pohon/sadap gas chromatographylflame ionization detector T-glutamil-ACC
glutation Gondang Tapen
hevea ethylene responsive, nama gen karet yang responsif etilen isoelectric focussing, pemisahan protein berdasarkan gradien pH
IP indeks penyumbatan, parameter aliran/koagulasi lateks
KAS kDa kgihalth kg/p/th KKK LAH LCB m MACC MGD
MH
MK mmm M
MnSOD MVA NBT nrn PAGE PH pHi PIPIR
PLD PR PTPN rpm RRIM R-SH % S SAE SAM SDS Serum-C SOD 2,4,5-T TCA TNB TPD UJGD UPBPkekeringan alur sadap, penyakit fisiologis pada bidang sadap karet kilo dalton, satuan berat molekul
satuan produksi lateks: kilogram per ha per tahun
satuan produksi lateks: kilogram per pohon per tahun
kadar karet kering (%)
lipolitik ail-hidrolase land caoutchouc bedrijven meter (satuan panjang) malonil-ACC
musim gugur daun karet, terjadi setahun sekali selama 2 3 bulan musim hujan
musim kemarau
milimeter (satuan panjang) satuan kadar senyawa: milimolar
gen SOD yang berikatan dengan
Mn
asam mevalonat nitroblue tetrazolium
mikrometer (satuan panjang), 1
o6
m nanometer (satuan panjang), 1o - ~
m polyacrilamide gel electrophoresis-
logm
pH pada titik isoelektrik
plugging index, indeks penyurnbatan perusahaan inti rakyat
fosfolipase-D
proefstation voor rubber
perseroan terbatas perkebunan nusantara rotation per minute (putaran per menit) rubber research institute of Malaysia tiol, gugus R dengan sulfhidril
'/Z spiral batang karet yang disadap setelah aplikasi etepon
S-adenosyl-L-methionine, Adomet, prekursor ACC sodium dodecyl sulphate
cairan sel, sitosol sel pembuluh lateks superoksida dismutase
2,4,5-trichlorophenoxyacetic acid (suatu auksin sintetik) asam trikloro asetat
thio-nitrobemoat
tapping panel dryness, KAS
' ujibeda jarak berganda Duncan
DAFTAR IS1
Halaman
DAFTAR TABEL
...
x...
DAFTAR GAMBAR
PENDAHULUAN ...
...
Latar Belakang
...
Perurnusan Masalah
...
Tujuan dan Manfaat Penelitian
...
Hipotesis
...
TINJAUAN PUSTAKA
Botani Tanaman Karet
...
Produksi Lateks Tanaman Karet ...
Pemberian Etepon ...
Biosintesis Etilen Tanaman ...
...
Protein Responsif Etilen padaTanaman Karet
...
Identifikasi Protein dengan Elektroforesis .
.
...
Karakter F~s~ologi
Kekeringan Alur Sadap (KAS)
...
BAI-IAN DAN METODE ...
Tempat dan Waktu
...
Bahan dan Alat ...
...
Metode Penelitian
...
HASIL
Kondisi Iklim dan Musim Gugur Daun
...
Hasil Analisis Statistik
...
Produksi Lateks (glp/s) ...
...
Kadar Karet Kering (KKK)
Indeks Penyurnbatan (IP)
...
Kekeringan Alur Sadap (KAS)
...
pH Lateks
...
...
Kadar Sukrosa
...
Kadar Fosfat Anorganik (FA)
Kadar Ti01
...
Aktivitas Superoksida Dismutase (SOD)
...
. .
[image:158.568.42.502.9.772.2]Produksi Etilen
...
...
Kadar ACC
Aktivitas ACC-oksidase ...
Pola Pita Protein
...
...
PEMBAHASAN
...
Produksi Lateks
...
Karakter Fisiologi dan Kaitan dengan Produksi Lateks
...
Variasi Musiman Produksi Lateks dan Karakter Fisiologi
...
Kejadian KAS sebagai Tolok Ukur Eksploitasi Berlebih
...
Sistem Eksploitasi yang Diskriminatif
...
Metabolisme Etilen dalam Jaringan Kulit
Protein Responsif Etilen pada Dua Fraksi Lateks ...
...
KESIMPULAN DAN SARAN
DAFTAR
TABEL
Nomor Halaman
1
.
Sifat beberapa klon skala besar (dalam nilai skor)...
222
.
Musim gugur daun tahun 1998 masing-masing klon karet danpengaruh perlakuan stimulasi pada sistem eksploitasi El dan E2
...
40...
3
.
Pengaruh klon dan sistem eksploitasi terhadap produksi lateks 444
.
Pengaruh klon dan sistem eksploitasi terhadap kadar karet kering...
46 5.
Pengaruh klon dan sistem eksploitasi terhadap indeks penyumbatan ... 49...
6
.
Pengaruh klon dan sistem eksploitasi terhadap kekeringan alur sadap 517
.
Pengaruh klon dan sistem . eksploitasi terhadap kadar sukrosa lateks...
53 8 . Pengaruh klon dan sistem eksploitasi terhadap kadar fosfatanorganik lateks
...
-55 9.
Pengaruh klon dan sistem eksploitasi terhadap kadar ti01 lateks ... 58...
10
.
Pengaruh klon terhadap aktivitas enzim superoksida dismutase 601 1
.
Pengaruh sistem eksploitasi terhadap aktivitas ACC-oksidase 2 klonkaret
...
6312
.
Pola pita protein yang muncul khas pada SDS-PAGE fraksi serum-Cbeberapa klon dengan perlakuan
by
El dan EZ...
-64 13.
Penyebaran pita protein dari fiaksi lutoid masing-masing klondengan perlakuan eksploitasi
...
... ...
6614
.
Kesesuaian sistem eksploitasi masing-masing klon berdasarkan...
Lampiran
Nomor Halaman
1. Perhitungan produksi kg/ha/tahun pada perlakuan klon dan sistem
eksploitasi; serta respons masing-masing klon terhadap etepon ... 109
2. Tetua masing-masing klon, perkiraan jumlah pohodha dan lilit
batang tanaman yang digunakan dalam penelitian (umur 1 1 tahun) ... .I10
3. Kecepatan awal(5 menit pertama) aliran lateks pada faktor tunggal
klon (4 jenis) dan sistem eksploitasi Eo, El dan Ez ... 110
4. Analisis ragam: produksi lateks (g/p/s), kadar karet kering (KKK),
indeks penyumbatan
(IP),
kadar sukrosa, kadar fosfat anorganik(FA), dan kadar ti01 ... 11 1
...
5. Analisis ragam: %KAS, pH lateks, dan aktivitas SOD 11 1
...
6 . Analisis ragam: etilen, kadar ACC, dan aktivitas ACC-oksidase 11 1
7. Rekapitulasi hasil ujibeda (UJGD pada P < 0.05) beberapa peubah
...
pada dua faktor perlakuan Klon x Eksploitasi 1 12
8. Rekapitulasi hasil ujibeda (UJGD pada P < 0.05) beberapa peubah
pada dua faktor perlakuan Bulan x Klon ... 1 12
9. Rekapitulasi hasil ujibeda (UJGD pada P < 0.05) beberapa peubah
...
pada dua faktor perlakuan Bulan x Eksploitasi 1 12
10. Hasil ujibeda OJJGD pada P < 0.05) produksi g/p/s pada perlakuan
sistem eksploitasi setiap klon setiap bulan ... 1 13
1 1. Hasil ujibeda (UJGD pada P
.
.
.
< 0.05) KKK pada perlakuan sistemeksplortasr setrap klon setiap bdan ... 113
12. Hasil ujibeda (UJGD pada P < 0.05)
IP
pada perlakuan sistem.
.
.
eksplo~tasr setiap klon setiap bulan ... 1 14
13. Hasil ujiberla (UJGD pada P < 0.05) sukrosa pada perlakuan sistem
14. Hasil ujibeda (UJGD pada P < 0.05) FA pada perlakuan sistem
...
eksploitasi setiap klon setiap bulan 1 15
15. Hasil ujibeda (UJGD pada P < 0.05) ti01 pada perlakuan sistem
...
eksploitasi setiap klon setiap bulan.. 1 15
16. Hasil ujibeda (UJGD pada P < 0.05) pH lateks pada perlakuan sistem
...
eksploitasi setiap klon setiap bulan 116
...
17. Koefisien korelasi (r) linear sederhana menurut Pearson 116
1 8. Analisis regresifkorelasi ganda produksi lateks (g/p/s) terhadap
peubah-peubah lain ... 117
DAFTAR GAMBAR
Nomor Halaman
1. Anatomi batang karet (Hebant, 198 1).
...
10 2. Fraksi lateks setelah sentrifugasi 50 000 g selama 60 menit (d'Auzac& Jacob, 1989)
...
1 1 3. Berbagai lintasan sintesis produk sekunder dalam tanaman (Taiz &...
Zeiger, 199 1) 13
4. Sintesis karet dalam sel pembuluh lateks Hevea brasiliensis (Jacob
& Prevot, 1992).
...
1 55. Pengaruh stimulasi etepon terhadap daerah aliran lateks
(Pakianathan et al., 1989)
...
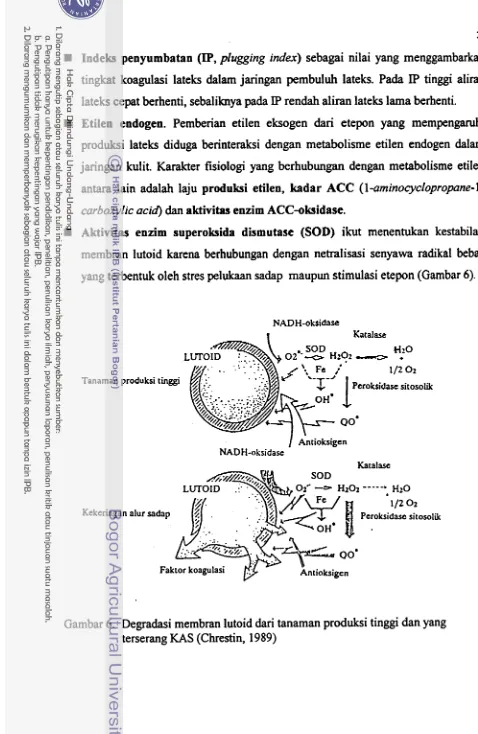
18 6. Degradasi membran lutoid dari tanaman produksi tinggi dan yang...
terserang KAS (Chrestin, 1989) 28
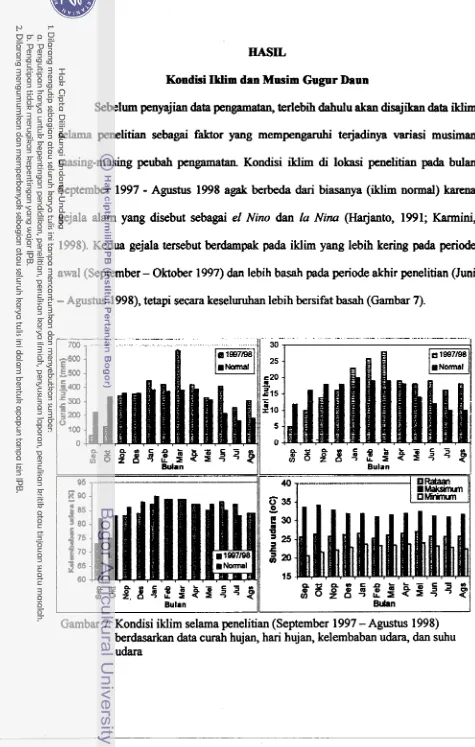
7. Kondisi iklim selama waktu penelitian (September 1997 - Agustus
1998) berdasarkan data curah hujan, hari hujan, kelembaban udara,
...
dan suhu udara 39
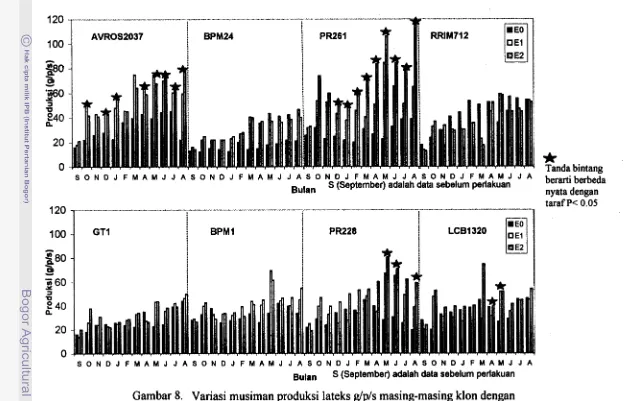
8. Variasi musiman produksi lateks g/p/s masing-masing klon dengan
...
sistem eksploitasi
&,
El dan E2 45.,
5'
9. Variasi musiman kadar karet kering masing-masing klon dengan
sistem eksploitasi Eo, El dan E2 ... 47
10. Variasi musiman indeks penyumbatan masing-masing klon dengan
...
sistem eksploitasi
&,
El dan E2 5011. Variasi musiman pH lateks 8 klon karet selama beberapa bulan
pengarnatan
...
5212. Variasi musiman kadar sukrosa lateks masing-masing klon dengan
sistem eksploitasi
&,
El dan E2...
5413. Variasi musiman kadar fosfat anorganik lateks masing-masing klon
14. Variasi musiman kadar ti01 lateks masing-masing klon dengan
dengan sistem eksploitasi EQ, El dan E2
... . . .
.. . .
59 15. Variasi musiman produksi etilen pada ketiga sistem eksploitasi... . . .
.
6 116. Variasi musiman kadar ACC bebas pada ketiga sistem eksploitasi ... .. ... ... 62
17. Variasi musiman aktivitas ACC-oksidase pada ketiga sistem
eksploitasi
...
6318. Pita protein hasil elektroforesis SDS-PAGE fraksi serum-C klon
RRLM 71 2, GT 1, dan BPM 1 masing-masing dengan perlakuan EQ,
El danE2
...
6519. Pita protein hasil elektroforesis SDS-PAGE fraksi lutoid beberapa
klon masing-masing dengan perlakuan EQ, El dan E2
...
6720. Pita protein hasil elektroforesis IEF fraksi lutoid klon GT 1, BPM 24
dan
RRIM
7 12 masing-masing dengan perlakuan&,
El dan E2... . .
6821. Protein hasil elektroforesis-2D (IEF-SDS-PAGE) dari fraksi lutoid
klon GT 1 dan BPM 24 dengan perlakuan
&,
El dan E2...
6822. Produksi lateks beberapa klon dalam satuan kghaftahun dan %.
Masing-masing klon adalah AVROS 2037, BPM 24, PR 261,
RRIM
712, GT 1, BPM 1, PR 228 dan LCB 1320..
. .. . .
.
..
.. .
. . .
.
. .
.
.. . .
. ..
.
.. .
..
7023. Hubungan penurunan IP terhadap peningkatan produksi g/p/s
berbagai klon dan perlakuan eksploitasi
...
7524. Pengaruh stimulasi terhadap IP dan produksi M s . Panjang dan ketegakan garis menggambarkan respons produksi lateks masing-
masing klon terhadap stimulasi etepon 2.5%
...
7625. Fluktuasi produksi lateks (a) dan kandungan etilen dalam jaringan
kulit pohon karet (b) klon AVROS 2037 dan BPM 24 pada 1
-
12hari SAE (setelah aplikasi etepon) 2.5%(E1) atau 5.0%(E2) ... 94
26. Skema hipotetik: Stimulasi etilen terhadap produksi lateks dalam
sistem sel pembuluh lateks
...
96Nomor
Lampiran
1
.
Lintasan biosintesis etilen dan jalur-jalur reaksi altematif lain(Mathooko. 1996) ... 119
2
.
TranskripHEVER
(Hevea ethylene.responsive). dari sel daun Hevea(a) dan sel pembuluh lateks Hevea yang diinduksi oleh etepon (b). serta dari jaringan Hevea yang diinduksi oleh asarn salisilat (c);
...
(Sivasubramaniarn et al.. 1995) 1 2 0
3
.
Denah percobaan di KP Ciomas. Bogor (250 m dpl)...
1214
.
Hubungan produksi glpls dan KKK ... 1225
.
Hubungan produksi g/p/s dan IP ... 1226
.
Hubungan produksi g/p/s dan FA...
122...
7
.
Hubungan KAS dan SOD 1238
.
Hubungan produksi g/p/s dan etilen ... 123PENDAHULUAN Latar Belakang
Sedikitnya telah seabad tanaman karet (Hevea brasiliensis Muell. Arg.)
mempunyai kedudukan penting dalam kebutuhan hidup manusia akan elastomer
karena nilai sosial-ekonominya yang cukup tinggi. Kebutuhan tersebut menyebabkan
karet dibudidayakan secara meluas sejak dari negeri asalnya di Brasil, Amerika Latin
hingga diintroduksi ke Asia dan Afiika. Saat ini negara produsen karet hevea yang
utama antara lain Thailand, Indonesia, Malaysia, Srilanka, dan Papua New Guinea.
Industri karet berkembang pesat baik di tingkat bahan baku maupun barang jadi,
bukan hanya di negara penghasil karet tetapi juga di negara-negara maju sebagai
konsumen. Perkembangan industri karet melibatkan banyak manusia yang
menggantungkan hidupnya di sektor ini, mulai dari pertanian, perdagangan maupun
perindustrian.
Karet alarn merupakan salah satu komoditas ekspor andalan bagi Indonesia,
karena menduduki urutan ekspor nonrnigas ketiga setelah kayu dan tekstil. Luas perkebunan karet di Indonesia pada tahun 1996 mencapai 3 448 5 10 ha dengan total
produksi 1 654 096 tonltahun, yang terdiri atas perkebunan rakyat 84.4%, perke-
bunan besar negara (PTP Nusantara) 8.7% dan perkebunan besar swasta 6.9%
(Ditjenbun, 1996).
Saat ini terdapat 20 klon unggul anjuran dan 18 klon unggul harapan hasil
pemuliaan (Puslit Karet, 1995). Di samping berlaku spesifik terhadap kondisi
terhadap sistem eksploitasi (suatu sistem yang meliputi tindak penyadapan, frekuensi
dan stimulasi; yang dapat dituliskan dalam notasi baku). Oleh karena itu di dalam
penerapannya harus menggunakan sistem eksploitasi yang paling tepat. Pada kondisi
krisis dewasa ini, peningkatan produksi tanarnan karet merupakan bidang kajian
yang makin mendapatkan prioritas terutama karena produktivitas tanaman masih
tergolong rendah. Beberapa usaha untuk meningkatkan produktivitas karet di
Indonesia telah dilakukan melalui berbagai pola pengembangan, yaitu pola unit
pelaksana proyek
(PRPTE,
PPKR, atau SRDP), perusahaan inti rakyat (PIR) dan polaswadaya berbantuan.
Hevea bukan satu-satunya tanaman penghasil karet, karena ada tanaman
lain seperti guayule (Parthenium argentantum), rambung (Ficus elastics), dan
jelutung (Dyera Spp), namun produksi karet dari hevea memiliki keunggulan
komparatif sehingga secara umum lebih menguntungkan. Bahkan meskipun industri
maju telah mampu membuat karet sintetik, eksistensi karet alam dari hevea tidak
dapat dihilangkan. Komplementasi antara karet alam dan karet sintetik dewasa ini
cenderung makin kokoh dibutuhkan dalam berbagai industri. Kelemahan karet
sintetik yang paling mendasar dan dimaklumi oleh semua pihak adalah sumber
bahannya berasal dari minyak bumi, sehingga tidak memiliki sifat dapat diperbarui
(renewable) seperti halnya karet alam khususnya Hevea brasiliensis.
Tanaman hevea sebagai penghasil karet tergolong produktif, terlebih setelah
melalui masa pembudidayaan yang panjang telah terpilih klon-klon unggul, dan
tanaman hevea lebih rendah dibandingkan dengan tanaman penghasil karet lain.
Partikel karet sangat mudah diperoleh dengan penyadapan kulit batang yang
mengeluarkan getah (lateks) yaitu cairan atau sitoplasma yang berisi sekitar 30%
partikel karet. Pada tanaman hevea, lateks dibentuk dan terakumulasi &lam sel-sel
pembuluh lateks yang tersusun pada setiap jaringan bagian tanaman, namun
penyadapan yang menguntungkan hanya dilakukan pada kulit batang dengan sistem
eksploitasi tertentu.
Secara fisiologis produksi lateks yang diperoleh dari hasil penyadapan
ditentukan antara lain oleh lamanya lateks mengalir dan kecepatan biosintesis atau
regenerasi lateks (Kekwick, 1989). Biosintesis lateks ditentukan oleh ketersediaan
bahan dasar pembentuk lateks berupa sukrosa dan oleh aktivitas enzim yang
berperan secara langsung, baik pada tahap glikolisis maupun anabolisme partikel
karet (cis-poliisoprena) (Jacob & Prevot, 1992).
Berhentinya aliran lateks disebabkan oleh adanya koagulasi partikel karet
yang menyumbat luka irisan sadap, sebagai akibat dari pecahnya organel di dalam
sel pembuluh lateks yang disebut sebagai lutoid dan partikel Frey Wyssling. Organel
tersebut akan membebaskan senyawa yang berperan secara langsung sebagai
penyebab koagulasi karet. Lutoid selain mengandung protein juga mengandung
serum asam, kation bervalensi dua seperti M$+, ca2+ sehingga muatan negatif dari
pertikel-partikel karet akan dinetralkan dan menyebabkan te rjadinya koagulasi.
Penambahan etepon (2-chloroethyl phosphonic acid, CEPA) sebagai bahan
dilakukan terutama pada perkebunan besar. Aplikasi etepon dimaksudkan untuk
menekan biaya eksploitasi clan memperoleh produksi yang tinggi. Narnun setiap
klon memiliki respons yang berbeda terhadap intensitas eksploitasi. Intensitas
eksploitasi mencakup faktor panjang irisan sadap, fiekuensi sadap dan aplikasi stimulasi etepon. Penggunaan etepon yang berlebihan atau intensitas sadapan yang
tinggi misalnya S/1 dl1 (irisan sadap 1 spiral dan dilakukan setiap hari) seperti yang
sering tejadi di perkebunan rakyat, biasanya diikuti oleh tingginya jumlah pohon
yang mengalami kekeringan alur sadap (KAS) (Darussamin et al., 1995; Siswanto,
1997). Sistem eksploitasi yang baik adalah yang memberikan produksi optimal,
tidak menekan perturnbuhan tanarnan, hemat kulit dan biaya murah serta tidak
menimbulkan KAS
--
yang merupakan gangguan fisiologis sehingga tanaman karettidak dapat mengalirkan lateks apabila disadap. Diperkirakan kerugian produksi yang
diakibatkan oleh KAS setiap tahunnya mencapai Rp 1.7 triliun; yakni pada
perkebunan besar negara (PTP Nusantara) Rp 1.4 triliun, perkebunan rakyat Rp 147
milyar dan perkebunan besar swasta Rp 1 16 milyar (Siswanto, 1998).
Untuk penetapan sistem eksploitasi yang optimal dikenal suatu program di
perkebunan karet yang disebut sebagai diagnosis lateks (Jacob et al., 1989). Di
Indonesia diagnosis lateks belum diterapkan secara luas. Perintisan baru dimulai di
perkebunan besar swasta asing, khususnya di Sumatra Utara yaitu pada konsorsiurn
PT Lonsum, Socfindo, Sipef dan Goodyear (Sembiring, 1994); sedangkan di
perkebunan besar negara (PTP Nusantara) masih pada tahap penyusunan data base.
solid content (TSC), pH lateks, kadar sukrosa, kadar fosfat anorganik dan kadar tiol.
Karakter fisiologi lain yang diperkirakan penting adalah indeks penyumbatan
(plugging index,
IP)
(Milford, Paardekooper &' Ho, 1969) dan aktivitas enzimsuperoksida dismutase (Miao & Gaynor, 1993).
Pembentukan etilen dalam jaringan tanaman antara lain terjadi karena
proses penuaan, pematangan buah, pelukaan, cekaman kekeringan atau
penggenangan, dan sebagainya, sudah merupakan fenomena umurn yang diketahui
pada berbagai tanaman (Mathooko, 1996; Harrison, 1997). Namun demikian,
pembentukan etilen dalam tanaman karet akibat pelukaan sadap atau aplikasi
stimulasi oleh sistem eksploitasi masih belum banyak diketahui (Sivakumaran,
Pakianathan & Abraham, 1984).
Kajian fisiologi perlakuan etepon pada tanaman karet umumnya tidak
dikaitkan dengan metabolisme etilen dalam jaringan kulit. Oleh karena itu fenomena
respons perubahan kadar etilen endogen terhadap pemberian etepon secara
kuantitatif perlu dipelajari secara simultan dengan berbagai karakter fisiologi
tanaman karet. Pembentukan etilen endogen dapat diduga melalui pengukuran
produksi etilen, kadar ACC ( 1 -aminocyclopropane- 1 -carboxylic acid) sebagai
prekursor etilen, dan aktivitas enzim ACC-oksidase sebagai enzim terakhir
pembentuk etilen (Paranjothy et al., 1979; Sivakumaran et al., 1984; Dominguez &
Vendrell, 1993, Katherisan, 1996). Pengdcuran tersebut penting mengingat ketiganya
berada pada penghujung metabolisme etilen, dan merupakan jalur yang berbeda
Peningkatan produksi oleh stimulasi etepon dapat diterangkan oleh
pengaruhnya terhadap aliran dan regenerasi lateks. Pengaruh etepon terhadap aliran
lateks antara .lain: meningkatkan influks air, sehingga memperluas daerah aliran
lateks (Pakianathan et al., 1989), dan meningkatkan kestabilan lutoid sehingga
indeks penyumbatan menurun (Jacob, Prevot & Kekwick, 1989). Pengaruh etepon
terhadap regenerasi lateks antara lain meningkatkan influks sukrosa melalui ATPase
clan pirofosfatase pompa proton sehingga meningkatkan metabolisme lateks serta meningkatkan fosforilasi oksidatif (Jacob et al., 1992; Siswanto, 1993).
Untuk menetapkan sistem eksploitasi sesuai dengan jenis klon dan variasi
musiman, perlu diidentifikasi faktor pembatas utarna serta kombinasi dari beberapa
k a r d e r fisiologi sebagai penanda produksi. Karakter fisiologi tersebut hendaknya
digunakan secara bersarna-sama karena produksi lateks tidak hanya ditentukan oleh
salah satu karakter saja namun ditentukan oleh keseimbangan yang harmonis antara
beberapa faktor pembatas yang potensial. Karakter fisiologi yang diarnati adalah
yang berkaitan dengan regenerasi dan aliran lateks antara lain kadar sukrosa, fosfat
anorganik, tiol, pH lateks, total solid content, atau kadar karet kering (Jacob et al.,
1989), indeks penyumbatan (Milford et al., 1969), termasuk etilen dan
metabolismenya (Paranjothy et al., 1979; Sivakumaran et al., 1984), serta aktivitas
enzim superoksida dismutase M a o & Gaynor, 1993) dan pola pita protein
Perumusan Masalah
Sampai saat ini perkebunan karet di Indonesia, baik perkebunan rakyat
maupun perkebunan besar, masih kurang mempertimbangkan jenis klon maupun
variasi musim &lam penerapan sistem eksploitasi. Padahal, kemarnpuan regenerasi
lateks berbeda untuk setiap klon dan khususnya ditentukan oleh karakter fisiologi.
Hal tersebut mengakibatkan klon yang tampalcnya memiliki produksi rendah tetapi
memiliki potensi produksi tinggi, tidak dieksploitasi secara optimal. Sebaliknya klon
yang sudah tinggi produksinya, jika masih dilakukan stimulasi berlebihan maka akan
memperbanyak tanaman terserang KAS. Variasi musiman dapat mempengaruhi
fluktuasi produksi karena adanya pengaruh musim gugur daun tahunan serta
perubahan musim kering dan musim hujan.
Tujuan dan Manfaat Penelitian
Penelitian ini bertujuan untuk mengetahui variasi musiman beberapa
karakter fisiologi tanaman karet akibat stimulasi etilen yang berasal dari etepon, dan
mengetahui keterkaitan antara karakter-karakter fisiologi pada lateks maupun etilen
endogen pada jaringan kulit terhadap tingkat produksi beberapa klon tanaman karet.
Adapun manfaat penelitian maupun perspektif penelitian yang diharapkan
antara lain adalah :
1) Hasil kajiap karakter fisiologi dapat digunakan dalam upaya mengoptimalkan
sistem eksploitasi tanaman karet secara diskriminatif terhadap jenis klon karet
2) Selanjutnya sistem eksploitasi yang tepat berdasarkan sifat suatu klon karet dan variasi musiman akan mendukung tingkat produksi karet yang optimal sekaligus
menglundari KAS.
3) Pada tingkat penelitian lebih lanjut dengan antibodi dari protein penanda tingkat
produktivitas, dapat dipergunakan untuk menduga potensi produksi Mon karet
atau untuk mendeteksi dini tanarnan karet muda hasil persilangan.
Hipotesis
Dalam penelitian ini hipotesis yang diajukan adalah :
1) Karakter fisiologi tanaman karet berupa kadar karet kering, indeks penyumbatan,
pH lateks, kadar sukrosa, kadar fosfat anorganik, kadar ti01 dan aktivitas enzim
superoksida dismutase; dapat digunakan untuk menilai tingkat yang optimal
suatu sistem eksploitasi. Stimulasi etepon dapat meningkatkan produksi lateks
yang diceminkan oleh perubahan karakter fisiologi yang mendukung.
2) Karakter-karakter fisiologi tanarnan karet tersebut memiliki pola respons spesifik berdasarkan perbedaan jenis klon dan variasi musimannya.
3) Stimulasi etepon sebagai generator etilen dapat menginduksi ekspresi tanaman
karet dalam sintesis suatu protein tertentu, dan kemudian akan mendukung
peningkatan produksi tanaman karet. Protein tersebut berperan sebagai protein
'I'INJAUAN PUSTAKA Botani Tanaman Karet
Tanaman karet (Hevea brasiliensis Muell. Arg.) merupakan tanaman
tahunan (perennial) dari famili Euphorbiaceae penghasil lateks yang telah lama
dibudidayakan. Tanaman turnbuh baik di dataran rendah hingga menengah (0
-
400m di atas permukaan laut) dengan curah hujan yang cukup sepanjang tahun (1500
-
2500 mrnltahun), dan marnpu hidup di lahan masam (pH 4.0
-
4.5) dengan tanah bersolum dalam dan miskin hara (Dijkman, 195 1; Webster & Baulkwill, 1989).Saat ini di Indonesia terdapat 20 klon unggul anjuran dan 18 klon unggul
harapan hasil pemuliaan (Puslit Karet, 1995). KIon unggul anjuran adalah klon-klon
yang direkomendasikan untuk pertanaman komersial dan dilepas secara resmi
men.& benih bina dengan Surat Keputusan Menteri Pertanian. Klon-klon dimaksud
adalah AVROS 2037, BPM 1, BPM 24, BPM 107, BPM 109, GT 1, PB 2 17, PB 235,
PB 260, PR 255, PR 261, PR 300, PR 303,
RRIC
100, RRIC 102, RRIC 110, RRIM600, RRIM 712, TM 2 dan
TM
9. Klon unggul harapan adalah klon yangdikembangkan secara terbatas di Perusahaan Perkebunan berdasarkan rekomendasi
dari Pusat Penelitian Karet, di antaranya adalah
IRR
2, IRR 5, IRR 7, PB 280, PB 330,RCG
2604,TM
6 danTM
8 (Woelan et al., 1995).Klon-klon anjuran tidak berlaku universal, tetapi perlu agroekosistem yang
tepat dalam ha1 curah hujan dan kelembaban udara, kedalaman solum tanah,
topografi, penyakit utarna, dan gangguan angin. Namun berdasarkan ciri genetik
10
setidaknya dibedakan dalam 6
-
8 kelompok pola pita protein (Pnmatillake & Yapa,1985; Lasminingsih et at., 1989). Di antara klon-klon anjuran, GT 1 misalnya
berbeda kelompok dengan kelompok PR 300
-
PR 303 dan kelompok RRIM 600.Pencirian genetik berdasarkan pola pita protein masih memiliki kelemahan, yaitu
adanya variasi tampilan elektroforegram suatu klon akibat perbedaan umur
penyadapan; atau dengan kata lain faktor penyadapan dapat mempengaruhi
kestabilan pola protein dari suatu klon yang diidentifikasi. Oleh karena itu pencirian
genetik berdasarkan pola pita isozim (enzim-enzim yang berbeda gugus
hgsionalnya tetapi aktivitas enzimatiknya sarna) juga sedang diupayakan
(Sitompul, 1995).
Keterannan: X- Xylem, Floern (U- h i t keras + ki- lculit lunak),
[image:174.574.31.515.29.789.2]L ~embuluh lateka C- karnbium dan PV- ~embuluh vaskuler.
Gambar 1. Anatomi batang karet (Hebant, 198 1)
Meskipun lateks terdapat dalarn daun, ranting dan percabangan, namun
penyadapan yang menguntungkan adalah pada kulit batang. Anatomi batang karet
dilakukan pada tanaman benunur 5
-
6 tahun, yakni apabila telah memenuhi kriteriamatang sadap. Kriteria matang sadap yang masih berlaku hingga saat ini adalah lilit
batang pada ketinggian 1.0 atau 1.3 m di atas 'kaki gajah' (bekas pertautan okulasi)
sebesar 45 cm atau lebih, dan lebih dari 60% populasi telah memenuhi ukuran lilit
batang tersebut. Penyadapan diharapkan dapat dilakukan selama 25
-
30 tahun.Oleh karena itu hams diusahakan agar kulit pulihan dapat terbentuk dengan baik,
sehingga pada saatnya dapat disadap lagi. Pengirisan kulit makin mendekati
kambium akan makin banyak mengeluarkan lateks, namun apabila mengenai
kambium
dan
kayu akan mengakibatkan buruknya kulit pulihan. Oleh karena itupenyadapan yang baik merniliki kedalaman be rjarak 1.0
-
1.5 rnm dari kambium(Junaidi & Kuswanhadi, 1992).
1. fraksi karet, berwarna putih
2. partikel Frey Wyssling, kuning-jingga
3. senun-C (sitosol), cairan bening
4. fraksi bawah (lutoid), endapan kuning
Gambar 2. Fraksi lateks setelah sentrifugasi 50 000 g selama 60 menit (d7Auzac &
Jacob, 1989)
Lateks yang keluar dari penyadapan pohon karet sebenarnya merupakan
sitoplasma dari sel-sel pembuluh lateks yang mengandung antara lain partikel karet,
lutoid, nuklek, mitokondria, ribosom dan partikel Frey Wyssling. Dengan cam
[image:175.568.44.506.0.767.2]12
tiga bagian utama yaitu lapisan atas berupa partikel karet yang berwama putih,
lapisan tengah merupakan cairan bening yang dinamakan serum-(: (sitosol), dan
endapan kuning pada dasar tabung disebut sebagai 'fraksi bawah' yang terdiri atas
partikel lutoid (Gambar 2).
Partikel karet berbentuk bulat berukuran antara 5 nm
-
3 pm, mengandungbeberapa ratus molekul cis- l,4-poliisoprena (d' Auzac dan Jacob, 1989). Partikel
karet dikelilingi oleh membran berupa fosfolipoprotein yang bermuatan
elektronegatif dan berperan untuk menjaga stabilitasnya. Enzim rubber transferase
berada pada membran tersebut dan berfimgsi dalam pembentukan molekul karet.
Serum C adalah cairan bening yang merupakan sitosol dari sel pembuluh lateks,
mengandung berbagai persenyawaan antara lain sukrosa, protein dan
asam-asam
organik. Adapun lutoid meliputi kurang lebih 10
-
20% dari volume lateks. Lutoidadalah organel yang disebut juga sebagai vakuolisosom karena sifatnya yang
menyerupai vakuola (pada sel tanaman) dan lisosom (pada sel hewan). Partikel
lutoid berdiarneter 2
-
10 pm, dikelilingi oleh satu lapisan membran yang tipis clanbersifat peka terhadap perbedaan tekanan osmosis sehingga dapat pecah pada
medium yang hipotonik.
Lutoid mengandung cairan (serum lutoid) dengan pH 5.0
-
5.5 yangmerupakan tempat akurnulasi senyawa atau ion yang bersifat racun, antara lain asam
sitrat, asam oksalat, M ~ ~ + , ca2+, FA, hevein, dan lisosim. Hevein mencakup 70%
dari total protein terlarut &lam serum lutoid dan dinyatakan bahwa hevein
13
ini diperkirakan berfhgsi sebagai suatu sistem proteksi tanaman karet untuk
menghambat terjadinya infeksi patogen pada saat pelukaan batang akibat
penyadapan. Namun, Kush et al. (1990) melaporkan bahwa hevein bila berasosiasi
dengan reseptor berukuran 23 kDa pada membran partikel karet akan menyebabkan
penggumpalan atau koagulasi lateks. Asosiasi hevein dengan reseptor tersebut
kemudian ternyata diduga kuat memerlukan N-asetil glukosamin (Jacob et al., 1998).
Produksi Lateks Tanaman Karet
Dalam metabolisme tanaman, fotosintesis merupakan proses awal penghasil
glukosa kemudian te rjadi proses-proses pembentukan sukrosa atau karbohidrat lain,
lipid, protein, dan metabolit sekunder melalui berbagai lintasan yang terkoordinasi
dengan proses respirasi (glikolisis, siklus trikarboksilat). Metabolit sekunder berupa
senyawa fenolik, terpen maupun produk sekunder mengandung N; secara spesifik
memiliki lintasan tertentu antara lain lintasan sikimat, malonat maupun mevalonat
(Gambar 3) (Taiz & Zeiger, 1991).
.
PEP
Senyawa fenolik
Gambar 3. Berbagai lintasan sintesis produk sekunder dalam tanaman (Taiz &
Lateks mengandung partikel karet (isoprena) yang dihasilkan oleh tanaman
karet dan merupakan produk sekunder yang tergolong sebagai terpen (politerpen),
yang disintesis melalui lintasan asam mevalonat (MVA). Sebagai prekursor dari
isoprena adalah asetil koA atau asam asetat, tetapi dalam jaringan dipersiapkan
berupa sukrosa yang mudah ditranslokasikan (Kekwick, 1989; West, 1990; Jacob et
al., 1998). Secara fisiologis produksi lateks pada tanaman karet dipengaruhl oleh dua
faktor utarna, yaitu biosintesis atau regenerasi lateks antara dua penyadapan, dan
lamanya lateks mengalir setelah penyadapan (Kekwick, 1989).
Biosintesis/Regenerasi Lateks
Biosintesis/regenerasi lateks berlangsung dalam sel pembuluh lateks
(laticferous cells) dkngan bahan dasar berupa sukrosa yarig berasal dari daun,
sebagai pusat fotosintesis. Metabolisme yang berlangsung terutama berotientasi
untuk pembentukan partikel karet (cis-poliisoprena atau (C&& yang mewakili 33
-
50% bobot segar atau 90% bobot kering lateks. Skema sintesis karet dalam sel
pembuluh lateks disajikan pada Gambar 4 (Jacob & Prevot, 1992). Ada dua tahap
utama pembentukan karet yaitu glikolisis dan anabolisme cis-poliisopre~.
Dalam sel pembuluh lateks, sebagian besar sakarida berbentuk sukrosa
(Tupy & Resing, 1968; d'Auzac & Pudarniscle, 1959). Aktivitas jaringan pembuluh
lateks tergantung pada ketersediaan sukrosa tersebut (Tupy, 1989). Dalam analisis,
kadar sukrosa merupakan residu dari jurnlah sukrosa tersedia dikurangi
15
sangat kompleks, menyangkut mekanisme tingkat membran sel yang memerlukan
energi biokimia (Jacob et al., 1995).
Molekul sukrosa melalui serangkaian reaksi enzimatik (invertase clan
utama dari cis-poliisoprena dengan pembentukan energi biokimia berupa ATP serta
akurnulasi senyawa pereduksi NAD(P)H. Asetil koA yang dihasilkan dari glikolisis
selanjutnya membentuk monomer isoprena yaitu isopentenil pirofosfat (IPP). Proses
pembentukan IPP ini memerlukan energi terutama dalam bentuk ATP dan tenaga
pereduksi NADH yang dihasilkan dari proses glikolisis. Dari IPP secara berturut-
turut terbentuk partikel karet (poliisoprena) yang makin besar. Biosintesis lateks
dikendalikan antara lain oleh pengaturan pH dan komposisi ion dalam sitosol lateks.
Adanya korelasi positif antara produksi lateks dan pH sitosol telah terbukti dengan r
= 0.639 (Brzozowska et al., 1979).
Biosintesis karet sebenarnya melalui reaksi biokimia yang melibatkan
sangat banyak enzim sebagai biokatalisator. Namun untuk lintasan utama dapatlah
disederhanakan dalam lima tahap sebagai berikut :
1. Pembentukan 3-hidroksi 3-metil ~lutaril koA
112
asetil koA
*
asetoasetil koA+
asetil koA,
3-5-3-hidroksi 3-metil glutaril koA El = asetil koA asil transferase (EC 2.3.1.9, serum)(H
MG-k0A) E2 = 3Wmksi 3-metil glutaril koA sinthase (K 4.1.3.5, serum)2. Reduksi 3-hidroksi 3-metil glutaril koA dm ~embentukan mevalonat pirofosfat
I S I 4
3-5-3-hidroksi 3-metil glutaril koA
+
2 NADPH+
2H++ 3-R-mevalonat+
mevalonat 5-PP(HMG-koA) (MVA) Is (MVA-PP)
E3 = 3 h imetil glutad koA reduktase (K 1.1 .I .3.4, fraksi bawah RE)
E4 = meMknat kinase (K 2.7.1.3.6, serum) E5 = mevabnat-PP base (EC 2.7.4.2, serum)
3. Pembentukan monomer isoprenoid: iso~entenil pirofosfat dan primer dimetilalil pirofosfat
.
E 6 I7
mevalonat-5PP
-+
isopentenil pirofosfat
<-
dimetilalil pirofosfat
(
MVA-PP)
(Ipp)
(DMAPP)
4. Pembentukan semua trans vrenol virofosfat vrimer
11 8
dimetilalil pirofosfat
+
3 isopentenil pirofosfat ->
geranil geranil pirofosfat
(DMApp)
(Ipp)
(GGPP)
E8 = prenil transferase (EC 2.5.1 .l, serum)
5. Pembentukan cis-wliiso~rena E 9 E 9 E 9
geranil geranil pirofosfat
+
isopentenil pirofosfat
+
-,
+
cis-poliisoprena
(GGPP)
(Ipp)
E9 = rubber transferase (K 2.5.1.20, permukaan partikel k tkecil)
(KARET)
Dari lima tahapan reaksi di atas terdapat sembilan enzim yang terlibat langsung dalarn biosintesis partikel karet (cis- l,4-poliisoprena). Menurut Kekwick
(1989) sembilan enzim tersebut, yang termasuk dalam kelas oksidoreduktase ialah
enzim HMG-koA reduktase, dalam kelas transferase ialah enzim asetil-koA-asil
transferase, MVA kinase, WA-PP kinase, prenil transferase, dan rubber transferase;
dan dalam kelas liase ialah enzim HMG-koA sinthase dan MVA-PP dekarboksilase.
Terdapat satu enzim dalam kelas isomerase yaitu enzim IPP isomerase. Sebagian
besar enzim-enzim utama pembentuk karet ini terdapat dalam serum atau sitosol sel
pembuluh lateks, kecuali enzim
HMG
koA reduktase yang terdapat pada fhksibawah (lutoid) dan enzim rubber transferase yang terdapat di permukaan partikel
karet. Kedudukan enzim ini berkaitan langsung dengan bgsinya dalam reaksi
biokimia dalam sel.
Lama Aiiran Lateks
Penyadapan dilakukan pada kulit pohon hingga mencapai dekat kambium.
Lateks berada dalam pembuluh lateks pada tekanan turgor 10-14 atmosfer. Segera
18
menembus dinding sel pembuluh lateks sehingga lateks mengalir sepanjang irisan
sadap. Lateks yang diperoleh dari penyadapan tidak saja berasal dari sel-sel
pembuluh lateks yang terlukai tetapi merupakan kumpulan lateks yang mengalir dari
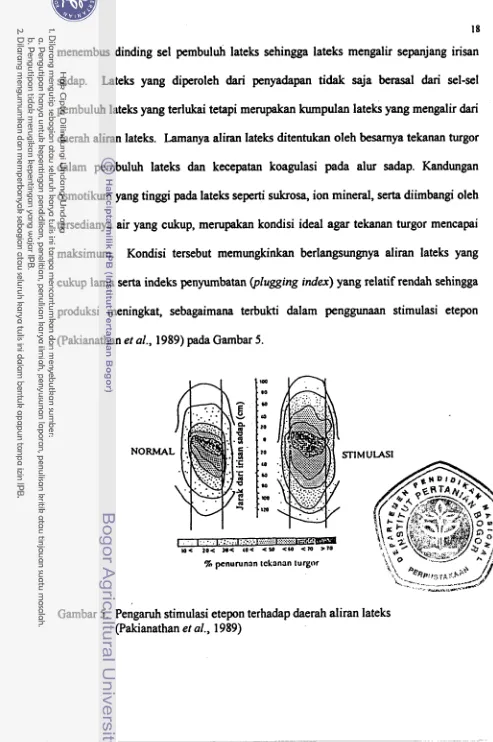
daerah aliran lateks. Lamanya aliran lateks ditentukan oleh besarnya tekanan turgor
&lam pembuluh lateks
dan
kecepatan koagulasi pada alur sadap. Kandunganosmotikurn yang tinggi pada lateks seperti sukrosa, ion mineral, serta diimbangi oleh
tersedianya air yang cukup, merupakan kondisi ideal agar tekanan turgor mencapai
maksimum. Kondisi tersebut memungkinkan berlangsungnya aliran lateks yang
cukup lama serta indeks penyumbatan (plugging index) yang relatif rendah sehingga
produksi meningkat, sebagaimana terbukti dalam penggunaan stimulasi etepon
(Pakianathan et al., 1989) pada Gambar 5.
[image:182.572.38.531.34.776.2]!% penurunan tckanan turgor
Beberapa jam setelah batang karet disadap, aliran lateks &an terhenti.
Berhentinya aliran lateks lebih disebabkan oleh adanya koagulasi partikel karet yang
menyurnbat luka irisan sadap sebagai akibat dari perusakan lutoid dan partikel Frey
Wyssling. Organel tersebut melepaskan beberapa senyawa yang berperan secara
langsung sebagai penyebab koagulasi karet.
Terdapat dua enzim penting yang terlibat dalam proses koagulasi karet,
yaitu 0-difenol oksidase yang terdapat dalarn partikel Frey Wyssling dan enzim
fosfolipase-D yang terdapat pada serum sitosol. 0-difenol oksidase mengkatalisis
tejadinya koagulasi apabila senyawa fen01 berasosiasi dengan oksigen udara.
Adapun fosfolipase-D (PLD) marnpu menghidrolisis komponen utama membran
lutoid, yakni fosfolipid. Enzim ini dapat terpacu aktivitasnya oleh ion ca2+ yang keluar dari lutoid yang mengalami kebocoran atau kerusakan, sehingga akan
mempercepat proses koagulasi (d'Auzac & Jacob, 1989). Degradasi fosfolipid pada
membran sel oleh PLD belakangan dilaporkan diinduksi oleh etilen (pada wortel),
yang diikuti oleh aktivitas enzim lipolitik asil-hidrolase (LAH) (Soo et al., 1998).
Pemberian Etepon
Penggunaan stimulan untuk meningkatkan produksi lateks pada mulanya
berawal dengan pemberian bahan-bahan tradisional, yaitu dengan memoleskan
campuran kotoran lembu dengan tanah liat ke bidang sadap, maupun minyak yang
mengandung hormon tanaman (Webster & Baulkwill, 1989); kemudian berkembang
sulfat yang diinjeksikan ke &lam kayu (Mainstone &
Tan,
1964, dan Lowe, 1964) ,gas etilen oksida, gas asetilen, dan etepon (CEPA) yang mampu melepaskan gas
etilen (Abraham et al., 1971% b, c) dan secara komersial diproduksi dengan nama
~threl", cephaa,
lot?'
dan ~thad@.(Lukrnan, 197 1 ; Tjasadihardja, 1976; 1977)Penelitian dan penerapan di perkebunan selama periode 1955 hingga 1980-
an membuktikan bahwa penggunaan stimulan sebaiknya dilakukan pada tanaman
karet yang cukup tua yaitu sudah disadap lebih dari 12 tahun. Penerapan stimulan
bagi tanaman yang lebih muda umurnnya lebih berisiko. Berbagai faktor yang
diketahui berpengaruh terhadap respons tanarnan antara lain adalah kualitas kulit,
umur pohon, kultivar (klon), sistem sadap, konsentrasi bahan aktif dalam campuran,
serta cara dan frekuensi aplikasi (Webster & Baulkwill, 1989).
Pengaruh stimulasi dengan etepon terutarna adalah meningkatkan produksi
lateks, menurunkan kadar karet keririg (KKK), namun juga berpengaruh terhadap
komposisi lateks, sifat teknis lateks dan karet (antara lain viskositas, plastisitas,
stabilitas mekanis dan warna), kekeringan batang, perturnbuhan lilit batang, kulit
pulihan, komposisi dam; dan memiliki pengaruh jangka panjang yang berkaitan
dengan kapasitas produksi (Sivakumaran et al., 1984).
Peningkatan produksi lateks berkisar antara 20
-
100% selama satu siklusstimulasi, terutarna disebabkan oleh lamanya aliran lateks. Pemanjangan waktu
aliran lateks disebabkan oleh turunnya tingkat penyumbatan lateks. Secara normal
pohon karet yang disadap akan mengeluarkan lateks dalam jangka waktu tertentu,
penyumbatan (IP). Semakin rendah IP semakin lama lateks mengalir. Proses
penyumbatan terjadi akibat pecahnya lutoid dalam sel pembuluh lateks, dan ini
berarti pemberian stimulan akan lebih menstabilkan lateks (lutoid tidak pecah),
sehingga Iateks tetap mengalir. Pemberian stimulan umurnnya memberi tambahan
hasil setelah lateks dikutip, yakni berupa lump mangkok pada hari berikutnya.
Etepon biasanya diaplikasikan pada pohon karet dalam bentuk produk
komersial ~threl" (Am.Chem.; Rhone Poullenc) dengan konsentrasi 10.0% dan
kemudian dapat diencerkan dengan minyak kelapa sawit sampai dengan konsentrasi
2.5%. Pada sistem eksploitasi dengan frekuensi sadap yang lebih rendah, konsentrasi
etepon dapat ditingkatkan sampai 5.0
-
10.0%. Aplikasi etepon pada pohon ada duamacam yaitu pada alur irisan sadap (Ga=groove application) atau pada kulit bidang
sadap dengan pengerokan (Ba=bark application). Bagi tanaman karet dengan
kondisi normal dan cukup urnur (lebih dari 10 tahun), sistem eksploitasi yang sering
dianjurkan adalah %S&3.ET2.5%.Ba1.0(1.5)9/y(m); yaitu sadapan % spiral, disadap
3 hari sekali dengan pemberian etepon 2.5% sebanyak 1.0 gram yang diaplikasikan
pada bidang sadap yang dikerok selebar 1.5 cm selama 9 bulan per tahun; berarti
selama 3 bulan gugur daun, tanaman diistirahatkan (Lukman,