STUD1 GENETIK KARAKTER HORTIKULTURA DAN
KETAHANAN TERHADAP CUCUMBEX MOSAIC WRUS
DAN CHILLI VEINAL MOTTLE WRUS
PADA
CABAI (Capsicurn cmnuurn L.)
AGUS RIYANTO
SEKOLAH PASCASARJANA
INSTITUT PERTANZAN BOGOR
BOGOR
PEKNYATAAN MENGENAI TESIS
DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Studi Genetik Karakter Hortikultura dan Ketahanan terhadap Cucumber Mosaic Virus dan Chilli Veinal n//otrle Virzrs pada Cabai (Capsictnn annuton L.) adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan
iinggi inanapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan d a l m
Daftar
Pustaka di bagian hkhir iesis ini.Bogor, September 2007
ABSTRACT
AGUS RIYANTO. Genetic Study of Horticulture Character and Resistance to
Cucumber iMosaic T'irzrs and Chilli Veinal A4ottle Virus in Chili (Capsicum unnuznn L.). Supervised by SRIANI SUJIPRIHATI, and SRI HENDRASTUTI NIDAYAT.
Czrantnber mosaic vjrus (ChlV) dan clzilli veinal mottle virus (ChiVMV) are iotlsidrred inajor viruses that mav cause significant yield losses in chili pepper (C. nnnuuin). The use of resistance variety is becoming impoltant fnr controlling virus infection since other methods were not effective. Genetic information. such as genetic parameter, combining ability, and heterosis is required in order to develop resistance variety.
The research involved three activities, i.e. (1) Evaluation of horticulture character; (2) Evaluation of chili response to CMV; and 3) Evaluation of chili response to ChiVMV.
The objectives of the 1" activity were (I) to evaluate genotypes for hodiculture character and (2) to estimate genetic parameter, general combining ability (GCA), specific combining ability (SCA) and heterosis of horticulture character. One set of population from half diallel mating, involving 6 parents and
15 hybrids were used in this research.
The objectives of the 2nd and the 3rd activities were (1) to estimate genetic parameters, general combining ability (GCA), specific combining ability (SCA) and heterosis of resistance to CMV and resistance to ChiVMV character and (2)
to evaluate genotypes for their response to CMV and ChiVMV infection. Two sets of population from ha!f diallel mating, involving 6 parent and 15 crosses for each set, were used in this research. One set of these population were inoculated with CMV isolate 02 and another set with ChiVMV isolate Cikabayan. Infection of the viruses were c o n f i i e d by DAS-ELISA. Furthermore, absorbance value of ELISA was used as an indicator of virus titer for calculation of combining ability and heterosis estimation.
It was evidenced that heterosis occurred due to interaction between genes controlling number of fruits and those controlling overdominance for fruit yield per plant, days of harvesting, resistance to Ch4V and resistance to ChiVMV. This gene interaction can be used for deve1opi;ig improved hybrid varities. General combing ability (GCA) effect will influence specific combining ability (SCA) in which the latter is responsible for improvement of heterosis. Narrow sense heritability values of horticulture character, resistance to CMV and resistance to ChiVMV were high and narrow sense heritability values were range from low to high. Resistance to CMV and ChiVMV were controlled by each recessive and dominant gene. Two parents i.e. IPB C14 and IPB C10 can be used as donor parents for developing both CMV and ChiVMV resistance chili varieties, whereas IPB C1 can be used as donor parents for developing ChiVMV resistance chili varieties only. Three hybrids showed resistance to CMV and 6 hybrids showed resistxlct only to Chi\ll\.lV.
RINGKASAN
AGUS RIYANTO. Studi Genetik Karakter Hortikultura dan Ketahanan terhadap
Cucunzber Mosaic Virus dan Chilli Veinal Mottle Virus pada Cabai (Cupsictmz annuurn L.). Dibimhing oleh SRIANI SUJIPRIHATI dan SRI HENDRASTUTI NIDAYAT.
Cucumber nzosuic virzrs (CMV) d m chil!i veil7ul tno//le virzrs ( ChiVMV) merupakaii dua v i m pcnting yang menginfe~si cabai di Asiz dan telah di1apod:an tersebar luas di Indonesia. Salali satu strategi untuk mengendalikau kedua virus tersebut adaleh penggunaan varietas tahan. Petdkitan varietas tahan CMV dan ChiVMV akm lebih mudah jika tersedia informasi kendali genetik karakter yang diinginkan. Studi genetik untuk mempelajari parameter genetik suatu sifat, lnenduga nilai daya gabung: menduga nilai heterosis dan menddgz nilai heritabilitas dapat dilakukan menggunakan analisis silang setengah dialel.
Penelitian meliputi tiga percobaan yaitu: (1) evaluasi karakter hortikultura,
(2) evalusi respon tanaman terhadap CMV, dan (3) evalusi respon tanaman terhadap ChiVMV.
Percobaan 1 bertujuan untuk memperoleh (1) genotipe yang memiliki penampilan hortikultura terbaik, dan (2) informasi tentang parameter genetik, nilai DGU, nilai DGK dan nilai heterosis karakter hotikultura. Percobaan ini menggunakan 21 genotipe yang terdiri atas 6 tetua dan 15 hibridanya. Analisis data dilakukan menggunakan pendekatan Nayman, Griffing dan Fehr.
Percobaan 2 dan 3 bertujuan untuk memperoleh (1) informasi tentang parameter genetik, nilai DGU, nilai DGK dan nilai heterosis karakter ketahanan terhadap CMV dan ChiVMV, dan (2) genotipe tnhan CMV dan ChiVMV. Kedua percobaan ini masing-masing menggunakan 21 genotipe yang terdiri atas 6 tetua dan 15 hibridanya. Isolat yang digunakan adalah CMV isolat 02 dan ChiVMV isolat Cikabayan. Analisis setengah dialel dan pendugaan heterosis karakter ketahanan terhadap CMV dan ChiVMV dilakukan menggunakan h a i l ELISA menggunakan pendekatan Hayman, Griffing dan Fehr. Pengelompokkan tipe ketahanan menggunakan metode yang dikemukan oleh Dolores.
Berdasarkan hasil percobaan diperoleh informasi bahwa interaksi gen pada karakter jumlah buah dan overdonzinance pada karakter bobot buah per tanaman: umur panen, ketahman terhadap CMV &n ketahanan terhadap ChiVMV menyebabkan heterosis dan dapat dimanfaatkan untuk pembentukan varietas hibrida. Efek DGU a k m mempengaruhi efek DGK dan efek DGK adalah konlponen yang bertanggungjawah pada peningkatan heterosis. Nilai heritabilitas arti luas pada semua karakter hortikultura yang diamati, ketahanan terhadap CMV dan ketahanan terhadap ChiVhN memiliki kriteria yang tinggi. Nilai heritabilitas arti sempit berkisar dari rendah sampai tinggi. Ketahanan cabai terhadap CMV dikendalikan 1 kelompok gen resesif dan ketahanan cabai terhadap ChiVMV dikendalikan 1 kelompok gen dominan. Tetua IPB C14 dan IPB C10 dapat dijadikan tetua donor ketahanan terhadap CMV dan ChiVMV dan IPB C1 dapat dijadikan tetua donor ketakaaad terhadap CMV. Tiga hhrida Serpotznsi dijadikan hibrida tahan CMV dan ChiVMV serta 6 hibiida berpotensi dijadikan hibrida tahan ChiVMV.
O
Hak Cipta milik Institut Pertanian Bogor, tahun 2007Hak cipta dilindungi Undang-undang
1 Dilarang mengutip sebagian atau selumh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b Pengutipan tidak memgikan kepentingan yang wajar IPB
STUD1 GENETIK KARAKTER HORTIKULTURA DAN
KETAHANAN TERHADAP CUCUMBER MOSAIC VIRUS
DAN CHILLI VEINAL MOTTLE VIRUS
PADA
CABAI (Capsicum annuutn
L.)
AGUS RIYANTO
Tesis
sebagai salah satu syarat untuk memperaleh gelar Magister Sains pada
Departemen Agronomi d m Hortikultura
SEKOLAH PASCASARJANA
INSTITUT P E R T A N M BOGOR
Judul Tesis : Studi Genetik Karakter Hortikultura dan Ketahanan terhadap
Cucumber Mosaic Jrirus dan Chilli Veinal Mottle J'irus pada Cabai (Capsicum anntiunz L.)
Nama Mahasiswa : Agus Riyanto
Nomor Pokok : A35 1040071
Disetujui, Komisi Pembimbing
Dr. Ir. Sriani Suiiprihati, MS. Dr. Ir. Sri Hendrastuti HidGat. MSc.
Ketua Anggota
Diketahui,
Ketua Progam Studi Agronomi Dekan Sekolah Pascasarjana IPB
PRAKATA
Puji syukur atas segala ridho, rahmat dan hidayah Allah SWT sehingga
tulisan ini dapat penulis selesaikan. Tesis yang berjudul Studi Genetik Karakter
fiorlikultura dan Ketahanan terhadap Cucunzber Mosaic Vil.lrs dan Chilli Veinal Moltle I*'iru.s pada Cabai (Capsicum annuun: L.) merupakan kelenkapan tugas akhir untuk men~peroleh ge:ar Magister Sains pada Sekolah Pascasrjana IPB.
Penelitian dan penulisan tesis ini di bawah bimbingan Dr. Ir. Sriani
Sujiprihati, MS. dan Dr. Ir. Sri Hendrastuti Hidayat, MSc. Terimakasih dan
penghargaan penulis sampaikan atas bimbingan dan arahan yang diberikan sejak
penyusunan, perencanaan dan pelaksanaan penelitian hingga selesai penulisar~.
Terimakasih dan penghargaan juga disampaikan kepada Dr. M. Syukur SP, MSi.
atas masukan dan saran sebagai penguji luar komisi szat sidang tesis.
Terimakasih juga diucapkan kepada SPMU TPSDP BATCH I11 Universitas
Jenderal Soedirman selaku pemberi beasiswa selama penulis menempuh studi dan
Tim Program Penelitian Kerjasama Faperta-AVRDC 2006 yang diketuai oleh Dr.
Ir. Sri Hendarstuti H, M.Sc. se& Dr. Ir. Sriani Sujiprihati, M.S. (anggota Bidang
Pemuliaan Tanaman) selaku penyandang dana penelitian.
2enghargaan penulis sampaikan kepada Rektor Universitas Jenderal
Soedirman, Dekan Fakultas Pertanian dan Ketua Program Studi Pemuliaan
Tanaman yang telah memberikan izin belajar serta Rektor Institut Pertanian
Bogor. Direktur Sekolah Pascasajana IPB dan Ketua Program Studi Agronomi
Sekolah Pascasajana IPB yang telah menerima penulis untuk melanjutkan S2.
Disatnpicg itu, terimakasih disampaikan kepada Kepala Bagian Genetika
dan Pe~nuliaan Tananan Departemen Agronomi dan Hortikultura IPB atas
bantuan bahan genetik dan fasilitas di Laboratorium Pendidikan Pemuliaan
Tanaman dan Kepala Laboratorium Virologi Departemen Proteksi Tanaman IPB
atas bantuan fasilitas penelitian di Laboratorium Virologi dan rumah kaca di
Cikabayan.
Tidak ketinggalan terimakasih dan penghargaan diucapkan kepada asisten
peneliti di Labdik Pemuliaan Tanaman Siti Manviyah, SP. dan Suhartini, SP.;
teknisi lapang di Kebun Tajur I1 Bu Ade d m Pak Kardi; teknisi Laboratorium
Rahmi Yunianti, Zahratul Millah, Ismi Yulianah, Latifah dan Siti Hafiah; dan
Staf fzngajar, Laboran dan Teknisi di Program Studi Pemuliaan Tanaman Faperta
UNSOED atas dukungai~ dan segala bantuan yang diherikan. Ungkapan
terimakasih dan penghargaan juga disampaikan kepada Ibunda Suripah, Ayahanda
Adi Pramono dan keluarga di Pasir serta Pekuncen atas doa. dukungan dan kasih
sayangnya.
Semoga karya tu!is ini dapat bermanfaat bagi pengembangan iimu
pengetahuan.
Bogor, September 2007
RIWAYAT
HIDUP
Penulis dilahirkan di Banyumas pada tanggal 2 Agustus 1977. Penulis
merupakan anak tunggal dari pasangan Adi Pramono dan Suripah.
Pendidikan sarjana ditempuh di Program Studi Pemuliaan Tanaman
Fekultas Pertanian Universitas Jenderal Soedirman Purwokerto, lulus tahun 2002.
Pada tahun 2004 ~ e n u l i s diterima di Program Studi Agronom; Sekolah
P a s c a s a j a n ~ IPB. Beasiswa pendidikan pascacajana diperoleh dari SPMU
TPSDP Eatch 111 Universitas Jenderal Soedirman.
Penulis bekeja sebagai staf pengajar kontrak pada Program Studi
Pemuliaan Tanaman Fakultas Pertanian Universitas Jenderal Soedirman
Purwokerto sejak tahun 2003 dan pada tahun 2004 diangkat sebagai staf pengajar
DAFTAR IS1
Halarnan
. .
DAFTAR TABEL...
XIIDAFTAR CiAMBAR
...
xivDAFTAR LAMPIRriN
...
xvI FENDAHULUAN
Latar Belakang
. .
...
1 Tujuan Penel~t~an .... .
3 Ruang Lingkup Penel~t~an...
4I1 TINJAUAN PUSTAKA
Klasifikasi; 3otani dan Syarat Tumbuh Cabai
...
6Cucumber Mosaic Virus (CMV)
...
8Chilli Veinal ,'doi/le Virus ( C h i V W )
...
9. . .
...
Analls~s Sllang Dialel 10
Heterosis
...
13111 ANALISIS SILANG SETENGAH DIALEL KARAKTER
HORTIKULTURA CABAI (Capsicun~ annuunz L.)
Abstrak
...
15 Pendahuluan...
15 Bahan dan Metode...
17...
Tempat dan Waktu 17
Bahan
...
17...
Metode 17
Hasil dan Pernbahasan
...
20...
Pendugaan Parameter Genetik 21
...
Daya Gabung 27
...
Heterosis Dan Heterobeltiosis 31
IV RESPON TANAMAY CABAI (Capsinta annuum L . ) TERHADAP
CUCUMBER I ~ ~ O S A I C VIRUS DAE CHILLI VEINAL MOTTLE VIRUS
Abstrak
...
36 Pendahuluan...
36 Bahan dan Metode ... 37...
Ternvat dan Waktu 37
Bahan
...
37 Metode...
37 Hasil dan Pembahasan...
397
S1mpulan
...
4-V ANALISIS SILANG SETENGAH DIALEL KARAKTER
KETAHANAN CABAI (Cupsicurn annuunz L.) TERHADAP
CUCUMBER MOSAIC J/(RUS DAN CHILI VEINAL MOTTLE VIRUS
Abstrak
...
44Pendahuluan ... 44
Bahan dan Metode
...
46Tempat dan Waktu
...
46Bahan
...
46Metode
...
46...
Ilasil Dan Pembahasan 48...
...
Pendugaan Parameter Genetik 1 48 Daya Gabung...
52Heterosis Dan Heterobeltiosis
...
54Simpulan
...
55Daftar Pustaka
...
56VI PEMBAHASAN UMUM
...
58VII SIMPULAN UMUM DAN SARAN Saran
...
66VIII DAFTAR PUSTAKA
...
68DAFTAR
TABEL
Halaman
1 Genotipe cabai bahan penelitian
...
172 Persilangan setengah dialel dan selfing enam tetua
...
IS3 Jenis dan dosis pup& serta ..valitu pemupukas tanaman cabai ... 20
4 Ku?.drat ten& karakter horiikultur; yalig riialuati pada 21 ger~otipe
cabai
...
205 Pendugaan parameter genetik karakter hotikultura menggunakan analisis silang setengah dialel
... ...
226 Nilai (Wr+Vr) karakter hortikultura 6 tetua cabai
...
247 Kuadrat tengah daya gabung karakter hortikultura yang diamati pada 21 genstipe cabai
...
288 Komponen ragam DGU, DGK dan proporsi ragaln DGK terhadap ragam DGU karakter hortikultura yang diamati pada 21 genotipe cabai
...
289 Nilai tengah, DGU dan DGK karakter hortikultura yang diamati pada 21 genotipe cabai
...
...
3010 Nilai duga heterosis dan heterobeltiosis karakter hortikultura yang diamati pada 15 genotipe cabai
...
3 111 Kespon 21 genotipe cabai terhadap CMV dan ChiVMV
...
4112 Kelas CMV dan kelas ChiVMV berdasarkan nisbah nilai absorban
ELISA ... 47 13 Kuadrat tengah analisis ragam karakter ketahanan berdasarkan kelas
CMV dan kelas ChiVMV pada cabai
...
4814 Pendugaan parameter genetik karakter ketahanan berdasarkan kelas CMV dan kelas ChiVMV
...
4915 Nilai (WrtVr) karakter ketahanan berdasarkan kelas CMV dan kelas
ChiVMV 6 tetua cabai
...
...
...
3016 Kuadrat tengah daya gabung karakter ketahanan berdasarkan kelas CMV dan kelas ChiVMV pada cabai
...
5217 Kornponen ragam DGU, DGK dan proporsi ragarn DGK terhadap ragam DGU karakter ketahanan berdasarkan kelas CMV dan kelas ChiVMV
pada cabai
...
5218 Respon 21 genotipe cabai terhadap CMV clan ChiVMV. nilai DGU Jan nilai DGK karakter ketahanan berdasarkan M a s CMV dan kelas
ChiVMV
...
5319 Nilai duga heterosis dan heterobeltiosis karakter ketchanal berdamkan
kelas CMV dan kelas ChiVMV pada 15 hibrida cabai
...
5421 Setengah dialel
...
75DAFTAR GAMBAR
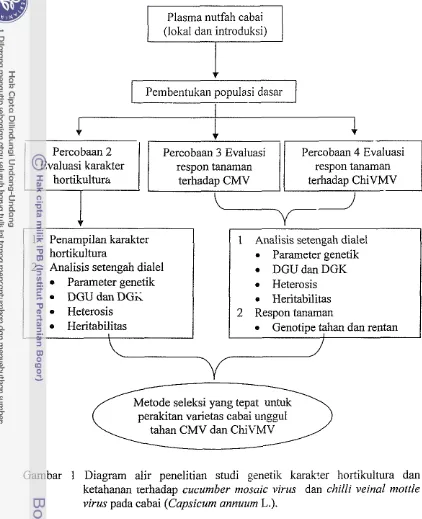
1 Diagram alir penelitian studi genetik karakter hortikultura dan ketahanan terhadap cucumber ntosuic virus dan cl7illi veinal moltle virus pada cabai
(Cqsicz!rn dnnz!!u~z
LG
...
52 S e v n g tetua ~nenggu~lakan sungkup kasa
... ... ... ... .
183 Teknik persilangan buatan pada cabai
...
194 Hubungan peragam (Wr) dan ragam (Vr) karakter bobot buah per tanaman, jumlah buah dan panjang buah
...
255 Hubungan peragam (Wr) dan ragam (Vr) karakter diameter buah dan umur panen
...
276 Teknik inokutasi virus secara mekanis pada cabai
...
387 Gejala infeksi CMV pada daun cabai
...
39o
Gejala infeksi ChiVMV pada daun cabai...
39DAFTAR LAMPIRAN
Halaman
. .
1 Metode a n a l ~ s ~ s setengah dialel
...
752 Korelasi karakter hortikultura, ketahanan terhadap CMV dan ketahanan terhadap ChiVMV yang diamati pada 21 genotipe cabai
...
803 Nilai tengah tetua, hibrida, ni!ai heterosis d2n heterobeltiosis karakter bobot buah per tanaman 15 hibrida yang diuji
...
904 Nilai tengah tetua, hibrida, nilai heterosis dan heterobeltiosis karakter
. . .
jumlah buah 15 hibrida yang dlujl...
815 Nilai tengah tetua, hibrida, nilai heterosis dan heterobeltiosis karakter
. . .
panjang buah 15 hibrida yang dlujl...
816 Nilai tengah tetua hibrida, nilai heterosis dan heterobeltiosis karakter diameter buah 15 hibrida yang diuji
...
827 Nilai tengah tetua hibrida, nilai heterosis
. .
. . .
dm heterobeltiosis karakter Umur panen 15 hlbnda yang ~ I U J I...
828 Nilai tengah tetua, hibridq nilai heterosis dan heterobeltiosis karakter ketahanan berdasarkan kelas CMV 15 hibrida yang diuji
...
839 Nilai Tengah Tetua, Hibrida, Nilai Heterosis dan Heterobeltiosis karakter ketahanan berdasarkan kelas ChiVMV 15 hibrida yang diuji
...
8310 Penampilan buah cabai hibrida tahan CMV dan ChiVMV
...
841 1 Penarnpilan buah cabai hibrida tahan ChiVMV
...
84I PENDAHULUAN
Latar Belakang
Cabai (Cnpsiczm~ spp) merupakan sayuran penting di dunia dan termasuk spesies pertama yang ditemukan telah digunakan manusia di seluruh dunia (Berke
200U). Luas pertanaman cabai dunia diperkirakan sekitar 1.25 juta hektar dzngan
pertumbuhan rata-rats 0.5%. Produksi cabai dunia mencapai 11 2 juta ton dan 4.3
juta ton dihasilkan ole11 nzgara-negara Asia seperti India, Indonesia, China dan
Korea (Xuefeng 1999).
Lima spesies cabai yaitu Capsicum pubescens, C. baccafum; C. chinense, C. ~ustescens dan C. annuum telah didomestikasi, dideskripsi dan dipelajari secara intensif (Greenleaf 1986; Pickersgill 1997; Berke 2000). Cabai merah
(C. annuum) merupakan spesies j.;ni, dibudidayakan paling luas dan merupakan
spesies yang bemilai ekonorni penting dari &:nus Capsicum, yang memiliki tipe buah rnanis hingga sangat pedas (Rubatzky & Yamaguchi 1997; Bosland &
Votava 2000; Zhang 2005).
Cabai dapat dikonsumsi dalam benfuk buah segar, kering atau bentuk
olahannya. Satu juta orang diperkirakan rnengkonsumsi cabai segar atau bentuk
olahannya dalam satu hari (Zhang 2005). Cabai kaya akan vitamin C, A dan B,
potasium, fosfor dan kalsium (Xuefeng 1999; Bosland & Votava 2000).
Kandungan kimia cabai merupakan bagian penting dalam obat-obatan, pewarna
makanan dan kosmetika (Taychasinpitak & Taylviya 2003; IISR 2006).
Luas panen cabai merah Indonesia rnencapai 110 170 hektar dengan
produksi 714 705 ton per tahun pada tahun 2004 dan menurun rnenjadi 103 531
hektar dengan produksi 661 730 ton per tahun pada tahun 2005 (Deptan 2007).
Luas panen cabai merah tidak diimbangi dengan produktivitas yang tinggi.
Produktivitas cabai merah Indonesia masih jauh dari potensi produksinya yang
mencapai 12 ton per hektar (Duriat 1996) dan mengalami penurunan dari 6.49 ton
per hektar pada tahun 2004 menjadi 6.39 ton per hektar pada tahun 2005.
Produktivitas cabai merah di Indonesia yang r~ndah dan mengalami penurunan
antara lain disebabkan o ~ e h pengguliaan varietas berdaya hasil rendah dan atau
Penyakit pada cabai dapat diakibatkan oleh infeksi bakteri, cendawan dan
virus. Infeksi virus pada cabai dapat menyebabkan kegagalan panen (Greenleaf
1986) dan paling sulit dikendalikan (Suzuki el 01. 2003). Sulyo el ul. (1993) menyebutkan bahwa sekitar 40 virus mampu menginfeksi tanaman cabai,
sedangkan Duriat el rrl. (1995) nlenyatakan bahwa sekitar 45 virus yang berbeda mampu menginfeksi tanaman cabai. Cuczmlher mosuk virus (CMV) dan chilli veitzul mottle virus (ChiVMV) merupakan 2 virus penting yang menginfcksi cabai di Asia (AVRDC 1994, 2006, 2001; Berke 2002) dan telah dilaporkan tersebar
luas di Indonesia (Sulyo el al. 1995; Taufik et 01.2006; Ditlinhorti 2066).
Infeksi CMV dan ChiVMV menjadi faktor pembatas produksi cabai dan
menyebabkan ketidakstabilan produktiviras serta rnenurunkan kualitas dan
kuantitas hasil cabai. Infeksi CMV menyebabkan buah berkerut, bergelombang,
benvarna pucat hijau kekuningan dan terkadang terbentuk lesiu kecil (Cerkauskas
2004a). Kehilangan hasil akibat infeksi CMV dapat rnencapai 90% tergantung
varietas tanaman dan strain CMV yang menginfeksi (AVRDC 2000). Hasil
penelitian Nilamsari et al. (1998) rnenyatakan bahwa infeksi CMV dapat menurunkan bobot buah per tanaman sarnpai 82.30%.
Infeksi ChiVMiJ rnenyebabkan bunga layu dan rontok sebelurn
pernbentukan buah. Jika terbentuk buah maka buah menjadi salah bentuk
(Cerkauskas 2004b). Infeksi ChiVMV dapat rnenyebabkan kehilangan hasil
san~pai 100% (AVRDC 2003), sernentara Subekti et al. (2006) melaporkan bahwa di Indonesia kehilangan hasil akibat infeksi ChiVMV mencapai 65%.
Sh-ategi pengendalian penyakit akibat infeksi CMV dan ChiVMV menjadi
bagian penting pada produksi cabai. Upaya pengendalian CMV dan ChiVMV
yang paling umum digunakan petani adalah penggunaan insektisida untuk
mengendalikan kutudaun. Cucumber mosaic virus dan ChiVMV adalah virus yang ditularkan secara non persisten oleh kutcdaun. Kutudaun membutuhkan
waktu makan akuisisi dan infeksi yang singkat untuk menularkan CMV dan
ChiVMV. Oleh karena itu, strategi pengendalian dengzn insektisida rnenjadi tidak
Ctictmzher t~zosuic 1-irzcr dan chilli 1leit7al nlortle virzrs marnpu n~enginfeksi cabai secara tunggal atau bersama-sama, sehingga pengendalian secara
konvensional selnakin sulit dilakukan. Salah satu strategi untuk niengendalikan
ked~la virus tersebut adalah penggunaan varietas tahan karena merupakan cara
yeng efektif, murah dan aman. Perakitan varietas baru yang meiniliki ketahanan
terhadap berbagai v i n ~ s (multiyle-virus resistance) n~enjadi penting sebagai pemecahan masalah (Grubs cl al. 2003). Sayangnya, \arietas cabai tahan CMV dan ChiVMV yang lnemiliki daya hasil yang baik belum tersedia (Taufik et al. 2006; Subekti et al. 2006).
Perakitan varietas tahan CMV dan ChiVMV akan lebih n~udah jika tersedia
informasi kendali genetik karakter yang diinginkan. Perilaku genetik gen-gen
karakter yang diinginkan dapat dipelajari melalui studi gendik menggunakan
metode analisis silang dialel. Metode ini secara eksperimental merupakan
pendekatan yang sistematik dan secara analitik merupakan evaluasi genetik
menyeluruh yang bergma dalarn mengidentifikasi persilangan bagi potensi seleksi
yang terbaik pada generasi awal (Johnson 1963; Khan & Habib 2003). Analisis silang dialel dapat digunakan untuk mempelajari kendali genetik suatu sifat,
menduga nilai daya gabung dalam hihrida serta membantu pernulia dalam
mcningkatkan dan menyeleksi populasi segregan (de Sousa & Maluf 2003).
Selain itu, dapat diperoleh informasi nilai heterosis dan heritabiiitas.
Tujuan Penelitian
Tujuan akhir dari seluruh percohaan yang dilakukan adala;~ untuk
memperoleh metgde seleksi yang tepat dalam perakitan varietas cabai unggul
tahan CMV dan ChiVMV. Tujuan tersebut dapat dicapai dengan melakukan
beberapa percobaan dengan tujuan khusus:
1 Menlpelajari paranleter genetik pada cabai untuk karakter hortikultura,
ketahanan terhadap Ch4V dan ketahanan terhadap ChiVh4V.
2 Menduga nilai daya gabung umum: daya gabung khusus dan nilai heterosis karakter hortikultura ketahanan terhadap CMV dan ketahanan terhadap
ChiVM;' pada cabai.
Ruang Lingkup Penelitian
Penelitian meliputi tiga percobaan yaitu: (1) evaluasi karakter hortikultura,
(2) evaluasi respon tanaman terhadap CMV. dan (3) evaluasi respon tanaman
terhadap CliiVMV. Diagram alir penelitian disajikan pada Gambar 1.
Infonnasi yang didapat dari percobaan 1 adalah ( I ) genotipe yang memiliki
penampila11 hortikultura terlaik, dan (2) informasi tentang parameter genetik, nilai
DGU, nilai DGK dan nilai heterosis karakter hot~k~~ltura. Pada percobaan 2 dan 3
diperoleli (1) infolliiasi tentang parameter genetik, nilai DGU, nilai DGK d m nilai
heterosis karakter ketahanan terhadap CMV dan ChiVMV, dan (2) genolipe tahan
CMV dan ChiVMV. Tujuan akhir selurnh percobaan yang dilakukan adalah untuk
mernperoleh metode seleksi yang tepat dalam perakitan varietas cabai unggul
Plasma nutfah cabai (lokal dan introduksi)
Pembentukan populasi dasar
I
Percobaan 3 Evaluasi Percobaan 4 Evaluasi
1
E z F z : t e r1
terhadap ChiVMV I
hortikultura
Analisis setengah dialel Parameter genetik
= DGU d a n D G i Heterosis Heritabilitas
1 Analisis setengah dialel Parameter genetik DGUdanDGK Heterosis Heritabilitas 2 Respon tanaman
Genotipe tahan dan rentan
Metode seleksi yang tepat untuk perakitan varietas cabai unggul
[image:21.539.36.458.31.550.2]tahan CMV dan ChiVMV
I1 TINJAUAN PUSTAKA
Klasifikasi, Botani dan Syarat Tumbuh Cabai
Cabai (Capsicurn annutrm L.) dibudidayakan pertama kali di Meksiko dan
A~nerika Tengah. Cabai diintroduksikan oleh Portugis ke Indonesia sekitar
350-500 tahun yang lalu (Berke 2004. Cabai merah diklasifikasikan daiam divisi
Magnoliophyts, kelas Magcoliopsidq ordo Sulanes, famili Soianaceae. gev.ls
Capsicum dan spesies Capsicum annuum (Boslanci & Votava 2000, Der-ra 2000).
Cabai mempakan jenis tanman setahun berbentuk perdu. Cabai memiliki
akar tunggang ( a k a primer) yarlg bercabang-cabang ke samping dengan akar-akar
rambut ( a k a sekunder). Akar tananlan menyebar dengan panjang 30 sampai
50 cm dan kedalaman 30 sampai 60 cm. Cabai merah menliliki batang tunggal
berdaun 8 sampai 15 helai sebelum membentuk bunga pertama (Bosland &
Votava 2000). D a m cabai merah mempakan daun tunggal yang tumbuh pada
tunas-tunas samping, batang utama dan c a b ~ z g tersusun secara spiral. Helai daun
cabai merah berbentuk ovate atau lanmolate dan benvama hijau atau hijau tua
(Kusandriani 1996b).
Cabai merah memiliki tipe bunga tunggal dan tumbuh pada uiung mas.
Mahkota bunga benvarna putih, memiliki lima sampai enam helai petal dan
berdiameter 8 sampai 15 mm. Cabai merah memiliki 5 sampai 7 benang sari
dengan kepala sari benvarna bim pucat sampai ungu. Putik terdiri atas indun2
telur, tangkai putik dan kepala putik. Indung telur berdiameter 2 sampai 5 mm.
Indung telur memiliki 2 sampai 4 carpel (locules). Panjang tangkai putik
3,s sampai 6,s mm. Diameter kepala putik lebih besar dari diameter tangkai sari.
Bunga pertama terbentuk pada umur 23 sampai 31 hari setelah tananl (HST).
Buah pertama terbentuk pada umur 29 sampai 40 HST dan buah matang pada
umur 45 hari setelah pembuahan. Tanaman cabai mempakan tanaman menyerbuk
sendiri, akan tetapi penyerbukan silang secara alami di lapang dapat terjadi
dengan bantuan kumbang dengan persentase kejadian antara 7.6% sampai 36.8%
Tanaman cabai memiliki jumlah kromosom somatik 2n=24 (Berke 2000).
Penyimpanpan jumlah kromosom 2n=26 ditnnukan pada spesies C. ciliaiun~ asal
Arnerika Selatan bagian barat, spesies liar di Brasil dan C. lanceolulutn asal
Guatemala (Nankui & Bosland 1997) serta cabai manis (Bosland & Votava 2000).
Buah cabai merah di Asia terutama bertipe cayenne dengan dua ukuran,
panjang atau pendek. Buah cabai merah berukuran panjang mzmiliki bentuk buah
lurus, wama buah muda hijau atau hijau gelap, warna buah masak merah menyala
atau merah gelap, panjang buah antara 9 sampai 15 cm dan kepedasannya berkisar
antara tidak pedas sampai pedas medium. Buah cabai merah berukuran pendek
memiliki bentuk buah lums, wama buah muda hijau atau hijau gelap, wama buah
m a s k merah menyala atau merah gelap, panjang buah antara 2 sampai7 cm d m
kepedasannya berkisar antara pedas medium sampai sangat pedas (Berke 2002).
Cabai merah membutuhkan suhu optimum 20 OC sampai 30 OC. Ketika suhu
turun menjadi 15 OC atau naik menjadi 30 OC untuk jangka w a h yang lama maka
pertumbuhan akan terhambat dan produksi akan menurun. Panjang hari tidak
menpengaruhi pernbentukan bunga atau buah. Pertumbuhan tanaman terbaik
diperoleh ketika cabai ditanam pada lahan dengan jenis tanah liat atau liat
berlumpur dengan kapasitas lapang yang baik. Pada tipe tanah lainnya, tanaman
ini memerlukan pengairan yang baik untuk dapat tumbuh. Cabai merah
membutuhkan pH tanah antara 5.5 dan 6.8 (Berke et a1 2005).
Cabai dapat terserang oleh hama dan penyakit. Hama utama cabai di
Indonesia adalah ulat tanah (Agrolis ipsilon), thrips (Thrips parvipnus). kutudaun
persik (Myz~is persicae), tungau teh h n i n g (Polyphagofarsonemus lalzrs). ulat
grayak (Spodoplera liftrra) dan ulat buah (Helieoverpa armigera). Penyakit utama
yang menyemng cabai di Indonesia adalah layu bakteri (Ralsonia solanacear~rm),
bercak daun serkospora (Cercospora capsici), penyakit patah batang
(Choanephora cucubilarum), penyakit busuk buah (Colletrotrichum capsici,
C. gloesporioides dun C. acutaium), CMV, ChiVMV, PVY d m TMV (Ditlinhorti
Cucurnber Mosaic Virus (CMV)
Cucumber mosaik virus (CMV) temlasuk dalam famili Bromoviridcie genus
Cztcumovirus. Partikel CMV berbentuk isohedral dengan diameter 29 nm. Tiap
partikel memiliki 180 subunit protein dengan berat molekul 24 kDa (Hull 2002).
Genom CM?' terdiri atas 5 komponen utas tunggal RNA dan 1 sub genom. Genom tersebut adalah RNAl {3,4 kbcb), RNA2 (3,l kb) dan RNA3 (2,2 kb). S L I ~
genom RNA4 n e m i l i i ukuran i,O kb (Agrios 1997). Virw ini memiliki titik
panas inaktivsi 55 sanlpai 70 OC dm ketahanan invitro kurang dari 1'3 hari
(Palukaitis & Garcia-Arenal 2003) dan titik batas pengenceran l:lo4 (Matthews
1991).
Cucumber mosuic virus tersebar di selurh dunia dengan kisaran inang
yang luas. Inang virus ini meliputi lebih dari 1200 spesies pada lebih dari 100
famili dikotiledon dan monokotiledon. Penularan CMV dapat dilakukan dengan
cara mekanis, melalui serangga vektor, benih pada lebih dari 20 spesies tanaman
dan tali putri (Palukaitis & Garcia-Arena1 2003). Pada cabai, CMV tidak dapat
ditularkan melalui benih (Cerkauskas 2004a). Cucumber mosaic virus ditularkan
secara non persisten oleh i e b i dari 800 sijesies kutudaun (Aphididae) dalam
33 genera. Myzuz persicae dan Aphis gossypii adalah 2 serangga vektor
terpenting. Serangga vektor memhutuhkan waktu makan akuisisi dan inokulasi
yang singkat.
Gejala CMV beragam tergantung spesies tanaman inang dan straimya.
Gejala umurn yang nampak adalah mosaik, belang, klorosis, nekrosis, kerdil serta
perubahan bentuk dam dan buah (Agrios 1997). Infeksi CMV pada cabai
menimbulkan gejala pengkerdilan, mosaik hijau gelap pada daun, perubahan
bentuk daun, pola nekrosis pada daun, perubahan warna dan bentuk buah serta
lesio pada buah cabai (Cerkauskas 2004a).
Pewarisan sifat ketahanan cabai terhadap CMV bersifat poligenik dan
beragam tergantung umur tanaman dan strain virus (Greenleaf 1986). Sifat
ketahanan cabai terhdap CMV dikendalikan oleh 1 hingga 3 gep resesif
(Rustikawati 2000). Herison er al. (2005) melaporkan bahwa ketahanan cabai
terhadap CMV dikendalikan oleh gen sederhana, resesif dengan aksi gen dominan
3 gen mayor pengendali karakter ketahanan CMV pada cabai dengan aksi gen
resesif tidak penuh yaitu 2 gen resesif dan 1 gen dominan yang bekerja secara
epistasis.
Clzilli Veinal Mottle firus (ChiVMV)
Chilir veinal niottle virus fChiVhlV) pertama kali di!aporkan pada
C. annulcnt di Malaysia oleh Bumett pada tahun 1947. Vilus ini tersebar di Korea, Malaysia, Filipina, Taiwan dan Thailand (Brunt 1992). Dilaporkan bahwa
ChiVMV telah tersebar luas di Indonesia (Sulyo et al. 1995; Taufik et al. 2006; Ditlinhorti 2006).
Chilli veinal nzoltle virus termasuk dalam genus potyvirus. Chilli veinal mottle virus memiliki partikei berbentuk filamentus; tidak memilid zelubung dan selall~ flexuous dengan panjang 720 nm dan lebar 12 nm; memiliki titik panas
inaktivasi 60 OC dan ketahanan invitro 7 hari (Brunt 1992). Chilli veinal mottle virus memiliki genom utas tunggal RNA yang diekspresikan sebagai poliprotein dengan berat molekul 9.7 kb. Chilli veinal mottle virus dapat membentuk badan inklusi berbentuk cakra (pinwhell) (Hull 2002).
Penularan ChiVMV dapat dilakukan dengan cam mekams, penyambungan
dan serangga vektor secara non persisten. Serangga vektor ChiVMV adalah Aphis craccivora, A. gossypii, A. spiraecola, &us persicae, Toxoptora citrida dan
Rhopalosiphum maidis (Brunt 1992).
Inang alarni ChiVMV adalah cabai (Capsicum spp), tomat (Lycopersicum esculentum) dan African eggplant (Solanutn aethiopicum) (Nono-Womdin 2005).
Chilli veinal mottle virus mampu menginfeksi tanaman C. annuunt, C. fiustescens, C. chinensis, Nikotiana tabacum, N. glutinosa, h:. megalosiphon, N. benthamiana, N. sylvestri.~, Physalis jloridana,
P.
minima, Datura stramonium, D. metel, Lycopersicon esculentunt, Nicandra physalodes. Petunia hybrida dan Solanum melongena.Infeksi ChiVMV pada cabai merah menimbulkan gejala belang hijau gelap
pada dam. penunman ukuran dan perubahan bentuk dam, perhunbuhan tanaman
terhmbat, timbul bintik hijau gelap pada cabang &an batang. bunga layu dan
rontok sebelum pembentukan buah dan pembahan bentuk buah (Cerkauskas
Karakter ketahanan terhadap ChiVMV pada cabai dilaporkan dikendalikan
oleh sepasang gen resesif dengan kemungkinan keterlibatan sejumlah gen resisten
independen (Chew dalam Green & Kim 1994), sedangkan Caranta & Palloix
(1995) melaporkan bahwa sifat ketahanan terhadap ChiVMV pada cabai
dikendalikan oleh 2 gen independen dengan efek gen dominan. Menurut Chew
dan Ong ketahanan terkadap ChiVMV dikendalikan oleh sepasang gen resesif
(Shah & Khalid 2031). Hasil penelitiai~ teitilru menyatakan bahwa kwakter
ketahanan cabai terhadap ChiVMV dikendalikan oleh 1 gen yang bersifat
dominan sempurna atau 1 gen dengan aksi gen ale1 ganda (Millah 2007).
Analisis Silang Dialel
Silang dialel adalah seluruh kombinasi persilaigan yang mungkin dilakukan
dalam satu kelompok individu untuk menghasilkan keturunan FI (Chahal & Gosal
2003). Sistem persiiangan dialel berarti 1 dari kelompok genotipe dipilih dan
disilangkan dengan genotipe lain dalam kelornpok tersebut. Prosedur tersebut
mernberikan kornbinasi persilangan yang rnaksimum. Hasil persilangan dibedakan
menjadi 3 kelompok, yaitu (1) tetua; (2) satu kelompok FI clan (3) satu kelompok
FI resiprokal (GriEng 1956).
Analisis silang dialel dapat dimanfaatkan untuk: (1) menduga nilai daya
gabung umum @GU) dan daya gabung khusus @GK), (2) rnenduga nilai
komponen ragam genetik dan (3) merupakan analisis genetik yang menyeluruh
(de Sousa & Maluf 2003; Dudley et ul. 1999).
Metode silan,: dialel yang dapat digunakan adalah Metode 1 (dialel penuh)
dengan melibatkan populasi tetua, F1 dan FI resiprok (total genotipe: p2);
Metode 2 (setengah dialel) dengan melibatkan populasi tetua, Fl dan tanpa FI
resiprok (total genotipe: [p@+1)/2]); Metode 3 dengan melibatkan F1, FI resiprok
dan tanpa tetua (total genotipe: p-p2); dan Metode 4 dengan melibatkan F1 tanpa
tetua dan tanpa FI resiprok (total genotipe: [p@-1)/2] (Griffig 1956; Singh &
Chaundhary 1979; Roy 2000)
Persilangan dialel yang melibatkan banyak tetua mengakibatkan jumiah
persilangan mznjadi banyak sehingga mengalami kesulitan dalam penanganan
tempat, waktu dan tenaga. Modifikasi persilangan dialel menjadi persilangan
permasalahan tersebut. Persilangan setengah dialel berarti persilangan yang dibuat
hanya untuk menghasilkan tanaman F1 tanpa membuat persilangan resiproknya.
Jika terdapat n tetua, dan masing-masing tetua meliputi s persilangan, maka
jumlah persilangannya adalah [(ns)/2].
Asumsi yans harus dipenuhi dalam penggunaan analisis silang dialel adalah:
(1) merupakan segregasi diploid, (2) tidak tcrdapat pengaruh tetua (tidak ada
perbedaan zntara persilhngan resiprokal), (3) tidak ada interaksi anvara gen-gen
yang tidak satu alel (independen), (4) tidak ada peristiwa multiple alel, (5) tetua
bersifat homozigot (6) gen-gen menyebar secara bebas dianrara tetua (Hayman
1 5 9 ; Singh & Chaundhary 1979; Roy 2000).
Tanaman cabai mempakan tanaman diploid (Greenleaf 1986): dengan
demikian segregasi gen-gen yang terjadi merupakan segregasi diploid. Hal
tersebut memenuhi asumsi segregasi diploid.
Perbedaan antar persilangan resiprokal menandakan bahwa ada pengaruh
tetua betina yang merupakan petunjuk bahwa pewarisan suatu karakter diwariskan
oleh gen-gen ekstrakromosomal (Mather dan Jinks 1971). Pewarisan sifat
ketahanan terhadap CMV tidak dipengaruhi oleh tetua betina (Herison et al.
2005) dan pewrisan sifat ketahanan terhadap ChiVMV tidak dipengaruhi oleh
tetua betina (Millah 2007). Oleh karena itu, pada penelitian ini digunakan analisis
silang setengah dialel melalui pendekatan metode Hayman (1954) dan metode
Griffing (1 956).
Interaksi antara gen-gen yang tidak sealel di dalarn analisis silang dialel
dapat diuji dengan nilai koefisien regresi b dari garis regresi antara Wr (peragam
antara tetua dan keturunan dari array ke-rj terhadap Vr (ragam di dalam array
ke-r). Jika nilai b = 1 maka tidak ada interaksi antara gen-gen tidak sealel (Singh
& Chaudhary, 1979).
Penggaluran genotipe-genutipe cakai dilakukan sedemikian rupa sehingga
asumsi tetua homozigot dapat terpenuhi. Tetua yang mewakili tetua tahan,
moderat dan rentan dipilih untuk memenuhi asumsi gen-gen yang mengendalikan
Jika asumsi silang dialel terpenuhi maka keluaran yang dapat diperoleh dari
suatu analisis silzng dialel Metode Hayman adalah: (1) D yaitu keragaman karena
pengaruh aditif, (2) F yaitu nilai tengah F, genotipe (rata-rata Fr untuk semua
urr-uy): peragam pengaruh aditif dan non aditif pada array ke-r, (3) HI yaitu
keragaman karena pengamh dominansi. (4) 1-11 yaitu perhitungan untuk menduga
proporsl gen negatif dan positif pada tetua, (5) h2 yaitu pengaruh dominan
(sebagai jumlah aljabar dari semur! persilangan saat heterozigous), (6) E yaitu
keragaman karena pengaruh lingkungan, (7) Rata-rata tingkat dominansi,
(8) Proporsi gen-gen dengan pengaruh positif dan negatif di dalam tetua,
(9) Proporsi ge11-gen dominx dan resesif di dalam tetua, (10) Jumlah kelompok
gen yang mengendalikan sifat dan menimbulkan dominansi, (1 1) Heritabilitas arti
luas dan heritabilitas arti sempit (Singh and Chaudhary 1979).
Daya gabung merupakan informasi yang diperoleh dari analisis silang dis;~cl
menggunakan metode Griffrng (1956). Daya gabung merupakan prosedur
pengujian dalam pembentukan varietas hibrida yang digunakan untuk
mempelajari dan mengidentifikasi kombinasi tetua yang menghasilkan hibrida
terhaik. Pendugaan daya gabung menjadi penting dalam identifikasi tetua yang
akan digunakan dalam pembentukan varistas hibrida (Welsh 198 1).
Daya gabung didefinisikan sebagai kemarnpuan tetua memindahkan
performa pada hibridanya jika disilangkan dengan tetua lain (Chahal & Gosal
2003). Daya gabung dibedakan menjadi daya gabung umum (DGU) dan daya
gabung khusus (DGK). Daya gahung umum merupakan ukuran performa
kert1runan suatu genotip; yang disilangkan dengan contoh acak atau genotipe
dengan jumlah besar. Daya gabung khusus merupakan ukuran performa ketumnan
suatu genotipe yang disilangkan dengan genotipe lainnya dan sering diekspresikan
sebagai simpangan perfova yang diduga dengan rata-rata atau daya gabung
umum (Stoskopf et 01. 1993).
Daya gabung umunl diekspresikan pada ketumrlan persilangan suatu galur
murni dengan beberapa genotipe dan terutama mempakan hasil aksi gen aditif.
Daya gabung khusus merupakan ekspresi performa diantara 2 galur murni dan
ditu.ju!:an untuk aksi gen dominan, epistasis dan aditif. Populasi yang telah
Heterosis
I-Ieterosis adalah informasi lain yang dapat diperoleh dari analisis silang
dialel. I-ieterosis dapat didefinisikan sebagai peningkatan perfomla keturunan
dibandingkan kedua tetuanya. Istilah heterosis pertama kali dikemukan oleh Shull
pada tahun 1908. Shull niengambarkan fenomena ini setelah nielakukan
pellgamatan terhadap dorongan heterosigositas pada pembelahan sel,
pertun~buhan dan karakter fisiologi tanaman jagung (Bud& 2002).
Dua hipotesis diajukan sebagai penyebab fenomena heterosis, yaitu
hipotesis dominan dan hipotesis overdonzinance (Fehr 1987). Hipotesis dominan menyatakan bahwa heterosis disebabkan oich gen-gen dominan yang
menguntungkan dari kedua lelua terkumpul pada hibridanya. Hipotesis ini
berdasarkan teori bahwa gen yang menguntungkan untuk tanaman bersifat
dominan dan gen yang merugikan bersifat resesif. Gen dominan yang berasal dari
satu tetua akan dilengkapi oleh gen dominan dari tetua lainnya sehingga tanaman
F1 memiliki kombinasi gen dominan yang menguntungkan dari kedua tetuanya
(Poehlman 1987).
Hipotesis overdominance menyatakan bahwa heterosigot (ala2) lebih vigor dan produktif dibandingkan homosigot (alal atau a2a2). Ale1 a, dan a2 memiliki
h g s i yang berbeda dan penggabungan a, dan az lebih superior jika dibandingkan
homosigotnya (alal atau a2a2). Semakin berbeda fungsi ale1 penyusun heterosigot,
semakin tinggi pembentukan superioritasnya (alaz < ala3 < a)%) (Allard 1960).
Hipotesis overdominunce terbukti pada sifat yang dikendalikan oleh gen tunggal atau sedikit (Virmani al. 2003).
Heterosis di-gmakan untuk perbaikan sifat penting tanaman seperti
peningkatan jumlah biji atau hasil biologis, penurunan umur panen peningkatan
ukuran biji atau buah d m kualitas nutrisi yang lebih baik. Heterosis juga
digunakan dalam perbaikan ketahanan tanaman terhadap lingkungan yang tidak
sesuai dan peningkatan ketahanan terhadap hama dan penyakit. Pemx~faatan
heterosis untuk karakter ketahanan kapas terhadap cotton leaf curl virus telah dilaporkan di India. Hibrida k a ~ a s LHH 144 dinyatakan memiliki tingkat
Heterosis telah dimanfaatkan dalam perakitan varietas hibrida pada tanaman
menyerbuk silang atau menyerbuk sendiri. Pemuliaan hibrida jagung dimulai
sejak tahun 1909 dan telah banyak mengalami kemajuan sampai sekarang (Allard
1960). Hasil penelitian lain bahwa varietas hibrida telah berhasil pada tanaman
gandum (Duvick 1997). sorgum (Wenzel & Pretorius 1999), padi (Virmani et al.
2002) dan sayuran (de Sousa & Maluf 2003).
Fenomena heterosis juga ditemukan pada cabai sehingga mzmungkinkan
dibuat hibrida cabai (Berke 2000). Hibrida cabai yang dirakit diharapkan memiliki
karakter yang lebih baik dari tetuanya untuk sifat kualitas, daya hasil, ketahanan
terhadap hama penyakit dan sifat baik lainnya (Kusandriani 1996a; Herison c: al.
2005). Hasil penelitian Eliyanti et al. (2005) menunjukkan bahwa persilangan antara beberapa genotipe cabai memperlihatkan efek heterosis yang baik dengan
nilai best parent heterosis (heterobeltiosis) berkisar antara 20% hingga 80%. Hasil penelitian laimya menunjukkan bahwa varietas hibrida cabai terbukti
mampu meningkatkan daya hasil dan ketahanan terhadap penyakit seperti CMV,
TMV, antraknosa, dan hawar (Zhang et al. 2003; Hong et al. 2004; Liu et al.
2004; Cui et al. 2004; Wang et al. 2004).
Heterosis dapat dibagi menjadi 2 tipe tergantung genotipe pembanding yang
digunakan (Fehr 1987). Kedua tipe heterosis tersebut adalah: (1) mid-parent heterosis (heterosis), yai'm peningkatan atau penurunan performa hibrida dibandingkan dengan nilai rata-rata kedua tetua; dan (2) best parent iteterosis
(heterobeltiosis), yaitu peningkatan atau penurunan perforrna hibrida
dibandingkan dengan nilai tetua terbaik yang digunakan dala~n k ~ n b i n a s i
111 ANALISIS SILANG SETENGAH DIALEL KARAKTER
WORTIKULTURA CABAI (Capsicutn annuum L.)
Abstrak
Persilangan setengah dialel dilakukan untuk mempelajari parameter genetik, daya gabung dan heterosis. Evaluasi karakter hortikultura dilakukan terhadap . - -
6 tetua dan 15 hibrida menggunakan RAKT. dengan 3 ulangan. Karakter hortikultura yang diamati adalah bobot baah per tanaman. jumlah buah. panjang buah, diameter buah d m um;lr panen. Hasil analisis menunjutkan tidak adz interaksi gen pada semila karakter kecuali pada jurnlah buah. Yarakter bobc: buah per tanaman dipengaruhi oleh aksi gen dominan, sedangkan karakter panjang buah, diameter buah serta umur panen dipengaruhi oleh aksi gen aditif dan dominan. Tingkat dominmsi berada antara dominan tidak lengkap sampai
overdominance. Gen-gen yang mengendalikan pewarisan sifat menyebar tidak merata diantara tetua. Gen-gen dominan lebih banyak pada populasi tetua diperoleh
ads
karakter bobot buah per tanaman. diameter buah dan umur panen. Gen-gen resesif lebih banyak pada populasi tetua diperoleh pada karakter jumlah buah dan panjang buah. Jumlah kelompok gen pengendali karakter bobot buah per tanaman, jumlah buah dan umur panen minimal 2 kelompok gen dan kelompok gen pengendali karakter panjang buah dan diameter buah minimal 1 kelompok gen. Nilai duga heritabilitas arti luas (h2b,) se;;;ua karakter tergolong tinggi. Nilai duga heritabilitas arti sempit (h2,,) berkisar dari rendah sampai tinggi. Kuantifikasi dominan ditunjukkan oleh bobot per tanaman tinggi, jumlah buah banyak, buah panjang, diameter buah kecil dan umur panen genjah. Berdasarkan pendugaan DGK, heterosis dan nilai tengah dengan karakter bobot buah per tanaman sebagai pertimbangan utarna maka 12 hibrida berpotensi untuk d~jadikan varietas hibrida berdaya hasil tinggi.Kata kunci: C. annuum, setengah dialel, parameter genetik, daya gabung, heterosis
Yendahuluan
Cabai (Capsicum annuurn L.) mempunyai nilai ekonomis tinggi dan hmyak
diusahakan. Luas areal pertanaman cabai mpncapai 19.12% dari se!uruh areal
pertanaman sayuran (Ditlinhorti 2006). Akan tetapi, luas areal pertanaman cabai
tidak diikuti dengan tingginya produktivitas yang hanya mencapai 6.39 ton per
hektar (Deptan 2007) dari potensi hasilnya yang dapat mencapai 12 ton per hektar
( P w a t i et al. 2000).
Penyebab rendahnya produktivitas cabai di Indonesia berkaitan dengan
kualitas benih, teknik budidaya, serangan hama penyakit serta sedikitnya varietas
berdaya h a i l tinggi (Yunianti et al. 2006). Persilangan merupakau rara yang lazim diiakukan untuk memperbaiki produktivitas cabai. Persilangan cabai
hibrida. Melalui persilangan buatan, diharapkan diperoleh gabungan gen-gen baik
yang herasal dari tetua-tetua yang disilangkan (Darlina et al. 1992).
Upaya meningkatkan kemajuan genetik ketumnall suatu persilangan akan
lebih efektif apahila didasarkan kriteria genetik yang jelas pada saat pemilihan
tetua persilangan (Suhendi el rrl. 2004). Informasi kriteria genetik suatu karakter
dcpat diduga menggunakan analisis dialel. Tujuan analisis dizlel adalah untuk
mempelajari kendali genetik sifat kuantitatif yang merupakan ha1 penting unt-k
merencanakan dan menyelesaikan program pemuliaan (Viana et a/. 2001). Selain itu, analisis dialel merupakan metode yang cepat uutuk mengetahui informasi
genetik pada generasi F1 dan telah digunakan secara luas untuk analisis karakter
kuantitatif pada berbagai organisme (Ukai 1991). Penggunaan silang dialel juga
memungkinkan untuk mernilih tetua dan memberikan informasi tentang daya
gabung tetua dalam hibrida sehingga dapat membantu pemulia untuk
meningkatkan dan menyeleksi populasi segregan (de Sousa & Maluf 2003).
Analisis silang dialel akan menghasilkan individu yang banyak pada
keturunannya. Penanganan individu yang banyak membutuhkan tenaga, ketelitian
dan biaya tinggi. Modifikasi silang dialel menjadi silang setengah dialel
merupakan cara untuk mengatasi ha1 tersebut.
Evaluasi daya gabung umum (DGU) dan daya gabung khusus (DGK)
merupakan uji keturunan yang sering digunakan sebagai evaluasi awal dalarn
perakitan varietas hibrida. Daya gabung @GU dan DGK) merupakan uji
keturunan yang diperlukan dalam mengidetifikasi kombinasi tetua untuk
menghasilkan hibrida berdaya hasil tinggi atau sifat baik laimya yang diinginkan.
Pemilihan tetua diarahkan kepada tetua yang memiliki efek DGU tinggi atau
kombinasi tetua dengan efek DGK nyata untuk karakter yang dikehendaki
sehingga keturunannya akan berpenampilan baik (Wahyuni el a1 2004). Selain itu,
hasil tinggi dapat di capai jika kombinasi tetua tersebut memiliki heterosis positif
(Darlina et al. 1992).
Heterasis adalah peningkatan yang terlihat dari ~ersilangan dua galw murni.
Pemurfaatan heterosis pada varietas yang relatif homosigot diawali dengan seleksi
tetua yang menghasilkan kombinasi karakTer terbaik sehingga diperoleh
Penelitian ini bertujuan untuk menduga parameter genetik, nilai daya
gabung umum, daya gabung khusus dan heterosis karakter hortikultura cabai.
Bahan dan Metode
Tempat dan Waktu
Penelitian diiaksanakan di Laboratorium Pendidikan Pemuliaan Tanaman
II'3, rumah plastik dan Kebun Percobadn Tajur Uiliversi(y Earnt IPB, Bogor. Pembentukan populasi dilaksanakan pada bulan Desemher 2095 sampai Juli 2006
dan evaluasi karakter hortikultura di lahan dilakukan pada bulan Agustus 2006
sampai Januari 2007.
Bahan
Bahan tanaman yang digunakan adalah 6 genotipe cabai koleksi Bagian
Genetika dan Pemuliaan Tanaman, Departemen Agronomi dan Hortikultura
Fakultas Pertanian IPB. Bahan tanaman berasal dari genotipe lokal dan introduksi
yang telah digalurkan (Tabel 1).
Tabel 1 Genotipe cabai bahan penelitian
No. Genotipe Tetua asal Daerah Keterangan -
asal/sumber
1 IPBCI PSPT C-17 IPB, Indonesia Produksi tinggi 2 IPB C2 PSPT C-1 1 IPB, Indonesia Produksi t i G i
3 IPB C4 Cilibangi 2 Malaysia Tahan ChiVMV
4 IPBC9 ICPN 12#4 AVRDC Tahan PVY, layu bakteri
5 IPB C10 PBC 495 AVRDC Tahan CMV, ChiVMV
6 IPB C14 CCA321 AVRDC Tahan CMV, ChiVMV
Metode
Pembentukan Populasi
Pembentukan populasi dilakukan di rumah plastik Kebun Tajur University
Farm IPB. Bogor. Enam tetua ditanam dalam kantong plastik berdiamater 35 cm
berisi 8 kg media tanam. Media tanam yang digunakan adalah campuran tanah
dan pupuk kandang dengan perbandingan 1:l (VN) yang disterilisasi dengan
suhu tinggi selama 8 jam. Tanaman dipelihara mengikuti teknik budidaya baicu.
Populasi yang dibentuk adalah Fi tanpa resiprok menggunakan persilangan
Tabel 2 Persilangan setengah dialel dan - - selfin~t "
-
enam tetuaTetua Tetua jantan
Betina IPB C2 IPB C14 IPB C4 IPB C9 IPB C10 IPB C1
IPB C2 8 X X X X X
IPB C14
c3
X X XX
IPB C4 8 X X X
IPB C9 @ X X
IPB C10
c3
XIPB C1 8
@ = SeFng; X = Persilansan
Selfing tetua dilakukan dengan menutup individu tanaman dengan sungkup kasa (Gambar 2). Peny~mgkupan dilakukan sebelum tanaman berbunga untuk
[image:34.536.44.460.45.735.2]menghindari masuknya serbuk sari dari tanaman lain.
Gambar 2 Se1Jin.g tetua lnenggunakan sungkup kasa.
Persilangan dilakukan pagi hari pada pukul 06.00 - 10.00. Emaskulasi
dilakukan sebelum bunga betina mekar untuk menghindari penyerbukan sendiri.
Emaskulasi dilakukan dengan membuang mahkota dan benang sari bunga betina
menggunakan pinset yang disterilisasi menggunakan alkohol 70%. Penyerbukan
menggunakan serbuk sari bunga yang bani mekar. Serbuk sari ditempelkan pada
bunga betina yang telah diemaskulasi. Bunga betina yang telah diserbuki ditutup
menggunakan selotipe dan diberi label yang berisi informasi nama tetua yang
Gambar 3 Teknik persilangan buatan pada cabai: A. Bunga betina siap diserbuki; B. Kastrasi bunga betina; C. Emaskulasi bunga betina; D. Bunga betina telah dikastrasi dan diemaskulasi; E. Bunga jantan sumber serbuk sari; F. Pengambilan anter; G. Anter yang terkumpul; H. Pengambilan serbuk sari; I. Penyerbukan menggunakan pinset; J. Penutupan menggunakan selotipe; K. Pelabelan; L. Buah hasil persilangan (Sumber: Yunianti (2007) yang dimodifikasi).
Evaluasi Karakter Hortikultura
Evaluasi karakter hortikultura dilakukan di Kebun Percobaan Tajur
Universify Farm IPB, Bogor. Evaluasi karakter hortikultura menggunakan Rancangan Acak Kelompok Lengkap (RAK), genotipe sebagai perlakuan dengan
tiga ulangan. Genotipe cabai yang digunakan berjumlah 21 (6 tetua dan 15 hibrida hasil persilangan).
Benih disemai pada media tanam dalam nampan. Bibit dipindahkan ke lahan
setelah berumur 5 minggu setelah semai. Bibit ditanam pada lahan yang ditutup
plastik hitam perak dengan jarak tanam 50 x 50 cm. Pemeliharaan ineliputi
penyiraman, pemupukan dan pengendalian hama penyakit. Pemupukan dilakukan
Tabel 3 Jenis d m dosis auauk serta waktu ~emunukan
. .
tanaman cabaiPupuk Waktu Pemupukan Total
Pupuk dasar 3 MST 6 MST' 9 MST
P u ~ u k Kandanrr
-
30 tonha . - - 30 ton/haTSP 200 kg/ha - - - 200 kg/ha
Urea - 50 kg/ha 50 kg/ha 50 kgha 150 kgha
Z A - 150 kg/ha 150 kgha 150 kgha 450 kgha
KCL
-
70 kg/ha 70 kg/ha 60 kg/ha 200 kglhaMST = rninggu setelah tanarn
Peubah yang diamati meliputi: bobot buah per tanarnan (g), jumlah buah,
panjang buah (cm), diameter buah (mm) dan umur panen (HST). Analisis data
dimulai dengan melakukan analisis sidik ragam. Analisis silang setengah dialel
dilakukan menggunakan dua pendekatan yaitu Hayman (1954) dan Grifing
(1956). Selain itu. dilakukan pendugaan nilai heterosis berdasarkan nilai tengah
kedua tetua dan i i a i tengah tetua terbaik menggunakan pendekatan Fehr (1987).
Metode analisis silang set~,igah dialel dan pendugaan heterosis disajikan secara
lengkap pada Lampiran 1.
Hasil dan Pembahasaa
Kuadrat tengah sidik ragam karakter hortikultura disajikan lengka~ pada
Tabel 4. Semua karakter menunjukkan perbedaan nilai kuadrat tengah sangat
nyata diantara genotipe. Hasil sidik ragam yang nyata mempakan syarat untuk
melakukan pendugaan parameter genetik menggunakan pendekatan Hayman dan
pendugaan daya gabung menggunakan pendekatan Grifing. Oleh karena itu
analisis dapat dilanjutkan.
Tabel 4 Kuadrat tenpah karakter hortikultura yang diamati aada
-
.
-
21-
~zenotiae cabaibush Jumlah Panjang Diameter Umur
SK db per tanaman buali buah panen
buah
(z)
(cm) (mm) (HST)Blok 1 35207.75 8629.78 0.06 0.09 0.44
Genotipe 20 151950.09** 54972.26** 20.84** 6.96** 116.89**
Galat 40 5146.79 2393.19 0.37 0.32 8.63
Pendugaan Parameter Genetik
Interaksi Gen
Nilai b (Wr, Vr) menunjukkan ada tidaknya interaksi gen (Tabel 5).
Berdasarkan uji t, jika nilai b berbeda nyata dengan 1 maka menunjukkan ada
interaksi gen dan jika nilai b tidak berbeda nyata dengm 1 maka tidal< ada
interaksi gen (Hzyman 1954; Singh & Chaundhary 1079: de Sousa & Maluf
2003) IIasil yang diperoleh pada pengujian nilai b adalah karakter bobot buah
per tanaman, panjang buah, diameter buah dan umur panen tidak berbeda nyata
dengan 1 menandakan tidak ada peran interaksi gen pada keempat kamkter
tersebut. Karakter jumlah buah menunjukan nilai h berbeda nyata dengan 1
menandakan ada peran interaksi gen pada karakter jumiah buah. Hal tersebut
menunjukkan bahwa asumsi pertanla ( t i d ~ k ada interaksi gen) terpenuhi pada
semua karakter hortikultura yang diamati kecuali jumlah buah.
Pengaruh Aditif (D), Dominan (HI) dan Tingkat Dominansi
Pengaruh aditif @) tidak berbeda nyata pada karakter bobot buah per
tanaman dan jumlah buah, tetapi berbeda nyata pada kder panjang buah,
diameter buah dan umur panen. Pengarub dominansi (HI) 'wrbeda nyata pada
semua karakter. Hal tersehut menunjukkan hahwa karakter bobot buah per
tanaman dipengaruhi oleh aksi gen dominan, sedangkan karakter panjang buah,
diameter buah serta umur panen dipengaruhi oleh aksi gen aditif dan dominan.
P e n g a d dominansi terlihat dari nilai rata-rata tingkat dominansi
[(HIID)'")]. Nilai (HIID)'" karakter bobot huah per tanaman. jumlah buah dan
umur panen lebih besar dari 1 sehingga dapat disimpulkan tingkat dominansi
karakter tersehut adalah overdominance. Nilai ( H I / D ) ' ~ karakter panjang buah dan diameter buah kurang dari 1 sehingga dapat disimpulkan tingkat dominansi
karakler tersebut adalah dominan parsial. Nilai
HI ID)'^
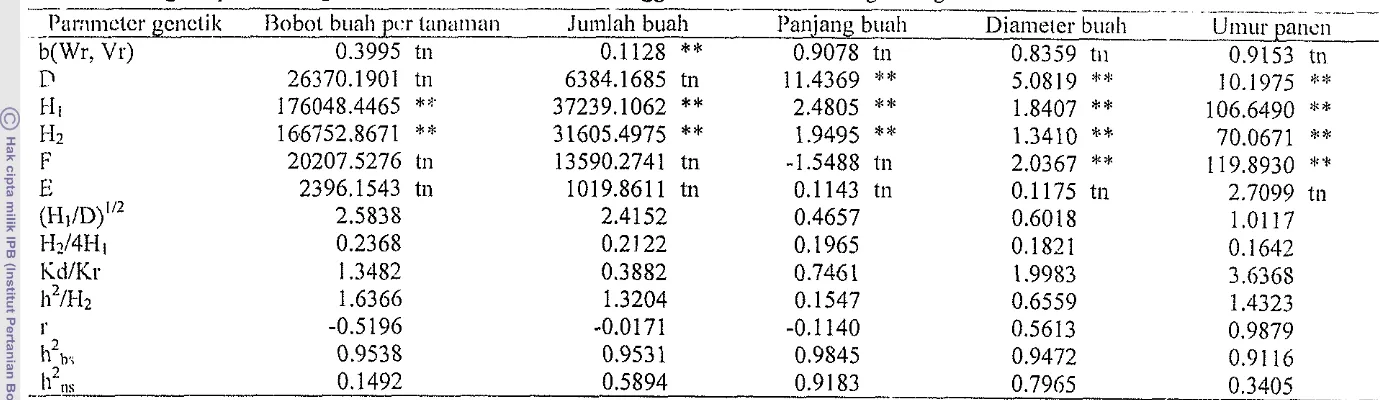
lebih dari 1 rnenunjukkanTabel5 Pcndugaan parameter genetik karakter hotikultura menggunakan analisis sila~ig setengah dialel
-
- I'nr:~inctcr gcnctik 13obot buall 1x.r lanoliian
b(Wr. Vr) 0.3995 tn
~ -
-- - .lumlah buah
0.1128
**
. .
I'ati,jang buah 0.9078 tn
-
...
Diameter b~leli 0.8359 ti1
2 -.. ..-
11 ,,s
-- 0.1492 0.5894 0.91 83 0.7965 0.3405
Distribusi Gen (HI) dalam Tetua
Nilai Hz digunakan untuk mengetahui distribusi &en dalain tetua. Hasil
pengujian menunjukkan bahwa nilai Hz semua karakter hortikultura yang diamati
berbeda nyata. Hal tersebut lnenandakan bahwa gen-gen pang menentukan
pewarisati karakter bobot buah per tanaman, jumlah buah, panjang buah, diameter
buah dan cmtlr panen tidak menyebar merata di dalam tetua.
Proporsi Gep Dominan terhrdap Gen Resesif
Porporsi gen dominan terhadap gen resesif tertihat dari nilai Kd/Kr. Jika
KdIKr > 1 maka gen-gen dominan lebih banyak di dalam tetua. Sebaliknya, jika
KdIKr < 1 maka gen-gzn resesif lebih banyak di clalam tetua. Nilai Kd/Kr >1
diperoleh pada karakter bobot buah per tanaman, diameter buah dan umur panen
menunjukkan gen-gen dominan lebih banyak di dalam tetua. Nilai Kd/Kr < 1
diperoleh pada karakter jumlah buah dan panjang buah menunjukkan gen-gen
resesif lebih banyak di dalam tetua.
Jumlah Kelompok Gen Pengendali
Jumlah kelompok gen pengendali ditunjukkan oleh nilai h2/H2. Jumlah
kelompok gen pengendali bobot buah per tanaman, jumlah buah dan umur panen
minimal 2 kelompok gen. Scdangkan jumlah kelompok gen pengendali panjang
buah dan diameter buah minimal 1 kelompok gen.
Heritabilitas
Nilai duga heritabilitas arti luas (h2h,) karakter bobot huah per tanamaq,
jumlah buah, panjang buah, diameter buah dan umur panen p d a percobaan ini
tergolong tinggi. Hal tersebut menunjukkan ragam gejala yang muncul terutama
dikendalikan oleh faktor genetik. Nilai heritabilitas arti sempit menggambarkan
peranan gen aditif dalam pengendalian . suatu karakter (Tenaya ef a!. 2003:
Agustina 7004). Nilai duga heritabilitas arti sempit (h2,,) karakter bobot buah per
tanaman tergolong rendah dan karakter umur panen tergolong sedang
menandakan pengamh ragam aditif yang rendah terhadap karakter tersebut. Pada
karakter jurnlah buah. panjang buah dan diameter buah memiliki nilai duga arti
sempit (h2,) tergolong tinggl menandakan pengamh ragam aditif yang tinggi
Arah d a n Urutan Dominansi
Nilai kuantifikasi karakter yang rendah aka11 domillan terhadap yang tinggi
apabila nilai r(Wr+Vr. Yr) positif. sebaliknya nilai kuantifikasi yang tinggi akan
dominan terhadap yang rcndah apabila nilai r(WrtVr, Yr) negatif (Agustina
2004). Berdasarkan nilai r(Wr-Vr. Yr) diketahui bahwa kuantifikasi dominan
ditunjukkan oleh bnbot ner tanamzn tinggi. jumlah buah banyak, buah panjang,
diameter buah kecil dati umur panen genjah (Tabel 5).
Urulan do~llinansi tetua dapat tercennin berdasarkan (WrtVr) atau posisi
tetua pada grafik hubungan antara Wr dan Vr. Jika pada grafik hubungan antara
Wr dan Vr posisi titik tetua makin dekat pada titik no1 maka tetua tersebut paling
banyak mengandung gen dominan, sebaliknya rnakin jauh dari titik no1 maka tetua
[image:40.532.36.453.0.762.2]tersebut paling banpak mengandung gen resesif. Nilai (Wr +Vr) disajikan pada
Tabel 6.
Tabel 6 Nilai (Wr+Vr) karakter hortikultura 6 tetua cabai
Bobot buah Jumlah Panjang Diameter Umur
Tetua
per tanaman buah buah buah panen
IPB C2 48632.53 14245.1 8 8.43 1.55 2.44
IPB C14 44855.27 14369.41 13.70 3.28 19.07
IPB C4 100639.56 24356.83 10.56 3.45 12.722
IPB C9 10705.71 26245.47 9.?5 2.73 25.93
IPB C10 75669.30 16071.17 9.13 2.97 234.68
IPB C1 52396.27 34659.56 9.07 6.06 -13.52
Bobot huah per tanaman. Dominansi tetua berdasarkan (WrtVr) untuk
karakter bobot bual? per tanaman secara berurutan adalah IPB C4, IPB C10.
IPB C1, IPR C2, IPB C14 dan IPR C9 (Tabel 6). Tetua yang paling banyak
mengandung gen resesif, karena paling jauh dari titik not adalah IPB C4. Tetua
yang paling banpak mengandung gen dominan, karena paling dekat dengan titik
no1 adalah IPB C9 (Gambar 4A). Garis lurus Wr memotong sumbu tegak (Wr) di
bawah pangkal sumbu menandakan terdapat overdominance pada karakter bobot buah per tanaman. Jika garis lurus menlotong sumbu tegak di bawah pangkal
Jumlah buah. Berdasarkan (WrWr) urutan dominansi tetua untuk karakter
jumlah buah adalah IPB C1, IPB C9, IPB C4, IPB C10. IPB C14 dan IPB C2.
Tetua IPB C1 merupakan tetua yang paling banyak mengandung gen resesif.
karena paling jauh dari titik nol. IPB C2 paling banyak mengandung gen
dominan, karena paling dekat dengan titik no1 (Gambar 4B). Interaksi gen yang
nyata pada jumlah buah tidak dapat diketahui berdcsarkan regresi (Wr+Vr).
Kegresi F; gaga1 mendeteksi interaksi non alelik dan uji regeresi tidak peka
terhadap interaksi gen (Dudley 1999).
Panjang buah. Secara berurutan dominansi tetua untuk karakter panjang
buah berdasarkan nilai (Wr+Vr) adalah IPB C14, IPB C4: IPB C9, IPB C10,
IPB C1 dan IPB C2. Tetua yang paling banyak mengandung gen resesif, karena
paling jauh dari titik no1 adalah IPB C14. Tetua yang paling banyak mengandung
gen dominan, karena paling dekat dengan titik no1 adalah IPB C2 (Gambar 4C).
Garis lurus Wr memotong sumbu tegak (Wr) di atas pangkal sumbu menandakan
gen dominan tidak lengkap mengendalikan karakter bobot buah per tanaman. Jika
garis lurus memotong sumbu tegak di atas pangkal sumbu menandakan aksi gen
dominan tidak lengkap (de Sousa & Maluf 2003).
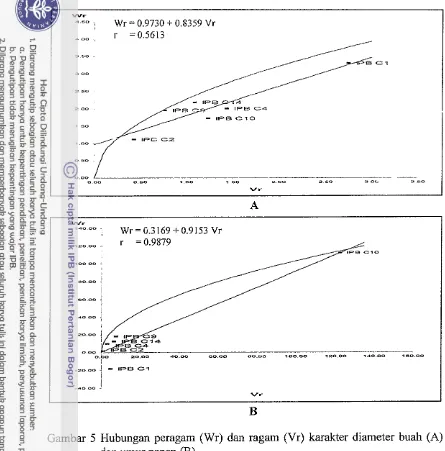
Diameter buah. Dominansi tetua berdasarkan nilai (Wr+Vr) karakter
diameter buah adalah IPB C2, IPB C9, IPB C10, IPB C14, IPB C4 dan IPB C1.
Tetua IPB C1 merupakan tetua yang paling banyak mengandung gen resesif,
karena paling jauh dari titik nol. IPB C2 paling banyak mengandung gen dominan,
karena paling dekat dengan titik no1 (Gambar 5A). Garis lurus Wr memotong
sumbu tegak (Wr) di atas pangkal sumbu menandakan gen dominan tidak lengkap
mengendalikan karakter bobot buah per tanaman.
Umur panen. Urutan dominansi tetua berdasarkan nilai (Wri-Vr) karakter
umur panen adalah IPB C2, IPB C4, IPB C1, IPB C14, IPB C9 dan IPB C10.
Tetua IPB C10 merupakan tetua yang paling banyak mengandung gen resesif,
karena paling jauh dari titik nol. IPB C2 paling banyak mengandung gen
uominan, karena paling dekat dengan titik no1 (Gambar 5B). Garis lurus Wr
memotong sumbu tegak (Wr) pada pangkal sumbu menandakan gen dominan
Gambar 5 Hubungan peragam (Wr) dan ragam (Vr) karakter diameter buah (A)
d m umur panen (B).
Daya Gabung
Efek daya gabung umum (DGU) dan daya gabung khusus (DGK) semua
karakter hotikultura berbeda sangat nyata (Tabel 7). Hal tersebut menunjukkan
bahwa aksi gen aditif d m aksi gen non aditif mempengaruhi ekspresi karakter
bobot buah per tanaman. jumlah buah, panjang buah, diameter buah dan umur
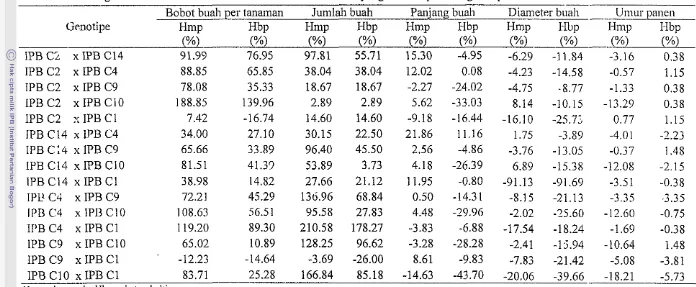
Tabel 7 Kuadrat tengah daya gabung karakter hortikultura yang diamati pada 21 genotipe cabai
SK db Bobot buah Jumlah Panjang Diameter Umur
per tanaman buah buah buah panen
(9) (cm) (mm)
DGU 2 34684.08** 40256.45** 25.88** 7.76** 71.74**
DGK 20 55972.01** 11013.30** 0.64** 0.51** 28.04**
Galat 40 174.93 797.73 0.12 0.11 2.88
SK = sun