SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
STUDI POLIEMBRIONI, VIABILITAS BENIH DAN
IDENTIFIKASI GENETIK SEMAIAN JERUK JAPANSCHE
CITROEN (
Citrus limonia
Osbeck.) MENGGUNAKAN SSR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Studi Poliembrioni, Viabilitas Benih dan Identifikasi Genetik Semaian Jeruk Japansche Citroen (Citrus limonia Osbeck.) menggunakan SSR adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
ANIS ANDRINI. Studi Poliembrioni, Viabilitas Benih dan Identifikasi Genetik Semaian Jeruk Japansche Citroen (Citrus limonia Osbeck.) menggunakan SSR. Dibimbing oleh TATIEK KARTIKA SUHARSI dan MEMEN SURAHMAN.
Jeruk Japansche Citroen (Citrus limonia Osbeck) atau JC merupakan batang bawah jeruk yang banyak digunakan petani Indonesia karena daya adaptasinya luas dan mempunyai ketahanan terhadap beberapa penyakit. Benih jeruk JC harus bermutu baik fisik, fisiologis, genetik dan kesehatan. Mutu benih maksimal saat mencapai masak fisiologis. Pendugaan tingkat kemasakan benih dapat dilakukan berdasarkan tingkat kemasakan buah jeruk JC yang umumnya terlihat dari perubahan warna kulit buah .
Benih JC bersifat poliembrioni sehingga dalam satu benih dapat menghasilkan lebih dari satu semaian asal embrio zigotik dan atau embrio nuselar. Adanya embrio nuselar dapat meningkatkan mutu genetik benih karena menghasilkan semaian yang sama dengan induknya. Namun demikian dalam persemaian JC masih ditemui semaian off type asal embrio zigotik yang pada umumnya dikenali dari morfologi semaian. Identifikasi genetik semaian berdasarkan penanda morfologi perlu dibuktikan lebih lanjut dengan penanda molekuler. Salah satu penanda molekuler yang efektif yaitu SSR karena dapat membedakan genotipe homozigot dan genotipe heterozigot.
Tujuan penelitian ini mendapatkan informasi poliembrioni pada jeruk JC, Mendeteksi tingkat kemasakan fisiologis benih jeruk JC berdasarkan warna kulit dan kekerasan buah jeruk, mengetahui pengaruh tingkat kemasakan fisiologis benih terhadap daya tumbuh benih, multiple seedling, semaian off type dan semaian true to type, mengidentifikasi semaian true to type dan off type berdasarkan penanda morfologi dan penanda SSR.
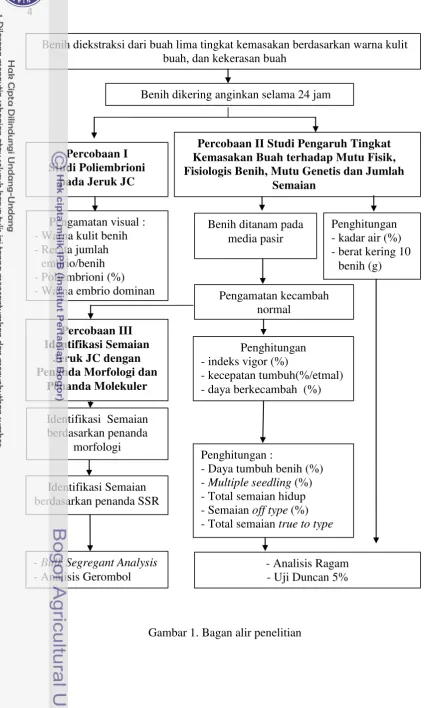
Percobaan dilakukan di Balai Penelitian Tanaman Jeruk dan Buah Subtropika pada bulan April 2012 sampai dengan Januari 2013. Penelitian terdiri atas 3 percobaan yaitu 1) Studi poliembrioni pada jeruk JC 2) Studi pengaruh tingkat kemasakan buah terhadap mutu fisik, fisiologis benih, mutu genetis dan jumlah semaian, serta 3) Identifikasi semaian jeruk dengan penanda morfologi dan penanda SSR.
Percobaan I dilakukan berdasarkan pengamatan visual pada masing-masing 30 benih dari lima tingkat kemasakan buah berdasarkan warna kulit buah dan kekerasan buah. Tingkat kemasakan buah dilihat berdasarkan warna kulit buah dan kekerasan buah yaitu 1) buah warna hijau tua, keras (> 7 kg/cm2) 2) buah warna hijau semburat kuning, agak keras (6-7 kg/cm2), 3) buah warna hijau kekuningan, agak lunak (5-5.9 kg/cm2), 4) buah warna kuning > 90% merata, lunak (4-4.9 kg/cm2) dan 5) buah warna kuning-orange, sangat lunak (< 4 kg/cm2). Tolak ukur yang diamati yaitu warna kulit benih, rerata jumlah embrio per benih, persentase benih poliembrioni (%) dan warna embrio dominan.
benih. Tolak ukur yang diamati yaitu kadar air benih (%), berat kering 10 benih (g), indeks vigor (%), kecepatan tumbuh benih (%/etmal), daya berkecambah (%), daya tumbuh benih (%), persentase multiple seedling, total semaian hidup, persentase semaian off type dan total semaian true to type. Data dianalisis dengan analisis ragam (ANOVA). Apabila diperoleh hasil analisis ragam yang berpengaruh nyata, maka dilakukan uji lanjut dengan Duncan Multiple Range Test (DMRT) pada taraf 5%.
Percobaan III yaitu identifikasi semaian true to type dan off type dengan penanda morfologi dan molekuler. Percobaan ini menggunakan 12 Sampel dari semaian pada percobaan II dan pohon induk JC sebagai pembanding. Identifikasi penanda morfologi berdasarkan descriptor list for Citrus dari IPGRI, dengan beberapa modifikasi. Tolak ukur yang diamati yaitu tinggi semaian, jumlah daun, rasio panjang dan lebar daun, LAMI (rasio panjang dan lebar pucuk daun), keberadaan duri, keberadaan stipula dan bentuk daun. Identifikasi penanda molekuler dengan 6 primer SSR. Secara garis besar, analisis DNA dengan penanda SSR terdiri atas ekstraksi DNA, uji kualitas dan kuantitas DNA dengan elektroforesis, amplifikasi dan separasi DNA dengan PCR, visualisasi hasil PCR dan analisis data molekuler. Bulk Segregant Analysis (BSA) digunakan untuk melihat keterpautan antara marka morfologi dan marka SSR. Analisis gerombol berdasarkan penanda morfologi dan SSR dilakukan dengan prosedur SIMQUAL dengan metode SAHN.
Hasil percobaan menunjukkan bahwa persentase poliembrioni benih jeruk JC pada lima tingkat kemasakan benih berdasarkan kemasakan buah yaitu 53.34-73.32% dengan jumlah embrio 1-6 per benih. Semakin masak benih warna embrio dominan semakin berwarna krem dan tidak terlihat adanya warna kehijauan.
Tingkat kemasakan fisiologis benih jeruk JC dicapai pada saat warna kulit buah kuning lebih dari 90%, lunak, warna kulit benih krem kecoklatan dan warna embrio dominan krem. Tingkat kemasakan benih berdasarkan warna kulit buah tidak berpengaruh nyata terhadap persentase multiple seedling dan off type. Multiple seedling pada benih jeruk JC dapat meningkatkan total semaian true to type.
Marka AG14 pada ukuran basa 170bp terpaut dengan marka morfologi LAMI < 0.75 dengan kemungkinan bias 40%. Karakter semaian jeruk JC true to type antara lain tipe daun tunggal, pupus daun hijau muda dan bentuk daun elliptic dengan pangkal daun cuncate, ujung daun obtusus dan tepi daun crenate. Sementara itu, tipe daun trifoliata, warna pupus daun merah keunguan dan bentuk daun lanceolate dengan ujung rounded dapat menjadi penanda yang efektif untuk semaian jeruk JC off type.
SUMMARY
ANIS ANDRINI. Study of Polyembryony, Seed Viability and Genetic Identification of Japansche Citroen (Citrus limonia Osbeck) seedling using SSR. Supervisor by TATIEK KARTIKA SUHARSI and MEMEN SURAHMAN.
Japansche Citroen (Citrus limonia Osbeck) or JC is citrus rootstock that is often used by Indonesian citrus farmer because its wide adaptation ability and has resistant to several diseases. JC seeds should have physical, physiological, genetic and health quality. Seed has maximum quality when it is reached at the time of physiological maturity. Seed maturity, in general is decided by physical appearance of fruit, such as rind colour changes.
JC seeds have polyembryony characteristic; threrefore one or more embryos per seed consist of either zygotic embryo and or nucellar embryo. Nucellar embryo can be useful for clonal propagation. Nevertheles, there were off type seedling. It is necessary to identify seedling base on morphology and molecular marker such as SSR marker. Morphology marker was easy to used and cheap, but the trully genetic must be identify with SSR. SSR could differenciate between homozygot and heterozigot genotype.
The aims of this research were to acquire information about poliembryony on JC citrus, to determine physiological maturity of JC seed base on rind colour and fruit hardness, to know the effect of fruits maturity to physical and physiological quality of seed, genetic quality and number of seedling, and to identify JC seedling base on morphology marker and SSR marker.
The research was conducted in Indonesian Citrus and Subtropical Fruits Research Institute from April 2012 to January 2013. The research consisted of three experiments, there were 1) study on polyembryony of JC , 2) study on effect of fruits maturity level to physical and physiological quality of seed, genetic quality and number of seedling, and 3) identification of JC seedling base on morphology marker and SSR marker.
Experiment I was conducted based on visual observations on each of 30 seeds of five maturity levels. Criteria of fruit maturity are 1) dark green rind, hard fruit (> 7 kg/cm2), 2) green yellow rind, rather hard (6-7 kg/cm2), 3) pieces of yellowish green rind, rather soft (5-5.9 kg/cm2), 4) >90% yellow rind, soft (4-4.9 kg/cm2) and 5) pieces of yellow-orange rind, very soft (< 4 kg/cm2). The experiment variable were seed coat colour, the mean number of embryos per seed, polyembryony seed percentage (%) and the dominant colour of the embryo.
of variance were significant different, then tested by Duncan's Multiple Range Test (DMRT) at 5% level.
Experiment III was the identification of true to type seedlings and off type seedling with morphology marker and molecular markers. This experiment used 12 samples of seedlings in experiments II and JC parent tree as a comparison. Identification of JC seedling based on descriptor list for Citrus from IPGRI with modified. Variable observed were high, the number of leaves, leaf length and width ratio, LAMI (the ratio of leaf length and width), the presence of spines, the presence of stipules and leaf shape. Identification of molecular markers with 6 SSR primers. The outline of DNA analysis with SSR markers were DNA extraction, test the quality and quantity of DNA by electrophoresis, amplification and separation of DNA by PCR, visualization of PCR results and analysis of molecular data. Bulk Segregant Analysis (BSA) was used to know the linkage between SSR markers and morphology markers. Cluster analysis of morphology marker and SSR marker used SIMQUAL procedure and SAHN method.
The result showed that JC citrus has 1-6 embyo per seed. The persentage of JC seed polyembryony was 53.34 to 73.32%. The riper the seed, the more cream- coloured the dominant embryo and no green colour.
JC seed physiological maturity was achieved in fruits with more than 90% rind is yellow, low hardness fruits, the color of seed skin was brownish cream and embryos color was cream. Seed maturity was not significantly different affect to the percentage of multiple seedling and percentage of off type seedling. Multiple seedling can increase the number of total of true to type seedling.
Seedling leaves type, leaves shape and colour of young leaves, in general use as morphology marker of true to type and off type seedling. SSR primer AG14 which amplified in 170bp have linkage to LAMI < 0.75 but there were 40% bias. Characteristic of JC true to type seedling were single leaf type, light green shoot tip colour, elliptic leaf shape with cuncate leaf base, obtusus leaf tip and crenate leaf margins. Meanwhile, trifoliata leaf type, purplish red shoot tip colour and lanceolate lamina shape with rounded tip were an effective marker to identify JC off type seedling.
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu dan Teknologi Benih
STUDI POLIEMBRIONI, VIABILITAS BENIH DAN
IDENTIFIKASI GENETIK SEMAIAN JERUK JAPANSCHE
CITROEN
(
Citrus limonia
Osbeck
.
) MENGGUNAKAN SSR
ANIS ANDRINI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Studi Poliembrioni, Viabilitas Benih dan Identifikasi Genetik Semaian Jeruk Japansche Citroen (Citrus limonia Osbeck.) menggunakan SSR
Nama : Anis Andrini NIM : A251100071
Disetujui oleh Komisi Pembimbing
Dr Dra Tatiek Kartika Suharsi, MS Ketua
Prof Dr Ir Memen Surahman, MScAgr Anggota
Diketahui oleh
Ketua Program Studi Ilmu dan Teknologi Benih
Prof Dr Ir Satriyas Ilyas, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Alhamdulillah. Puji syukur penulis panjatkan kehadirat Allah SWT atas berkah dan karunianya sehingga penulis dapat melalui semua hambatan dan rintangan dan pada akhirnya dapat menyelesaikan tesis ini.
Ucapan terima kasih penulis sampaikan kepada Dr DraTatiek Kartika Suharsi, MS dan Prof Dr Ir Memen Surahman, MScAgr sebagai komisi pembibing yang telah berkenan memberikan bimbingan dan saran-saran selama penulis melakukan penelitian dan penulisan tesis. Kepada Kepala Badan Litbang Pertanian, Kepala Puslitbang Hortikultura dan Kepala Balai Penelitian Tanaman Jeruk dan Buah Subtropika penulis mengucapkan terima kasih karena telah memberikan kesempatan kepada penulis untuk melanjutkan studi. Selain itu kepada Kepala Kebun Tlekung dan Kepala Kebun Punten yang telah memberikan fasilitas dan saran-sarannya sehingga penulis dapat melakukan penelitian ini. Terima kasih pula kepada Chaireni Martasari, Dita Agisimanto, Farida, Arisah, Baiq Dina, Ibu Emi Budiyati, Lizia, teknisi dan tenaga harian Balitjestro yang banyak memberikan bantuan. Kepada Cici Tresniawati, Ikrarwati, Ita Yustina, Noflindawati, Ratri, Reren, Candra, Via dan teman-teman ITB 2010 serta semua pihak yang tidak dapat penulis sebutkan satu persatu atas saran dan bantuannya penulis ucapkan terima kasih.
Terima kasih untuk almarhum ayahanda tercinta yang selalu menggerakkan hati penulis melanjutkan studi. Kepada bunda, suami dan anak tercinta, penulis sangat mengagumi, mencintai dan berterimakasih atas pengorbanan, cinta kasih kalian sehingga penulis dapat menyelesaikan studi ini. Penulis menyadari bahwa tulisan ini masih jauh dari sempurna. Meskipun demikian, penulis berharap semoga tesis ini dapat bermanfaat dan penelitian masih dapat dilanjutkan sehingga kemanfaatannya lebih luas.
DAFTAR ISI
DAFTAR TABEL xi
DAFTAR GAMBAR xi
DAFTAR LAMPIRAN xii
1 PENDAHULUAN 1
Latar Belakang 3
Tujuan Penelitian 3
Hipotesis 3
2 TINJAUAN PUSTAKA 5
3 METODE PENELITIAN 9
Waktu dan Tempat Penelitian 9
Bahan dan Alat 9
Metodologi 9
5 HASIL DAN PEMBAHASAN 18
Percobaan I Studi poliembrioni Jeruk JC 18
Percobaan II Studi Pengaruh Tingkat Kemasakan Buah terhadap Mutu Fisik, Fisiologis Benih, Mutu Genetis dan Jumlah
Semaian 22
Percobaan III Identifikasi Semaian Jeruk dengan Penanda Morfologi
dan Penanda Molekular 27
6 SIMPULAN DAN SARAN 35
Simpulan 35
Saran 35
DAFTAR PUSTAKA 35
DAFTAR TABEL
1 Urutan basa enam primer SSR (forward dan reverse) 17 2 Karakteristik benih dan poliembrioni jeruk JC pada lima tingkat
kemasakan buah 19
3 Rekapitulasi hasil analisis ragam kadar air, berat kering 10 benih, indeks
vigor, kecepatan tumbuh dan daya berkecambah 22 4 Pengaruh tingkat kemasakan buah terhadap mutu fisik dan fisiologis
benih JC 23
5 Rekapitulasi hasil analisis ragam daya tumbuh benih, persentase multiple seedling, total semaian hidup, persentase semaian off type, dan
total semaian true to type 25
6 Pengaruh tingkat kemasakan benih terhadap daya tumbuh benih, persentase multiple seedling, total semaian hidup, persentase semaian off
type, dan total semaian true to type 26
7 Identifikasi semaian jeruk JC berdasarkan karakter kuantitatif 28 8 Identifikasi semaian jeruk JC berdasarkan karakter kualitatif 29 9 Hasil Bulk Segregant Analysis penanda SSR pada penanda morfologi
LAMI < 0.75, vigor tinggi, keberadaan duri, bentuk daun elliptic, bentuk daun lanceolate, pangkal daun acute, ujung daun obtusus dan tepi daun
dentatus 32
6. Embrio-embrio pada benih rudimenter yang ditemukan pada buah tingkat kemasakan V terdiri atas embrio fase globular (G) dan fase
hearth (H) 22
7. Semaian jeruk JC true to type (no semaian 1, 2 dan 3) 28 8. Keragaan tipe daun trifoliata pada semaian no 4 saat tiga bulan setelah
tanam (A) dan enam bulan setelah tanam (B) 30
9. Keragaan warna pupus daun merah keunguan pada semaian no 6 saat berumur satu minggu setelah berkecambah (A), tiga bulan setelah
tanam (B) dan enam bulan setelah tanam (C) 30
DAFTAR LAMPIRAN
1. Keragaan semaian tunggal dan multiple seedling jeruk JC 40 2. Sidik ragam perlakuan tingkat kemasakan buah terhadap kadar air benih 40 3. Sidik ragam perlakuan tingkat kemasakan buah terhadap berat kering
10 benih 40
4. Sidik ragam perlakuan tingkat kemasakan buah terhadap indeks vigor
benih 40
5. Sidik ragam perlakuan tingkat kemasakan buah terhadap indeks vigor
benih setelah transformasi 41
6. Sidik ragam perlakuan tingkat kemasakan buah terhadap kecepatan
tumbuh benih 41
7. Sidik ragam perlakuan tingkat kemasakan buah terhadap daya
berkecambah 41
8. Sidik ragam perlakuan tingkat kemasakan buah terhadap daya tumbuh
benih 41
9. Sidik ragam perlakuan tingkat kemasakan buah terhadap persentase
multiple seedling 41
10. Sidik ragam perlakuan tingkat kemasakan buah terhadap persentase
multiple seedling hasil transformasi 42
11. Sidik ragam perlakuan tingkat kemasakan buah terhadap total semaian
hidup 42
12. Sidik ragam perlakuan tingkat kemasakan buah terhadap persentase
semaian off type 42
13. Sidik ragam perlakuan tingkat kemasakan buah terhadap persentase
semaian off type hasil transformasi 42
14. Sidik ragam perlakuan tingkat kemasakan buah terhadap total semaian
true to type 42
15. Visualisasi pita DNA dengan primer AG14 43 16. Visualisasi pita DNA dengan primer ATC09 43
17. Visualisasi pita DNA dengan primer TAA15 43
18 Visualisasi pita DNA dengan primer TAA27 43
19 Deskripsi varietas batang bawah jeruk Japansche Citroen (JC) 44
20 Riwayat Hidup 45
1
PENDAHULUAN
Latar Belakang
Benih jeruk diperbanyak secara vegetatif dengan okulasi maupun penyambungan sehingga benih jeruk terdiri atas dua bagian penting yaitu batang atas dan batang bawah. Batang bawah jeruk dapat mempengaruhi lebih dari 20 karakteristik yang berhubungan dengan karakter hortikultura dan hama/penyakit pada tanaman dan buah jeruk batang atas (Castle et al. 1993). Hal tersebut menunjukkan bahwa penggunaan benih batang bawah yang unggul dan bermutu tinggi adalah penting dalam budidaya jeruk dikarenakan adanya pengaruh yang besar dari batang bawah terhadap keberhasilan budidaya jeruk di berbagai lokasi dan cara budidayanya.
Japansche Citroen (JC) (Citrus limonia Osbeck) merupakan salah satu batang bawah jeruk varietas unggul yang telah banyak digunakan di Indonesia. Keunggulan batang bawah jeruk JC yaitu mempunyai daya adaptasi yang luas, kompatibel dengan berbagai varietas jeruk batang atas, meningkatkan vigor batang atas, dan dapat bertahan dengan baik pada kondisi lahan rawa daerah pasang surut (Putri 2002, Supriyanto dan Setiono 2005 dan Dwiastuti et al. 2007). Jeruk JC pada umumnya diperbanyak secara generatif menggunakan benih. Namun demikian, sampai saat ini benih jeruk JC diperjualbelikan secara bebas tanpa melalui sertifikasi benih sehingga tidak terdapat jaminan mutu benih. Menurut Ilyas (2012), mutu benih yang harus dipenuhi mencakup empat hal yaitu mutu fisik, fisiologis, genetis dan patologis.
Mutu fisik dan fisiologis benih dipengaruhi oleh tingkat kemasakan benih. Mutu benih mencapai maksimum pada saat masak fisiologis yang dicirikan oleh berat kering benih maksimum karena cadangan makanan benih sudah terbentuk sempurna, dan vigor benih maksimum (Ilyas 2012).
Bakal benih berkembang di dalam bakal buah dan pembentukan benih pada sebagian besar spesies dimulai sejak fertilisasi (Copeland dan McDonald 2001). Menurut Sedgley and Griffin (1989), pada saat buah masak fisiologis biasanya kulit buah yang berwarna hijau menjadi mengkilap dan secara perlahan-lahan klorofil akan hancur sehingga berubah menjadi warna kuning, merah atau jingga. Bonner (1972) mengemukakan adanya korelasi yang kuat antara perubahan warna yang terjadi pada buah yang masak dengan fase kemasakan benih. Dengan demikian tingkat kemasakan fisiologis benih dapat diduga berdasarkan mutu fisiologis buah.
Selain mutu fisiologis benih, sifat benih JC yang dapat mempengaruhi mutu benih yaitu adanya sifat poliembrioni. Menurut Kepiro dan Roose (2007) poliembrioni pada jeruk umumnya yaitu terdapat satu embrio zigotik dan atau satu atau lebih embrio nuselar di dalam satu benih.
2
Benih poliembrioni pada jeruk JC disebabkan adanya embrio nuselar yang berasal dari jaringan nuselus (jaringan cadangan makanan di luar kantong embrio). Adanya embrio nuselar merupakan keuntungan dalam perbanyakan tanaman batang bawah karena menghasilkan semaian yang secara genetik seragam dan identik dengan induknya (true to type) (Kepiro dan Roose 2007). Sementara itu embrio zigotik merupakan hasil fusi gamet jantan dengan sel telur sehingga dapat menghasilkan semaian true to type atau off type. Adanya semaian off type sangat tidak diinginkan dalam perbenihan jeruk batang bawah karena umumnya tanaman kurang vigor dan menurunkan produksi buah batang atas (Andrade-Rodríguez et al. 2004, Hussain et al. 2011).
Semaian off type harus diroguing sehingga meningkatkan mutu genetik benih. Penangkar benih pada umumnya menggunakan penanda morfologi untuk mengidentifikasi semaian off type. Penanda morfologi relatif mudah dan murah, tetapi kebenarannya perlu diuji dengan alat dan metode yang lebih akurat.
Penanda molekuler berbasis protein yaitu analisis isozim dapat membedakan antara semaian off type dan true to type lebih akurat daripada penanda morfologi. Hasil penelitian Farida (2005) menggunakan penanda morfologi dan penanda isozim dapat mengidentifikasi semaian JC off type dan true to type. Namun demikian terdapat perbedaan visualisasi pita yang diperoleh dari sampel umur dua bulan dengan sampel umur 4 bulan. Hal ini menunjukkan bahwa hasil analisis isozim dipengaruhi oleh perkembangan tanaman.
Penanda molekuler berbasis DNA dapat menyempurnakan kelemahan penanda isozim karena visualisasi pita tidak dipengaruhi oleh perkembangan tanaman. Penanda RAPD dapat membedakan semaian true to type dan semaian off type pada batang bawah Volkameriana (Andrade-Rodríguez et al. 2004). Kelemahan penanda RAPD yaitu bersifat dominan sehingga keberadaan genotipe heterozigot dan homozigot tidak dapat dibedakan.
Sementara itu penanda SSR yang digunakan dalam penelitian ini merupakan penanda molekuler yang bersifat kodominan sehingga keberadaan genotipe heterozigot dan homozigot dapat dibedakan. Menurut Ruiz et al. (2000) penanda SSR ini lebih efisien daripada penanda isozim untuk mengidentifikasi semaian batang bawah dari embrio zigotik baik dari penyerbukan sendiri maupun penyerbukan silang.
Meskipun penanda SSR lebih akurat, penggunaan penanda SSR relatif mahal bagi petani. Untuk itu perlu dicari marka yang terpaut dengan karakter morfologi sebagai penanda semaian off type dan true to type. Dengan demikian, penangkar benih jeruk batang bawah JC dapat mengidentifikasi semaian off type dengan penanda yang relatif murah dan mudah tetapi secara genetik telah terbukti berbeda dengan tanaman induknya.
Tujuan
1 Mendapatkan informasi poliembrioni pada jeruk JC
2 Mendeteksi tingkat kemasakan fisiologis benih jeruk JC berdasarkan warna kulit dan kekerasan buah jeruk
3 Mengetahui pengaruh tingkat kemasakan fisiologis benih terhadap daya tumbuh benih, multiple seedling, semaian off type dan semaian true to type 4 Mengidentifikasi semaian true to type dan off type berdasarkan penanda
morfologi dan SSR
Hipotesis
1 Persentase poliembrioni JC lebih dari 85% dengan jumlah embrio 5-10 per benih
2 Benih telah mencapai masak fisiologis bila berat kering dan vigor maksimal, diperoleh pada buah warna kulit lebih dari 90% kuning, lunak.
3 Benih JC masak fisiologis menghasilkan daya tumbuh benih dan multiple seedling terbanyak serta semaian off type paling rendah
4 Multiple seedling dapat meningkatkan total semaian true to type
4
Gambar 1. Bagan alir penelitian
Benih diekstraksi dari buah lima tingkat kemasakan berdasarkan warna kulit buah, dan kekerasan buah Fisiologis Benih, Mutu Genetis dan Jumlah
Semaian
2 TINJAUAN PUSTAKA
Deskripsi Tanaman Jeruk JC
Berbagai genus jeruk (Citrus) berasal dari daerah tropik dan sub-tropik Asia dan kepulauan Malaya, kemudian menyebar ke seluruh bagian dunia. Tanaman jeruk telah dibudidayakan sejak lama dan sebagian besar spesies utama bentuk aslinya tidak diketahui dengan pasti. Klasifikasi Jeruk cukup sulit karena banyaknya jumlah kultivar, hibrida, poliploidi, mutasi dan poliembrioni yang secara alami menyebabkan keragaman. Hal yang sulit dijelaskan adalah hubungan antara satu varietas dengan varietas yang lain, banyak nama lokal dan perubahan nomenklatur dalam klasifikasi (Ray 2002).
Menurut Swingle dan Reece (1967) Japansche Citroen (JC) sebenarnya adalah Rangpur Lime berasal dari India atau Canton Lemon, di Jepang disebut Hime Lemon, dan di Brazil disebut Cravo Lemon. Klasifikasi jeruk JC disesuaikan dengan klasifikasi USDA (2013) :
Tanaman jeruk mempunyai bunga yang tersusun dalam rangkaian bunga majemuk (inflorescentia) tipe cymose dimana inisiasi bunga di mulai dari ujung ke bagian axilar sehingga bunga tidak mekar serentak (Spiegel-Roy dan Goldschmidt 1996). Secara umum struktur bunga jeruk terdiri atas: 1) kelopak yang berbentuk seperti mangkuk terdiri atas 5 sepal, 2) mahkota terdiri atas 5 petal yang terletak berselang seling di atas sepal, 3) 20-40 benang sari yang masing-masing terdiri atas filament dengan ujung anther yang berwarna kuning, 4) putik yang terdiri atas bakal buah (8-14 carpel), stylus dan stigma. Terdapat kelenjar madu seperti subang di atas bakal buah dan melingkari tangkai kepala putik. Pada bagian sudut lokulus pada tiap-tiap carpel berkembang plasenta yang berhubungan dengan bakal benih (Spiegel-Roy dan Goldschmidt 1996, Tjitrosoepomo 2007)
Buah Jeruk merupakan buah basah tipe hesperidium (Tjitrosoepomo 2007). Pericarp jeruk terdiri atas tiga jaringan. Jaringan terluar yang berwarna adalah epicarp atau eksocarp disebut flavedo. Jaringan ditengah putih atau berwarna seperti spon adalah mesocarp disebut albedo. Jaringan terdalam yaitu endocarp tipis seperti selaput. Selaput ini membentuk sekat-sekat, bagian dalamnya membentuk kantong-kantong juice, dan benih terdapat bebas diantara kantong-kantong jus (Spiegel-Roy dan Goldschmidt 1996)
6
merupakan membran tipis yang terbentuk dari integumen bagian dalam bakal benih serta sisa nuselus dan endosperm (tegmen). Sebagian besar volume embrio yang masak dipenuhi oleh kotiledon. Benih poliembrioni mempunyai ukuran kotiledon bervariasi, beberapa embrio mungkin kecil dengan kotiledon yang kurang berkembang (Spiegel-Roy dan Goldscmidt 1996).
Poliembrioni Benih Jeruk
Beberapa varietas jeruk benihnya bersifat poliembrioni, dan beberapa varietas lain seperti jeruk besar benihnya bersifat monoembrioni (Ray 2002). Sifat poliembrioni pada jeruk terjadi karena adanya embrio nuselar. Embrio nuselar berkembang dari jaringan maternal benih yang berkembang bersamaan dengan embrio zigotik sehingga dalam satu benih bisa muncul lebih dari satu semaian (Frost dan Soost 1968). Embrio zigotik merupakan embrio yang terbentuk dari fusi antara gamet jantan dan gamet betina sedangkan embrio nuselar terbentuk dari jaringan nuselar tanpa melalui fertilisasi (Salisbury dan Ross 1992).
Embrio nuselar merupakan bentuk adventif dari reproduksi dimana sel somatik dari jaringan nuselus diinisiasi untuk memasuki lintasan perkembangan embrionik. Embrio nuselar berkembang dari sel inisial nuselus yang berasal dari jaringan nuselus yang mengelilingi kantong embrio. Tidak terdapat kontribusi gamet jantan dalam pembentukan embrio nuselar. Dengan demikian, semaian asal embrio nuselar bersifat identik dengan induknya kecuali terdapat variasi somatik (Kepiro dan Roose 2007).
Berdasarkan hasil pengamatan histologis jeruk manis Valensia, sel inisial embrio nuselar terdapat pada jaringan nuselus di bakal benih, terbentuk sekitar waktu antesis. Sel inisial embrio nuselar dicirikan dengan ukuran sel yang besar, bereaksi kuat pada tes pewarnaan, rerata hanya 2-3 per bakal benih (Koltunow et al. 1995). Embrio nuselar dapat berkembang sampai dengan tingkat awal kotiledon. Namun demikian, embrio nuselar membutuhkan endosperm untuk berkembang lebih lanjut sehingga polinasi dan fertilisasi tetap dibutuhkan (Dhillon et al. 1993 dalam Andrade-Rodriguez et al. 2004) .
Jumlah benih per buah dan benih utuh berkorelasi secara signifikan dengan jumlah embrio per benih. Buah dengan banyak benih mempunyai embrio lebih kecil daripada buah dengan sedikit benih yang menghasilkan beberapa embrio yang besar. Ukuran embrio dipengaruhi oleh asal usul embrio, dari benih monoembrioni atau benih poliembrioni. Embrio yang berkembang pada benih monoembrionik lebih besar daripada yang berkembang pada benih poliembrioni. (Andrade-Rodriguez et al. 2004).
Perkembangan Buah, Benih dan Perkecambahan Benih Jeruk
Proses pemasakan benih adalah perubahan-perubahan morfologik, fisiologik dan fungsional yang terjadi dalam bakal buah dan bakal benih sejak fertilisasi sampai terbentuk benih yang masak fisiologis (Mugnisjah dan Setiawan 1990).
setelah antesis. Pada awal perkembangan buah, flavedo tampak berwarna hijau tua, merupakan jaringan yang aktif berfotosintesis (berklorofil) dengan jumlah stomata relatif sedikit. Saat mendekati waktu masak, klorofil pada kulit buah bagian flavedo perlahan hilang dan kloroplas menjadi kromoplas dengan banyak karotinoid. Bagian dalam flavedo bergabung dengan albedo yang berwarna putih dan membentuk jaringan spon (Spiegel Roy dan Goldscmidt 1996)
Sementara itu perkembangan benih di dalam buah secara fisiologi dibagi menjadi tiga tahapan yaitu 1) histodiferensiasi, 2) pembesaran sel karena terjadi akumulasi cadangan makanan, dan 3) penurunan kadar air. Fase 1 merupakan fase pembelahan sel yang dimulai setelah fertilisasi sehingga terbentuk zigot. Zigot berkembang menjadi embrio serta berkembangnya endosperm. Embrio mencapai fase awal perkembangan kotiledon (berat basah dan berat kering embrio meningkat). Fase 2 merupakan fase pembesaran sel secara cepat disebabkan akumulasi cadangan makanan. Pada fase III benih mencapai tingkat kemasakan fisiologis maksimal yang ditandai dengan berat kering, viabilitas dan vigor maksimal. Pada fase ini kadar air menurun dan hubungan antara benih dengan funikulus terputus (Hartman et al. 1997).
Pembentukan benih tergantung pada kondisi fisiologi yang kompleks yang tergantung asupan air, nutrisi dan hormon, dan sangat dipengaruhi oleh kompetisi diantara bunga dan buah muda. Kondisi pada saat polinasi, fertilisasi dan pembentukan embrio merupakan proses reproduksi yang sangat penting mempengaruhi pembentukan buah dan benih (Frost dan Soost 1968)
Hampir pada semua species citrus benang sari dan putik masak secara bersamaan. Ciri bunga jeruk pada fase antesis adalah kepala putik mengeluarkan eksudat yang menyebabkan tepung sari bisa menempel dipermukaan stigma (Ray 2002). Penyerbukan bunga jeruk terjadi dengan bantuan serangga. Lebah madu merupakan polinator utama terjadinya penyerbukan silang selain thrips dan tungau. Angin juga dapat membantu penyerbukan tetapi merupakan faktor minor. Penyerbukan sendiri dapat terjadi pada genotip yang self-compatible dengan bantuan angin atau kontak langsung antara polen dan stigma (Spiegel-Roy dan Goldscmidt 1996).
Kenampakan cincin berwarna coklat di antara ovarium dan benang sari adalah tanda pertama fruit set. Fertilisasi terjadi 3 hari setelah terjadi polinasi atau bahkan beberapa varietas jeruk dapat mencapai 4 minggu setelah polinasi. Pembelahan zigot dimulai segera dan saat itu pula endosperm sudah multiseluler (Spiegel-Roy dan Goldscmidt 1996)
Benih yang masak mempunyai empat komponen yang secara fisiologis maupun ekologis penting bagi kelangsungan hidupnya yaitu kulit benih sebagai pelindung, embrio, cadangan makanan serta enzim dan hormon yang diperlukan untuk mencerna cadangan makanan dan untuk menyusun jaringan baru dalam semai selama perkecambahan (Gardner et al. 1991). Volume terbesar benih jeruk yang masak merupakan kotiledon yang mengandung lemak benih berkisar 53-54% (Khan et al. 2003).
8
bagian mikrofilar. Sementara itu setelah mengalami fase istirahat, epikotil memanjang ke arah atas dan ujung epikotil berkembang menjadi dua daun pertama (Frost dan Soost 1968).
Penanda Molekuler
Potensi penggunaan penanda sebagai alat untuk melakukan karakterisasi genetik telah dikenal sejak puluhan tahun yang lalu. Penanda bisa dikategorikan sebagai penanda morfologi, sitologi dan yang terbaru adalah penanda molekuler (Moritz dan Hillis 1996; Sessions 1996).
Penanda morfologi merupakan penanda yang telah banyak digunakan karena penanda ini dapat dengan mudah diamati, seperti bentuk daun, warna bunga, warna batang, warna kulit benih, bentuk benih dan sebagainya. Namun penanda ini memiliki kelemahan dapat dipengaruhi oleh kondisi lingkungan, memperlihatkan sifat menurun dominan/resesif, dan mempunyai tingkat keragaman (polimorfisme) rendah atau jumlah yang sedikit (Tanksley et al. 1989).
Penanda Sitologi adalah penanda yang berhubungan dengan kromosom. Contoh penanda sitologi yang telah dipergunakan di dalam pemuliaan tanaman adalah jumlah kromosom, ukuran kromosom dan morfologi set kromosom (Sessions 1996).
Penanda isoenzim merupakan penanda molekuler di tingkat protein yang lebih awal berkembang dan paling sederhana. Polimorfisme protein dideteksi dengan cara elektroforesis, dan perbedaan yang terdeteksi antar alel bergantung pada pergantian asam-asam amino. Analisis genetik organisme tingkat tinggi dengan isoenzim mempunyai keterbatasan apabila tanaman tidak mempunyai penanda genetik yang spesifik. Hal ini disebabkan banyak isoenzim dipengaruhi oleh perkembangan organisme (Soemartono et al. 1992).
Penanda molekuler di tingkat DNA mempunyai kelebihan dibanding penanda molekuler isoenzim. Penanda ini mencerminkan perubahan pada tingkat DNA sehingga menunjukkan jarak genetik yang sesungguhnya antara individu secara akurat. Penanda molekuler di tingkat DNA yang telah berhasil digunakan untuk mengidentifikasi semaian asal embrio zigotik dan semaian asal embrio nuselar antara lain Inter-Simple Sequence Repeat Polymorfism (ISSR), Random Amplified Polymorfic DNA (RAPD) dan Simple Sequence Repeat (SSR) (Ruiz et al. 2000, Tusa et al. 2002, dan Andrade-Rodríguez et al. 2004)
Penanda SSR
Umumnya komposisi DNA eukariotik terdapat empat kelompok urutan nukleotida, yaitu 1) unik (terdapat satu kopi genom), 2) berulang sedikit (terdapat satu sampai sepuluh kopi), 3) berulang sedang (terdapat sepuluh sampai beberapa ratus kopi per genom) dan 4) berulang sangat banyak (beberapa ratus sampai beberapa juta kopi per genom (Yuwono 2005)
Variasi jumlah pengulangan untuk suatu batasan lokus di antara genotipe-genotipe yang berbeda dengan mudah dapat dideteksi dengan PCR (Polimerase Chain Reaction) (Hamada et al. 1982).
Alel-alel DNA mikrosatelit berekspresi dalam bentuk kodominan. Sifat tersebut memungkinkan untuk membedakan antara genotip homozigot dan heterozigot,(Ruiz et al. 2000)
Penanda SSR telah dimanfaatkan untuk studi keragaman genetik dan filogenetik jeruk (Gulsen dan Roose 2001, Golein et al. 2005, dan Hvarleva et al. 2008), identifikasi jeruk hibrida Tangor dan Murcot yang bersifat poliembrioni (Oliveira et al. 2002), dan membedakan semaian dari embrio zigotik dan embrio nuselus pada batang bawah Poncirus trifoliata (L.) Raf. var. ‘Flying Dragon’,
jeruk tangor ‘Ortanique’ dan Fortunella crassifolia Swing. Hasil penelitian pada
jeruk Clementin menunjukkan bahwa 6.6% dari 228 semaian yang dianalisis menunjukkan genotipe yang off type (asal embrio zigotik).
3
METODE PENELITIAN
Waktu dan Tempat Pelaksanaan
Penelitian dilaksanakan di kebun percobaan, nurseri dan laboratorium terpadu Balai Penelitian Tanaman Jeruk dan Buah Subtropika di Tlekung, Batu, Jawa Timur pada bulan April 2012 sampai dengan Januari 2013.
Bahan dan Alat
Bahan yang digunakan yaitu benih dari lima tingkat kemasakan buah berdasarkan warna kulit buah dan kekerasan buah. Pasir, pupuk dan pestisida digunakan dalam persemaian benih JC. Bahan-bahan kimia digunakan untuk pembuatan preparat metode irisan dan analisis DNA.
Alat yang digunakan yaitu alat-alat pertanian, mikroskop binokuler, alat-alat laboratorium untuk analisis DNA, alat ukur dan kamera.
Metodologi
Percobaan I. Studi Poliembrioni Jeruk JC
10
Gambar 2 Buah jeruk JC tingkat kemasakan satu sampai dengan tingkat kemasakan lima
Buah jeruk JC dipotong kemudian diperas sehingga dapat dipisahkan antara benih dan sari buahnya. Benih digosok dengan menggunakan kasa untuk menghilangkan musil yang menyelimuti benih dan dicuci bersih. Benih yang digunakan hanya benih-benih dengan warna kulit yang seragam pada tiap tingkat kemasakan. Benih-benih yang tidak berkembang diamati untuk melihat perkembangan embrionya. Sebanyak 30 benih dikupas dan diamati di bawah mikroskop binokular. Tolak ukur yang diamati adalah sebagai berikut :
1 Warna kulit benih
Benih yang digunakan sebagai sampel yaitu benih yang diambil dari benih-benih yang lebih banyak mempunyai persamaan warna kulit dalam setiap tingkat kemasakan buah.
2 Rerata jumlah embrio per benih
Jumlah embrio masing-masing benih dihitung dan dijumlahkan kemudian dibagi dengan jumlah sampel benih.
Keterangan:
e1 = jumlah embrio benih ke satu en = jumlah embrio benih terakhir 3 Persentase poliembrioni
Persentase poliembrioni dihitung berdasarkan persentase benih yang mempunyai embrio lebih dari satu dari total sampel benih yang diamati.
Persentase semaian poliembrioni dijabarkan menjadi persentase dua embrio per benih, tiga embrio per benih, empat embrio per benih, lima embrio per benih dan enam embrio per benih.
4 Warna embrio dominan
Percobaan II Studi Pengaruh Tingkat Kemasakan Buah terhadap Mutu Fisik, Fisiologis Benih, Mutu Genetis dan Jumlah Semaian Percobaan menggunakan Rancangan Acak Kelompok (RAK) satu faktor lima tingkat kemasakan benih berdasarkan kemasakan buah. Kriteria kemasakan buah seperti halnya percobaan I. Setiap perlakuan diulang tiga kali sehingga terdapat 15 satuan percobaan dengan unit percobaan terdiri 100 benih.
Analisis statistika yang digunakan adalah sidik ragam dengan model Rancangan Acak Kelompok satu faktor sebagai berikut:
Yij = µ + αi + ßj + Ɛij
Keterangan :
Yij = Nilai pengamatan perlakuan tingkat kemasakan ke-i, dan blok ke-k
µ = Rataan umum
αi = Pengaruh perlakuan tingkat kemasakan benih ke-i
βj = Pengaruh blok
εijk= Galat perlakuan tingkat kemasakan ke i
Benih disemai di dalam kotak persemaian yang berisi media pasir dengan jarak tanam 10 cm x 10 cm kemudian diberi fungisida dan pestisida. Penyiraman, dan pengendalian hama penyakit dilakukan sejak semai sampai dengan akhir percobaan.
Mutu Fisik dan Fisiologis Benih
Pengujian benih untuk menentukan saat masak fisiologis dilakukan dengan mengamati kecambah normal yang muncul sejak mulai tanam sampai dengan 30 hari setelah tanam. Benih yang menghasilkan kecambah normal lebih dari satu tetap dihitung satu kecambah normal (ISTA 2010)
Mutu fisik dan fisiologis benih diduga menggunakan tolak ukur : Kadar Air Benih (KA), Berat Kering 10 Benih (BKB), Indeks Vigor (IV), Kecepatan Tumbuh Benih (KCT) dan Daya Berkecambah (DB).
1 Kadar Air Benih (KA)
Pengujian kadar air benih dihitung dengan metode langsung menggunakan oven 103±2 oC selama 17±1 jam (ISTA 2010). Jumlah benih yang diuji 10 benih, sebanyak tiga ulangan dari masing-masing perlakuan tingkat kemasakan buah. Benih diris-iris tipis, dimasukkan dalam wadah aluminium kemudian dioven. Benih yang telah dioven dimasukkan ke dalam desikator selama 30 menit kemudian ditimbang berat keringnya. Rumus pengukuran kadar air sebagai berikut :
Keterangan :
KA = Kadar Air Benih M1 = Berat cawan
12
2 Berat Kering 10 Benih (BKB)
Pengukuran berat kering benih dilakukan dengan mengeringkan 10 benih dalam oven suhu 60 oC selama 3x24 jam, setelah didinginkan dalam desikator beratnya ditimbang. Pengukuran dilakukan empat ulangan untuk setiap satuan percobaan
3 Indeks Vigor (IV)
Indeks Vigor merupakan tolok ukur vigor yang dinilai berdasarkan persentase kecambah normal yang muncul pada pengamatan hitungan pertama (KN1) (Copeland dan McDonald 2001)
4 Kecepatan Tumbuh (KCT)
Pengamatan dilakukan dengan mengamati jumlah kecambah normal yang muncul setiap hari (interval 24 jam) hingga pengamatan kecambah hitungan terakhir.Kecepatan tumbuh benih dihitung berdasarkan jumlah pertambahan persentase kecambah normal/etmal (Sadjad et al. 1999).
Keterangan :
KCT = Kecepatan Tumbuh
N = kurun waktu perkecambahan
d = persentase kumulatif kecambah normal per etmal 5 Daya Berkecambah (DB)
Pengukuran daya berkecambah benih berdasarkan ISTA (2010). Sebanyak 100 benih dari masing-masing perlakuan dikecambahkan dalam bak pasir. Kecambah normal dihitung pada umur 21 hari (hitungan pertama) dan 30 hari (hitungan kedua) setelah tanam (Mulsanti 2002).
Keterangan :
DB = Daya Berkecambah Benih
KN I = jumlah kecambah normal pada hitungan pertama KN II = jumlah kecambah normal pada hitungan kedua Mutu Genetis dan Jumlah Semaian
Setelah pengujian perkecambahan (DB, IV, KCT) selesai dilakukan,
1 Daya Tumbuh Benih
Daya tumbuh dihitung saat tiga bulan setelah tanam untuk memberi kesempatan benih-benih poliembrioni menghasilkan semaian. Pengamatan daya tumbuh dilakukan berdasarkan jumlah benih yang tumbuh menjadi semaian dibagi dengan jumlah benih yang ditanam. Multiple seedling hanya dihitung satu semaian.
2 Persentase Multiple Seedling
Persentase multiple seedling dihitung berdasarkan persentase benih yang menghasilkan semaian lebih dari satu pada setiap unit percobaan.
Keterangan :
∑ BTMS = Jumlah benih tumbuh menjadi multiple seedling.
∑ BTS = Jumlah benih yang tumbuh menjadi semaian (multiple seedling dan
semaian tunggal).
Persentase multiple seedling jeruk JC dijabarkan menjadi persentase dua semaian per benih, tiga semaian per benih dan empat semaian per benih. 3 Total Semaian Hidup
Total semaian hidup dihitung berdasarkan total semaian yang tumbuh pada setiap unit percobaan. Multiple seedling dihitung secara keseluruhan.
4 Persentase Semaian Off type
Persentase semaian off type dihitung berdasarkan persentase semaian off type dari total semaian hidup. Semaian off type yang dihitung yaitu semaian yang secara morfologi berbeda dengan semaian JC pada umumnya (true to type)
5 Total Semaian True to type
Persentase semaian true to type dihitung berdasarkan persentase semaian true to type dari total semaian hidup.
Analisis Data
14
PERCOBAAN III Identifikasi Semaian Jeruk dengan Penanda Morfologi dan Penanda Molekuler
Identifikasi Semaian Jeruk dengan Penanda Morfologi
Sebanyak 12 sampel semaian dari percobaan II digunakan dalam percobaan III. Semaian no 1, 2, dan 3 merupakan sampel dari semaian true to type (semaian dengan karakter morfologi yang sama dengan semaian JC pada umumnya) sedangkan semaian no 4-12 merupakan sampel dari semaian off type (semaian yang secara morfologi berbeda dengan semaian JC pada umumnya). Semaian no 3 dan no 11 berasal dari satu benih. Kedua semaian tersebut dipisahkan saat semaian berumur dua bulan setelah tanam. Identifikasi secara morfologi terdiri atas karakter kuantitatif dan kualitatif.
Karakter kuantitatif diamati saat semaian berumur enam bulan setelah tanam untuk memberi kesempatan pada semaian yang tumbuh dari benih poliembrioni. Tolak ukur yang diamati yaitu : tinggi semaian, jumlah daun, rasio panjang dan lebar daun, dan LAMI [Leaf Apex Morphometric Index] yaitu rasio panjang dan lebar pucuk daun.
1 Tinggi Semaian
Tinggi semaian diukur mulai pangkal semaian sampai dengan ujung semaian. Pangkal semaian ditandai dengan spidol permanen satu bulan setelah tanam, dan tinggi semaian diukur tiga bulan setelah tanam.
2 Jumlah Daun
Jumlah daun dihitung saat tiga bulan setelah tanam. Daun yang dihitung merupakan daun yang telah membuka.
3 Rasio Panjang dan Lebar Daun
Pengukuran dilakukan pada daun keempat dari ujung semaian. Panjang daun diukur dari pangkal ke pucuk daun. Lebar daun diukur pada bagian tengah daun. Rasio panjang dan lebar daun dihitung dengan rumus :
4 LAMI (Rasio Panjang dan Lebar Pucuk Daun)
LAMI (rasio panjang dan lebar pucuk daun) diukur berdasarkan Oliveira et al. (2002). Pengukuran dilakukan pada daun keempat dari ujung semaian. Helai daun dibagi menjadi tiga kemudian pada 1/3 bagian ujung daun diukur lebarnya. Panjang daun diukur dari 1/3 bagian ujung daun bagian tengah ke ujung daun. LAMI dihitung dengan rumus :
Karakter kualitatif diamati 1 minggu setelah semaian muncul dan 3 bulan setelah tanam. Pengamatan dilakukan sejak awal untuk melihat kemungkinan adanya perubahan karakter morfologi. Tolak ukur yang diamati yaitu :
1 Keberadaan Duri
Pengamatan dilakukan pada semaian untuk mengetahui apakah sampel semaian mempunyai duri atau tidak.
2 Keberadaan Stipula
Stipula yaitu daun tambahan pada petiol sehingga disebut juga sebagai sayap petiol. Pengamatan dilakukan pada semaian untuk mengetahui apakah sampel mempunyai stipula atau tidak
3 Tipe Daun
Berdasarkan Descriptor List for Citrus (IPGRI 1999) tipe daun dibagi menjadi dua yaitu tipe daun tunggal dan tipe daun trifoliata atau yang menyerupai tipe daun trifoliata.
4 Warna Pupus Daun dan Daun Masak
Pengamatan dilakukan secara visual dengan membandingkan warna pupus daun dan daun masak sampel semaian dengan semaian JC true to type
5 Bentuk Daun
Bentuk helai daun, ujung daun, dan tepi daun diamati pada daun keempat dari ujung (IPGRI 1999, Tjitrosoepomo 2007). Tipe daun, bentuk helai daun dan tepi daun diamati berdasarkan Descriptor list for Citrus (IPGRI 1999). Gambar skematik bentuk helai dan tepi daun tampak dalam Gambar 3 dan 4.
Gambar 3 Bentuk helai daun jeruk berdasarkan Descriptor list for citrus, IPGRI 1999
16
Gambar 4 Bentuk tepi daun jeruk berdasarkan Descriptor list for citrus, IPGRI 1999
Keterangan : 1. Crenate (bergerigi halus), 2. Dentate (beringgit), 3. Entire (rata), 4. Sinuate (bergelombang)
Identifikasi Semaian dengan Penanda SSR
Secara garis besar, analisis DNA dengan penanda SSR terdiri atas ekstraksi DNA, uji kualitas dan kuantitas DNA dengan elektroforesis, amplifikasi dan separasi DNA dengan PCR, visualisasi hasil PCR dan analisis data molekuler. Ekstraksi DNA
Ekstraksi DNA dilakukan terhadap masing-masing sampel daun muda dari 2 pohon induk, 3 sampel semaian true to type dan 9 sampel semaian off type. Ekstraksi DNA dilakukan dengan metode CTAB yang dimodifikasi (Triani 2010). Buffer ekstraksi sebanyak 1 ml (60 ml CTAB 3%, 47.6 ml NaCl 1.4 M; 8.l ml EDTA 20 mM dan 20 ml Tris-HCl 100 mM) dipanaskan dalam waterbath 65
o
C selama 15 menit. Daun muda dari pohon induk dan semaian JC diambil dan dibersihkan dengan alkohol 70% lalu ditimbang tanpa tulang daunnya sebanyak 0.3 gram. Sampel daun ditambah 0.5 gram PVP digerus, ditambahkan sebagian buffer kemudian dimasukkan dalam tabung ependorf yang telah berisi sebagian buffer ekstraksi dan ditambahkan 5µl mercaptoethanol.
Larutan kemudian dihomogenisasi dengan vortex dan diinkubasi dalam water bath dengan suhu 65 oC selama 30 menit. Setiap 10 menit tabung eppendorf yang berisi larutan dibolak balik. Setelah 30 menit inkubasi, ditambahkan 700 µl Chlorofom:Isoamylalcohol (CHISAM) (24:1), dihomogenisasi dengan vortex kemudian larutan dipisahkan dengan sentrifuse kecepatan 6000 rpm selama 10 menit.
Supernatan (bagian atas) diambil dan dimasukkan ke dalam ependorf yang baru, ditambahkan 1ml CHISAM dan dipisahkan dengan sentrifuge kecepatan 6000 rpm selama 10 menit. Supernatan diambil kembali dan dimasukkan dalam ependorf baru, ditambah 1ml isopropanol dingin kemudian diaduk secara perlahan. Larutan dimasukkan dalam freezer selama 30 menit. Setelah 30 menit larutan dipisahkan dengan sentrifuse kecepatan 6000 rpm selama 10 menit.
disuspensi dengan 50 µl buffer TE dan simpan dalam suhu -4 oC sampai digunakan dalam amplifikasi DNA.
Uji Kualitas dan Kuantitas DNA
Kualitas DNA diamati dengan metode Sambrook et al. (1989) yang dimodifikasi. Langkah-langkah elektroforesis sebagai berikut:
1. Agarose 1% dipanaskan sampai mendidih, dibiarkan sampai hangat kuku kemudian ditambahkan EtBr 1/10.000 volume. Setelah dikocok rata larutan langsung dituang ke dalam plate sampai keras.
2. Gel direndam ke dalam elektroforesis chamber kemudian campuran 5µl DNA dan 1 µl loading dye dimasukkan dalam sumur (wells).
3. Elektroforesis dilakukan dengan tegangan 50 Volt selama 1 jam.
4. Visualisasi hasil elektroforesis dilakukan di atas lampu ultraviolet dengan mesin Biodoc Analize.
Amplifikasi dan Separasi DNA
Sebelum dilakukan amplifikasi DNA dengan PCR, terlebih dahulu dilakukan optimasi program reaksi PCR untuk memperoleh kondisi optimum PCR yang dapat digunakan untuk amplifikasi DNA dengan primer mikrosatelit yang telah ditentukan. reverse) seperti tampak pada Tabel 1.
Reaksi amplifikasi PCR dilakukan dengan menggunakan alat Biometra Thermocycler. Setiap tabung PCR berisi 25 µl total volume yang terdiri atas 12.5 µl PCR mix (Fermentas Dream Taq TM Green Master Mix 2x), 2.5 µl air bebas nuklease, 3 µl primer forward, 3 µl primer reverse dan DNA template 4 µl
Tabel 1 Urutan basa enam primer SSR (forward dan reserve)
Primer Urutan Basa
Forward Reverse
AG14 5’AAAGGGAAAGCCCTAATCTCA3’ 5’CTTCCTCTTGCGGATGTTC3’
CAC23 5’ATCACAATTACTAGCAGCGCC3’ 5’TTGCCATTGTAGCATGTTGG3’
AMB5 5' CCCTGCACAAAAACTCACAC3' 5' TGGGGGTGTTGAATGGTAAT3'
ATC09 5’TTCCTTATGTAATTGCTCTTTG3’ 5’TGTGACTGTTTGTGCGTCTG3’
TAA15 5’GAAAGGGTTACTTGACCAGGC3’ 5’CTTCCCAGCTGCACAAGC
TAA27 5’GGATGAAAAATGCTCAAAATG3’ 5’TAGTACCCACAGGGAAGAG3’
Sumber : Scarano et al. 2003, Jannati et al. 2009, Triani 2010.
Visualisasi Hasil PCR
18
Visualisasi dilakukan diatas lampu ultraviolet dengan menggunakan alat Biodoc Analyze.
Bulk Segregant Analysis (BSA)
Metode Bulk Segregant Analysis (BSA) digunakan untuk menyeleksi marka-marka yang terpaut dengan karakter semaian asal embrio zigotik dan semaian true to type. Hasil identifikasi asal semaian dengan penanda morfologi dibuat pool DNA semaian off type dan pool DNA semaian true to type.
Analisis data dilakukan berdasarkan persentase kehadiran pita pada kedua pool. Marka yang 100% teramplifikasi pada pool satu dan tidak teramplifikasi pada pool yang lain berarti marka tersebut spesifik untuk karakter morfologi yang diamati, dan juga terpaut dengan semaian off type atau semaian true to type. Analisis Gerombol
Hasil pengamatan morfologi semaian diterjemahkan dalam bentuk data biner. Skoring pita DNA dilakukan berdasarkan keberadaan pita pada setiap sampel semaian. Profil pita DNA selanjutnya diterjemahkan ke dalam data biner berdasarkan keberadaan pita (1) dan tidak ada pita (0) untuk membangun matriks kemiripan. Analisis gerombol berdasarkan penanda morfologi dan penanda SSR dilakukan dengan menggunakan prosedur SIMQUAL dan dihitung berdasarkan metode SAHN pada program NTSysSPc versi 2.02. Hasil analisis disajikan dalam bentuk dendrogram.
4 HASIL DAN PEMBAHASAN
Percobaan I Studi Poliembrioni pada jeruk JC
Karakter poliembrioni benih JC perlu dipelajari untuk melihat pengaruhnya terhadap mutu benih. Hasil pengamatan karakteristik benih dan embrio JC pada lima tingkat kemasakan buah disajikan dalam Tabel 2.
Warna kulit benih pada umumnya dapat menjadi penanda tingkat kemasakan embrio di dalam benih karena warna embrio sangat mempengaruhi warna kulit benih. Hasil pengamatan menunjukkan bahwa 87.85% benih dari buah tingkat kemasakan I berwarna krem kehijauan. Buah tingkat kemasakan II menghasilkan ± 80.42% benih berwarna krem, sedangkan buah tingkat kemasakan III menghasilkan ± 91.31% benih berwarna krem. Buah dari tingkat kemasakan IV menghasilkan ± 87.3% benih berwarna krem kecoklatan sedangkan buah dari tingkat kemasakan V hanya terdapat ± 80.54% benih berwarna krem kecoklatan. Hasil penelitian Hamilton (2007) pada Citrus garrawayi juga menunjukkan bahwa di dalam buah jeruk dengan ukuran besar (panjang 7 cm) hanya terdapat 76% benih yang terlihat dari warna kulit benih dan warna embrio dalam benih cenderung berwarna krem.
Persentase poliembrioni benih JC tertinggi diperoleh pada buah dari tingkat kemasakan III. Jeruk JC mempunyai 1-6 embrio per benih dengan rerata 1.67-3 embrio per benih. Meskipun benih JC dapat menghasilkan sampai enam embrio per benih kejadiannya sangat jarang bahkan sebagian besar benih JC poliembrioni hanya mempunyai dua embrio per benih. Terdapat keragaman rerata jumlah embrio per benih pada lima tingkat kemasakan benih berdasarkan kemasakan buah. Hal ini diduga disebabkan perbedaan waktu polinasi antara kelima tingkat kemasakan buah. Menurut Andrade-Rodriguez et al. (2004), faktor-faktor lingkungan yang mempengaruhi polinasi dan fertilisasi dapat mempengaruhi persentase poliembrioni dan jumlah embrio per benih.
Tabel 2 Karakteristik benih dan poliembrioni jeruk JC pada lima tingkat kemasakan buah
Tolak ukur pengamatan Tingkat Kemasakan
I II III IV V
Hasil pengamatan menunjukkan bahwa sebagian besar embrio yang dominan di dalam benih tingkat kemasakan I mempunyai kotiledon berwarna krem kehijauan dan plumula, radikula yang berwarna hijau muda. Sementara itu beberapa embrio lain dalam benih yang sama menunjukkan warna kotiledon, plumula dan radikula hijau muda (Gambar 5A).
Perkembangan tingkat kemasakan embrio dapat diduga berdasarkan perubahan warna embrio. Menurut Werker (1997), warna hijau pada embrio menunjukkan adanya klorofil dan pada tahap akhir kemasakan benih terjadi perubahan warna yang disebabkan klorofil mengalami degradasi. Dengan demikian, embrio yang masih berwarna hijau menunjukkan bahwa embrio tersebut belum masak.
20
Gambar 5 Keragaan poliembrioni benih jeruk JC dari buah tingkat kemasakan I -V
Keterangan : A. tingkat kemasakan I, B. Tingkat kemasakan II, C. Tingkat kemasakan III, D. tingkat kemasakan IV, E. tingkat kemasakan V, e1= embrio dominan, e2-e5 = embrio selain embrio dominan, pr = plumula dan radikula, m = mikrofilar
Hasil pengamatan pada benih dari tingkat kemasakan buah II dan III menunjukkan bahwa embrio yang mendominasi benih mempunyai kotiledon yang telah berwarna krem, sedangkan plumula dan radikula masih berwarna hijau muda
m
m
m
m
(Gambar 5B, 5C). Sementara itu embrio-embrio selain embrio dominan dalam benih tingkat kemasakan II menunjukkan kotiledon yang berwarna krem kehijauan dengan plumula radikula berwarna hijau muda. Embrio-embrio selain embrio dominan dalam benih dari buah tingkat kemasakan III menunjukkan bahwa embrio-embrio selain embrio dominan masih berwarna hijau muda (Gambar 5C).
Benih dari buah tingkat kemasakan buah IV dan V menunjukkan warna embrio dominan yang sama yaitu kotiledon, plumula dan radikula berwarna krem. Embrio selain embrio dominan di dalam benih dari buah tingkat kemasakan IV cenderung mempunyai tingkat kemasakan yang hampir sama dengan embrio dominannya (Gambar 5D). Hal yang berbeda ditunjukkan pada benih dari buah tingkat kemasakan V yaitu terdapat beberapa embrio selain embrio dominan yang berwarna coklat. Embrio-embrio yang berwarna coklat menunjukkan bahwa embrio tersebut telah mati (Gambar 5E)
Hasil pengamatan embrio pada benih dari tingkat kemasakan I-V seperti tersebut diatas menunjukkan bahwa poliembrioni menyebabkan keragaman tingkat kemasakan embrio di dalam benih sehingga menyebabkan keragaman kemasakan benih dalam suatu lot benih. Spiegel-Roy dan Goldschmidt (1996) juga menyatakan bahwa pada benih jeruk poliembrioni, ukuran kotiledon sering beragam dan beberapa embrio kecil dengan perkembangan kotiledon yang lambat. Sampel benih poliembrioni dari buah tingkat kemasakan I –V menunjukkan bahwa embrio dominan dan embrio-embrio lainnya tampak bergerombol di mikrofilar dengan bentuk yang kadang-kadang tidak beraturan (Gambar 5). Menurut Kishore et al. (2012), posisi embrio di mikrofilar diduga disebabkan ketergantungan embrio nuselar terhadap perkembangan endosperm. Keberadaan endosperm mendukung perkembangan embrio nuselar pada mikrofilar tetapi menekan pertumbuhannya di khalazal.
Selain dari 30 benih yang digunakan sebagai sampel, pada buah tingkat kemasakan V ditemukan satu benih dengan embrio-embrio yang berkembang hanya sampai dengan fase hearth (jantung) (Gambar 6). Aleza et al. (2010) juga menemukan benih rudimenter pada jeruk hasil persilangan. Embrio-embrio berkembang sampai dengan fase globular dan torpedo.
22
Gambar 6 Embrio-embrio pada benih rudimenter yang ditemukan pada buah tingkat kemasakan V terdiri atas embrio fase globular (G) dan fase hearth (H)
Percobaan II. Studi Pengaruh Tingkat Kemasakan Buah terhadap Mutu Fisik, Fisiologis Benih, Mutu Genetik dan Jumlah Semaian
Mutu Fisik dan Fisiologis Benih
Viabilitas dan Vigor benih diukur berdasarkan perhitungan kecambah normal yang ditandai dengan munculnya dua daun pertama berwarna hijau dengan bentuk daun sempurna dan akar primer yang lurus. Hasil pengamatan ada dua kriteria kecambah abnormal yang muncul dalam persemaian antara lain kecambah yang tidak berklorofil (bule), daun melengkung dan daun melilit. Kecambah yang tidak berklorofil, daun melengkung dan daun melilit sampai dengan umur enam bulan tidak mengalami perkembangan yang signifikan bahkan beberapa mengalami kematian.
Rekapitulasi hasil analisis sidik ragam (Tabel 3) menunjukkan bahwa tingkat kemasakan benih berpengaruh nyata pada taraf 5% terhadap kadar air benih (%), indeks vigor (%), kecepatan tumbuh (%/etmal) dan daya berkecambah (%). Hasil analis ragam menunjukkan pula bahwa koefisien keragaman kadar air benih JC 10.31. Menurut Chin 1988 dalam Adimargono 1997, kadar air benih individu-individu dalam lot benih rekalsitran sangat bervariasi sehingga koefisien keragamannya tinggi (5-13.4) dan lebih besar dibandingkan dengan benih ortodoks (2.36-2.86).
Tabel 3 Rekapitulasi hasil analisis ragam kadar air, berat kering 10 benih, indeks vigor, kecepatan tumbuh dan daya berkecambah
Sumber keragaman
Tingkat kemasakan Benih berdasarkan
kemasakan buah
Koefisien Keragaman
(%)
Kadar air benih (%) * 10.31
Berat kering 10 benih (g) tn 6.63
Indeks vigor (%) * 20.73
Kecepatan tumbuh (%/etmal) * 8.66
Daya berkecambah (%) * 7.07
Koefisien keragaman indeks vigor sangat tinggi hingga 20.73 meskipun telah dilakukan transformasi. Tingginya keragaman diduga disebabkan keragaman tingkat kemasakan embrio dalam satu benih sebagaimana hasil percobaan I. Keragaman tingkat kemasakan embrio per benih meningkatkan keragaman kemasakan benih di dalam lot benih sehingga benih berkecambah tidak serentak. Benih dengan embrio yang lebih masak dapat muncul terlebih dahulu dibanding dengan benih dengan embrio-embrio yang belum masak.
Hasil analisis ragam yang berbeda nyata diuji lanjut dengan DMRT. Rerata tolak ukur kadar air, berat kering 10 benih, indeks vigor, kecepatan tumbuh dan daya berkecambah disajikan dalam Tabel 4.
Tabel 4 Pengaruh tingkat kemasakan buah terhadap mutu fisik dan fisiologis Keterangan : Angka yang diikuti huruf yang sama dan pada kolom yang sama tidak berbeda nyata
dengan Duncan Multiple Range Test (DMRT) pada taraf 5%. KA : Kadar Air benih, BKB : Berat Kering 10 Benih, IV : Indeks Vigor ; KCT : Kecepatan Tumbuh, DB :
Daya Berkecambah
Kadar Air Benih
Tabel 4 menunjukkan bahwa benih dari tingkat kemasakan I yang merupakan fase buah termuda yang digunakan dalam penelitian ini mempunyai kadar air tertinggi yaitu 48.21%. Semakin masak benih kecenderungan kadar air menurun. Kadar air benih JC paling rendah yaitu pada buah tingkat kemasakan V yang tidak berbeda nyata dengan buah tingkat kemasakan IV. Kadar air benih tingkat kemasakan III tinggi dan tidak berbeda nyata dengan tingkat kemasakan I. Tingginya kadar air benih pada buah tingkat kemasakan I diduga disebabkan rerata jumlah embrio per benih paling tinggi dibandingkan dengan keempat tingkat kemasakan benih (Percobaan I). Sebagian embrio di dalam benih tingkat kemasakan III masih belum masak (embrio masih berwarna hijau) sehingga kadar air benih di dalam embrio masih tinggi.
Berat Kering 10 Benih
Hasil analisis ragam menunjukkan berat kering 10 benih tidak berbeda nyata pada lima tingkat kemasakan benih berdasarkan kemasakan buah. Pada umumnya berat kering maksimum dapat menjadi indikator tercapainya saat masak fisiologi (Hartmann et al. 1997). Namun demikian dalam kasus benih poliembrioni seperti pada benih JC, berat kering benih tidak dapat digunakan dalam penentuan tingkat kemasakan fisiologis benih.
24
berdasarkan hasil pengamatan pada percobaan I terdapat keragaman jumlah dan ukuran embrio di dalam benih serta tingkat kemasakan embrio yang tidak seragam. Keragaman ukuran dan tingkat kemasakan embrio di dalam benih diduga menyebabkan hasil pengukuran berat kering 10 benih pada lima tingkat kemasakan buah tidak berbeda nyata. Berdasarkan hal tersebut, pengukuran berat kering benih poliembrioni untuk menentukan tingkat kemasakan fisiologis benih dapat lebih akurat apabila hanya menggunakan sampel embrio dominan saja tanpa menyertakan embrio lainnya di dalam satu benih.
Vigor Benih
Pengamatan vigor benih dalam penelitian ini menggunakan tolak ukur indeks vigor dan kecepatan tumbuh. Indeks vigor tertinggi dicapai pada benih tingkat kemasakan IV berbeda nyata dengan keempat tingkat kemasakan yang lain. Menurut Copeland dan McDonald (2001) benih yang berkecambah normal pada hitungan pertama mengindikasikan kekuatan benih dalam menghadapi kompetisi.
Nilai kecepatan tumbuh tertinggi pada tingkat kemasakan IV berbeda nyata dengan tingkat kemasakan I dan V, tapi tidak berbeda nyata dengan tingkat kemasakan II. Menurut Sadjad (1993), tolak ukur kecepatan tumbuh mengidikasikan vigor kekuatan tumbuh karena benih yang cepat tumbuh lebih mampu menghadapi kondisi lapang yang suboptimum.
Berdasarkan tolak ukur indeks vigor dan kecepatan tumbuh, buah tingkat kemasakan IV benih yang dihasilkan mempunyai vigor paling tinggi. Penelitian Hamilton 2007 pada Citrus garrawayi juga menunjukkan bahwa benih masak lebih cepat berkecambah daripada benih belum masak.
Daya Berkecambah
Viabilitas benih maksimal dicapai pada benih dari buah tingkat kemasakan IV dengan nilai daya berkecambah 74.33 % berbeda nyata dengan tingkat kemasakan I, III, dan V tetapi tidak berbeda nyata dengan benih dari buah tingkat kemasakan II. Nilai daya berkecambah terendah pada benih dari tingkat kemasakan I berbeda nyata dengan keempat tingkat kemasakan yang lain.
Benih dari buah tingkat kemasakan IV mempunyai daya berkecambah, indeks vigor dan kecepatan tumbuh paling tinggi dibanding benih dari buah tingkat kemasakan yang lain. Berdasarkan hal tersebut dapat disimpulkan bahwa tingkat kemasakan fisiologis benih dicapai pada tingkat kemasakan buah dengan ciri warna kulit buah kuning 90% merata (tingkat kemasakan IV). Benih yang telah masak fisiologis telah mempunyai cadangan makanan yang sempurna yang dapat mendukung pertumbuhan kecambah. Menurut Qodir dalam Widajati et al. (2012), benih yang mempunyai viabilitas awal benih yang tinggi akan mempunyai daya simpan yang lebih lama daripada benih dengan viabilitas awal rendah. Dengan demikian, panen buah batang bawah JC sebaiknya dilakukan pada saat benih mencapai masak fisiologis sehingga diperoleh benih dengan mutu fisik dan fisiologis yang tinggi.
daya berkecambah, kecepatan tumbuh dan kemampuan berkecambah pada hitungan pertama.
Benih dari buah tingkat kemasakan I mempunyai vigor dan viabilitas yang terendah. Benih yang dihasilkan dari buah dengan karakteristik warna kulit buah hijau merata seperti tingkat kemasakan I belum memasuki kemasakan fisiologis sehingga tidak disarankan digunakan dalam perbenihan. Hasil studi morfologi benih pada percobaan I menunjukkan sebagian besar benih dari buah tingkat kemasakan I mempunyai embrio dengan warna krem kehijauan sedangkan benih dari buah tingkat kemasakan IV sebagian besar didominasi embrio berwarna krem.
Benih dari buah tingkat kemasakan II dan III mempunyai daya kecambah, indeks vigor dan kecepatan tumbuh yang tidak berbeda nyata. Buah yang dipanen pada tingkat kemasakan II dan III dapat menghasilkan benih dengan tingkat kemasakan yang sama meskipun dari segi warna kulit buah, warna daging buah dan kekerasan buah berbeda. Percobaan I menunjukkan bahwa buah tingkat kemasakan II dan III menghasilkan benih-benih yang sebagian besar mempunyai warna kulit, warna kotiledon dominan dan warna radikula dan plumula yang sama.
Vigor dan viabilitas benih dari buah tingkat kemasakan V lebih rendah daripada benih dari tingkat kemasakan IV sehingga benih pada fase ini diduga telah melewati masak fisiologi. Benih yang dipanen pada tingkat kemasakan buah V dapat merugikan karena dapat menghasilkan benih dengan mutu fisik dan fisiologis yang rendah.
Mutu Genetik dan Jumlah Semaian
Hasil analisis ragam (Tabel 5) menunjukkan bahwa tingkat kemasakan benih tidak berpengaruh nyata terhadap persentase multiple seedling dan persentase semaian off type. Tingkat kemasakan benih berpengaruh nyata terhadap daya tumbuh benih, total semaian hidup dan total semaian true to type. Tabel 5 Rekapitulasi hasil analisis ragam daya tumbuh benih, persentase multiple
seedling, total semaian hidup, persentase semaian off type dan total
Keterangan : *) berbeda nyata pada taraf 5%; tn=tidak berbeda nyata