i
PENINGKATAN MUTU BENIH PADI MELALUI
PELLETING
MENGGUNAKAN BAKTERI PROBIOTIK UNTUK
MENEKAN
Xanthomonas oryzae
pv. o
ryzae
ANAK AGUNG KESWARI KRISNANDIKA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
iii
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK
CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Peningkatan Mutu Benih Padi Melalui Pelleting Menggunakan Bakteri Probiotik untuk Menekan Xanthomonas oryzae pv. oryzae adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2016
A. A. Keswari K
RINGKASAN
ANAK AGUNG KESWARI KRISNANDIKA. Peningkatan Mutu Benih Padi Melalui Pelleting Menggunakan Bakteri Probiotik untuk Menekan
Xanthomonas oryzae pv. Oryzae. Dibimbing oleh ENY WIDAJATI, WAWAN HERMAWAN dan GIYANTO.
Bakteri probiotik yang berasal dari dalam jaringan tanaman padi (endofit 467 dan endofit 748), rhizosfer (Ralstonia pickettii TT47) serta berasal dari tanah (aktinomiset 6) diketahui mampu mengendalikan patogen. Kemampuan bakteri bertahan dalam bentuk pelet dan menekan patogen
Xanthomonas oryzae pv. oryzae serta mempertahankan viabilitas benih padi terinfeksi belum diketahui, sehingga diuji pada penelitian ini.
Uji in vitro menunjukkan isolat yang memiliki kemampuan antagonis terhadap X. oryzae pv. oryzae adalah R. pickettii TT47, endofit 467 dan aktinomiset 6. Isolat yang kompatibel yaitu endofit 467 dan aktinomiset 6. Isolat-isolat tersebut kemudian ditambahkan ke dalam formula pelet untuk diuji daya simpannya. Formula pelet (talek + CMC 1.5% + gliserol 1%) terbukti mampu mempertahankan daya simpan bakteri probiotik R. pickettii
TT47, endofit 467, aktinomiset 6 dan endofit 467 + aktinomiset 6 selama tiga minggu penyimpanan. Selanjutnya, formula tersebut diaplikasikan pada benih padi yang telah diinfeksi patogen Xanthomonas oryzae pv. oryzae. Populasi patogen dan viabilitas benih dihitung tiap 2 minggu. Formula pelet (talek + CMC 1.5% + gliserol 1%) dengan penambahan aktinomiset 6 tunggal maupun kombinasi dengan endofit 467 pada benih padi Ciherang yang terinfeksi, merupakan formula terbaik dalam menekan populasi X. oryzae pv.
oryzae selama 6 minggu penyimpanan. Persentase daya berkecambah dan kecepatan tumbuh tertinggi pada benih padi terinfeksi selama 6 minggu penyimpanan dihasilkan oleh perlakuan pelet dengan penambahan R. pickettii
TT47 yaitu 86.67% dan 17.17% etmal-1 dan nyata lebih tinggi dibanding dengan benih terinfeksi X. oryzae pv. oryzae yang tidak diberi perlakuan pelet (62.67% dan 11.02% etmal-1).
Aplikasi bakteri probiotik R. pickettii TT47, aktinomiset 6 atau aktinomiset 6 + endofit 467 dalam bentuk pelet terbukti efektif menurunkan populasi patogen X. oryzae pv. oryzae dan mempertahankan viabilitas benih padi terinfeksi selama 6 minggu penyimpanan.
v
SUMMARY
ANAK AGUNG KESWARI KRISNANDIKA. Rice Seed Quality Enhancement by Pelleting using Probiotic Bacteria to Suppress
Xanthomonasoryzae pv. oryzae. Supervised by ENY WIDAJATI, WAWAN HERMAWAN and GIYANTO.
Probiotic bacteria included endophytic bacteria (strain 467 and 748), rhizosphere bacteria (Ralstonia pickettii TT47) and soil bacteria (Actinomycetes 6) has been reported as biocontrol agents for the disease. Since the bacteria's ability to survive in pellets form, suppressing X. oryzae
pv. oryzae and maintaining viability of rice seeds infected with X. oryzae pv.
oryzae has not been known thoroughly, this research was carried out to evaluate it.
In vitro test revealed that isolates R. pickettii TT47, endophytic 467 and actinomycetes 6 showed antagonistic activities against X. oryzae pv.
oryzae. Among them, only endophytic 467 and actinomycetes 6 that showed compatibility. Those isolate then adding into pellets formulation to know their shelf life. Pellets formulation proved capability to maintain the shelf life of isolate R. pickettii TT47, endophytic 467, actinomycetes 6 and actinomycetes 6 + endophytic 467 for three weeks of storage. Subsequently, the formula was applied to the rice seeds that have been infected by the pathogen
Xanthomonas oryzae pv. oryzae. Populations of pathogens in seeds were counted every 2 weeks. Pellets formulation (talc + CMC 1.5% + glycerol 1%) with adding actinomycetes 6 in single or in combination with endophytic 467 on Ciherang infected rice seeds is the best formulation to suppress the population of X. oryzae pv. oryzae as long as 6 weeks of storage. The highest germination percentage and growth rate in infected rice seeds during 6 weeks of storage was produced by pellets treatment with the addition of R. pickettii
TT47 i.e. 86.67% and 17.17% etmal-1, respectively and significantly higher than non-pelleted rice seed infected (62.67% and 11.02% etmal-1).
In conclusion, the application of probiotic bacteria R. pickettii TT47, actinomycetes 6 and actinomycetes 6 + endophytic 467 in pellet formulation effective to decrease X. oryzae pv. oryzae and maintain viability of infected rice seed during 6 weeks storage.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
vii
Tesis
sebagai salah satu syarat untuk mendapatkan gelar Magister Sains
pada
Program Studi Ilmu dan Teknologi Benih
ANAK AGUNG KESWARI KRISNANDIKA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PENINGKATAN MUTU BENIH PADI MELALUI
PELLETING
ix
Judul : Peningkatan Mutu Benih Padi Melalui Pelleting Menggunakan Bakteri Probiotik untuk Menekan
Xanthomonas oryzae pv. oryzae
Nama : Anak Agung Keswari Krisnandika NIM : A251130031
Disetujui oleh Komisi Pembimbing
Dr Ir Eny Widajati, MS Ketua
Dr Ir Wawan Hermawan, MS Anggota
Dr Ir Giyanto, MSi Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu dan Teknologi Benih
Dr Ir Endah Retno Palupi, MSc Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian dengan judul Peningkatan Mutu Benih Padi Melalui Pelleting Menggunakan Bakteri Probiotik untuk Menekan Xanthomonas oryzae pv. Oryzae ini dilaksanakan sejak bulan Agustus 2014.
Terima kasih penulis ucapkan kepada Dr Ir Eny Widajati MS, Dr Ir Wawan Hermawan MS dan Dr Ir Giyanto MSi selaku pembimbing, serta Dr Ir Sugiyanta MSi sebagai penguji luar komisi. Penulis juga menyampaikan terima kasih kepada Direktorat Jenderal Pendidikan Tinggi (DIKTI) atas Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN) yang telah penulis terima selama ini. Terima kasih juga penulis sampaikan kepada Balai Besar Tanaman Padi Sukamandi atas isolat patogen X. oryzae pv. oryzae
patotipe IV yang digunakan pada penelitian ini. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, seluruh keluarga, serta teman-teman atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
xi
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
2 TINJAUAN PUSTAKA 3
Pelet Benih 3
Bakteri Probiotik 4
Xanthomonas oryzae pv. oryzae 5
3 METODE 7
Waktu dan Tempat 7
Sumber Benih 7
Metode Percobaan 7
Percobaan 1. Uji Antagonis Bakteri Probiotik terhadap Patogen X. oryzae pv. oryzae 8 Percobaan 2. Uji Kompatibilitas Bakteri Probiotik 8 Percobaan 3. Uji Daya Simpan Bakteri Probiotik dalam
Formula Pelet 8
Percobaan 4. Uji Formula Pelet pada Benih Padi terinfeksi
X. oryzae pv. oryzae 9
4 HASIL DAN PEMBAHASAN 12
5 SIMPULAN DAN SARAN 21
Simpulan 21
Saran 21
DAFTAR PUSTAKA 22
DAFTAR TABEL
1 Kompatibilitas antar bakteri probiotik ditandai dengan pembentukan zona bening di sekitar kertas saring steril pada media NA 13 2 Pengaruh komposisi formulasi pelet terhadap viabilitas bakteri
probiotik selama 4 minggu simpan 14
3 Pengaruh perlakuan benih terhadap populasi patogen X. oryzae pv.
oryzae pada benih padi 15
4 Pengaruh perlakuan benih terhadap daya berkecambah benih padi
selama 6 minggu penyimpanan 16
5 Pengaruh perlakuan pelet terhadap berat kering kecambah normal 17 6 Pengaruh perlakuan benih terhadap kecepatan tumbuh benih padi
selama 6 minggu penyimpanan 18
7 Pengaruh faktor tunggal periode simpan dan perlakuan terhadap
tolok ukur vigor serta viabilitas benih 18
DAFTAR GAMBAR
1 Diagram alir kegiatan penelitian 7
2 Uji antagonis antar bakteri probiotik terhadap X. oryzae pv.oryzae
1
1
PENDAHULUAN
Latar Belakang
Patogen Xanthomonasoryzae pv. oryzae merupakan patogen utama penyebab penyakit pada padi, mulai dari fase bibit atau umum disebut kresek hingga menjelang panen yang dikenal dengan Hawar Daun Bakteri (HDB). Pada tahun 2009-2013, sekitar 94 246 ha lahan pertanaman padi di Indonesia terserang HDB
dengan 27.6 ha diantaranya puso (Ditlitanpang 2014). Penyebaran patogen
X. oryzae pv. oryzae luas dan cepat terutama karena terbawa melalui benih. Inokulum patogen yang terbawa melalui benih dapat berkembang seiring dengan perkembangan tanaman dan menyebabkan penyakit pada tanaman sehat lainnya melalui udara, gesekan fisik maupun air. Patogen tersebut sulit dikendalikan karena bersifat soilborne (bertahan dalam tanah), seedborne (terbawa benih) dan dapat bertahan pada sisa-sisa tanaman serta memiliki kisaran inang yang luas.
Pengendalian X. oryzae pv. oryzae umumnya melalui penyemprotan tanaman menggunakan bakterisida sintetik. Namun, teknik ini dinilai kurang ramah lingkungan dan dapat menyebabkan resistensi patogen. Oleh sebab itu, pengendalian X. oryzae pv. oryzae secara biologi menggunakan bakteri probiotik mulai dikembangkan khususnya pada tingkat benih. Bakteri probiotik mampu menghasilkan antibiotik dalam jumlah yang cukup untuk menekan perkembangan patogen sehingga meminimalisir terjadinya resistensi patogen (Nawangsih 2006). Pemberian inokulan pada benih sebelum tanam perlu dilakukan untuk memastikan ketersediaan mikroorganisme yang diinginkan dan mengurangi kemungkinan kolonisasi flora tanah indogenous ketika benih berkecambah (Ruiza et al. 2011).
Bakteri probiotik adalah mikroba hidup yang memiliki manfaat bagi inangnya melalui asosiasi dengan inang atau mikroba sekitar, meningkatkan penggunaan makanan atau nilai nutrisi, resistensi penyakit atau dengan memperbaiki kualitas lingkungan (Verschuere et al. 2000). Bakteri kelompok Bacillus, Pseudomonas dan
Streptomyces diketahui mampu menghasilkan antibiotik maupun metabolit sekunder lainnya untuk menekan X. oryzae pv. oryzae seperti difficidin, bacilysin, iturin, siredofor dan hidrogen sianida, serta hormon pertumbuhan seperti indole aceticacid/IAA (Miliute dan Buzaite 2011; Beric et al. 2012; Lukkani dan Reddy 2014; Harikrishnan et al. 2014; Wu et al. 2015). Pemanfaatan Bacillus subtilis
dalam matriconditioning benih padi terinfeksi terbukti mampu menurunkan populasi X. oryzae pv. oryzae serta meningkatkan pertumbuhan bibit padi (Agustiansyah et al. 2010).
Masalah yang dihadapi dalam pemanfaatan bakteri probiotik adalah sulitnya menyediakan bakteri yang viabel dalam jumlah banyak di lapangan. Upaya alternatif yang dapat dilakukan yaitu dengan aplikasi bakteri langsung pada benih dalam bentuk formula kering (pelet). Pelet yang baik mampu melindungi benih dan bakteri dari kondisi ekstrim (sub optimal ruang hidup, nutrisi, tekanan potensial, pH, suhu, predator) ketika di penyimpanan, transportasi dan ditabur ke lapangan sehingga viabilitas benih dan bakteri dapat dipertahankan (van Overbeek et al.
1995; Copeland dan Mc Donald 1995; van Veen et al. 1997). Pelet dapat mempermudah penanaman benih menggunakan mesin tanam, sehingga meminimalisir masuknya patogen X. oryzae pv. oryzae akibat luka pada akar saat pindah tanam.
2
viabilitas bakteri Pseudomonas flourescens RRb-11 hingga 90 hari (30.1 x 107 cfu g-1) serta dapat digunakan sebagai seed treatment yang efektif
mereduksi keparahan penyakit HDB hingga 83.87% dibandingkan kontrol (Jambhulkhar dan Sharma 2014).
Penelitian ini menggunakan bakteri probiotik yang berasal dari dalam jaringan tanaman/endofit (467 dan 748), tanah (aktinomiset 6) dan rhizosfer (Ralstonia pickettii TT47). Aktinomiset 6 diisolasi dari tanah sawah dan diketahui memiliki kemampuan antagonis terhadap X. oryzae pv. oryzae serta dapat diinokulasikan ke benih padi dalam bentuk formula kering (Amalia 2014). Sementara bakteri endofit dan R. pickettii TT47 akan diuji kemampuannya dalam menekan X. oryzae pv. oryzae pada penelitian ini. Pelet diharapkan mampu menekan pathogen pada benih selama di penyimpanan serta melindungi dan membawa bakteri probiotik sedini mungkin ke lapangan sehingga aktinomiset dan
R. pickettii dapat mengkolonisasi tanah serta rhizosfer perakaran dengan cepat, sementara bakteri endofit masuk dan mengkolonisasi jaringan tanaman. Kombinasi bakteri-bakteri ini tentunya dapat memberikan perlindungan lebih optimum kepada tanaman dari luar dan dalam tanaman.
Informasi mengenai kemampuan pelet dalam membawa bakteri probiotik dan pengaruhnya terhadap viabilitas benih padi yang terinfeksi X. oryzae pv. oryzae
juga belum tersedia. Oleh sebab itu, penelitian ini bertujuan untuk mengevaluasi pengaruh pelet dengan penambahan bakteri probiotik dalam menekan patogen
X. oryzae pv. oryzae serta mempertahankan viabilitas benih padi terinfeksi di penyimpanan.
Tujuan
Penelitian ini bertujuan untuk:
1. Menguji bakteri probiotik yang antagonis terhadap patogen X. oryzae pv. oryzae
2. Menguji bakteri probiotik yang saling kompatibel
3
2
TINJAUAN PUSTAKA
Pelet Benih
Pelet benih didefinisikan unit-unit berbentuk bulat yang dibuat untuk meningkatkan presisi tanam, dimana ukuran dan bentuk asli benihnya tidak terlihat lagi (ISTA 2014). Tujuan dari pelet adalah untuk merubah bentuk, berat dan ukuran benih menjadi lebih spesifik sehingga memudahkan penanaman mengunakan mesin tanam (Ilyas 2012). Berat benih yang dipelet umumnya meningkat 50-200% (Gregg dan Billups 2010). Pelet terdiri atas inokulan, bahan pembawa inokulan dan perekat. Penelitian mengenai bahan pembawa dan perekat terbaik untuk benih padi telah dilaporkan oleh Palupi et al. (2012), dimana pelapisan menggunakan talek 1% + CMC 1.5% tidak menghambat perkecambahan dan memiliki daya berkecambah benih (89%) yang tidak berbeda nyata dengan tanpa pelapisan (80.50%). Formula tersebut juga merupakan formula terbaik dibandingkan formula lainnya (alginat,
arabic gum, gypsum dan gambut) untuk semua tolok ukur vigor benih padi yaitu indeks vigor (55.50%), keserempakan tumbuh (79.50%) dan kecepatan tumbuh (16.46%KN etmal-1).
Faktor lingkungan seperti suhu pengeringan, suhu penyimpanan dan kadar air juga mempengaruhi kemampuan formula pelet dalam mempertahankan viabilitas benih dan bakteri. Pada benih Vigna radiata misalnya, pelapisan menggunakan substrat talek ber-pH 7 (1 kg talek + perekat 1 g CMC + 2.5 L air) + 400 mL suspensi bakteri Pseudomonas aeruginosa (3.1 x 109 cfu mL-1) mampu mempertahankan viabilitas bakteri hingga 120 hari penyimpanan (Ali et al. 2001). Pada pH yang sama, P. flourescens UTPF6 dengan formulasi bubuk bakteri (terbuat dari suspensi bakteri + Na alginat + talek) yang dikemas dalam botol kaca dan disimpan pada suhu 4 0C dengan kandungan air 15%, mampu mempertahankan viabilitas bakteri hingga 90 hari penyimpanan (Sadi dan Masoud 2012). Dengan pH dan suhu yang sama, penyimpanan P. flourescens pada kantong polyethylene dengan kandungan air 35% menggunakan media talek dan wood flour mampu mempertahankan viabilitas bakteri hingga 5 bulan penyimpanan (2 x 108 cfu mL-1 di awal penyimpanan dan 5 x 105cfu mL-1 di bulan ke 5 penyimpanan) (Sallam et al. 2013).
Kloepper dan Schroth (1981) melaporkan formulasi pelet menggunakan campuran talek dengan 20% xanthan gum mampu mempertahankan populasi PGPR tanpa mengalami decline pada benih kentang. Formula tersebut dapat mempertahankan populasi PGPR sebesar 8.2 x 107 cfu mL-1 selama dua bulan penyimpanan pada 4 0C. Adapun formulasi tersebut: 5 mL XG + 5 mL PGPR (109
cfu mL-1) diaduk selama 20 menit, kemudian ditambah talek (< 5x formula awal), lalu disimpan selama 3-4 hari (sampai kering) pada 12 0C, 0.5 kg formula (dust) untuk 46 kg benih. Suslow dan Schroth (1982) menggunakan cellulose methyl ether
4
Vidhyasekaran dan Muthamilan (1995) melaporkan formulasi: 10 g CMC + 1 kg talek/vermikulit diaduk dan diukur pH, apabila pH < 7 maka ditambahkan kalsium karbonat hingga pH larutan menjadi 7. Larutan lalu diautoklaf. Setelah dingin, ditambahkan 400 mL bakteri (9 x 108 cfu mL-1) + 4 g kg-1 benih, lalu dikeringanginkan selama 2 jam. Formulasi yang diaplikasikan pada benih Chickpea
ini mampu mempertahankan koloni P. flourescens (107 cfu g-1) selama 240 hari penyimpanan.
Bakteri Probiotik
Bakteri probiotik merupakan mikroba hidup yang memiliki manfaat bagi inangnya melalui asosiasi dengan inang atau mikroba sekitar, meningkatkan penggunaan makanan atau nilai nutrisi, resistensi penyakit atau dengan memperbaiki kualitas lingkungan (Verschuere et al. 2000). Bakteri ini mampu menghasilkan antibiotik dalam jumlah yang cukup untuk menekan perkembangan patogen sehingga meminimalisir terjadinya resistensi patogen (Nawangsih 2006). Bakteri dapat dibagi menjadi tiga kelompok berdasarkan lingkungan hidupnya, yaitu 1) bakteri yang hidup bebas. Bakteri ini berinteraksi dengan tanaman apabila kondisi menguntungkan; 2) rhizosfer dan filosfer, bakteri terlokalisasi pada zona tertentu seperti tanah hingga perakaran atau di permukaan epidermis daun tanaman; 3) bakteri endofit, bakteri yang mampu mengkolonisasi jaringan dan organ tanaman dengan penetrasi melalui celah interseluler (Maksimov et al. 2011).
Bakteri endofit merupakan bakteri saprofit yang hidup dan berasosiasi dengan jaringan tanaman yang sehat tanpa menimbulkan gejala penyakit (Backman dan Sikora 2008). Mikroorganisme endofit memberikan manfaat ke tanaman inang dengan meningkatkan aktivitas fisiologi dari tanaman atau aktivitas lainnya dan mungkin menyediakan sumber dari komponen aktif, agen biokontrol atau promotor pertumbuhan tanaman (Hastuti et al. 2012). Penelitian Mattos et al. (2008) menggunakan Burkholderia kururiensis strain KP23T (108 sel mL-1) yang diinokulasikan pada planlet padi varietas Guarani menunjukkan adanya perbedaan nyata pada aktivitas auxin-inducible reporter DR5-GUS planlet yang diberi perlakuan inokulasi bakteri dan tidak, pada 24 jam setelah inokulasi. Hal ini mungkin diakibatkan ekspresi DR5-GUS yang berhubungan dengan kemampuan
B. kururiensis memproduksi auksin dan sekresi dalam jaringan tanaman. Penelitian ini juga menunjukkan dugaan mekanisme bakteri endofit masuk ke dalam tanaman. Tahap awal yaitu masuknya bakteri ke sel-sel epidermis dari permukaan akar, dimana rambut akar merupakan zona kolonisasi terbesar munculnya bakteri. Tahap kedua ditandai oleh proliferasi bakteri melalui jaringan basal rambut akar, di mana kerusakan dangkal yang mudah terdeteksi menunjukkan sebuah proses aktif mungkin diakibatkan oleh enzim pendegradasi dinding sel, yang memungkinkani infeksi kortikal akibat dispersi apoplastic oleh bakteri. Tahap ketiga adalah invasi interseluler jaringan internal oleh bakteri yang kemudian masuk ke dalam sistem vaskular akar, di mana bakteri mungkin akan ditranslokasikan ke batang bawah di xilem. Lokalisasi utama B. kururiensis dalam pembuluh xilem menunjukkan bahwa bakteri menyebar sistematis ke tunas melalui jaringan pembuluh.
5 nitrogen menghasilkan panjang dan berat kering yang lebih tinggi dibandingkan tanpa penambahan nitrogen.
Aktinomiset termasuk bakteri Gram positif berfilamen, bersifat saprofitik, mampu menjelajah jaringan tanaman dan menghasilkan spora bertahan yang dapat bertahan lama di tanah. Bakteri ini dapat memproduksi metabolit antibiotik dan senyawa antimikroba sehingga dapat membatasi serangan organisme patogen (Patil
et al. 2010). Pemanfaatan bakteri ini sebagai bio-protektan dilaporkan oleh Zarandi
et al. (2009), padi (varietas Kazemi) yang diinokulasi (spray) dengan Magnaporthe oryzae (penyebab penyakit rice blast) 4 x 105 konidia mL-1 saat fase tiga daun menghasilkan gejala spesifik blast pada daun sebesar 8%. Sementara tanaman yang diberi perlakuan M. oryzae + Streptomyces sindeneusis isolat 263 pada konsentrasi 3.125 mg mL-1 hanya sebesar 0.5%.
R. pickettii TT47 merupakan salah satu bakteri probiotik yang koloninya berukuran 1.5 mm, berbentuk sirkular dengan pinggiran rata, agak cembung, berwarna kuning dengan permukaan yang licin. Bakteri Gram negatif ini memiliki sel berbentuk batang berukuran 1.0 x 4.0 μm, tidak memiliki endospora serta mampu menghasilkan siderofor, enzim kitinase dan melarutkan fosfat (Rustam 2012). Rustam (2012) melaporkan penggunaan R. pickettii TT47 sebagai agen biokontrol penyakit hawar pelepah yang disebabkan oleh cendawan Rhizoctonia solani dan mampu menekan penyakit hingga 79.6%. Bakteri ini diduga mampu mendegradasi dinding sel cendawan. Park et al. (2002) melaporkan penggunaan R. pickettii PKO1 sebagai bioremidioator yaitu untuk mendegradasi Trichloroethylene
(TCE).
Peranan bakteri probiotik dalam memacu pertumbuhan tanaman mungkin terkait dengan substansi yang dihasilkan atau kemampuan bakteri tersebut. Mbai et al. (2013) menunjukkan isolat M31 (bakteri endofit CO3) dan M32 (Pseudomonas fluorescens Mc07/d3) yang diisolasi dari benih padi Mwea Basmati 370, kemudian diaplikasikan pada tanah dengan konsentrasi 1 × 105 lebih baik dibandingkan 1 × 1010. Isolat tersebut juga secara signifikan lebih tinggi dibandingkan kontrol negatif pada tolok ukur tinggi tanaman (42.025, 34.275, 24.525 cm) dan berat kering tanaman (1.567, 1.4967, 0.363 g). Kedua isolat tersebut sama-sama Gram negatif, mampu melarutkan fosfat, fiksasi nitrogen dan memproduksi auksin. Kannan et al.
(2014) mengaplikasikan isolat bakteri endofit CSR-M-16 (108 cfu (OD595 = 0.3)
yang diisolasi dari mangga (Mangifera indica) aksesi ML-2 dan GPL-3 yang ditumbuhkan pada tanah salin pada benih padi yang ditanam dalam kondisi salin juga (pH 9.35 dan EC 4.2 dS/m). Isolat CSR-M-16 menunjukkan kemampuan yang signifikan dalam meningkatkan perkecambahan benih 93.33% dibanding kontrol 6.67%, vigor benih (4675.83 : 80.22), panjang akar (13.6 : 3.7 cm), panjang tunas (36.5 : 8.31 cm), berat kering akar (0.38 : 0.14 g) dan berat kering tunas (1.75 : 0.12 g). Isolat tersebut juga menunjukkan kemampuan menghasilkan IAA tertinggi (74
6
Xanthomonas oryzae pv. oryzae
X. oryzae pv. oryzae merupakan bakteri Gram negatif, tidak membentuk spora, aerobik, sel berbentuk batang lurus dengan panjang 0.7-2.9 μm dan lebar 0.4-0.7 μm, bergerak menggunakan satu bulu cambuk polar. Koloni bakteri X. oryzae
pv. oryzae yang tumbuh pada media agar biasanya berwarna kuning, mengkilap, bulat, cembung, mukoid, dan sebagian besar tumbuh dengan lambat. Warna kuning dikarenakan bakteri memproduksi pigmen xanthomonadin (Nino-Liu et al. 2006).
X. oryzae pv. oryzae adalah patogen vaskular yang dapat langsung masuk ke pembuluh melalui luka yang dihasilkan selama tanam atau dengan hujan angin.
X. oryzae pv. oryzae dapat terbawa cairan gutasi dan masuk melalui hidatoda (pori air). Setelah X. oryzae pv. oryzae memasuki epithem (ruang bawah pori air), bakteri berkembang biak lalu bergerak melalui pembuluh vaskular dan masuk ke dalam pembuluh xilem. X. oryzae pv. oryzae kemudian berkembang biak kembali dan menyebar ke seluruh sistem vaskular (Leach dan Corral 2013). Pada benih padi, X. oryzae pv. oryzae dapat dengan mudah menginfeksi glume, namun gejala baru tampak pada cabang panikel. Pada kondisi penyimpanan alami (temperatur 25 - 35 0C) bakteri dapat terdeteksi hingga 2 bulan (Kauffman dan Reddy 1975).
Patogen X. oryzae pv. oryzae dapat menyerang pertanaman padi mulai dari fase bibit (kresek) hingga menjelang panen (hawar) dengan cara mengaktifkan transkripsi gen Xa-13. Gen tersebut kemudian mengkode protein di membran plasma. Protein Xa-13 bekerjasama dengan protein COPT1 dan COPT5 untuk menghilangkan copper dari pembuluh xilem, kemudian X. oryzae pv. oryzae
berduplikasi dan menyebar menyebabkan penyakit (Yuan et al. 2010).
Serangan tertinggi patogen X. oryzae pv. oryzae cenderung terjadi di daerah Jawa Barat terutama pada bulan Februari (Ditlitanpang 2014). Selain itu, di Inonesia, varietas padi yang ditanam umumnya rentan terhadap X. oryzae pv. oryzae, seperti Ciherang yang hanya memiliki ketahanan terhadap patotipe III dan rentan (keparahan penyakit 25-50%) terhadap patotipe IV dan VIII (Susanto dan Sudir 2012). Penurunan hasil hingga 70% dapat terjadi ketika varietas rentan ditanam pada kondisi lingkungan yang menguntungkan patogen X. oryzae pv. oryzae (IRRI 2014). Fase kresek merupakan fase terparah karena dapat menyebabkan kematian tanaman akibat keringnya daun tanaman (Jambhulkar dan Sharma 2014). Patogen
7
3
METODE
Waktu dan Tempat
Percobaan dilaksanakan pada Agustus 2014 hingga Oktober 2015 di Laboratorium Fisiologi dan Kesehatan Benih, Departemen Agronomi dan Hortikultura serta Laboratorium Bakteriologi Tumbuhan, Departemen Proteksi Tanaman, Institut Pertanian Bogor.
Sumber Benih
Benih yang digunakan adalah benih padi komersial varietas Ciherang yang diproduksi oleh PT. Sang Hyang Seri, Indonesia, telah disimpan selama dua tahun (suhu 160 C) dengan kadar air 13% dan daya berkecambah 82%.
Metode Percobaan
Percobaan terdiri atas empat bagian: 1) uji antagonis isolat bakteri probiotik terhadap patogen X. oryzae pv. oryzae; 2) uji kompatibilitas antar bakteri probiotik; 3) uji daya simpan bakteri probiotik dalam formula pelet; 4) pengaruh perlakuan pelet dengan penambahan bakteri probiotik pada benih padi. Adapun diagram alir kegiatan penelitian ditunjukkan pada Gambar 1.
8
Pembuatan Suspensi Bakteri Probiotik dan X. oryzae pv. oryzae
Isolat bakteri probiotik maupun patogen dikulturkan kembali pada Nutrient Agar/NA (endofit 467, endofit 748, R. pickettii TT47), water yeast extract/WYE (aktinomiset 6) atau Yeast Dekstrose Carbon Agar/YDCA (X. oryzae pv. oryzae). Setelah berumur 48 jam, satu ose koloni tunggal endofit, R. pickettii atau X. oryzae
pv. oryzae yang muncul diinokulasikan ke dalam 10 mL media cair Nutrient Broth. Inkubasi dilakukan dalam shaker (100 rpm) selama 48 jam.
Koloni tunggal aktinomiset 6 yang diperoleh pada kultur media padat WYE, dimasukkan ke dalam air destilata, lalu divortex. Suspensi tersebut digunakan untuk menginokulasi beras yang telah dicuci dan disterilisasi (autoklaf). Setelah inkubasi selama 7 hari pada suhu ruang, 1 g beras yang telah terselimuti spora aktinomiset 6 dimasukkan ke dalam air destilata hingga mencapai volume 10 mL kemudian divortex sehingga bakteri dan spora yang menempel pada permukaan beras dapat lepas (suspensi aktinomiset siap digunakan) (Amalia 2014).
Percobaan 1. Uji Antagonis Isolat Bakteri Probiotik terhadap Patogen X. oryzae pv. oryzae
Kemampuan bakteri probiotik dalam menghasilkan zat metabolit yang mampu menghambat X. oryzae pv. oryzae diuji pada percobaan ini. Suspensi X. oryzae pv. oryzae (107-108cfu mL-1) sebanyak 100 μL diambil menggunakan pipet mikro kemudian disebar dengan segitiga penyebar di atas media agar YDCA. Kertas saring steril berdiameter 0.5 cm diletakkan di atas media padat YDCA. Suspensi bakteri probiotik (108-109 cfu mL-1) sebanyak 10 μL, diteteskan di atas kertas saring tersebut (Amalia 2014). Inkubasi 3-5 hari pada suhu ruang. Pengamatan terhadap ada tidaknya zona bening di sekitar kertas saring dilakukan setiap hari. Zona bening di sekitar kertas saring menunjukkan bakteri probiotik menghasilkan metabolit yang menghambat perkembangan X. oryzae pv. oryzae. Percobaan dilakukan duplo.
Percobaan 2. Uji Kompatibilitas Bakteri Probiotik
Aplikasi beberapa bakteri dalam satu formula hanya bisa dilakukan jika bakteri-bakteri tersebut tidak saling menghambat perkembangan satu dengan yang lain (kompatibel). Kompatibilitas antar bakteri probiotik dapat diuji menggunakan metode biakan ganda (Putra dan Giyanto 2014)melalui ada tidaknya zona bening yang terbentuk di sekitar kertas saring. Suspensi bakteri probiotik 1 (100 μL, 108-109 cfu mL-1) disebar di atas media padat NA, kemudian kertas saring steril berdiameter 0.5 cm diletakkan di atas media tersebut. Kertas saring steril tersebut lalu ditetesi 10 μL (108-109 cfu mL-1) suspensi bakteri probiotik 2. Apabila terbentuk zona bening di sekitar kertas saring steril, berarti bakteri 2 menghambat perkembangan bakteri 1 (tidak kompatibel). Untuk mengetahui apakah bakteri 1 antagonis terhadap bakteri 2, dilakukan metode yang sama. Bakteri yang menunjukkan sifat kompatibel kemudian diaplikasikan secara bersama pada percobaan 3. Percobaan diulang sebanyak dua kali.
Percobaan 3. Uji Daya Simpan Bakteri Probiotik dalam Formula Pelet
9 yang digunakan yaitu talek, talek + dekstrose (2%) dan talek + gliserol (1%). Suspensi bakteri (108-109 cfu mL-1) sebanyak 2 mL dibutuhkan untuk menginokulasi 5 g talek. Pengamatan terhadap populasi bakteri dalam satu gram bahan pembawa dilakukan setiap minggunya selama 4 minggu. Formula yang mampu mempertahankan viabilitas bakteri paling stabil digunakan dalam percobaan 4. Populasi bakteri dihitung dengan metode pengenceran berseri.
Pengenceran Berseri
Satu gram sampel dimasukkan dalam tabung lalu ditambahkan air steril hingga 10 mL (pengenceran 100). Setelah divortex, sebanyak 1 mL suspensi bakteri yang sudah homogen tersebut diambil menggunakan mikro pipet, lalu dimasukkan ke dalam tabung yang telah berisi 9 mL air steril (10-1). Pengenceran terus dilakukan hingga mencapai 10-7. Suspensi dari tabung pengenceran ke-5 disebar pada media padat (NA/WYE/YDCA) sebanyak 0.1 mL, begitu juga dengan tabung pengenceran ke-6 dan 7. Penyebaran suspensi pada media padat dari masing-masing tabung dilakukan duplo. Inkubasi dilakukan selama 24-48 jam pada suhu ruang. Koloni-koloni yang terbentuk kemudian dihitung.
Rancangan Percobaan
Percobaan 3. dilaksanakan menggunakan Rancangan Acak Kelompok (RAK) petak tersarang. Petak utama adalah periode simpan dengan anak petak formula pelet. Periode simpan terdiri atas lima taraf (0, 1, 2, 3, 4 minggu), sementara anak petak terdiri atas tiga taraf (talek, talek + dekstrose 2%, talek + gliserol 1%). Masing-masing perlakuan diulang sebanyak dua kali, sehingga diperoleh 30 satuan percobaan. Berikut merupakan model linier rancangan yang digunakan dalam percobaan ini:
Yijk = μ + τi+ (ατ)ik+ βj+ (τβ)ij + αk + εijk
Dimana:
Yijk = nilai pengamatan pada penyimpanan ke-i, formula pelet taraf ke- j, ulangan ke-k
μ = nilai rataan umum
τi = pengaruh utama periode simpan
(ατ)ij = kelompok tersarang dalam periode simpan
Βj = pengaruh fomula pelet ke- j
(τβ)ik = pengaruh interaksi periode simpan ke-i, formula pelet ke-j dan ulangan ke-k
αk = pengaruh kelompok ke-k
εijk = pengaruh acak pada periode simpan ke-I, formula pelet ke-j dan ulangan ke-k.
Percobaan 4. Uji Formula Pelet pada Benih Padi Terinfeksi
X. oryzae pv. oryzae
Benih padi diinfeksi patogen X. oryzae pv. oryzae terlebih dahulu sebelum diberi perlakuan pelet. Infeksi dilakukan dengan merendam benih selama 24 jam dalam suspensi X. oryzae pv. oryzae berumur 48 jam dimana kerapatan bakteri
10
plastik poliprofilen untuk disimpan pada suhu ruang selama 6 minggu. Bobot benih yang dipelet meningkat 10 kali lipat atau berkisar 900%.
Rancangan Percobaan
Percobaan ini juga menggunakan rancangan acak kelompok (RAK) petak tersarang. Petak utama adalah periode simpan (0, 2, 4, 6 minggu). Anak petak yaitu perlakuan pelet (8 taraf): (1) Kontrol (benih sehat), (2) X (X. oryzae pv. oryzae), (3)
benih direndam dalam air selama 24 jam pada suhu ruang, (4) K + pelet, (5) X + pelet + aktinomiset 6 + endofit 467, (6) X + pelet + aktinomiset 6, (7) X +
pelet + endofit 467, (8) X + pelet + R. pickettii TT47. Perlakuan diulang sebanyak tiga kali sehingga diperoleh 96 satuan percobaan.
Pengamatan dilakukan terhadap populasi X. oryzae pv. oryzae pada benih padi terinfeksi menggunakan metode pengenceran berseri. Viabilitas benih terbagi menjadi dua yaitu viabilitas potensial dan vigor. Tolok ukur viabilitas potensial yaitu daya berkecambah (DB), berat kering kecambah normal (BKKN)dan potensi tumbuh maksimum (PTM). Tolok ukur vigor meliputi: indeks vigor (IV) dan kecepatan tumbuh (KCT). Penghitungan kadar air benih dilakukan tiap periode
simpan. Adapun rumus dari kadar air dan masing-masing tolok ukur viabilitas benih adalah sebagai berikut:
Kadar Air (KA)
Benih padi dari masing-masing ulangan dalam satu perlakuan digabung menjadi satu, dipisahkan benih dari peletnya. Benih lalu dihaluskan menggunakan blender hingga menjadi serbuk. Serbuk padi disaring kembali menggunakan ayakan hingga diperoleh butiran yang lebih halus. Sekitar 1-1.5 g benih yang telah dihaluskan dimasukkan ke dalam cawan untuk diukur kadar airnya menggunakan oven suhu 105 0C selama ±17 jam. KAdihitung dengan menggunakan rumus:
KA = M2 - M3 × 100%
Daya berkecambahnmenggambarkan viabilitas potensial, dimana hasil diperoleh dari persentase kecambah normal (KN) pada pengamatan 1 (hari ke-5) dan
pengamatan 2 (hari ke-14). Rumus yang digunakan:
DB = Σ KN hitungan I + Σ KN hitungan II × 100% mengindikasikan viabilitas potensial (Vp) dengan menggambarkan laju
11 Potensi Tumbuh Maksimum (PTM)
Potensi tumbuh maksimum (PTM) mengindikasikan viabilitas total. Penghitungan PTMdidasarkan pada benih yang tumbuh (berkecambah) sampai hari ke-14 setelah tanam. Rumus untuk menghitung PTM adalah:
PTM= Σ KNΣ B+ Σ KAb × 100%
T
Dimana:
ΣKN = jumlah kecambah normal sampai akhir pengamatan
ΣKAb = jumlah kecambah abnormal sampai akhir pengamatan
Σ BT = jumlah benih yang ditanam
Kecepatan Tumbuh (KCT)
Kecepatan tumbuh (KCT) merupakan tolok ukur yang mengindikasikan
vigor kekuatan tumbuh. Perhitungan KCT didasarkan pada akumulasi KCT harian
dalam unit tolok ukur persentase per hari dengan rumus:
KCT = %Ketmal N ke-2 +…..+ %Ketmal N ke-n
Dimana:
1 etmal = 24 jam
%KN = persentase kecambah normal
Indeks Vigor (IV)
Perhitungan didasari pada persentase kecambah normal (KN) di hitungan
pertama untuk padi yaitu hari ke-5 pada uji daya berkecambah, dengan rumus: IV = Σ KN hitungan I X 100%
Σ BT
Dimana:
ΣKN = persentase kecambah normal
Σ BT = jumlah benih yang ditanam
Populasi X. oryzae pv. oryzae
Populasi X. oryzae pv. oryzae tiap periode simpan dihitung dengan pengenceran berseri. Benih padi dilepas dari peletnya terlebih dahulu. Sebanyak 1 g benih dari masing-masing perlakuan, disterilisasi permukaannya menggunakan klorox 1%, kemudian dibilas menggunakan air steril tiga kali. Benih lalu dihaluskan menggunakan mortar, kemudian dimasukkan ke dalam tabung, lalu ditambahkan air steril hingga volume mencapai 10 mL (pengenceran ke 0), selanjutnya, tabung-tabung diinkubasi ke dalam suhu 15 0C selama dua jam, kemudian tabung di-shaker
sekitar 30 menit. Baru dilakukan pengenceran berseri.
Analisis Data
12
4
HASIL DAN PEMBAHASAN
Aktivitas Antagonis Isolat Bakteri Probiotik terhadap Patogen X. oryzae pv. oryzae
Aktivitas antagonisme isolat bakteri probiotik yang digunakan (endofit 467, endofit 748, aktinomiset 6 dan R. pickettii TT47) dapat diketahui menggunakan metode biakan ganda, yaitu dengan ada atau tidaknya zona bening yang terbentuk di sekitar kertas saring. Hasil percobaan menunjukkan R. pickettii TT47 menghasilkan zona bening di sekitar kertas saring yang terbesar, diikuti oleh aktinomiset 6 dan endofit 467 (Gambar 2). Zona bening dihasilkan ketika bakteri yang berada di kertas saring menghasilkan senyawa yang bersifat menekan/antagonis terhadap perkembangan X. oryzae pv. oryzae yang berada di media padat YDCA. Isolat endofit 748 tidak menunjukkan kemampuan menghasilkan senyawa antagonis karena tidak terbentuk zona bening disekitar kertas saring, namun koloni bakteri tersebut dapat dengan cepat mengelilingi kertas saring. Apabila isolat endofit 748 dapat kompatibel dengan isolat bakteri probiotik lain yang digunakan pada penelitian ini, diharapkan dapat membantu memperluas spektrum penekanan patogen X. oryzae pv. oryzae melalui kompetisi ruang dan nutrisi.
Gambar 2 Uji antagonis antar bakteri probiotik terhadap X. oryzae pv.oryzae
(koloni berwana kuning pekat) ditandai dengan pembentukan zona bening di sekitar kertas saring pada media YDCA
Isolat bakteri digolongkan antagonis apabila mampu menunjukkan aktivitas antagonis terhadap patogen. Aktivitas antagonisme tersebut dapat digolongkan menjadi 3 yaitu: (a) langsung melalui Hyperparasitism/predasi, (b) Mixed-path antagonism : bakteri dapat memproduksi senyawa yang bersifat antagonis terhadap patogen seperti antibiotik, enzim lisis maupun metabolit sekunder, (c) tidak langsung : bakteri mampu berkompetisi dengan patogen dalam kolonisasi tempat hidup/ruang maupun dalam penyerapan nutrisi; beberapa jenis bakteri dapat menginduksi ketahanan sistemik tanaman sehingga tidak mudah terserang penyakit (Pal dan Gardener 2006). Interaksi antagonis antara bakteri Gram negatif dan Gram positif dipengaruhi oleh produksi toksin yang disebut bakteriosin (Dardick et al. 2003).
13 mengandung rantai samping dan grup fungsional, serta mampu menyediakan afinitas tinggi pada ligan yang berkoordinasi dengan ion besi (Crosa dan Walsh 2002). Siderofor disekresikan untuk melarutkan zat besi, membentuk ferric-siderophore komplek yang dapat bergerak dengan difusi dan dikembalikan ke permukaan sel (Andrews et al. 2003). Mikroorganisme dalam kondisi aerob membutuhkan zat besi untuk berbagai siklus dalam sel mulai dari energi untuk regenerasi, fotosintesis, respirasi, termasuk pengurangan oksigen untuk sintesis ATP, pengurangan prekursor ribotida DNA, untuk pembentukan heme, dan proteksi dari stress oksidatif (Neilands 1995, Andrews et al. 2003, Skaar 2010).
Sifat Kompatibilitas Antar Bakteri Probiotik
Uji kompatibilitas dilakukan agar antar bakteri probiotik yang akan diaplikasikan bersama tidak saling menghambat sehingga menurunkan kemampuan antagonis mereka terhadap patogen (Mishra et al. 2013). Kompatibilitas antar isolat bakteri probiotik yang digunakan dapat diketahui menggunakan metode biakan ganda yaitu ada tidaknya zona bening yang terbentuk di sekitar kertas saring. Zona bening yang terbentuk di sekitar kertas saring menunjukkan bakteri probiotik yang berada di kertas saring menghasilkan senyawa antagonis yang menekan perkembangan bakteri probiotik di media agar (tidak kompatibel). Tabel 1 menunjukkan bahwa aktinomiset 6 dan endofit 467 bersifat kompatibel, sehingga
dapat diaplikasikan secara bersama. Isolat endofit 748 tidak antagonis terhadap
X. oryzae pv. oryzae dan tidak kompatibel dengan tiga isolat bakteri probiotik lain yang diuji sehingga tidak digunakan kembali dalam penelitian berikutnya.
Tabel 1 Kompatibilitas antar bakteri probiotik ditandai dengan pembentukan zona bening di sekitar kertas saring steril pada media NA
Keterangan: + terbentuk zona bening; - tidak terbentuk zona bening; x tidak diuji kompatibilitas.
Daya Simpan Bakteri Probiotik dalam Formula Pelet
14
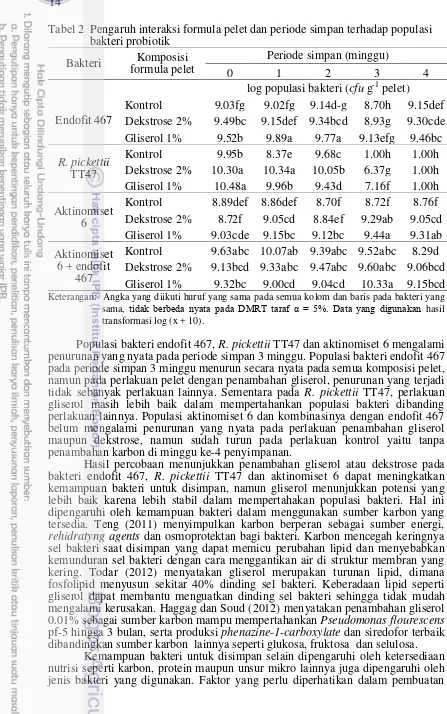
Tabel 2 Pengaruh interaksi formula pelet dan periode simpan terhadap populasi bakteri probiotik Dekstrose 2% 9.13bcd 9.33abc 9.47abc 9.60abc 9.06bcd Gliserol 1% 9.32bc 9.00cd 9.04cd 10.33a 9.15bcd
Keterangan: Angka yang diikuti huruf yang sama pada semua kolom dan baris pada bakteri yang sama, tidak berbeda nyata pada DMRT taraf α = 5%. Data yang digunakan hasil transformasi log (x + 10).
Populasi bakteri endofit 467, R. pickettii TT47 dan aktinomiset 6 mengalami penurunan yang nyata pada periode simpan 3 minggu. Populasi bakteri endofit 467 pada periode simpan 3 minggu menurun secara nyata pada semua komposisi pelet, namun pada perlakuan pelet dengan penambahan gliserol, penurunan yang terjadi tidak sebanyak perlakuan lainnya. Sementara pada R. pickettii TT47, perlakuan gliserol masih lebih baik dalam mempertahankan populasi bakteri dibanding perlakuan lainnya. Populasi aktinomiset 6 dan kombinasinya dengan endofit 467 belum mengalami penurunan yang nyata pada perlakuan penambahan gliserol maupun dekstrose, namun sudah turun pada perlakuan kontrol yaitu tanpa penambahan karbon di minggu ke-4 penyimpanan.
Hasil percobaan menunjukkan penambahan gliserol atau dekstrose pada bakteri endofit 467, R. pickettii TT47 dan aktinomiset 6 dapat meningkatkan kemampuan bakteri untuk disimpan, namun gliserol menunjukkan potensi yang lebih baik karena lebih stabil dalam mempertahakan populasi bakteri. Hal ini dipengaruhi oleh kemampuan bakteri dalam menggunakan sumber karbon yang tersedia. Teng (2011) menyimpulkan karbon berperan sebagai sumber energi,
rehidratyng agents dan osmoprotektan bagi bakteri. Karbon mencegah keringnya sel bakteri saat disimpan yang dapat memicu perubahan lipid dan menyebabkan kemunduran sel bakteri dengan cara menggantikan air di struktur membran yang kering. Todar (2012) menyatakan gliserol merupakan turunan lipid, dimana fosfolipid menyusun sekitar 40% dinding sel bakteri. Keberadaan lipid seperti gliserol dapat membantu menguatkan dinding sel bakteri sehingga tidak mudah mengalami kerusakan. Haggag dan Soud (2012) menyatakan penambahan gliserol 0.01% sebagai sumber karbon mampu mempertahankan Pseudomonas flourescens
pf-5 hingga 3 bulan, serta produksi phenazine-1-carboxylate dan siredofor terbaik dibandingkan sumber karbon lainnya seperti glukosa, fruktosa dan selulosa.
15 pelet dengan penambahan inokulan bakteri menurut Jansen et al. (1994) serta Elzein et al. (2008) yaitu kemampuan pelet dalam membawa inokulan, kompatibilitasnya dengan inokulan, ukuran granul, tipe dan bentuk inokulum serta kandungan air. Isolat R. pickettii TT47 menunjukkan kemampuan terbaik dalam menekan perkembangan X. oryzae pv. oryzae secara in vitro namun bakteri ini kurang dapat bertahan dalam formula pelet yang kering. Populasi R. pickettii dalam formula talek kering hanya dapat dipertahankan selama 3 minggu. Hal ini dikarenakan R. pickettii TT47 merupakan bakteri Gram negatif yang tidak dapat membentuk spora, tidak seperti endofit 467 dan aktinomiset 6 yang termasuk bakteri Gram positif serta dapat membentuk spora. Spora merupakan struktur bertahan bakteri terhadap kondisi lingkungan yang kurang menguntungkan, misalnya penyimpanan dengan kadar air yang rendah. Kadar air maksimum yang aman untuk peyimpanan benih padi adalah 13% (BSN 2003). Pada penelitian ini kadar air benih yang dipelet maupun tidak dipelet berkisar antara 10.3 - 10.5%.
Pengaruh Formula Pelet pada Benih Padi Terinfeksi X. oryzae pv. oryzae
Berdasarkan hasil percobaan sebelumnya, komposisi formula pelet yang digunakan pada percobaan ini adalah talek + CMC 1.5% + gliserol 1% + bakteri probiotik (108-109cfu mL-1). Perlakuan pelet menggunakan bakteri probiotik pada benih padi terinfeksi secara nyata menurunkan populasi X. oryzae pv. oryzae dalam benih terutama pada penyimpanan minggu ke-6 dibandingkan dengan benih terinfeksi yang tidak diberi perlakuan pelet (Tabel 3). Pada awal penyimpanan, penekanan terbaik ditunjukkan oleh perlakuan pelet menggunakan R. pickettii TT47 dan endofit467. Populasi patogen X. oryzae pv. oryzae pada benih terinfeksi yang diberi perlakuan pelet dengan bakteri tersebut nyata lebih rendah dibandingkan benih terinfeksi yang tidak diberi perlakuan pelet (X). Pada minggu ke 6, pelet menggunakan aktinomiset 6 dan kombinasi aktinomiset 6 + endofit 467 menunjukkan kemampuan penekanan X. oryzae pv. oryzae terbaik, ditunjukkan dari populasi patogen X. oryzae pv. oryzae yang nyata lebih rendah dibandingkan perlakuan lainnya. Benih terinfeksi yang tidak diberi perlakuan pelet terus mengalami peningkatan populasi patogen selama penyimpanan.
Tabel 3 Pengaruh interaksi perlakuan benih dan periode simpan terhadap populasi X. oryzae pv. oryzae pada benih padi
Keterangan: Angka yang diikuti huruf yang sama pada semua kolom dan baris tidak berbeda nyata pada DMRT taraf α = 5%; Data yang digunakan adalah hasil transformasi log (x + 10).
Formula pelet (talek + CMC 1.5% + gliserol 1%) terbukti dapat
mempertahankan kemampuan antagonis bakteri probiotik terhadap patogen
16
bottromycin A2 dan dunaimycin D3S yang dihasilkan Streptomyces bottropensis
(Park et al. 2011); 2.4-diacetylphloroglucinol (DAPG) oleh Pseudomonas fluorescens PDY7 (Velusamy et al. 2013); kitinase, fosfatase dan siredofor dari
Streptomyces sp. (AB131-1 dan LBR02) (Hastuti et al. 2012); produksi siderofor, enzim fosfatase, enzim peroksidase dan hidrogen sianida oleh P. diminuta A6 (Agustiansyah et al. 2013). Antibiotik merupakan molekul bermasa rendah (<1500kDa) hasil dari metabolisme sekunder, umumnya diproduksi selama
idiophase (Sanchez et al. 2010). Antibiotik dapat menghambat sintesis dinding sel, protein maupun asam nukleat patogen serta mengacaukan metabolisme dan membran sel patogen (Dzidic et al. 2008).
Perlakuan pelet benih menggunakan bakteri probiotik aktinomiset 6,
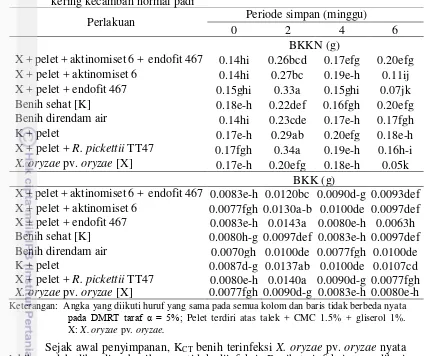
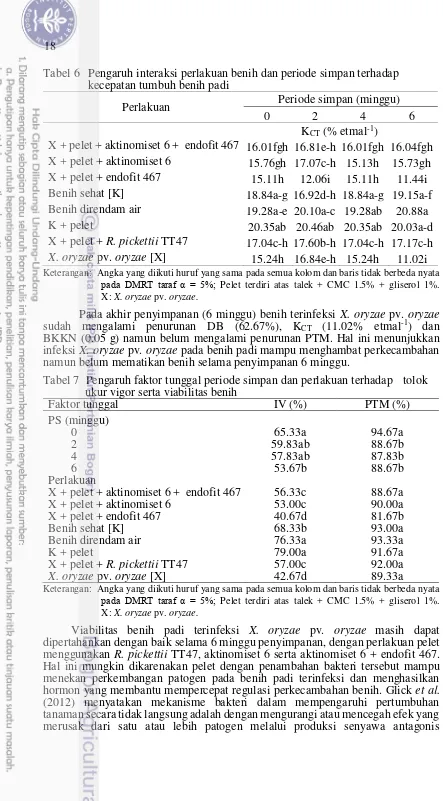
R. pickettii TT47, serta endofit 467 + aktinomiset 6 juga mampu mempertahankan viabilitas benih padi terinfeksi X. oryzae pv. oryzae lebih baik dibanding tanpa perlakuan pelet, berdasarkan tolok ukur daya berkecambah (DB) (Tabel 4), berat kering kecambah normal (BKKN) (Tabel 5) dan kecepatan tumbuh (KCT) (Tabel 6).
Hasil ini sejalan dengan penelitian sebelumnya yang dilakukan oleh Putra dan Giyanto (2014), dimana aplikasi Bacillus galur B12 dan aktinomiset pada benih padi dapat menekan populasi X. oryzae pv. oryzae pada bibit padi serta meningkatkan pertumbuhan bibit.
Tabel 4 Pengaruh interaksi perlakuan benih dan periode simpan terhadap daya berkecambah benih padi Benih direndam air 89.33ab 85.33abc 86.67abc 94.67a
K + pelet 93.33a 85.33abc 89.33ab 90.67a
X + pelet + R. pickettii TT47 90.67a 84.00abc 89.33ab 86.67abc
X. oryzae pv. oryzae [X] 89.33ab 86.67abc 80.00abc 62.67e
Keterangan: Angka yang diikuti huruf yang sama pada semua kolom dan baris tidak berbeda nyata
pada DMRT taraf α = 5%; Pelet terdiri atas talek + CMC 1.5% + gliserol 1%.
X: X. oryzae pv. oryzae.
Daya berkecambah benih terinfeksi X. oryzae pv. oryzae yang diberi perlakuan pelet menggunakan bakteri probiotik R. pickettii TT47, aktinomiset 6 atau aktinomiset 6 + endofit 467 tidak berbeda nyata dibandingkan kontrol (benih yang tidak diinfeksi) hingga 6 minggu penyimpanan. Sebaliknya, benih terinfeksi
X. oryzae pv. oryzae yang tidak diberi perlakuan pelet dan benih terinfeksi yang diberi perlakuan pelet menggunakan endofit 467 sudah mengalami penurunan DB yang nyata pada minggu ke-6 penyimpanan.
Berat kering kecambah normal benih terinfeksi X. oryzae pv. oryzae maupun benih yang tidak terinfeksi X. oryzae pv. oryzae dengan atau tanpa perlakuan pelet belum menunjukkan perbedaan yang nyata hingga minggu ke-4 penyimpanan. Penurunan BKKN baru terjadi pada minggu ke-6 penyimpanan. Penurunan yang
17 Tabel 5 Pengaruh interaksi perlakuan pelet dan periode simpan terhadap berat kering kecambah normal padi
Perlakuan Periode simpan (minggu)
0 2 4 6
BKKN (g)
X + pelet + aktinomiset 6 + endofit 467 0.14hi 0.26bcd 0.17efg 0.20efg X + pelet + aktinomiset 6 0.14hi 0.27bc 0.19e-h 0.11ij X + pelet + endofit 467 0.15ghi 0.33a 0.15ghi 0.07jk
Benih sehat [K] 0.18e-h 0.22def 0.16fgh 0.20efg
Benih direndam air 0.14hi 0.23cde 0.17e-h 0.17fgh
K + pelet 0.17e-h 0.29ab 0.20efg 0.18e-h
X + pelet + R. pickettii TT47 0.17fgh 0.34a 0.19e-h 0.16h-i
X. oryzae pv. oryzae [X] 0.17e-h 0.20efg 0.18e-h 0.05k
BKK (g)
X + pelet + aktinomiset 6 + endofit 467 0.0083e-h 0.0120bc 0.0090d-g 0.0093def X + pelet + aktinomiset 6 0.0077fgh 0.0130a-b 0.0100de 0.0097def X + pelet + endofit 467 0.0083e-h 0.0143a 0.0080e-h 0.0063h Benih sehat [K] 0.0080h-g 0.0097def 0.0083e-h 0.0097def Benih direndam air 0.0070gh 0.0100de 0.0077fgh 0.0100de
K + pelet 0.0087d-g 0.0137ab 0.0100de 0.0107cd
X + pelet + R. pickettii TT47 0.0080e-h 0.0140a 0.0090d-g 0.0077fgh
X. oryzae pv. oryzae [X] 0.0077fgh 0.0090d-g 0.0083e-h 0.0080e-h
Keterangan: Angka yang diikuti huruf yang sama pada semua kolom dan baris tidak berbeda nyata
pada DMRT taraf α = 5%; Pelet terdiri atas talek + CMC 1.5% + gliserol 1%.
X: X. oryzae pv. oryzae.
Sejak awal penyimpanan, KCT benih terinfeksi X. oryzae pv. oryzae nyata
lebih rendah dibanding benih yang tidak diinfeksi. Benih terinfeksi yang diberi perlakuan pelet mengalami penurunan KCT namun tidak berbeda nyata dengan
benih tidak terinfeksi sampai minggu ke 2. Perlakuan pelet menggunakan bakteri probiotik R. pickettii TT47 mampu mempertahankan KCT benih paling baik di
antara perlakuan benih terinfeksi lainnya selama 6 minggu penyimpanan yaitu sebesar 17.17% KN etmal-1.
Pemeletan benih padi terinfeksi X. oryzae pv. oryzae dengan bakteri probiotik R. pickettii TT47, aktinomiset 6 dan endofit 467 + aktinomiset 6 memiliki IV nyata lebih tinggi dibanding tanpa pelet (Tabel 7). Perlakuan pelet dengan bakteri endofit 467 saja belum mampu mempertahankan vigor benih terinfeksi X. oryzae pv. oryzae di penyimpanan, terlihat dari KCT dan IVyang tidak berbeda nyata
18
Tabel 6 Pengaruh interaksi perlakuan benih dan periode simpan terhadap kecepatan tumbuh benih padi Benih direndam air 19.28a-e 20.10a-c 19.28ab 20.88a
K + pelet 20.35ab 20.46ab 20.35ab 20.03a-d
X + pelet + R. pickettii TT47 17.04c-h 17.60b-h 17.04c-h 17.17c-h
X. oryzae pv. oryzae [X] 15.24h 16.84e-h 15.24h 11.02i
Keterangan: Angka yang diikuti huruf yang sama pada semua kolom dan baris tidak berbeda nyata
pada DMRT taraf α = 5%; Pelet terdiri atas talek + CMC 1.5% + gliserol 1%.
X: X. oryzae pv. oryzae.
Pada akhir penyimpanan (6 minggu) benih terinfeksi X. oryzae pv. oryzae
sudah mengalami penurunan DB (62.67%), KCT (11.02% etmal-1) dan
BKKN (0.05 g) namun belum mengalami penurunan PTM. Hal ini menunjukkan infeksi X. oryzae pv. oryzae pada benih padi mampu menghambat perkecambahan namun belum mematikan benih selama penyimpanan 6 minggu.
Tabel 7 Pengaruh faktor tunggal periode simpan dan perlakuan terhadap tolok ukur vigor serta viabilitas benih
Benih direndam air 76.33a 93.33a
K + pelet 79.00a 91.67a
X + pelet + R. pickettii TT47 57.00c 92.00a
X. oryzae pv. oryzae [X] 42.67d 89.33a
Keterangan: Angka yang diikuti huruf yang sama pada semua kolom dan baris tidak berbeda nyata
pada DMRT taraf α = 5%; Pelet terdiri atas talek + CMC 1.5% + gliserol 1%.
X: X. oryzae pv. oryzae.
Viabilitas benih padi terinfeksi X. oryzae pv. oryzae masih dapat dipertahankan dengan baik selama 6 minggu penyimpanan, dengan perlakuan pelet menggunakan R. pickettii TT47, aktinomiset 6 serta aktinomiset 6 + endofit 467. Hal ini mungkin dikarenakan pelet dengan penambahan bakteri tersebut mampu menekan perkembangan patogen pada benih padi terinfeksi dan menghasilkan hormon yang membantu mempercepat regulasi perkecambahan benih. Glick et al.
19 (antibiotik, enzim lisis, siderofor, ethylene) maupun menginduksi resistensi sistemik tanaman (ISR). Chung et al. (2015) menunjukkan aplikasi Bacillus strain YC7007 (107 cfu mL-1) pada benih padi yang baru berkecambah meningkatkan ISR dan pertumbuhan tanaman sampe ke booting stage.
Bakteri secara langsung mampu menyediakan nutrisi tambahan seperti N, P dan Fe serta mengatur level fitohormon (sitokinin, giberelin, IAA) yang dapat
menunjang perkecambahan, pertumbuhan dan perkembangan tanaman (Glick 2012). Bakteri probiotik seperti aktinomiset, Bacillus sp. dan R. pickettii
dilaporkan mampu menghasilkan hormon pertumbuhan seperti Indole Acetic Acid
(IAA) dan giberelin (Velusamy et al. 2013; Lestari et al. 2014).
Giberelin menginduksi sintesis enzim α-amilase yang berperan dalam perombakan pati untuk digunakan sebagai energi dalam perkecambahan benih (Palmiano dan Juliano 1972). Hormon IAA dibutuhkan tanaman setelah berkecambah untuk perpanjangan sel (Miransari dan Smith 2014). Subash et al.
(2015) menyatakan aplikasi 2 mg L-1 GA3 dan IAA pada benih Sesamum indicum
TVM-1 mampu meningkatkan perkecambahan, panjang akar dan tunas dibandingkan kontrol. Chithrashree et al. (2011) membuktikan seed treatment
menggunakan talek + CMC (0.2%) + Bacillus sp. mampu secara nyata meningkatkan perkecambahan (82%) dan vigor indeks (1309) benih padi terinfeksi
X. oryzae pv. oryzae dibanding kontrol (71% dan 850).
Mutu benih terbagi atas 4 kelompok yaitu mutu genetik, fisik, fisiologis dan kesehatan benih. Mutu fisiologis merujuk pada kemampuan benih berkecambah, meliputi viabilitas benih (Ilyas 2012). Viabilitas benih menunjukkan daya hidup benih, aktif secara metabolis dan memiliki enzim yang dapat mengkatalisis reaksi metabolis yang diperlukan untuk perkecambahan dan pertumbuhan kecambah (Ilyas 2012). Parameter viabilitas benih yaitu viabilitas potensial dan vigor. Viabilitas potensial merupakan kemampuan benih untuk tumbuh dan berproduksi normal dalam keadaan optimum sementara vigor dalam keadaan sub optimum (Widajati et al. 2013). Pada penelitian ini, perlakuan pelet dengan penambahan inokulan bakteri probiotik aktinomiset 6, R. pickettii TT47, endofit 467 atau kombinasi endofit 467 + aktinomiset 6 tidak hanya dapat mempertahankan mutu fisik benih, namun juga kesehatan (menekan perkembangan patogen terbawa benih
X. oryzae pv. oryzae) dan mutu fisiologis benih padi terinfeksi X. oryzae pv. oryzae
(viabilitas benih terinfeksi selama 6 minggu penyimpanan).
Pembahasan Umum
Pengendalian patogen terbawa benih X. oryzae pv. oryzae menggunakan agen antagonis seperti bakteri probiotik merupakan alternatif yang menjanjikan dan ramah lingkungan. Bakteri probiotik dapat diaplikasikan langsung pada benih melalui teknik pelet untuk memudahkan penyimpanan dan distribusi dalam skala luas. Pengembangan formula pelet yang diharapkan adalah dapat membawa serta mempertahankan viabilitas bakteri probiotik dan mutu benih menjadi fokus penelitian ini.
Penelitian ini menunjukkan formula pelet dengan penambahan bakteri probiotik R. pickettii TT47, aktinomiset 6, endofit 467 maupun kombinasi aktinomiset 6 dengan endofit 467 dapat mempertahankan viabilitas benih padi terinfeksi X. oryzae pv. oryzae hingga 6 minggu penyimpanan. Namun, formula pelet yang paling baik dalam menekan X. oryzae pv. oryzae pada benih padi
terinfeksi adalah formula pelet dengan penambahan bakteri probiotik aktinomiset 6 dalam bentuk tunggal maupun kombinasi dengan endofit 467.
Formula pelet ini terbukti efektif menekan perkembangan patogen X. oryzae pv.
20
dan panas (Emmert dan Handelsman 1999) sehingga mampu bertahan dalam formula pelet dengan lebih baik dibandingkan R. pickettii TT47 yang tidak memiliki sruktur bertahan.
Kemampuan antagonis beberapa spesies bakteri golongan aktinomiset dan endofit telah dilaporkan sebelumnya oleh Koberl et al. (2013), dimana
Streptomyces peucetius, S. scabiei, S. subrutilus memiliki kemampuan antagonis terhadap Verticillium dahlia, Rhizoctonia solani, Fusarium culmorum, Ralstonia solanacearum dan Meloidogyne incognita. Yuliar (2014) menyatakan bakteri endofit yang diisolasi dari tanaman Avicenia alba, A.marina dan Bruguiera ghymnorhiza terbukti antagonis terhadap R. solani (7 strain), F. oxysporum (5 strain) dan antagonis terhadap X. oryzae pv. oryzae (2 strain). El-shakh et al. (2015) menunjukkan 5 isolat Bacillus endofit yang diisolasi dari benih padi mampu menghambat perkembangan X. oryzae pv. oryzae secara in vitro dan di greenhouse, bakteri ini juga mampu menghasilkan IAA dan siderofor.
Kombinasi endofit dan aktinomiset dalam formula pelet juga tidak menurunkan kemampuan pelet dalam menekan patogen dan mempertahankan viabilitas benih padi terinfeksi X. oryzae pv. oryzae. Raupach dan Kloepper (1998) menyatakan kombinasi agen hayati yang memiliki mekanisme berbeda lebih menguntungkan bagi tanaman dibandingkan aplikasi tunggal mengingat kondisi di lapangan yang lebih ekstrim. Xu dan Jegger (2013) membuktikan aplikasi tunggal agen biokontrol yang memiliki mekanisme berbeda lebih efektif dibandingkan kombinasi agen bikontrol dengan mekanisme yang sama. Nagendran et al. (2013) menyimpulkan bakteri endofit tepat diaplikasikan bersama dengan aktinomiset karena memiliki lingkungan hidup yang berbeda di lapangan sehingga tidak terjadi kompetisi ruang/nutrisi antar bakteri yang digunakan. selain itu, endofit diharapkan dapat masuk dan mengkolonisasi jaringan tanaman dengan cepat agar mampu menekan patogen yang berada di sekitar jaringan vaskular tanaman seperti patogen
X. oryzae pv. oryzae.
Hasil penelitian menunjukkan bakteri Gram positif seperti aktinomiset 6 + endofit 467 dapat bertahan dalam formula pelet dan tidak menekan perkembangan salah satu bakteri. Kombinasi bakteri ini dalam formula pelet efektif menekan perkembangan patogen X. oryzae pv. oryzae pada benih serta mempertahankan viabilitas benih. Sehingga, aplikasi formula pelet dengan penambahan bakteri probiotik aktinomiset 6 + endofit 467 pada benih padi dapat digunakan sebagai salah satu cara untuk mengendalikan patogen terbawa benih X. oryzae pv. oryzae.
21
5
SIMPULAN DAN SARAN
Simpulan
Bakteri probiotik R. pickettii TT47, endofit 467 dan aktinomiset 6 terbukti antagonis terhadap patogen X. oryzae pv. oryzae. Isolat yang kompatibel adalah endofit 467 dan aktinomiset 6. Formulasi pelet terbaik terdiri dari: talek + CMC 1.5% + gliserol 1%. Formulasi ini mampu mempertahankan populasi bakteri probiotik endofit 467, aktinomiset 6 dan endofit 467 + aktinomiset 6 selama 4 minggu simpan serta populasi R. pickettii TT47 selama 3 minggu simpan. Perlakuan pelet dengan penambahan bakteri probiotik R. pickettii TT47, aktinomiset 6 atau aktinomiset 6 + endofit 467 efektif menekan patogen X. oryzae pv. oryzae dan mempertahankan mutu benih padi terinfeksi selama 6 minggu penyimpanan.
Perlakuan pelet menggunakan bakteri probiotik aktinomiset 6 serta kombinasi
aktinomiset 6 dan endofit 467 terbukti paling efektif dalam menekan patogen
X. oryzae pv. oryzae serta mempertahankan viabilitas benih terinfeksi berdasarkan tolok ukur DB, BKKN, IV dan KCT.
Saran
Perlakuan pelet dengan bakteri probiotik R pickettii TT47 lebih efektif digunakan untuk penekanan patogen pada benih yang langsung akan ditanam. Perlakuan pelet untuk benih yang akan disimpan atau didistribusikan, lebih disarankan menggunakan bakteri probiotik aktinomiset 6 tunggal maupun kombinasi dengan endofit 467.
22
DAFTAR PUSTAKA
Agustiansyah, Ilyas S, Sudarsono, Machmud M. 2010. Pengaruh perlakuan benih secara hayati pada benih padi terinfeksi Xanthomonas oryzae pv. oryzae
terhadap mutu benih dan pertumbuhan bibit. J Agron Indonesia. 38:185-191. Agustiansyah, Ilyas S, Sudarsono, Machmud M. 2013. Karakterisasi rizobakteri yang berpotensi mengendalikan bakteri Xanthomonas oryzae pv. oryzae dan meningkatkan pertumbuhan tanaman padi. J HPT Tropika. (13)1:42-51. Ali NI, SiddiQui IA, Shaukat SS dan Zaki MJ. 2001. Survival of Pseudomonas
aeruginosa in various carriers for the inhibition of root rot-root knot disease complex of mungbean. Phytopath Mediterr. 40:108–112.
Amalia, A. F. 2014. Formulasi aktinomiset menggunakan beberapa jenis tanah sebagai agens hayati penyakit kresek (Xanthomonas oryzae pv. oryzae) pada padi. Skripsi. Departemen Proteksi Tanaman, Fakultas Pertanian. Institut Pertanian Bogor.
Andrews SC, Robinson AK and Rodríguez-Quiñones F. 2003. Bacterial iron homeostasis. FEMS Microbiol Rev. 27:215-237.
Backman PA, Sikora RA. 2008. Endophytes: An Emerging Tool for Biological Control. Biological Control (July):1-8. DOI: 10.1016/j.biocontrol.2008.03. 009
Beric T, Kojic M, Stankovic S, Topisirovic L, Degrassi G, Myers M, Venturi V, Fira D. 2012. Antimicrobial activity of Bacillus sp. natural isolates and their potential use in the biocontrol of phytopathogenic bacteria. Food Technol Biotechnol. 50(1):25-31.
[BSN] Badan Standardisasi Nasional. 2003. Benih padi kelas dasar (BD). BSN [internet]. [diunduh 23 November 2015]. Tersedia pada: http://sisni.bsn.go.id/index.php?/sni_ main/sni/detail_sni/5774.
Chithrashree AC. Udayashankar, Nayaka SC, Reddy MS, Srinivas C. 2011. Plant growth-promoting rhizobacteria mediate induced systemic resistance in rice against bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. Bio Control. 59:114–122. DOI: 10.1016/j.biocontrol.2011.06.010.
Copeland LO, McDonald MB.1995. Seed Science and Technology. Chapman and Hall, NY.
Crosa JH, Walsh CT. 2002. Genetics and assembly line enzymology of siderophore biosynthesis in bacteria. Microbiol Mol Biol Rev. 66:223-249.
Dardick C, da Silva FG, Shen Y, Ronald P. 20013. Antagonistic interactions between strains of Xanthomonas oryzae pv. oryzae. APS 93(6):705-711. [Ditlitanpang] Direktorat Perlindungan Tanaman Pangan. 2014. Data opt padi.
http://tanamanpangan.pertanian.go.id/ditlintp/statis-17-dataoptpadi.html [diakses 2 Maret 2014].
Dzidic S, Suskovic J, Kos B. 2008. Antibiotic resistance mechanisms in bacteria: biochemical and genetic aspects. Food Technol Biotechnol. 46(1):11–21. El-shakh ASA, Kakar KU, Wang X, Almoneafy AA, Ojaghian MR, Li B, Anjum
SI, Xie GL. 2015. Controlling bacterial leaf blight of rice and enhancing the plant growth with Endophytic and Rhizobacterial Bacillus strains. Toxicol environt chem. :1-36. DOI:10.1080/02772248.2015.1066176.
Elzein A, Kroschel J, Cadisch G. 2008. Efficacy of Pesta granular formulation of
Striga-mycoherbicide Fusarium oxysporum f. sp. strigae Foxy 2 after 5-year of storage. J Plant Dis Protect. 115(6):259–262.