NIKEN SUBEKTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyataan dalam disertasi yang berjudul: Kelimpahan, Sebaran, dan Arsitektur Sarang serta Ukuran Populasi Rayap Tanah Macrotermes gilvus Hagen (Blattodea : Termitidae) di Cagar Alam Yanlappa, Jawa Barat adalah karya saya sendiri dan belum pernah penulis ajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Agustus 2010
NIKEN SUBEKTI. Distribution Patterns, Architecture and Population Density of Subterranean Termites Macrotermes gilvus Hagen (Blattodea : Termitidae) in Yanlappa Nature Reserve, West Java. Supervised by DEDY DURYADI SOLIHIN, DODI NANDIKA, SURJONO SURJOKUSUMO, and SYAIFUL ANWAR.
A study was conducted to distribution patterns, mound architecture and population density in Yanlappa Nature Reserve, West Java. The data collected included the geographic position mound of M. gilvus by using the GPS, mound distribution patterns, slope, Leaf Area Index (LAI). The mound of subterranean termite are classified into the types based on size, namely small mounds (≤ 0,49 m tall), medium mounds (0,5-0,99 m tall), and large mounds (≥ 1 m tall). In addition, a vegetation was also analysis with a plot method of 20 x 20 m for trees, 10 x 10 m for small trees, and 5 x 5 m for seedling. The soils and queens room soils each mounds from outside and inside the mounds from 0-20 cm deep (outer zone), 20-40 medium zone, and 40 - 60 cm (inner zone), were also sampled for chemical and physical analyses. The physical analyses included soil texture, porosity, bulk density, water content, temperature, aggregate stability tests, soil humidity, and mounds material pressure test. Meanwhile, the chemical observation included pH, CO2, CH4, organic matter contents, organic C content, and total N. The results of this study showed that the mounds distribution pattern is clustered in the slopes of 3-8% and canopy coverage Leaf Area Index of (0-2). The shape of M. gilvus mounds are dome-shaped with a tall and wide base diameter that depend on the colony age. The whole mounds surface is coverred with porous litter layer. The main architecture in the mounds is a wall with ventilation, mounds center, mold plantation, queen room, and nomad holes, and closed air circulation system. The average diameter of the mounds base is 2.06 ± 0,67 m, the average height of mounds center from the ground is 71,33 ± 4,50 cm, while the average diameter of mounds center is 32,33 ± 4,17 cm, the average mounds height is 1,00 ± 0,64 m, preassure resistance 25,6 ± 3,62 kg/cm2, and mounds hardiness 46,66 ± 9,81 kg/cm2. The queen room wall is stronger than the other mounds walls. Population M. gilvus in large mound in the natural forest was 183.825 ± 0.71 individuals, middle mound is 46.267 ± 0.17 individuals, and small mound is 20.223 ± 0.50 individuals.
NIKEN SUBEKTI. Kelimpahan, Sebaran, dan Arsitektur Sarang serta Ukuran Populasi Rayap Tanah Macrotermes gilvus Hagen (Blattodea : Termitidae) di Cagar Alam Yanlappa, Jawa Barat. Dibimbing oleh DEDY DURYADI SOLIHIN, DODI NANDIKA, SURJONO SURJOKUSUMO, dan SYAIFUL ANWAR.
Macrotermes gilvus Hagen (Blattodea: Termitidae) merupakan spesies rayap tanah yang paling luas sebaran geografisnya di Indonesia, khususnya di hutan alam dataran rendah. Disamping itu, keberadaan rayap ini juga paling mudah dikenali kerena sarangnya berbentuk bukit kecil (mound), mencuat di atas permukaan tanah. Namun demikian pengetahuan tentang kelimpahan, sebaran, dan arsitektur sarang serta ukuran populasi spesies rayap tersebut di Indonesia belum pernah dilaporkan. Suatu penelitian telah dilakukan untuk mengetahui kelimpahan, sebaran, dan arsitektur sarang rayap M. gilvus serta ukuran populasinya di Cagar Alam Yanlappa, Jawa Barat. Untuk mengetahui keberadaan sarang rayap M. gilvus di lokasi penelitian dibuat transek melintang dari arah Tenggara ke Barat Laut dengan jarak antar transek 50 meter dan panjang keseluruhan 6.800 meter. Setiap sarang M. gilvus yang ditemukan ditentukan koordinat dan ketinggiannya dari permukaan laut (altitude) dengan menggunakan Global Position System (GPS), kemudian dicatat morfologinya (bentuk dan dimensi sarang termasuk tinggi, diameter, luas bidang dasar, kemiringan dinding sarang, kondisi permukaan dinding sarang, dan kedalaman dasar sarang dari permukaan tanah). Disamping itu, untuk mengetahui arsitektur bagian dalam sarang, termasuk tata ruang sarang, sembilan sarang contoh yang terdiri dari 3 sarang berukuran besar (tinggi sarang ≥ 1 m), 3 sarang berukuran sedang (tinggi sarang 0,5-0,99 m), dan 3 sarang berukuran kecil (tinggi sarang ≤ 0,49 m) dibongkar (dismantled) secara bertahap, baik secara vertikal maupun horisontal. Pembongkaran sarang dilakukan sampai dasar sarang. Pada saat pembongkaran sarang contoh, seluruh spesimen rayap pada masing-masing sarang dikumpulkan dengan menggunakan vacum cleaner (3,5 kVA), kemudian dihitung dan dikelompokkan berdasarkan kastanya. Contoh spesimen per kasta juga dikoleksi dalam botol koleksi berisi alkohol 70%. Sementara itu, contoh bahan pembentuk dinding sarang dan komponen bagian sarang lainnya, serta tanah di luar sarang diuji sifat fisis (tekstur, porositas, bobot jenis, kadar air, kemantapan agregat, kelembaban), kimiawi (pH, kandungan bahan organik, N total), serta sifat mekanisnya (kekuatan tekan). Analisis vegetasi dilakukan pada plot-plot pengamatan berukuran 20 x 20 m untuk tingkat pohon, 10 x 10 m untuk tingkat tiang, 5 x 5 m untuk tingkat semai dengan intensitas pengambilan contoh sebesar 0,38%. Disamping itu, dilakukan analisis indeks tutupan tajuk (leaf area index) dan pencatatan tingkat kemiringan lereng di sekitar sarang M. gilvus.
Hasil penelitian menunjukkan bahwa di Cagar Alam Yanlappa, Jawa Barat ditemukan 155 sarang M. gilvus terdiri dari 56 sarang berukuran besar (36,13%), 94 sarang berukuran sedang (60,65%), dan 5 sarang berukuran kecil (3,22%). Kelimpahan sarang pada ketinggian 100-150 m dpl mencapai 10 sarang/Ha, sementara pada ketinggian 50-100 m dpl adalah 3 sarang/Ha. Sarang-sarang rayap tersebut menyebar secara berkelompok (cluster) pada kemiringan lereng 3% - 8% dan tutupan tajuk terbuka (LAI = 0) sampai tertutup ringan (LAI = 2). Tapak sarang berada pada lahan berdrainase baik, tetapi tidak terkait dengan tipe vegetasi tertentu.
Sarang M. gilvus berukuran besar dihuni oleh 183.825 ± 7.742 individu (kasta pekerja, kasta prajurit dan nimfa), sementara itu sarang ukuran sedang dihuni oleh 46.267 ± 2.073 individu, dan sarang ukuran kecil dihuni oleh 20.223 ± 1.919 individu. Struktur populasi koloni rayap tanah M. gilvus terdiri dari kasta reproduktif, kasta prajurit, kasta pekerja serta telur dan nimpa yang kelimpahannya bervariasi sesuai dengan ukuran sarang (umur koloni rayap). Struktur populasi tersebut didominasi oleh kasta pekerja yang memang merupakan inti dari koloni. Jumlah nimfa pada sarang berukuran besar dan berukuran sedang pada umumnya sangat banyak (82,5%). Hal ini diduga karena sarang berukuran sedang dan berukuran besar berada pada fase perkembangbiakan koloni yang sangat pesat. Ciri lain dari sarang berukuran besar dan sarang berukuran sedang adalah adanya kasta reproduktif sekunder (neoten). Hasil penelitian menunjukkan bahwa biomassa rayap M. gilvus terbesar (46%) adalah kasta prajurit mayor, disusul oleh kasta prajurit minor (26%), kasta pekerja (18%), dan nimfa (10%). Secara keseluruhan ukuran populasi rayap M. gilvus di Cagar Alam Yanlappa, Jawa Barat adalah 14.744.912 ± 2.391 individu. Keberadaan koloni rayap tanah M. gilvus di Cagar Alam Yanlappa Jawa Barat diyakini sangat berpengaruh terhadap eksistensi dan kelestarian ekosistem hutan tersebut. Penelitian dan pengembangan tentang ekologi populasi rayap tanah M. gilvus sangat strategis untuk mendukung pengelolaan ekosistem hutan, termasuk untuk mengembangkan teknologi bioremediasi lahan-lahan kritis.
© Hak cipta milik IPB, tahun 2010
Hak cipta dilindungi
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tujuan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
NIKEN SUBEKTI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Nama : Niken Subekti NIM : G. 361050021
Disetujui Komisi Pembimbing
Dr.Ir.Dedy Duryadi Solihin DEA
Ketua Anggota
Prof.Dr.Ir.Dodi Nandika, M.S
Prof.(Em).Ir.Surjono Surjokusumo,MSF,PhD Dr.Ir.Syaiful Anwar, M.Sc
Anggota Anggota
Ketua Program Studi Biologi Dekan Sekolah Pascasarjana
Dr.Ir.Dedy Duryadi Solihin, DEA Prof.Dr.Ir. Khairil Anwar Notodiputro, M.S
memberikan hidayah dan rahmat-Nya sehingga penyusunan disertasi ini dapat diselesaikan. Disertasi dengan judul “KELIMPAHAN, SEBARAN, DAN ARSITEKTUR SARANG SERTA UKURAN POPULASI RAYAP TANAH Macrotermes gilvus Hagen (Blattodea : Termitidae) DI CAGAR ALAM YANLAPPA, JAWA BARAT” merupakan salah satu syarat untuk memperoleh gelar Doktor di Program Studi Biologi Sekolah Pascasarjana IPB. Penelitian ini memuat beberapa topik seperti: (1) Pola Sebaran Rayap Tanah M. gilvus (2) Ukuran Populasi Rayap M. gilvus (3) Struktur dan Arsitektur Bangunan Sarang Rayap M. gilvus. Sebagian penelitian ini dibiayai oleh Direktorat Pendidikan Tinggi melalui Program Hibah Pascasarjana dan Program Sandwich.
Pada kesempatan ini saya menyampaikan rasa terima kasih dan penghargaan pada berbagai pihak yang telah membantu dengan caranya masing-masing dalam studi, penelitian maupun penyempurnaan tulisan disertasi:
(1) Kepada keluarga: suami dan Ibunda yang saya cintai, rasa hormat dan terima kasih yang tidak pernah habis kepada keluarga besar Maskuri (Alm), AKBP.Ir. Slamet Iswanto dan keluarga, Mbak Yetty sekeluarga, dan Mbak Tutik sekeluarga. (2) Kepada komisi pembimbing, Dr.Ir. Dedy Duryadi Solihin, DEA. Yang telah banyak
memberikan pengetahuan, ketajaman dalam menganalisis, penulisan dan saran-saran dalam disertasi. Prof.Dr.Ir. Dodi Nandika, MS. yang ahli dibidang Termitologi sangat banyak memberikan arahan ketajaman dalam detail-detail penelitian, motivasi yang tinggi untuk kemajuan berfikir ilmiah dan saran-saran dalam penulisan disertasi. Prof. Surjono Surjokusumo, MSF,PhD. yang selalu memberikan nasehat, masukan dan pengarahan dengan penuh kasih sayang dan kesabaran dalam penulisan disertasi. Dr.Ir. Syaiful Anwar M.Sc. yang telah memberikan saran, motivasi dan masukan dalam disertasi.
(3) Kepada Rektor, Ketua LPPM, Dekan dan Ketua Jurusan Biologi FMIPA Universitas Negeri Semarang yang telah memberikan kesempatan studi dan mendukung perjalanan ilmiah di Sekolah Pascasarjana IPB
(4) Dekan Sekolah Pascasarjana IPB beserta staf yang telah banyak membantu memperoleh kesempatan dalam Program Sandwich di Jepang dan program Hibah Doktoral serta pelayanan administrasi dalam penyelesaian studi S3
(5) Kepada Prof. Tsuyoshi Yoshimura dan Dr. Kunio Tsunoda yang telah membimbing selama di RISH, Kyoto University Japan. Teman-teman di Research Institute Sustainable and Humanosphere: Prof.Dr.Ir. Yusuf Sudo Hadi, Dr. Joko Sulistyo, Motoko Fujita PhD, Nakai-kun, Tashiro-san, Toyomi-san, dan Nakayama-san yang selalu membantu dalam penelitian dan telah memperkenalkan kebudayaan dan kehidupan orang Jepang.
(6) Kepada Prof. Chow Yang Lee dan Neoh Kok Boon, PhD dari USM Malaysia, David T. Jones PhD dari NHM Inggris, Akinori Yamada PhD dari Kyoto University, Dr. Anggoro Prasetyo (Alm) dari LIPI. Terima kasih atas bantuan identifikasi rayap dan jurnal-jurnal ilmiahnya.
(7) Kepada sahabat dan teman di Lab PPSHB IPB: Mbak Arinana dan keluarga, Mbak Diba dan keluarga, Mas Yudi dan keluarga, Efendi, Mas Agus Nuryanto, Mbak Rosa, Mbak Fahma, Mbak Suriana, Nana, Mas Tri, Pak Heri. Terima kasih atas kebersamaan, semangat, dan persaudaraan yang indah selama penelitian.
dosen
(10) Keluarga Bapak Soleh (Alm) dan Satria di Cagar Alam Yanlappa, keluarga Abah dan Solihin di Taman Nasional Ujung Kulon, Mas Inyong di Taman Nasional Gunung Halimun Salak, Budi, Iwan, Rudi, Tukul, Beny, Ryan, Agus, Rizal, Cepy, Irwandi, yang membantu dalam survey, pembongkaran sarang dan pengambilan sampel rayap di hutan
(11) Akhirnya kepada sahabat, teman dan pihak-pihak yang tidak bisa saya sebutkan satu per satu, terima kasih atas bantuan yang diberikan selama saya studi S3 dan saya berharap semoga tulisan ini bermanfaat bagi pihak yang berkepentingan
Bogor, Agustus 2010
Penulis dilahirkan di Jepara pada tanggal 14 Februari 1973 sebagai anak keempat dari empat bersaudara dari pasangan yang berbahagia Maskuri (Alm) dan Siti Qodri. Penulis menamatkan Sekolah Dasar Prambatan Kidul 2 Kudus pada tahun 1984, kemudian melanjutkan ke SMP Negeri 1 Kudus dan lulus pada tahun 1987. Pada tahun 1990 penulis lulus dari SMA Negeri 2 Kudus kemudian melanjutkan studi di Fakultas Biologi, Jurusan Biologi Lingkungan Universitas Jenderal Soedirman Purwokerto.
Pada tahun 1995 penulis lulus dan mendapat gelar Sarjana Sains dan diterima sebagai staf pengajar di jurusan Biologi FMIPA Universitas Negeri Semarang pada tahun 1999. Pada tahun 2001 penulis mendapat Beasiswa Pendidikan Pascasarjana dari Direktorat Pendidikan Tinggi, Kementerian Pendidikan Nasional untuk mengikuti Program Magister pada Sekolah Pascasarjana Institut Pertanian Bogor Program Studi Biologi, lulus dengan mendapat gelar Magister Sains pada tahun 2004. Pada tahun 2005 penulis kembali mendapatkan beasiswa BPPS dari Ditjen DIKTI Kementerian Pendidikan Nasional untuk melanjutkan pendidikan Program Doktor di IPB dengan bidang studi Biologi minat Entomologi Hutan. Penulis diberi kesempatan penelitian di RISH (Research Institute Sustainable and Humanosphere) Kyoto University, Jepang pada tanggal 4 Oktober 2008 – 13 Maret 2009 oleh pascasarjana IPB melalui program sandwich yang diselenggarakan oleh DIKTI.
Sebagian hasil penelitian ini telah dipresentasikan pada Seminar Internasional Termite Research Group (TRG), MAPEKI (Masyarakat Peneliti Kayu Indonesia), dan PEI (Perhimpunan Entomologi Indonesia).
Penguji Luar Komisi:
Ujian Tertutup : tanggal pelaksanaan 8 Maret 2010
1. Dr.Ir. Sulaeman Yusuf, M.Agr (Peneliti Utama LIPI)
2. Dr.Ir. Idham Sakti Harahap
(Departemen Hama dan Penyakit Tanaman IPB)
Ujian Terbuka : tanggal pelaksanaan 9 Juli 2010
1. Prof.(Em). Rudy C.Tarumingkeng, M.SF, PhD (Departemen Teknologi Hasil Hutan IPB) 2. Prof. Dr. Rochadi Abdulhadi
xii Latar belakang penelitian………... 1
Tujuan penelitian………... 2
III. KELIMPAHAN DAN SEBARAN SARANGMacrotermes gilvus Hagen Abstrak……… 12
IV. ARSITEKTUR BANGUNAN SARANG RAYAP Macrotermes gilvus Hagen
xiii
DAFTAR PUSTAKA……… 82
xiv
Halaman
1. Kelimpahan sarang rayap tanah M. gilvus berdasarkan ketinggian,
nilai LAI dan kemiringan lahan areal di Cagar Alam Yanlappa... 21
2. Karakteristik sarang rayap tanah Macrotermes gilvus Hagen
pada berbagai tipe sarang di Cagar Alam Yanlappa, Jawa Barat... 30
3. Karakteristik konstruksi sarang rayap tanah Macrotermes gilvus Hagen
di Cagar Alam Yanlappa, Jawa Barat………. 33
4. Jumlah individu dalam koloni pada berbagai tipe sarang
di Cagar Alam Yanlappa, Jawa Barat... 66
5. Berat basah rayap tanah Macrotermes gilvus Hagen
dengan menggunakan uji Tukey………... 68
6. Berat kering rayap tanah Macrotermes gilvus Hagen
xv
1. Kasta dalam koloni rayap tanah Macrotermes gilvus Hagen... 7
2. Peta sebaran rayap Macrotermes gilvus di dunia... 13
3. Peta sebaran rayap tanah Macrotermes gilvus
di areal Cagar Alam Yanlappa Jawa Barat... 20
4. Kondisi tutupan tajuk areal Cagar Alam Yanlappa, Jawa Barat:
tertutup ringan dan terbuka ...………... 21
5. Tiga tipologi bentuk dan bidang dasar sarang M. gilvus
di Cagar Alam Yanlappa, Jawa Barat... 31
6. Irisan vertikal sarang rayap tanah M. gilvus Hagen
di Cagar Alam Yanlappa... 32
7. Contoh material pembentuk dinding sarang (A), dan dinding bilik ratu (B) pada sarang rayap tanah M. gilvus
di Cagar Alam Yanlappa... 34
8. Irisan vertikal sarang tipe kecil rayap tanah M. gilvus
di Cagar Alam Yanlappa... 36
9. Irisan vertikal sarang tipe sedang rayap tanah M. gilvus
di Cagar Alam Yanlappa... 37
10. Irisan vertikal sarang tipe besar rayap tanah M. gilvus
di Cagar Alam Yanlappa... 39 .
11. Irisan horisontal sarang tipe kecil rayap tanah M. gilvus
di Cagar Alam Yanlappa... 41
12. Irisan horisontal sarang tipe sedang rayap tanah M. gilvus
di Cagar Alam Yanlappa... 42
13. Irisan horisontal sarang tipe besar rayap tanah M. gilvus
di Cagar Alam Yanlappa... 43
14. Permukaan dinding pusat sarang rayap tanah M. gilvus Hagen
xvi
16. Tampak luar bilik ratu pada sarang rayap tanah M. gilvus
di Cagar Alam Yanlappa... 49
17. Kebun jamur di dalam (A) dan di luar pusat sarang rayap tanah
M. gilvus ... 51
18. Irisan vertikal liang kembara di dalam sarang rayap tanah
M. gilvus... 52
19. Ukuran liang kembara di dalam sarang rayap M. gilvus yang merupakan jalan rayap untuk melakukan aktivitas
foraging... 53
20. Penampang melintang lubang ventilasi di dalam sarang rayap
M. gilvus.... 55
21. Lempeng-lempeng kebun jamur (LK) di dalam sarang rayap tanah M. gilvus yang membentuk rongga-rongga
udara (RU)... 55
22. Tipologi bangunan sarang rayap tanah Macrotermes gilvus Hagen di Cagar Alam Yanlappa (A), dan rayap tanah Macotermes jeanneli
di Afrika (B)... 59
23. Kondisi salah satu sarang rayap tanah M. gilvus sebelum dan pada saat pembongkaran untuk penghitungan kelimpahan
populasi:... 65
24. Peranan rayap tanah M. gilvus sebagai bioprospektor dalam
xvii
Halaman
1. Peta lokasi Cagar Alam Yanlappa ... 91
2. Peta tanah kawasan Cagar Alam Yanlappa... 92
3. Peta vegetasi Cagar Alam Yanlappa... 93
4. Daftar koordinat dan ketinggian lokasi sarang rayap tanah
M. gilvus di Cagar Alam Yanlappa, Jawa Barat... 94
5. Identifikasi rayap tanah M. gilvus Hagen di Natural History Museum, London………. 98
6. Tinggi dan luas bidang sarang rayap tanah M. gilvus
di Cagar Alam Yanlappa... 99
7. Lokasi sarang rayap M. gilvus berdasarkan kemiringan lereng, indeks
Penutupan tajuk (LAI) dan jarak ke sarang terdekat... 103
8. Analisis pola sebaran sarang rayap M. gilvus
di Cagar Alam Yanlappa... 107
9. Korelasi antara jumlah total individu rayap tanah M. gilvus
dengan ukuran sarangnya di Cagar Alam Yanlappa... 108
10. Spesies pohon dengan index nilai penting (INP) tertinggi
yang tumbuh di kelompok tapak sebaran... 110
11. Famili pohon dengan index nilai penting (INP) tertinggi
yang tumbuh di kelompok tapak... 111
12. Hasil penghitungan kelimpahan rayap tanah M. gilvus
di Cagar Alam Yanlappa... 112
13. Morfologi tiga tipe sarang rayap tanah M.gilvus
xviii 19. Daftar jenis dan famili pancang
pada wilayah sedikit sarang M. gilvus... 119
20. Daftar jenis dan famili tiang
pada wilayah sedikit sarang M. gilvus... 121
21. Daftar jenis dan famili pohon
pada wilayah sedikit sarang M. gilvus... 122
22. Daftar jenis dan famili semai
pada wilayah banyak sarang M. gilvus... 123
23. Daftar jenis dan famili pancang
pada wilayah banyak sarang M. gilvus... 124
24. Daftar jenis dan famili tiang
pada wilayah banyak sarang M. gilvus... 125
25. Daftar jenis dan famili pohon
Latar Belakang
Salah satu spesies rayap tanah yang paling luas sebaran geografisnya di Indonesia adalah Macrotermes gilvus Hagen (Blattodea : Termitidae). Spesies rayap ini dapat ditemukan hampir di seluruh pulau, dari Sumatera sampai Papua, dari Miangas sampai Pulau Rote. Bahkan di beberapa kawasan konservasi di Jawa Barat, spesies rayap ini ditemukan di dataran rendah sampai dataran tinggi (1500 m dpl), dari pantai sampai pegunungan1
Keunikan lain dari rayap tanah M. gilvus adalah hubungan simbiotiknya dengan jamur, khususnya jamur Termitomyces, yang “dibudidayakan” oleh koloni rayap tersebut di dalam sarangnya. Oleh karena itu di dalam sarang M. gilvus terdapat “kebun-kebun jamur” (fungus combs) sebagai sumber makanan bagi anggota koloni rayap tersebut. Jamur Termitomyces yang ditanam memiliki kandungan enzim selulase, glukose, galaktose, mannose, arabinose, dan xylose yang sangat berguna bagi rayap untuk mendegradasi bahan organik (Ohkuma 2003). Sementara itu, serasah, liur, kandungan karbon dan nitrogen serta kelembaban yang tinggi merupakan kondisi optimal bagi jamur Termitomyces untuk tumbuh dan berkembang. Nodul dari jamur Termitomyces merupakan sumber makanan bagi nimfa, kasta pekerja mayor, dan kasta pekerja minor untuk pertumbuhannya (Pearce 1997). Sejalan dengan itu, Miura & Matsumoto (1995) melaporkan bahwa spora jamur dibawa oleh rayap dari luar sarang ke dalam sarang dengan menggunakan tungkai dan antennanya. Tugas menseleksi dan “menanam” spora yang diperlukan koloni dilakukan oleh kasta pekerja minor.
)
. Kehadiran rayap tersebut di ekosistem alaminya sangat mudah diketahui karena bangunan sarangnya sangat unik, berbentuk gundukan tanah atau bukit kecil (mound) yang ukurannya beragam tergantung umur koloni rayap yang hidup di dalamnya. Konstruksi sarang tersebut sangat kokoh, tahan terhadap curahan air hujan, teriknya sinar matahari, dan hempasan angin, serta gangguan alam lainnya termasuk gempa. Bahkan keberadaan sarang M. gilvus pada suatu tapak atau hamparan lahan dapat mencapai puluhan tahun dengan ukuran yang terus “tumbuh” membesar sejalan dengan umur koloni rayap yang hidup didalamnya.
1)
Sementara itu untuk membawa spora jamur dari luar ke dalam sarang dilakukan oleh kasta pekerja mayor. Bahan utama pembentuk sarang M. gilvus adalah serasah, tanah, dan liur rayap (Krishna & Weesner 1969). Sarang berfungsi tidak saja sebagai tempat kawin ratu dan raja tetapi juga sebagai tempat memperbanyak koloni yang dihasilkan pasangan rayap tersebut. Lebih dari itu sarang berfungsi melindungi koloni rayap dari pengaruh lingkungan luar dan musuh alaminya.
Penelitian di berbagai negara telah mengungkapkan keunikan dan kompleksitas ekosistem sarang Macrotermes spp. (Meyer 2000, Turner 2000, Korb 2003, dan Inoue et al. 2006). Namun demikian pengetahuan tentang arsitektur dan ekosistem sarang M. gilvus di Indonesia belum pernah dilaporkan. Padahal diduga kuat kehadiran rayap ini memiliki peran ekologis (ecological role) yang sangat penting khususnya di ekosistem alaminya. Spesies rayap tersebut juga diduga berpotensi untuk dikembangkan sebagai agen biologis dalam bioremediasi lahan kritis.
Berdasarkan hal-hal tersebut di atas dirasa perlu melakukan penelitian tentang kelimpahan, sebaran, dan arsitektur sarang serta ukuran populasi rayap tanah M. gilvus di ekosistem alaminya.
Tujuan Penelitian
Penelitian bertujuan untuk mengetahui kelimpahan, sebaran, dan arsitektur sarang rayap tanah M. gilvus di Cagar Alam Yanlappa, Jawa Barat, serta ukuran populasinya di kawasan konservasi tersebut.
Manfaat Penelitian
Ruang Lingkup Penelitian
II. TINJAUAN PUSTAKA
Habitat Rayap Tanah
Hutan alam di daerah tropika merupakan habitat alami dari rayap tanah
genus Macrotermes. Tanah dengan kandungan bahan organik yang tinggi di
hutan tropis merupakan habitat yang sangat disukai oleh rayap ini (Lavelle &
Spain 2001). Demikian pula iklim yang hangat dengan curah hujan sekitar
1500-3000 mm/th, kelembaban sekitar 80-90%, suhu ambient antara 28-29 o
Habitat alami rayap tanah Macrotermes natalensis adalah di savanna
Afrika. Rayap jenis ini sama dengan rayap dari genus Macrotermes lainnya yaitu
menyukai daun dan ranting yang telah lapuk. Selain itu, rayap jenis ini sangat
menyukai kondisi iklim dengan curah hujan tidak terlalu tinggi yaitu antara
600-700 mm/th, tanah tipe granitik dan ketinggian tempat 81-375 m dpl (Mayer et al.
2001). Namun demikian, pada rayap tanah Macrotermes michaelseni yang juga
menyukai habitat di daerah savanna di Afrika dengan ketinggian 92-368 m dpl
menyenangi curah hujan yang sangat rendah yaitu hanya 200 mm/th, tipe tanah
pasir kemerahan dengan spesifikasi vegetasi rumput savanna (Turner 2000). C dan
keanekaragaman jenis tumbuhan yang sangat tinggi sangat mendukung
keberadaan rayap jenis ini. Hal tersebut menjadikan rayap jenis ini mendapatkan
bahan makanan berupa serasah, kayu dan ranting yang banyak mendukung
kehidupannya (Traniello & Leuthold 2000).
Keberadaan rayap tanah pada suatu area salah satunya tergantung pada
tipe tanah yang terdapat pada areal tersebut (Meyer et al. 2003). Secara umum
rayap tanah lebih menyukai tipe tanah yang banyak mengandung liat. Serangga
ini tidak menyukai tanah berpasir karena tipe tanah ini memiliki kandungan bahan
organiknya rendah (Traun dan Perry 2000). Di samping itu, konstruksi liang
kembara rayap tanah yang terbuat dari tanah liat akan lebih kokoh (Evans
2003).
Sarang rayap tanah utamanya terbuat dari tanah, kotoran, dan serasah
yang berfungsi sebagai tempat memperbanyak koloni dan melindungi koloni dari
lingkungan ekstrim. Kehidupan di dalam sarang inilah yang menyebabkan rayap
berhasil hidup di daerah tropik atau subtropik. Hal ini disebabkan terdapat sistem
pengendalian iklim mikro yang dibuat rayap di dalam sarang untuk
mempertahankan kehidupannya (Noirot & Darlington 2000). Sarang sebagai
memerlukan tipe tanah dan topografi tertentu. Topografi dengan tipe tanah
bergelombang dan tutupan tajuk yang terbuka merupakan habitat yang disukai
oleh rayap tanah Macrotermes natalensis dan Macrotermes bellicosus di Afrika
Selatan (Holt & Lepage 2000; Meyer et al. 2001).
Biologi Macrotermes gilvus
Krishna dan Weesner (1969) menyatakan rayap dapat diklasifikasikan ke
dalam 6 famili yang meliputi : Mastotermitidae, Kalotermitidae, Hodotermitidae,
Rhinotermitidae, Serritermitidae dan Termitidae. Rayap tanah Macrotermes
termasuk ke dalam famili Termitidae sub famili Macrotermitinae. Menurut
Nandika et al. (2003), Macrotermes gilvus merupakan rayap tanah yang banyak
tersebar di Indonesia. Klasifikasi rayap tanah M. gilvus sebagai berikut :
phylum : Arthropoda kelas : Insecta sub-klass : Pterigota ordo : Blattodea famili : Termitidae sub-famili : Macrotermitinae genus : Macrotermes
species : Macrotermes gilvus Hagen
Termitidae adalah famili terbesar diantara famili yang lain. Hal ini
disebabkan 80% anggota ordo Blattodea termasuk dalam famili ini (Ruelle
(1989). Menurut Khrishna & Weesner (1969) rayap hidup dalam kelompok sosial
dengan sistem kasta yang berkembang sempurna. Kasta rayap terdiri dari 3
yaitu kasta reproduktif yang terdiri dari ratu dan raja, kasta prajurit dan kasta
pekerja.
Di dalam koloni, rayap tanah M. gilvus terdapat polimorfisme pada kasta
prajurit dan kasta pekerja. Pada kasta prajurit dikenal prajurit mayor dan kasta
prajurit minor. Sementara pada kasta pekerja dikenal kasta pekerja mayor dan
pekerja minor (Miura & Matsumoto 1995).
Ciri-ciri kasta prajurit M. gilvus Hagen memiliki kepala yang berwarna
coklat tua. Mandibel berkembang dan berfungsi, mandibel kanan, dan kiri
simetris dan tidak memiliki gigi marginal. Mandibel melengkung pada ujungnya
dan digunakan untuk menjepit. Ujung dari labrum tidak jelas, pendek dan
melingkar. Labrum ini memiliki hyallin pada ujungnya. Antena terdiri dari 16-17
ruas. Ada dua jenis kasta prajurit dari M. gilvus Hagen yaitu kasta prajurit yang
besar (mayor) dan kasta prajurit yang kecil (minor) dengan ciri-ciri sebagai
1. Kasta prajurit mayor
Kepala berwarna coklat kemerahan, dengan lebar 2,88 - 3,10 mm. Panjang
kepala dengan mandibel 4,80 - 5,00 mm. Antena 17 ruas, ruas ketiga sama
panjang dengan ruas kedua, ruas ketiga lebih panjang dari ruas keempat.
2. Kasta prajurit minor
Kepala berwarna coklat tua, dengan lebar 1,52 - 1,71 mm, panjang kepala
dengan mandibel 3,07 - 3,27 mm, panjang kepala tanpa mandibel 1,84 - 2,08
mm. Antena 17 ruas, ruas kedua sama panjang dengan ruas keempat.
Kasta pekerja merupakan anggota yang sangat penting dalam koloni
rayap. Tidak kurang dari 80-90% populasi dalam koloni rayap merupakan
individu-individu kasta pekerja. Kasta pekerja umumnya berwarna pucat dengan
kutikula hanya sedikit mengalami penebalan sehingga tampak menyerupai nimfa.
Kasta reproduktif terdiri dari betina (ratu) yang tugasnya bertelur dan jantan
(raja) yang tugasnya membuahi betina. Kasta ini dibedakan menjadi kasta
reproduktif primer dan kasta reproduktif suplementer atau neoten. Kasta
reproduktif primer terdiri atas serangga-serangga dewasa yang bersayap dan
merupakan pendiri koloni. Kasta reproduktif dari Macrotermes dapat
menghasilkan telur setelah satu minggu sejak melakukan swarming (Harris
1971). Menurut Berenbaum (1996), neoten muncul segera setelah kasta
reproduktif primer mati atau terpisah karena pemisahan koloni. Neoten dapat
terbentuk beberapa kali dalam jumlah besar sesuai dengan perkembangan
koloni. Selanjutnya, neoten menggantikan fungsi kasta reproduktif primer untuk
perkembangan koloni. Secara lengkap, pembagian kasta dalam koloni rayap
(A) (B)
(C) (D)
(E) (F)
Ekologi Macrotermes gilvus
Faktor lingkungan yang mempengaruhi perkembangan populasi rayap
meliputi curah hujan, suhu, dan kelembaban. Faktor-faktor tersebut saling
berinteraksi dan saling mempengaruhi satu sama lain. Kelembaban dan suhu
merupakan faktor yang secara bersama-sama mempengaruhi aktivitas rayap.
Perubahan kondisi lingkungan menyebabkan perubahan perkembangan,
aktivitas, dan perilaku rayap.
Curah Hujan
Curah hujan umumnya memberikan pengaruh fisik secara langsung pada
kehidupan koloni rayap, khususnya yang membangun sarang di dalam ataupun
di permukaan tanah (Pearce 1997). Pada rayap genus Macrotermes, memiliki
toleransi curah hujan yang lebar antara 250-3000 mm/th (Meyer et al. 2001). Hal
ini disebabkan rayap jenis ini memiliki sistem mikroklimat di dalam sarang.
Curah hujan merupakan faktor lingkungan yang berguna untuk mendorong
keluarnya kasta reproduksi (laron) dari dalam sarang (Bignell & Eggleton 2000).
Laron tidak akan keluar jika curah hujan rendah. Namun demikian, curah hujan
yang terlalu tinggi dapat juga menurunkan aktivitas wilayah jelajah rayap (Krebs
& Davis 1989).
Kelembaban
Rayap tanah Macrotermes gilvus memerlukan kelembaban yang tinggi.
Perkembangan optimumnya dicapai pada kisaran kelembaban 75-90% (Evans
2003). Perubahan kelembaban sangat mempengaruhi aktivitas jelajah rayap.
Pada kelembaban yang rendah, rayap bergerak menuju daerah dengan suhu
yang lebih rendah. Namun demikian, rayap memilki kemampuan untuk menjaga
kelembaban di dalam liang-liang kembaranya sehingga tetap memungkinkan
rayap bergerak ke daerah yang lebih kering (Holt & Lepage 2000). Di pihak lain,
jika permukaan air tanah rendah, serangga ini hanya sedikit dipengaruhi oleh
Kadar Garam
Beberapa rayap memiliki toleransi terhadap kondisi garam dibanding
dengan rayap yang lain. Genus Coptotermes heimi lebih toleran dibanding
Odontotermes obesus dan Microcerotermes championi. Sementara itu rayap
genus Macrotermes cenderung tidak toleran terhadap kadar garam (contoh:
CaCl, MgCl). Rayap pada umumnya tidak ada dalam kondisi kadar garam yang
ekstrim. Hal ini disebabkan wilayah dengan kadar garam yang tinggi cenderung
memiliki tipe vegetasi tertentu sehingga jenis rayap yang dapat hidup hanya
jenis-jenis tertentu pula (Pearce 1997).
Suhu
Suhu merupakan faktor penting yang mempengaruhi kehidupan
serangga, baik terhadap perkembangan maupun aktivitasnya. Pengaruh suhu
terhadap serangga terbagi menjadi beberapa kisaran. Pertama suhu maksimum
dan minimum, yaitu kisaran suhu terendah atau tertinggi yang dapat
menyebabkan kematian serangga; kedua adalah suhu aestivasi dan hibernasi,
yaitu kisaran suhu di atas atau suhu di bawah suhu optimum yang dapat
mengakibatkan serangga mengurangi aktivitasnya atau dorman; dan ketiga
adalah kisaran suhu optimum. Pada sebagian besar serangga suhu optimumnya
adalah 15-38o
Menurut Lisa dan Conacher (2000), suhu dan kelembaban juga
mempengaruhi kondisi vegetasi yang pada gilirannya mempengaruhi rayap di
sekitarnya. Di tempat terbuka dimana sinar matahari langsung menyinari
permukaan tanah pada tengah hari hingga awal sore hari ketika suhu berada
pada puncaknya, rayap sering berada di bawah tanah atau berada di dalam
sarang (Wood & Lee 1971). Namun mereka tetap dapat berada di permukaan
tanah bila terdapat naungan yang besar yang menciptakan suhu optimum
(thermal shadow).
C (Pearce 1997).
Mekanisme pengaturan suhu pada sarang rayap dapat dilakukan dengan
beberapa cara: Pertama dengan cara isolasi, yaitu dengan membangun sarang
yang tebal, gudang makanan, dan ruangan lain di sekitar sarang. Dengan isolasi
ini, suhu sarang menjadi terkontrol dan transfer panas dari luar ke dalam sarang
diperlambat. Kedua, pengaturan suhu dengan cara mengatur termoregulasi.
dan mampu dikendalikan oleh rayap. Ketiga dengan mempertahankan
kandungan air tanah penyusun sarang (Bonabeau 2000). Pada rayap
Macrotermes metabolisme makanan yang dikumpulkan dari kebun jamur (fungus
comb) mampu menghasilkan karbondioksida, panas dan air. Panas yang
dihasilkan dapat memelihara suhu sarang sehingga suhu dapat dipertahankan
pada kisaran optimum, yaitu 29-32o
Sumber Makanan
C (Korb dan Aanen 2003).
Distribusi sumber makanan dalam habitat rayap berhubungan dengan
ukuran sarang dan wilayah jelajah. Jika rayap memerlukan sumber makanan
yang lebih, maka akan memperluas wilayah jelajahnya dengan mencari sumber
makanan baru untuk kebutuhan koloninya. Wilayah yang kering merupakan
wilayah yang sedikit sumber makanan dibandingkan dengan wilayah yang
lembab.
Komponen yang dapat digunakan untuk memprediksi wilayah jelajah dan
tingginya populasi antara lain 1). Komponen lingkungan yaitu tipe tanah, curah
hujan/sistem irigasi dan sumber makanan, 2). Komponen dalam koloni rayap
yaitu peningkatan ukuran populasi, peningkatan migrasi seperti laron serta
aktivitas foraging (Pearce 1997).
Sarang Rayap Tanah Macrotermes
Model sistem perkembangan yang alami dari suatu organisme
merupakan informasi penting untuk mempelajari kehidupan organisme tersebut.
Konsep biologi dalam arsitektur adalah adaptasi, homeostatis, kondisi darurat
dan sistem pengaturan sendiri kemungkinan dapat diterapkan dalam aplikasi
arsitektur (Linardou 2008).
Arsitektur dan tata ruang sarang rayap M. gilvus akan menghasilkan
suatu mekanisme konstruksi yang mendukung sistem pengaturan energi,
penyimpanan makanan, serta penyempurnaan sarang untuk penyesuaian
terhadap lingkungan luar. Bentuk dan fungsi tata ruang yang kompleks di dalam
sarang M. gilvus menunjukkan bahwa arsitektur sarang mengadaptasi dari
lingkungan luar. Bentuk bangunan sarang rayap M. gilvus adalah bentuk kubah
(dome) yang kokoh. Sistem ventilasi dalam sarang M. gilvus adalah sistem
M. gilvus memiliki sistem tata ruang unik, merupakan bentuk adaptasi dari cuaca
yang ekstrim dan mencegah serangan oleh organisme predator (Ladley &
Bullock 2005).
Pada rayap tanah jenis Macrotermes jeanneli di Afrika memiliki arsitektur
sarang yang menjulang tinggi keatas dengan ketinggian mencapai puluhan
meter. Sarang rayap tanah M. jeanneli berbentuk cerobong dengan posisi
puncak cenderung kearah sinar matahari (building thermotaxis). Sarang rayap
ini memiliki sistem ventilasi terbuka (open ventilation system). Hal ini berkaitan
dengan upaya adaptasi rayap tersebut dengan lingkungannya (Leuthold et al.
2004).
Hal yang unik dari struktur dan arsitektur sarang rayap genus
Macrotermes adalah koloni rayap ini mampu membentuk iklim mikro di dalam
sarang yang bersifat stabil yaitu 30oC dan kandungan CO2 yang tidak lebih dari
1,5 ppm (Noirot & Darlington 2000). Di dalam pembangunan sarang rayap
Macrotermes terdapat pola pembangunan berkelanjutan. Hal ini dapat dimengerti
karena proses pembangunan sarang rayap Macrotermes berlangsung secara
terus menerus tanpa berhenti. Proses pembangunan sarang tidak pernah
membongkar bangunan sebelumnya, tetapi melanjutkan proses pembangunan
sebelumnya. Efisiensi energi (homeostatis) terjadi di dalam proses mekanisme
konstruksi pembangunan sarang. Hal ini dapat dibuktikan dengan pengamatan
jumlah ventilasi yang dibuat dalam sarang disesuaikan dengan jumlah anggota
koloni di dalamnya. Sementara itu tanah yang digunakan untuk membangun
sarang berasal dari dalam sarang dibawa oleh kasta pekerja mayor menuju ke
Abstrak
Penelitian ini bertujuan untuk mengetahui kelimpahan dan sebaran sarang rayap M. gilvus di Cagar Alam Yanlappa, Jawa Barat. Untuk mengetahui keberadaan sarang rayap M. gilvus di lokasi penelitian dibuat transek dengan arah Barat Laut ke Tenggara dari batas patok cagar alam. Jarak antar transek 50 meter dan total panjang transek 6.800 meter. Setiap sarang M. gilvus yang ditemukan ditentukan koordinatnya dengan Geophisical Position System (GPS). Sarang rayap diklasifikasikan kedalam tiga tipe berdasarkan ukurannya, yaitu sarang kecil (tinggi sarang ≤ 0,49 m), sarang sedang (tinggi sarang 0,5-0,99 m), dan sarang besar (tinggi sarang ≥ 1 m). Kemiringan lereng dan indeks tutupan tajuk (Leaf Area Index, LAI) di sekitar rayap M. gilvus juga dicatat. Disamping itu, dilakukan analisis vegetasi dengan membuat plot pengamatan berukuran 20 x 20 m untuk tingkat pohon, 10 x 10 m untuk tingkat tiang, 5 x 5 m untuk tingkat semai dengan intensitas pengambilan contoh sebesar 0,38%. Hasil penelitian menunjukkan bahwa pola sebaran dan kelimpahan sarang rayap adalah berkelompok (cluster) dengan mengikuti kemiringan lereng (3-8%) dan penutupan tajuk terbuka (LAI 0-1) hingga terbuka ringan (LAI 1-2). Sementara itu tapak sarang tidak terkait dengan tipe vegetasi di permukaan tanah.
Kata Kunci: Pemetaan, GPS, habitat, Macrotermes gilvus Hagen.
Abstract
A study was conducted to find out the distribution pattern and number of subterranean termite M. gilvus based on the spatial, combined with physical factors (height, slope, soil type) and biotic factors (vegetation and Leaf Area Index) which significantly influence on M. gilvus. The data collected included the geographic position mound of M. gilvus by using the GPS, mound distribution patterns, slope, Leaf Area Index (LAI) which is determined with hemipot method by taking photographs with fisheyecamera for the ground cover from above. The mound of subterranean termite are classified into the types based on size namely, small mounds (≤ 0,49 m tall), medium mounds (0,5 -0,99 m tall), and large moundss (≥ 1 m tall). In addition, a vegetation was also analysed with a plot method of 20 x 20 m for trees, 10 x 10 m for small trees, and 5 x 5 m for seedling. The results of this study showed that the mounds distribution pattern is grouped following the slopes of 3-8% and open canopy coverage of LAI 0-2.
Pendahuluan
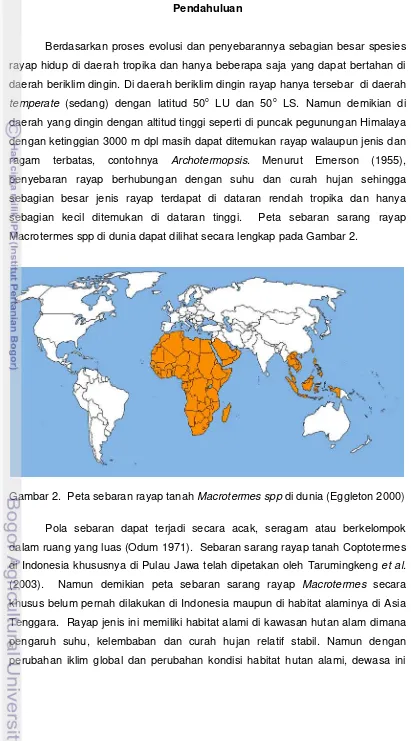
Berdasarkan proses evolusi dan penyebarannya sebagian besar spesies rayap hidup di daerah tropika dan hanya beberapa saja yang dapat bertahan di daerah beriklim dingin. Di daerah beriklim dingin rayap hanya tersebar di daerah temperate (sedang) dengan latitud 50o LU dan 50o LS. Namun demikian di daerah yang dingin dengan altitud tinggi seperti di puncak pegunungan Himalaya dengan ketinggian 3000 m dpl masih dapat ditemukan rayap walaupun jenis dan ragam terbatas, contohnya Archotermopsis. Menurut Emerson (1955), penyebaran rayap berhubungan dengan suhu dan curah hujan sehingga sebagian besar jenis rayap terdapat di dataran rendah tropika dan hanya sebagian kecil ditemukan di dataran tinggi. Peta sebaran sarang rayap Macrotermes spp di dunia dapat dilihat secara lengkap pada Gambar 2.
Gambar 2. Peta sebaran rayap tanah Macrotermes spp di dunia(Eggleton 2000)
memungkinkan telah terjadi perubahan sebaran sarangnya. Informasi ini belum dieksplorasi secara rinci dan dalam.
Pada hutan tropik, rayap memiliki keragaman yang tinggi. Hal ini disebabkan pada kawasan hutan alam memiliki ekosistem yang beragam (Indrawan et al. 2007). Ada beberapa alasan tingginya keragaman ekosistem mempengaruhi keragaman rayap. Pertama, spesies tropik dapat lebih tenggang terhadap relung ekologi yang tumpang tindih, dengan menganggap relung itu sebagai corak ragam lingkungan mikro pada habitat tropik yang majemuk, terutama di dalam hutan. Hal ini berdampak pada daerah tropik banyak memberikan kesempatan pembentukan spesies dibandingkan pada iklim sub tropik. Kedua, tersedianya sumberdaya yang melimpah, terutama dalam hal pakan. Ketiga, banyaknya pemangsaan dan parasitisme dalam lingkungan tropik cenderung membatasi berlimpahnya spesies tertentu dan mempersulit spesies tertentu untuk menambah besaran populasinya. Dengan demikian lebih banyak jumlah spesies yang menghuni habitat itu dibandingkan dengan kelimpahannya (Odum 1971).
hutan hujan tropis seperti Indonesia dengan vegetasi hutan yang heterogen jumlah sarangnya adalah > 4 sarang/ha (Vongkaluang et al. 2006), sementara di Afrika terutama pada tipe ekosistem savanna dengan vegetasi yang homogen memiliki jumlah sarang < 4 sarang/ha (Schuurman 2000; Gromadzki 2003).
Faktor lingkungan yang utama mempengaruhi sebaran sarang rayap antara lain temperatur dan kelembaban, sementara itu faktor lain yang mendukung adalah curah hujan, struktur tanah dan vegetasi (Cookson & Trajstman 2002). Hal ini dapat dimengerti, karena rayap adalah serangga yang memiliki kulit tipis yang rentan terhadap proses dehidrasi oleh angin/udara kering sehingga rayap membutuhkan kelembaban yang stabil. Sebagai contoh keberadaan rayap Reticulitermes sp (Rhinotermitidae) di Florida sangat dipengaruhi oleh faktor suhu, kelembaban dan sifat tanah. Suhu berperan dalam sebaran dan aktivitas rayap saat mencari makan. Bilamana suhu permukaan tanah terlalu panas atau terlalu dingin rayap tidak melakukan foraging. Suhu ideal bagiReticulitermes spadalah 15-35 o
Rayap Macrotermes merupakan rayap yang banyak tersebar di Asia Tenggara terutama banyak ditemukan di Indonesia, Malaysia, Singapura, Philiphina dan Thailand (Tho 1992), namun sebaran sarang berdasarkan garis lintang (latitude) dan ketinggian (altitude) tempat belum pernah dilakukan. Oleh karena itu informasi mengenai sebaran spasial dari rayap Macrotermes ini di habitat alaminya penting untuk diketahui karena belakangan rayap jenis ini telah tersebar diluar habitat alaminya.
C. Adapun kelembaban mendekati RH 100%, terutama pada daerah kering, memberikan kondisi yang baik pada rayap jenis ini. Tanah liat merupakan tipe tanah yang baik bagi kehidupannya karena pada tanah jenis ini saat hujan cenderung menahan kelembaban (Suiter et al. 2000). Pada rayap yang hidup di daerah subtropis, jenis-jenis kayu yang lebih disukai adalah kayu lunak (seperti : redwood, cypress, juniper) sedangkan kayu keras dan kayu-kayu yang mengandung banyak senyawa protektan kurang disukai karena bersifat toksik bagi rayap. Menurut Yoshimura et al. (2003) rayap subtropik lebih menyukai bahan berselulosa yang telah terdegradasi dengan urutan kesukaan sebagai berikut : serat selululose > kertas > kayu lapis > balok kayu lunak > balok kayu keras.
analog. Belakangan ini dengan perkembangan teknologi digital dan tata ruang maka telah berkembang sistem pemetaan menggunakan Sistem Informasi Geografis (SIG). SIG merupakan seperangkat fungsi dengan kemampuan canggih, yang dapat digunakan oleh para profesional untuk : menyimpan, menampilkan, dan memanipulasi/mengoreksi data geografis/spasial. Sistem ini memiliki beberapa keunggulan antara lain : (1) Dapat dilakukan pada seluruh kemampuan pemetaan konvensional; (2) Dapat menentukan jarak tertentu dari titik atau garis; (3) Dengan analisis overlay, dapat digunakan lebih dari satu peta tematik dan menghasilkan coverage/peta baru; dan (4) Adanya visualisasi 3 dimensi yang dapat dikombinasikan dengan citra satelit. Dari keunggulan yang telah dijelaskan, peta spasial sebaran sarang rayap tanah Macrotermes gilvus pada penelitian ini akan menggunakan metode SIG karena akan lebih bermanfaat dibandingkan dengan pemetaan secara konvensional. Selain itu, hasilnya secara cepat dapat memvisualisasikan data sebaran sarang spasial organisme target.
Penelitian ini bertujuan untuk mengetahui pola sebaran sarang rayap M. gilvus Hagen dan faktor-faktor yang mempengaruhinya yaitu kemiringan lereng, tutupan tajuk, dan tipe tanah di Cagar Alam Yanlappa, Jawa Barat.
Bahan dan Metode
Waktu dan Tempat Penelitian
Penelitian dilakukan bulan April 2007 - Oktober 2009 di Cagar Alam Yanlappa, Jawa Barat dengan ketinggian (50-150 mdpl) dan luas kawasan 32 Ha. Analisis sebaran spatial dilakukan di Laboratorium PPLH–IPB, analisis LAI (Leaf Area Index) dilakukan di Laboratorium Konservasi Hutan-IPB, identifikasi rayap dilakukan di Laboratorium Biologi Hasil Hutan PPSHB–IPB, sementara itu sebagai pembanding dilakukan identifikasi rayap Macrotermes di Natural History Museum, London.
Prosedur Penelitian
alam dengan interval lebar 50 meter dan panjang sampai batas patok wilayah cagar alam. Setiap titik awal jalur pengamatan diberi tanda patok dengan arah lintasan pengamatan menggunakan kompas. Data yang dikumpulkan meliputi posisi sarang rayap Macrotermes menurut GPS, pola sebaran sarang, data kemiringan lereng yang diperoleh dengan menggunakan Abneylevel, data Leaf Area Index (LAI) yang diperoleh dengan metode hemipot yaitu dengan cara mengambil foto menggunakan kamera fisheye dibawah lantai hutan tepat diatas sarang rayap. Sarang rayap yang ditemukan diklasifikasikan kedalam tiga tipe berdasarkan ukurannya yaitu sarang kecil (tinggi sarang ≤ 0,49 m), sarang sedang (tinggi sarang 0,5-0,99 m), dan sarang besar (tinggi sarang ≥ 1 m). Disamping itu, dilakukan analisis vegetasi pada 3 area yaitu area dengan kelimpahan sarang padat (5 sarang/ha), area dengan kelimpahan sarang sedang (3 sarang/ha), dan area dengan kelimpahan sarang rendah (tidak ada sarang) dengan membuat transek 20 x 20 m untuk tingkat pohon, 10 x 10 m untuk tingkat tiang, 5 x 5 m untuk tingkat semai pada masing-masing wilayah kelimpahan sarang (Surasana 1990).
Posisi sarang rayap yang ditemukan dibuat “overlay”nya (metode tumpang tindih) dengan menggunakan software ArcView GIS 3.3 dan ERDAS Imagine 8.5, Sementara itu pengolahan LAI dilakukan dengan software Hemiview 2.1. Variabel vegetasi yang diamati meliputi jumlah famili, spesies, individu, nilai kerapatan mutlak (KM), frekuensi mutlak (FM) dan dominasi mutlak (DM). Index Nilai Penting (INP) setiap spesies dihitung menurut Cox (1978). Pola sebaran sarang spasial dari sarang rayap Macrotermes ditentukan dengan mengukur indeks penyebaran (dispersion index). Penentuan indeks dispersi (ID), dengan formula sebagai berikut:
Pola penyebaran spasial dari data sarang yang dikumpulkan diuji menggunakan uji Square untuk menentukan tipe penyebaran. Uji Chi-Square yang digunakan untuk N<30, adalah persamaan λ2
1. Jika λhit2 < λ2 0.975
2. Jika λ
, maka pola sebaran sarang yang terjadi adalah seragam.
2
0.975 ≤ λhit2 ≤ λ2 0.025
3. Jika λhit
, maka pola sebaran sarang yang terjadi adalah acak.
2 > λ2
0.025, maka pola sebaran sarang yang terjadi adalah
Hasil Penelitian
Sebaran Sarang Rayap Tanah M. gilvus
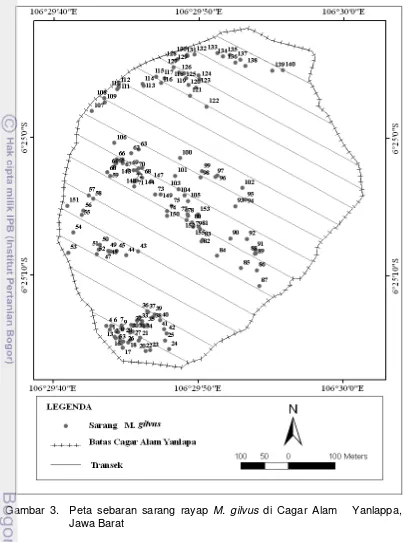
Hasil penelitian menunjukkan bahwa di Cagar Alam Yanlappa, Jawa Barat dengan luas 32 Ha ditemukan 155 tapak sarang dengan rincian 56 sarang tipe besar (tinggi ≥ 1 m), 94 sarang tipe sedang (tinggi 0,5 – 0,99 m), dan 5 sarang tipe kecil (tinggi ≤ 0,49 m). Sarang -sarang tersebut menyebar secara berkelompok karena λ2 hitung > λ2 0.025 (Lampiran 4). Posisi keberadaan sarang
Gambar 3. Peta sebaran sarang rayap M. gilvus di Cagar Alam Yanlappa, Jawa Barat
kelas. Penentuan kelas ketinggian menjadi 2 kelas berdasarkan rentang penyebaran sarang rayap tanah M. gilvus (50-100 m dpl dan 100-150 m dpl). Jumlah sarang rayap M. gilvus pada masing-masing kelas ketinggian, nilai LAI dan kemiringan lereng disajikan pada Tabel 1.
Tabel 1. Kelimpahan sarang rayap tanah M. gilvus pada kelas ketinggian, nilai LAI, dan kemiringan lahan di Cagar Alam Yanlappa
Kelas ketinggian Luas area (m2) Jumlah sarang Kelimpahan (sarang/Ha) 50-100 m dpl 258.124 75 3 100-150 m dpl 80.835 80 10
Leaf Area Index (LAI)
0-1 75.260 128 2 1-2 35.790 27 8 >2 227.911 0 0 Kemiringan lereng (%)
3-8 288.026 155 5 > 8 110.935 0 0
Data pada Tabel 1 di atas mengungkapkan bahwa jumlah dan densitas sarang rayap tanah M. gilvus berada paling banyak pada kelas ketinggian 100-150 m dpl dibandingkan dengan ketinggian 50-100 m dpl. Hal ini dapat dikatakan bahwa sarang M. gilvus menyukai ketinggian antara 100-150 m dpl. Selain itu pada ketinggian 100-150 m dpl kondisinya bergelombang, sehingga rayap M. gilvus lebih menyukai tipe topografi tersebut. Densitas sarang per hektar adalah 3 sarang untuk kelas 50-100 m dpl dan 10 sarang untuk kelas 100-150 m dpl.Foto penutupan tajuk hasil pengambilan gambar hemipot disajikan pada Gambar 4.
(A) (B)
Nilai LAI semakin besar menunjukkan bahwa penutupan tajuk semakin tertutup, sehingga radiasi matahari ke bawah tajuk semakin kecil dan kelembaban dibawah tajuk semakin meningkat. Nilai dari Leaf Area Index (LAI) merupakan presentasi dari penutupan kanopi pada areal dibawah tajuk yang diproyeksikan secara vertikal. Nilai ini digunakan untuk memprediksi besarnya cahaya dan kelembaban dengan asumsi jika kanopi semakin tertutup maka cahaya yang masuk ke lantai hutan semakin kecil dan kelembabannya akan semakin tinggi.
Nilai LAI di Cagar Alam Yanlappa dapat diklasifikasikan dalam 2 kelas yaitu kelas terbuka dan kelas tertutup ringan (Tabel 1). Berdasarkan hasil pengambilan data dan pengolahan data maka dapat diketahui bahwa sarang rayap tanah M. gilvus Hagen di Cagar Alam Yanlappa terletak pada tempat yang memiliki nilai LAI 0,211-1,722. Jumlah sarang M. gilvus lebih banyak pada kelas dengan nilai LAI 0-1 yakni 128 sarang sedangkan jumlah lebih kecil yaitu 27 sarang pada nilai 1-2, sedangkan pada nilai (LAI > 2) tidak ditemukan sarang. Densitas sarang per hektar adalah 2 sarang untuk kelas LAI 0-1 dan 8 sarang untuk kelas LAI 1-2.
Sama halnya dengan ketinggian, kemiringan lereng pun merupakan salah satu faktor habitat yang berpengaruh terhadap keberadaan rayap (Traniello & Leuthold 2000), sehingga kelerengan merupakan salah satu variabel yang diuji dalam penelitian ini. Berdasarkan hasil pengolahan kontur, kemiringan lereng di Cagar Alam Yanlappa dibagi menjadi 2 kelas yaitu 3-8 % dan >8 %.
Berdasarkan hasil pengolahan data dapat diketahui bahwa penyebaran rayap tanah M. gilvus di Cagar Alam Yanlappa adalah sebagai berikut: pada kelas dengan kemiringan lereng 3-8% terdapat 155 jumlah sarang, kelas kemiringan lereng >8% tidak dijumpai sarang. Semua sarang rayap M. gilvus terdapat pada kelas 3-8% dengan luasan 288.026 m2
Tanah merupakan tubuh alam yang terbentuk pada permukaan bumi yang dapat menunjang pertumbuhan tanaman dengan sifat yang dihasilkan dari interaksi antara bahan induk, iklim, organisme dan waktu (Purwowidodo 1998).
Berdasarkan peta satuan lahan daerah kawasan hutan alam provinsi Jawa Barat dan Banten yang dibuat oleh pusat penelitian tanah dan agroklimat Badan Penelitian dan Pengembangan Pertanian tahun 1991, dan peta digital Bakorsurtanal 2006 diketahui bahwa seluruh kawasan Cagar Alam Yanlappa memiliki tipe tanah podsolik merah kekuningan, sehingga sarang rayap M. gilvus menyebar pada tanah tipe tersebut.
Hasil analisis vegetasi menunjukkan bahwa dari areal pengamatan 1200 m2
Berdasarkan komposisi famili, maka pada daerah dimana kelimpahan sarang rayapnya padat pada tingkat pohon dan tiang didominasi oleh famili Myrtaceae, pada tingkat pancang Arecaceae (INP = 50%), dan pada tingkat semai Lycopodiaceae (INP = 32,13%). Pada area dengan kelimpahan sarang sedang, pada tingkat pohon didominasi oleh famili Moraceae (INP = 57,41%), sedangkan untuk tiang, pancang dan semai masing-masing didominasi oleh famili Ebenaceae (INP = 49,59%), Euphobiaceae (INP = 28,69%), Araceae (INP = 82,22%). Secara lengkap, indeks nilai penting tertinggi dari famili pohon, tiang, pancang dan semai dapat dilihat pada Lampiran 10.
terdapat area sarang rayap dengan kelimpahan padat (8 sarang/ha), spesies pohon yang dominan adalah Artocarpus elastica (INP = 34,33%), tiang (Syzygium zollingerianum = 109,64%), pancang (Glycosmis pentaphylla =
Pembahasan
Pola sebaran sarang rayap tanah M. gilvus di Cagar Alam Yanlappa, cenderung membentuk pola sebaran sarang berkelompok (cluster), dengan pembentukan 3 cluster yaitu cluster dengan kelimpahan sarang padat, kelimpahan sedang dan tidak ada sarang (Gambar 4). Hal ini menunjukkan bahwa sebaran sarang rayap tanah M. gilvus sangat unik. Posisi keberadaan sarang rayap M. gilvus di Cagar Alam Yanlappa memanjang dari arah Barat Laut ke Tenggara. Posisi tersebut diduga berkaitan dengan sifat cryptobiotik (tidak menyukai sinar) pada rayap. Dalam membangun sarang, terlihat bahwa posisi sebaran sarang M. gilvus dari Barat Laut ke Tenggara adalah wilayah yang terhindar dari sinar matahari secara langsung dari Timur dan Barat. Sistem bangunan seperti ini sangat cocok didirikan pada daerah tropis yang memiliki suhu panas dan kelembaban yang tinggi. Posisi bangunan seperti ini memiliki fungsi antara lain dapat mengurangi pemanasan matahari serta memanfaatkan angin agar terjadi pendinginan karena penguapan.
Parameter abiotik yang diamati dalam penelitian antara lain ketinggian tempat, jenis tanah, elevasi kelerengan, dan penutupan tajuk. Dari empat parameter tersebut terlihat bahwa elevasi kelerengan dan penutupan tajuk sangat berpengaruh pada pola sebaran sarang. Rayap M. gilvus sangat menyukai tutupan tajuk yang terbuka dengan kelas Leaf Area Index 0-1 dan elevasi kelerengan kelas 3-8%. Hasil yang sama juga dilaporkan oleh Meyer et al. (2000) yang mengatakan bahwa sebaran sebaran sarang rayap M. natalensis di Kruger National Park, Afrika Selatan sangat dipengaruhi oleh topografi tipe bergelombang dengan ketinggian kawasan 250-400 m dpl dan warna tanah merah dan kuning (Meyer et al. 1999).
Sinar matahari mutlak diperlukan oleh organisme hidup dalam suatu ekosistem. Namun demikian, suhu, radiasi sinar matahari dan kelembaban merupakan faktor pembatas bagi kehidupan rayap M. gilvus (Donovan et al. 2007).
Gangguan terhadap hutan yang berupa penebangan pohon akan mengakibatkan terbukanya tajuk pohon. Terbukanya tajuk pohon akan menyebabkan terjadinya perubahan faktor lingkungan seperti suhu udara, penguapan, kelembaban dan intensitas cahaya matahari pada ekosistem hutan tersebut (Indrawan 2000). Pada hutan sangat terganggu yang didominasi oleh tumbuhan bawah memberikan indikasi bahwa struktur vegetasinya merupakan struktur vegetasi dengan penutupan tajuk terbuka dan jarang (Alcaraz & Avilla 2000). Nilai LAI pada hutan yang sangat terganggu diperkirakan rendah dengan tutupan tajuk terbuka.
Simpulan
Abstrak
Untuk mengetahui struktur populasi pada sarang rayap M. gilvus dilakukan pembongkaran sarang secara bertahap dan hati-hati baik secara vertikal maupun horisontal pada setiap tipe sarang (masing-masing 3 sarang). Pembongkaran dilakukan sampai kedalaman 1 m di bawah permukaan tanah. Namun apabila kedalaman sarang contoh lebih dari 1 m pembongkaran dilanjutkan hingga seluruh anggota koloni terambil. Setelah dilakukan pembongkaran, sarang contoh ditutup dengan menggunakan plastik berwarna gelap selama ± 3 jam untuk memberikan waktu rayap yang sedang foraging kembali ke sarang. Pengambilan spesimen dilanjutkan sampai tidak ada lagi rayap tersisa di titik sarang. Hasil penelitian menunjukkan bahwa jumlah total individu rayap tanah M. gilvus di Cagar Alam Yanlappa adalah 17.423.695 individu dengan rincian Sarang tipe besar dihuni 183.825 ± 7742 individu, sarang tipe sedang dihuni 46.267 ± 2073 individu dan sarang tipe kecil dihuni 20.223 ± 1919 individu. Jumlah nimfa pada sarang tipe besar dan tipe sedang pada umumnya besar (82,5%). Hal ini diduga sarang sedang dan besar berada pada fase perkembangan koloni yang sangat pesat. Ciri lain dari koloni sarang sedang dan sarang besar adalah ditemukannya kasta reproduktif sekunder (neoten). Biomassa rayap M. gilvus paling besar berturut-turut adalah kasta prajurit mayor, kasta prajurit minor dan nimfa. Biomassa terkait dengan peran masing-masing kasta dalam koloni. Semakin besar ukuran sarang maka biomassa anggota setiap kastanya akan berukuran lebih besar.
Kata kunci: Ukuran populasi, biomasa, pembongkaran, M. gilvus Hagen
Abstract
A study was conducted on subterranean termites Macrotermes gilvus Hagen population structure, by carefully and subcequently demoliting the mounds vertically and horizontally for the large, medium, and small mounds. Observation of the termite population size was carried out by the demolition of the mounds little by little and carefully both vertically and horizontally in each type of the mounds (respectively 3 mounds). The demolition was carried out in until the depth of 1 m below land surface. However if the sample mounds is more than 1 m deep, the demolition it was continued as far as all the colony member was taken. After demolition, the mounds was coverred by using dark coloured plastic for 3 hours, to give time of termite that was foraging returned to the mounds. Sampling was continued by using the vacuum cleaner until all termites finished in the mounds. Specimen counting was carried out in each caste of each type of the mounds. The results of observation of population size showed that there are three mounds groups namely: large mounds with population size of 183.825 ± 7742 individuals, medium mounds with 46.267 ± 2073 individuals, and small mounds with 20.223 ± 1919 individuals. The main characteristics of medium and large mounds are the nymphs are very big, which indicate that these mounds are in the best development stage of the colony.
Pendahuluan
Di Indonesia ditemukan tidak kurang dari 300 jenis rayap (Tarumingkeng 2000). Dari jenis yang ada ternyata yang paling banyak menimbulkan kerusakan pada bangunan dan perkebunan adalah golongan rayap tanah (subterranean termites), anggota dari famili Termitidae dan famili Rhinotermitidae (Nandika 2003). Rayap Macrotermes termasuk dalam famili termitidae. Namun kerusakan yang ditimbulkan belum sebanyak jenis lainnya. Kemampuan merusak serangga tersebut erat hubungannya dengan ukuran populasi, daya jelajah, dan daya adaptasinya terhadap lingkungan. Besarnya ukuran populasi seringkali menyebabkan upaya pengendaliannya menjadi sangat sukar (Su 2002).
Struktur populasi sarang merupakan kelompok-kelompok yang terdapat dalam sarang rayap termasuk didalamnya ukuran populasi dan biomassanya. Ukuran yang terdapat dalam populasi tersebut, antara lain Ukuran populasi koloni akan bervariasi menurut status tingkatan rayap, jenis spesies, ukuran sarang, habitat, mikroklimat (kelembaban, suhu, curah hujan), jenis tanah dan umur ratu (Krishna & Weesner 1969). Pada umumnya ukuran populasi dalam satu koloni rayap tingkat rendah (yaitu ditandai dari adanya hubungan mutualisme dengan bakteri) memiliki ukuran populasi lebih besar dibandingkan dengan rayap tingkat tinggi (hubungan mutualisme tidak dengan bakteri tetapi dengan organisme protozoa) (Wood & Lee 1971). Sebagai contoh ukuran populasi dalam satu koloni rayap tingkat rendah Coptotermes curvignathus dapat mencapai sebesar 1.536.190-1.834.400 ekor (Diba & Nandika 1999), sementara itu pada rayap tingkat tinggi (famili Kalotermitidae) seperti Kalotermes flavicollis hanya 600 - 1000 individu per koloni. Walaupun demikian ukuran populasi tingkat tinggi lainnya seperti Macrotermes di Afrika ada yang kisarannya lebih luas yaitu 5.000 - 200.000 individu per sarang (Lepage & Darlington 2000).
tidak produktif lagi apabila telah terjadi fragmentasi koloni. Ratu dapat terbentuk beberapa kali sesuai dengan perkembangan koloni. Dalam satu sarang pada kondisi normal hanya terdiri dari satu ratu dan satu raja. Ratu dibuahi oleh raja. Telur-telur hasil pembuahan tersebut akan berkembang menjadi nimfa yang kemudian berkembang menjadi kasta-kasta (pekerja, prajurit, dan kasta reproduktif). Krishna dan Weesner (1969) melaporkan bahwa koloni rayap Macrotermes diduga berjumlah satu juta individu (berbagai kasta) dalam satu sarang dan menghasilkan kasta reproduktif di luar ratu dan raja yaitu berupa laron sebanyak 60.000 setahun. Ukuran koloni Trinervitermes geminatus (Termitidae) di savanna Afrika Barat diduga antara 19.000 sampai 52.000 individu/sarang dan rata-rata menghasilkan laron sebanyak 950 ekor per koloni (Lepage & Darlington 2000). Beberapa peneliti lain melaporkan bahwa persentase jumlah laron yang dihasilkan dalam satu koloni rayap adalah sebagai berikut : 30% (Pterotermes occidentis; Famili Kalotermitidae); 14-16% (Paraneotermes simplicicornis; Famili Kalotermitidae); 39% (Zootermopsis laticeps; famili Termopsidae); 17,5% (Stolotermes victoriensis Hill; Famili Termopsidae) dan 28-43,3% (Odontotermes obesus; Famili Termitidae) dari ukuran populasi rayap tersebut (Pearce 1997).
Meskipun ukuran populasi rayap tanah sulit dipelajari mengingat hampir semua aktivitas serangga ini dilakukan dibawah permukaan tanah namun penelitian dalam bidang tersebut telah berkembang dengan menggunakan berbagai macam metode baru. Sebagai contoh untuk mengetahui ukuran populasi dapat dilakukan dengan menggali lempeng-lempeng rayap atau dengan menggunakan tracer radioisotop. Hal ini telah digunakan untuk mempelajari ukuran populasi rayap tanah di Amerika Serikat dan Thailand (Meyer et al. 2001).
ukuran populasi koloni rayap tanah Macrotermes gilvus Hagen (Blattodea : Termitidae) di komunitas hutan alam.
Bahan dan Metode
Waktu dan Tempat Penelitian
Penelitian dilakukan pada bulan Desember 2006 sampai dengan Agustus 2007 pada areal pengamatan seluas ± 32 Ha. Pemilihan areal pengamatan didasarkan tingginya kepadatan sarang rayap tanah Macrotermes gilvus Hagen di areal pengamatan setelah dilakukan sensus sarang rayap M. gilvus.
Prosedur Penelitian
1. Penghitungan Koloni
(A) (B)
(C) (D)
Gambar 23. Kondisi salah satu sarang rayap tanah M. gilvus pada saat penghitungan kelimpahan populasi: sarang utuh (A), sarang yang telah dibongkar (B), sarang yang telah dibongkar ditutup plastik untuk beberapa hari, agar memberi kesempatan anggota koloni yang sedang diluar sarang (foraging) kembali ke sarang (C) dan individu rayap dan bagian sarang ditampung dalam wadah untuk proses penghitungan kelimpahan populasi (D).
2. Biomasa Rayap
Rayap tanah Macrotermes gilvus Hagen dikoleksi sebanyak 300 ekor untuk setiap kasta (kasta pekerja, kasta prajurit mayor, kasta prajurit minor, dan nimfa) pada setiap tipe sarang (masing-masing 3 sarang). Setelah dilakukan pemisahan, dilakukan penghitungan berat basah, berat kering dan rasio biomassa.
3. Analisis Data
Hasil Penelitian
A. Ukuran Populasi
Ukuran populasi rayap tanah Macrotermes gilvus Hagen pada masing masing sarang dari tiga kali ulangan didapat hasil bahwa sarang ukuran besar penghuninya 10 kali lebih besar dari sarang ukuran kecil, sedangkan sarang ukuran sedang hanya empat kalinya. Perbandingan antar kasta terlihat bahwa pada sarang ukuran kecil dan sarang ukuran besar konsisten, sedangkan pada sarang ukuran sedang jumlah kasta prajurit mayornya sangat besar. Sarang ukuran kecil didominasi oleh kasta pekerja, sementara itu pada sarang ukuran sedang dan sarang ukuran besar didominasi oleh nimfa (Tabel 4). Berdasarkan uji ANOVA diperoleh F>Fcrit dan P value
Tabel 4. Jumlah individu dalam koloni pada berbagai tipe sarang di Cagar Alam Yanlappa, Jawa Barat
<<α=0,05, sementara itu pada uji korelasi dihasilkan nilai sebesar 92,92% artinya terdapat hubungan yang erat antara jumlah total individu dengan ukuran sarang (Lampiran 8).
Kasta
Tipe sarang P Mayor P.Minor Pekerja Nimfa Total
Kecil 1.297 ± 179 261 ± 91 10.196 ± 1455 8.468 ± 210 20.223 ± 1919
Sedang 4.021 ± 289 906 ± 573 3.167 ± 706 38.173 ± 645 46.267 ± 2073
Besar 2.964 ± 158 360 ± 423 29.277 ± 1,83 151.233 ± 8,24 183.825 ± 7742
Adanya polimorfisme pada kasta prajurit berupa terbentuknya dua bentuk prajurit yang berbeda yaitu prajurit mayor dan prajurit minor. Beda keduanya adalah secara morfologi kasta prajurit mayor lebih besar dibandingkan prajurit minor. Pada kasta prajurit mayor kepala berwarna coklat kemerahan, dengan lebar kepala 2,80-3,10 mm, panjang kepala dari mandibel 4,8-5,0 mm, ruas ketiga antenna lebih panjang dari ruas keempat. Sementara itu, pada kasta prajurit minor kepala berwarna coklat tua dengan lebar kepala 1,52-1,71 mm, panjang kepala dari mandibel 1,84-2,08 mm. Ruas antena kedua sama panjang dengan ruas antena keempat.