KUALITAS NUTRIEN DAN KECERNAAN IN VITRO
HIJAUAN FERMENTASI (Hi-Fer
) RUMPUT GAJAH
DENGAN PENAMBAHAN INOKULAN ATAU
CRUDE ENZYME ASAL Trichoderma reesei
DEA JUSTIA NURJANA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Kualitas Nutrien dan Karakteristik In vitro Hijauan Fermentasi (Hi-Fer) Rumput Gajah dengan Penambahan Inokulan atau Crude Enzyme Asal Trichoderma reesei adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2016
Dea Justia Nurjana
iv
Ketersediaan hijauan yang berkualitas dan berkelanjutan diperlukan untuk usaha peternakan ruminansia. Teknologi pengawetan hijauan yang tepat dapat mengatasi keterbatasan hijauan di musim kemarau. Hijauan fermentasi (Hi-fer) merupakan teknologi pengawetan hijauan melalui proses fermentasi anaerob dengan menggunakan cairan aditif fermentasi dan dikemas dalam kantong plastik kedap udara. Peningkatan nilai guna Hi-fer dapat dilakukan dengan memanfaatkan aktivitas kapang tanah, yaitu Trichoderma reesei (T. reesei). Kapang T. reesei memproduksi enzim selulase cukup banyak dan dapat tumbuh pada pH 4 – 6. Penelitian ini bertujuan untuk mengevaluasi pengaruh inokulan T. reesei atau crude enzyme asal T. reesei terhadap kualitas nutrien Hi-fer rumput gajah, kualitas fermentasi Hi-fer, kecernaan dan produk fermentasi rumen in vitro.
Cairan rumen berasal dari sapi Peranakan Ongole fistula yang berada di Lembaga Ilmu Pengetahuan Indonesia, Cibinong, Bogor. Kapang T. reesei QM6a diperoleh dari IPB Culture Collection (IPBCC), Bogor. Kajian kualitas fermentasi Hi-fer rumput gajah menggunakan rancangan acak lengkap dengan tiga perlakuan dan enam ulangan. Kajian fermentasi rumen menggunakan rancangan acak kelompok dengan tiga perlakuan dan enam ulangan. Waktu pengambilan cairan rumen sebagai ulangan dan kelompok. Perlakuan untuk kedua rancangan penelitian ini adalah (T0) kontrol Hi-fer rumput gajah, (T1) Hi-fer rumput gajah+ inokulan T. reesei sebanyak 2.13 x 107 cfu/kg, (T2) Hi-ferrumput gajah + crude enzyme T. reesei sebanyak 11.4 unit/kg. Hi-fer disimpan pada suhu ruang dan dibuka setelah 21 hari fermentasi. Peubah yang diamati adalah karakteristik fisik (warna, aroma, tekstur, dan keberadaan jamur), komposisi nutrien Hi-fer, karakteristik fermentasi Hi-fer (pH Hi-fer, asam laktat, asam asetat, asam propionat, asam butirat, dan NH3), dan karakteristik fermentasi
rumen (pH rumen, degradasi bahan kering dan organik, koefisien cerna bahan kering dan organik, konsentrasi volatile fatty acid total dan parsial, NH3 dan
estimasi produksi gas metan). Data yang diperoleh diuji dengan sidik ragam (ANOVA) dan dilakukan uji Duncan untuk data yang berbeda nyata.
kandungan protein kasar (PK) dan total digestible nutrient (TDN) Hi-fer. Sementara perlakuan penambahan inokulan T. reesei hanya berpengaruh (P<0.01) menurunkan kandungan NDF Hi-fer. Kandungan lemak kasar (LK), bahan ekstrak tanpa nitrogen (BETN), acid detergent fiber (ADF), selulosa, lignin dan hemiselulosa tidak dipengaruhi (P>0.05) oleh perlakuan.
Perlakuan penambahan inokulan T. reesei nyata meningkatkan (P<0.05) pH Hi-fer, konsentrasi asam asetat dan asam propionat Hi-fer. Perlakuan penambahan crude enzyme berpengaruh meningkatkan (P<0.05) asam asetat dan asam propionat. Kandungan asam laktat, asam butirat dan NH3 tidak dipengaruhi
(P>0.05) oleh perlakuan. Perlakuan inokulan T. reesei dan crude enzyme tidak berpengaruh (P>0.05) terhadap nilai fleigh, populasi bakteri asam laktat, dan kecernaan dan kecernaan dan produk fermentasi rumen in vitro. Perlakuan crude enzyme menurunkan (P<0.05) degradasi NDF in vitro. Estimasi produksi gas metan tidak dipengaruhi (P>0.05) oleh perlakuan inokulan T. reesei dan crude enzyme. Berdasarkan penelitian ini dapat disimpulkan bahwa semua perlakuan menghasilkan Hi-fer rumput gajah berkualitas baik dengan nilai pH Hi-fer
yang rendah, tinggi konsentrasi asam laktat dan rendah konsentrasi asam butirat. Penambahan crude enzyme lebih meningkatkan kualitas nutrien Hi-fer rumput gajahdengan menurunkan kandungan NDF dan meningkatkan TDN. Penambahan inokulan T. reesei dan crude enzyme menurunkan degradasi NDF in vitro namun belum berpengaruh terhadap degradasi dan kecernaan bahan kering dan bahan organik serta produk fermentasi rumen.
Kata kunci: crude enzyme, in vitro, hijauan fermentasi, rumput gajah,
vi
SUMMARY
DEA JUSTIA NURJANA. Nutrient Quality and In vitro Digestibility of Napier Grass Fermented Forage (Hi-fer) by Addition Inoculant or Crude Enzyme from
Trichoderma reesei. Supervised by SURYAHADI and SRI SUHARTI.
The qualified and sustainable forages availability is needed in the ruminant production. An appropriate forage preservation technology is needed to overcome the lack of forage in dry season. Fermented forage (Hi-fer) is a forage preservation technology through ensiling process added with special fermentation additive and packed in airtight plastic bags. To improve fermentation quality and digestibility of Hi-fer, it can be done by using Trichoderma reesei (T. reesei)
activity. T. reesei is as cellulase producer and can grow at pH 4 – 5. This study was aimed to to evaluate the effect of T. reesei inoculant or crude enzyme derived from T. reesei on nutrient quality of fermented forage, fermentation quality of fermented forage, dry matter and organic matter digestibility and rumen fermentation product determined by in vitro.
The rumen fluid was collected 2 h after morning feeding from three fistulated Ongole Crossbred beef cattle’s in Indonesia Institute of Science, Cibinong, Bogor. Trichoderma reesei QM6a was collected from IPB Culture Collection (IPBCC), Bogor. Experimental design for fermentation characteristics of Hi-fer napier grass used a completely randomized design with three treatments and six replications. Experimental design for rumen fermentation characteristics of Hi-fer used a randomized block design with three treatments and six replications. Collect time of rumen fluid as replication and block. The treatments were (T0) control Hi-fer napier grass, (T1) Hi-fer napier grass +
T.reesei inoculant at level 2.13 x 107 cfu/kg, and (T2) Hi-fer napier grass + crude enzyme from Trichoderma reesei at level 11.4 unit/kg. Fermented forages were kept at room temperature and opened 21 days after fermentation. Variables observed were physical characteristics (color, odor, texture, and visible molds), nutrient composition, fermentation characteristics of Hi-fer (pH Hi-fer, lactic acid, acetic acid, propionic acid, butyric acid, and NH3) and rumen fermentation
characteristics (ruminal pH, dry matter degradability, organic matter degradability, NDF degradability, dry matter digestibility, organic matter digestibility, total and molar proportion of VFA, NH3 and also estimation of methane production). Data
were analyzed using the analysis of variance (ANOVA) and the differences between means were compared using Duncan Multiple Range Test.
(P<0.01) NDF content of Hi-fer . Treatments did not affected (P>0.05) crude fat, nitrogen free extract (NFE), acid detergent fiber, cellulose, lignin and hemicellulose contents of Hi-fer.
Based on fermentation characteristics of Hi-fer, the addition of T. reesei
inoculant significantly increased (P<0.05) pH value, acetic acid, propionic acid of Hi-fer. Addition of crude enzyme only increased (P<0.05) propionic acid content of Hi-fer. There were no significant differences in lactic acid, butyric acid, and NH3 contents of Hi-fer. The addition of T. reesei inoculant and crude
enzyme did not affect (P>0.05) fleigh point, lactic acid bacteria population, digestibility and rumen fermentation product determined by in vitro. The addition of crude enzyme decreased (P<0.05) degradability NDF in vitro. Estimation of methane productions did not differ (P>0.05) among treatments. It was concluded that all treatments produced well-preserved napier grass fermented forage with low pH, high concentration of lactic acid and low concentration of butyric acid. The addition of crude enzyme more improved nutrient quality of napier grass fermented forage with decreasing NDF and increasing TDN. The addition of T. reesei inoculant and crude enzyme decreased degradability NDF in vitro but has not affected degradability and digestibility of dry matter and organic matter and also rumen fermentability products.
viii
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KUALITAS NUTRIEN DAN KECERNAAN IN VITRO
HIJAUAN FERMENTASI (Hi-Fer
) RUMPUT GAJAH
DENGAN PENAMBAHAN INOKULAN ATAU
CRUDE ENZYME ASAL Trichoderma reesei
DEA JUSTIA NURJANA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
x
xii
PRAKATA
Puji syukur kehadirat Allah SWT atas rahmat, hidayah dan ridho-Nya sehingga penulis dapat menyelesaikan studi dan tesis yang berjudul “Kualitas Nutrien dan Kecernaan In vitro Hijauan Fermentasi (Hi-Fer) Rumput Gajah dengan Penambahan Inokulan atau Crude Enzyme Asal Trichoderma reesei”. Sebagian hasil penelitian ini sedang dalam proses penerbitan di jurnal ilmiah
Media Peternakan dengan judul “Improvement of Fermented Forage Nuritive
Value Using Inoculant and Crude Enzyme from Trichoderma reesei and Its Effect on In vitro Rumen Fermentation”.
Penulis berterima kasih kepada Dr Ir Suryahadi, DEA dan Dr Sri Suharti, SPt MSi selaku komisi pembimbing atas waktu, bimbingan dan masukan dalam penyelesaian tesis ini. Ucapan terima kasih juga disampaikan Dr Ir Muhammad Ridla, MAgr selaku penguji luar komisi atas kritik dan sarannya, Prof Dr Ir Toto Toharmat MAgr Sc atas bimbingannya, Dr Ir Dwierra Evvyernie A, MS MSc, Prof Dr Yuli Retnani, MSc selaku ketua program studi INP, staf program studi INP (Mas Supri dan Bu Ade) atas bantuan pengurusan administrasinya, Ibu Dian Anggraeny dan Ibu Adriani atas bantuan dan bimbingan di laboratorium, mba Saprilian, Lilis, CENTRAS, Umul, Infitria, Nining, Ardi dan teman-teman seperjuangan INP 2013, INP 2012, anggota laboratorium Biokimia Fisiologi dan Mikrobiologi Nutrisi dan laboratorium Nutrisi Ternak Perah, teman-teman kos Cibanteng atas motivasi, bantuan dan kebersamaan yang telah terjalin serta kepada semua pihah yang telah membatu selama penelitian. Ungkapan terima kasih tak terhingga penulis sampaikan kepada kedua orang tua (Bapak Sumarno dan Ibu Eti), adik-adik tersayang (Qori dan Sofa) serta seluruh keluarga besar atas nasihat, doa, bantuan dan motivasinya.
Semoga tesis ini bermanfaat dan dapat dijadikan referensi dalam bidang pertanian dan peternakan Indonesia.
Bogor, Februari 2016
DAFTAR ISI
DAFTAR TABEL xiv
DAFTAR GAMBAR xiv
DAFTAR LAMPIRAN xiv
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan 3
Manfaat 3
2 METODE 3
Lokasi dan Waktu Penelitian 3
Materi 3
Metode 4
Rancangan Percobaan 9
3 HASIL DAN PEMBAHASAN 10
Uji Aktivitas Crude enzyme asal Trichoderma reesei 10 Karakteristik Fisik Hi-fer Rumput Gajah 10 Komposisi Nutrien Hi-fer Rumput Gajah 11 Karakteristik Fermentasi Hi-ferRumput Gajah 14 Nilai Fleigh dan Populasi Asam Laktat Hi-ferRumput Gajah 16 Karakteristik Fermentasi Rumen 17
4 SIMPULAN 20
DAFTAR PUSTAKA 21
LAMPIRAN 25
xiv
DAFTAR TABEL
1 Karakteristik fisik Hi-fer rumput gajah setelah 21 hari fermentasi 11 2 Komposisi nutrien rumput gajah dan Hi-fer rumput gajah setelah 21
hari fermentasi 12
3 Karakteristik fermentasi Hi-fer rumput gajah setelah 21 hari
fermentasi 14
4 Nilai fleigh dan populasi bakteri asam laktat Hi-fer rumput gajah 17 5 Karakteristik fermentasi rumen 17
DAFTAR GAMBAR
1 Warna Hi-fer rumput gajah sebelum proses fermentasi 10 2 Warna Hi-fer rumput gajah setelah proses fermentasi selama 21 hari 11
DAFTAR LAMPIRAN
1 Hasil sidik ragam komposisi nutrient Hi-fer rumput gajah 26 2 Hasil uji lanjut Duncan perlakuan terhadap bahan kering Hi-fer
rumput gajah 27
3 Hasil uji lanjut Duncan perlakuan terhadap bahan organik Hi-fer
rumput gajah 27
4 Hasil uji lanjut Duncan perlakuan terhadap protein kasar Hi-fer
rumput gajah 27
5 Hasil lanjut Duncan perlakuan terhadap serat kasar Hi-fer rumput
gajah 27
6 Hasil uji lanjut Duncan perlakuan terhadap TDN Hi-fer rumput
gajah 28
7 Hasil uji lanjut Duncan perlakuan terhadap NDF Hi-fer rumput
gajah 28
8 Hasil sidik ragam karakteristik fermentasi Hi-fer Hasil sidik ragam
karakteristik fermentasi Hi-fer rumput gajah 28 9 Hasil uji lanjut Duncan perlakuan terhadap pH Hi-fer rumput gajah 29 10 Hasil uji lanjut Duncan perlakuan terhadap asam asetat Hi-fer
rumput gajah 29
11 Hasil uji lanjut Duncan perlakuan terhadap asam propionat Hi-fer
rumput gajah 29
12 Hasil sidik ragam nilai fleigh dan populasi bakteri asam laktat
Hi-ferrumput gajah 29
13 Hasil sidik ragam pH, degradasi bahan kering/bahan organik,
14 Hasil uji sidik ragam total VFA, proporsi molar VFA, gas metana
dan amonia 31
1
1 PENDAHULUAN
Latar Belakang
Ketersediaan hijauan pakan secara kontinyu dan berkualitas baik sangat diperlukan pada usaha peternakan ruminansia. Pemberian hijauan yang cukup untuk ternak ruminansia sangat penting untuk mengoptimalkan fungsi rumen yang berkaitan dengan kesehatan dan produksi ternak. Perubahan musim berpengaruh terhadap ketersediaan hijauan. Ketika musim hujan ketersediaan hijauan melimpah, sementara masalah kelangkaan hijauan sering terjadi di musim kemarau. Keterbatasan hijauan pakan berkualitas dapat mengakibatkan produksi ternak ruminansia tidak optimal. Pemanfaatan teknologi pengewatan hijauan diperlukan untuk menangulangi hal tersebut.
Hijauan fermentasi (Hi-fer) merupakan teknologi pengawetan hijauan melalui proses fermentasi dengan menggunakan cairan aditif fermentasi dan dikemas dalam kantong plastik pada kondisi anaerob (Suryahadi 2014). Prinsipnya seperti proses ensilase dimana proses pengawetan hijauan terjadi secara fermentasi anaerob dan bakteri asam laktat (BAL) yang ada di hijauan akan memfermentasikan gula menjadi asam organik terutama asam laktat (McDonald
et al. 1991). Proses ensilase menurunkan pH melalui proses homofermentasi sebagian besar water soluble carbohydrates (WSC) menjadi laktat sehingga mikroorganisme perusak dapat dihambat dan kandungan nutrien bisa terjaga (Sun
et al. 2009).
Teknologi Hi-fer merupakan teknologi pengawetan hijauan yang dikembangkan oleh Pusat Studi Hewan Tropika (Centras) LPPM-IPB. Kecepatan dan efisiensi fermentasi anaerob bergantung pada jenis dan populasi BAL yang ada di hijauan dan kandungan WSC (Amanullah et al. 2014, McDonald et al. 1991) sehingga penambahan cairan aditif pada Hi-fer dapat meningkatkan kualitas fermentasi. Komponen aktif aditif fermentasi terdiri atas growth promotor
bagi mikroba, palatability enhancer, mineral dari asam-asam organik dan antioksidan. Penambahan aditif fermentasi bertujuan untuk mempercepat proses fermentasi anaerob sehingga Hi-fer dapat segera digunakan, meningkatkan palatabilitas Hi-fer yang dihasilkan dan meningkatkan daya simpan (Suryahadi 2014). Proses pengawetan pada Hi-fer terjadi karena BAL menggunakan WSC yang tersedia dengan cepat sehingga penurunan pH terjadi secara cepat dan dapat menjaga nutrient dan menghambat pertumbuhan mikroorganisme perusak.
Selama ini Hi-ferdiarahkan pada pengawetan, penyimpanan, pengemasan dan peningkatan palatabilitas bahan pakan. Perlu adanya upaya untuk meningkatkan kecernaan pakan, sehingga proses pembuatan Hi-fer diharapkan selain untuk mengawetkan juga dapat meningkatkan kualitas bahan pakan. Proses ensilase hijauan tropis cukup sulit karena hijauan tropis memiliki kandungan serat kasar yang tinggi, kandungan bahan kering yang rendah, rendahnya kandungan protein kasar dan konsentrasi WSC (Avellaneda et al. 2009; Bureenok et al. 2012). Akibatnya kecernaan hijauan tropis cukup rendah. Enzim fibrolitik, molasses dan glukosa telah digunakan sebagai aditif untuk meningkatan kualitas silase (Dean et al. 2005; Li et al. 2014). Peningkatan kualitas fermentasi dan kecernaan Hi-fer
2
Trichoderma reesei (T. reesei) merupakan kapang yang dapat
mensekresikan enzim selulase cukup banyak (Juhász et al. 2005). Potensi T. reesei menghasilkan enzim selulase dapat bermanfaat dalam mendegradasi dinding sel tumbuhan sehingga akan meningkatkan kecernaanya. Kapang T. reesei dapat tumbuh pada sekitar pH 4 – 7 (Bailey et al. 1993), sehingga diharapkan dapat bekerja dalam proses pembuatan Hi-fer. Sementara produksi enzim selulase oleh T. Reesei terjadi pada pH 4 – 5 (Bailey et al. 1993, Juhász et al. 2004). Inokulan T. reesei diharapkan dapat bekerja di awal fermentasi Hi-fer., dimana masih terdapat sedikit oksigen sehingga T. reesei dapat memanfaatkan substrat yang ada untuk memproduksi selulase dan mempercepat kondisi anaerob. Ketika kondisi fermentasi menjadi anaerob, enzim selulase yang diproduksi dapat meningkatkan degradasi serat tanaman yang belum terdegradasi oleh T. reesei
menjadi WSC sebagai substrat yang dapat dimanfaatkan BAL.
Pemanfaatan aktivitas enzim T. reesei dapat pula dilakukan melalui pembuatan crude enzyme. Penggunaan enzim bergantung pada aktivitas enzim dan pH. Colombatto et al. (2004) menyatakan beberapa enzim fibrolitik lebih stabil bekerja pada suhu rendah dan pH asam. Penggunaan aditif crude enzyme
dapat digunakan untuk meningkatkan kualitas nutrisi silase. Hidayat et al. (2005) melaporkan bahwa penambahan crude enzyme asal Trichoderma viride dapat meningkatkan kecernaan bahan kering dan bahan organik silase jerami padi. Ketika enzim fibrolitik ditambahkan sebelum proses ensilase, dinding sel tanaman akan terhidrolisis menjadi substrat yang mudah difermentasi dan dapat dimanfaatkan oleh BAL sehingga meningkatkan kualitas fermentasi silase (Dehghani et al. 2012). Proses tersebut akan meningkatan asam laktat dan menurunkan pH sehingga menghambat proses proteolysis dari aktivitas bakteri selain BAL dan enzim tanaman (Li et al. 2014).
Pembuatan crude enzyme selulase membutuhkan substrat sebagai media tumbuh mikroorganisme. Ketika T. reesei dikultivasi dengan substrat selulosa dan lignoselulosa sebagai sumber karbon dapat menghasilkan enzim seperti selulase, amilase, hemiselulase ligninase, peptidase dan proteinase (Adav et al. 2012; Saloheimo & Pakula 2012). Jerami padi mengandung selulosa, hemiselulosa dan lignin. Kandungan jerami padi tersebut dapat dimanfaatkan sebagai substrat dalam pembuatan crude enzyme.
Penggunaan crude enzyme asal T. reesei dalam pembuatan silase masih jarang dilakukan. Kinerja crude enzyme tentunya lebih rendah dibandingkan dengan enzim komersial, tetapi biaya produksinya lebih murah. Sementara itu, penggunaan inokulan T. reesei pada Hi-fer diharapkan enzim selulase dapat disekresikan dan mendegradasi dinding sel tanaman menjadi substrat yang dapat dimanfaatkan BAL.
Beradasarkan uraian tersebut, penggunaan inokulan T. reesei atau crude enzyme asal T. reesei pada Hi-fer rumput gajah diharapkan dapat meningkatkan kualitas nutrient dan fermentasi Hi-fer rumput gajah. Pada penelitian ini juga dilakukan evaluasi pengaruh penggunaan inokulan T. reesei atau crude enzyme
3
Tujuan
Penelitian ini bertujuan mengevaluasi pengaruh inokulan T. reesei atau
crude enzyme asal T. reesei terhadap kualitas nutrien Hi-fer rumput gajah, kualitas fermentasi Hi-fer, kecernaan dan produk fermentasi rumen in vitro.
Manfaat
Manfaat penelitian ini diharapkan dapat memanfaatkan inokulan kapang
Trichoderma reesei dan crude enzyme asal Trichoderma reesei sebagai bahan aditif untuk meningkatkan kualitas nutrisi Hi-ferrumput gajah. Selain itu, dapat memperkenalkan Hi-fersebagai hijauan awet fermentasi yang praktis.
2 METODE
Lokasi dan Waktu Penelitian
Penelitian ini dilaksanakan di Laboratorium Biokimia, Fisiologi dan Mikrobiologi Nutrisi, Laboratorium Nutrisi Ternak Perah, Laboratorium Ilmu dan Teknologi Pakan, Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor. Analisa kandungan nutrien silase dilakukan di Pusat Penelitian Sumberdaya Hayati dan Bioteknologi, IPB. Analisa fraksi serat silase dilakukan di Laboratorium Balai Penelitian Ternak, BPPT Bogor. Analisa proporsi molar volatile fatty acid (VFA) di Laboratorium Kimia Pusat Studi Pangan dan Gizi, Universitas Gadjah Mada, Yogjakarta. Penelitian dilaksanakan pada bulan November 2014 sampai Juni 2015.
Materi Alat
Peralatan yang digunakan adalah chopper, timbangan, oven 60°C, oven 105°C, tanur 600°C, autoclave, laminar, jarum ose, tabung rekasi, cawan petri, erlenmeyer, shaker inkubator, sentrifuge, botol scott, pH meter, eksikator, stirrer,
shaker waterbath, termos vakum, pipet volumetik, tabung eppendorf, pompa vakum, lemari es, freezer, cawan porselen, cawan conway, buret, tabung fermentor, tutup karet, vortex, beaker glass dan wadah uap. Alat untuk uji aktivitas enzim adalah spektrofotometer. Alat untuk uji proporsi molar VFA adalah gas chromatography.
Bahan
Bahan pembuatan Hi-fer adalah rumput gajah (Pennisetum purpureum), aditif fermentasi yang diproduksi CENTRAS LPPM, dan kantong plastik
4
Agar (PDA). Penyiapan crude enzyme dibutuhkan jerami padi, NaOH 2%, media nutrisi untuk produksi crude enzyme terdiri dari ekstrak ragi (Oxoid-England),
bacteriological peptone (Oxoid-England), (NH4)2SO4, KH2PO4, FeSO4.7H2O,
larutan CMC 1% dan larutan buffer asetat 0.1 M dengan pH 5.0, larutan salin 0.85%, larutan 1% Tween 80, dan buffer asetat 01 M dengan pH 5.5. Cairan rumen diperoleh dari sapi Peranakan Ongole berfistula di Lembaga Ilmu Pengetahuan Indonesia (LIPI). Uji in vitro dibutuhkan larutan Mc Dougall, larutan pepsin, gas CO2, HgCl2 jenuh, kertas saring Whatman No.41, aquades, asam borat,
Na2CO3 jenuh, dan H2SO4 0.005 N, NaOH 0.5 N, HCl 0.5 N, H2SO4 15%,
indikator PP (Phenolpthaline) dan vaseline. Uji aktivitas enzim FP-ase dibutuhkan kertas saring Whatman No. 1, larutan dinitrosalicylic acid (DNS), 0.05 M Na sitrat pH 5, dan glukosa standar dalam buffer.
Metode
Peremajaan Inokulan Trichoderma reesei (Modifikasi Prabavathy et al. 2006). Biakan murni T.reesei diperbanyak dengan membiakkan ulang di dalam tabung media potatoe dextrose Agar (PDA). Sebanyak 3.9 g PDA dilarutkan dalam 100 ml aquades steril dan diaduk rata. Larutan tersebut dipanaskan hingga seluruhnya larut. Media PDA tersebut dituangkan pada tabung reaksi sebanyak 4 - 5 ml kemudian tabung reaksi ditutup kapas dan disterilkan pada suhu 121 °C selama 15 menit dalam autoclave. Setelah tabung reaksi dingin sekitar 45-50 °C, medium yang masih cair ditempatkan dalam keadaan miring dengan kemiringan 30° dan tidak mengenai kapas. Biakan murni T.reesei ditanam pada media agar miring di dalam laminar dan disimpan pada suhu kamar (28 °C) selama 7 hari. Setelah 7 hari inokulum berupa spora ditambahkan aquades steril sebanyak 10 ml per tabung reaksi. Selanjutnya, spora dipisahkan dengan cara permukaan biakan yang berisi biakan kapang pada agar miring digores-gores dengan penganduk kemudian dikocok dengan vortex agar suspensi spora menjadi homogen dan siap dilakukan perhitungan jumlah koloni.
Pada perhitungan jumlah koloni dipersiapkan media PDA steril dituangkan ke dalam cawan petri. Tabung reaksi yang berisi 5 ml media PDA dipersiapkan sebanyak 7 tabung. Sebanyak 0.5 ml larutan spora dimasukkan ke dalam tabung pengenceran 10-1, kemudian dari tabung tersebut diambil sebanyak 0,5 ml untuk dituangkan pada tabung 10-2. Selanjutnya, tabung pengenceran 10-1 dituangkan dalam cawan petri. Proses tersebut dilakukan sampai pengenceran 10-7. Sebelum pengambilan larutan spora dalam tabung harus dihomogenkan dengan vortex. Cawan petri disimpan pada suhu kamar (28 °C) dan diinkubasi selama 48-72 jam. Pengamatan jumlah koloni dilakukan setiap 6 jam.
⁄
Produksi Crude enzyme asal T.reesei (Modifikasi Anwar et al.2010)
5
Penyiapan Crude enzyme. Kapang T.reesei dikembangbiakkan pada media PDA miring selama 7 hari. Enzim dipersiapkan dengan cara menginkubasi T.reesei
dalam media padat jerami padi dengan larutan nutrisi yang mengandung 1.0 g ekstrak ragi (Oxoid-England), 1.5 g bacteriological peptone (Oxoid-England), 1.4 g (NH4)2SO4, 2.0 g KH2PO4, 0.005 g FeSO4.7H2O, 5 ml larutan CMC 1% dalam
tiap liter larutan buffer asetat 0,1 M dengan pH 5.0.
Sebanyak 5 g serbuk jerami padi dimasukkan ke dalam erlenmeyer 250 ml dan ditambahkan 25 ml larutan nutrisi kemudian ditutup kapas yang dibalut kain kasa. Campuran tersebut disterilisasi pada suhu 121 °C selama 15 menit pada autoclave. Bibit T.reesei dalam agar miring disuspensikan dalam larutan salin 0.85% yang mengandung 0.1% Tween 80. Suspensi spora T.reesei yang mengandung 1.8 x 108 cfu/ml diinokulasikan secara aseptik ke dalam erlenmeyer. Media yang berisi
T.reesei diinkubasi selama 6 hari. Enzim dipanen menggunakan 100 ml larutan 1% Tween 80 dalam buffer asetat 0,1 M dengan pH 5.5 dan diaduk pada 175 rpm selama 135 menit. Selanjutnya campuran enzim disentrifugasi pada 8.000 rpm selama 10 menit dengan suhu 4 °C dan disaring untuk mendapatkan supernatan yang disebut crude enzyme. Pada saat tidak digunakan enzim disimpan pada suhu 4 °C.
Uji Aktivitas Enzim Filter Paper Activity (FPA) (Modifikasi Ghose 1987) Aktivitas enzim FP-ase dapat mencerminkan aktivitas eksoglukanase. Aktivitas enzim dilakukan dengan cara mengukur jumlah glukosa yang dibebaskan dari kertas saring Whatman No. 1 ukuran 1 x 6 cm dari enzim tersebut pada suhu 30 °C dan pH 5. Kadar glukosa yang terbentuk ditetapkan dengan prosedur Ghose (1987) dengan spektrofotometri menggunakan larutan
dinitrosalicylic acid (DNS) dan diukur pada panjang gelombang 550 nm.
Pembuatan Hi-fer (Suryahadi 2014)
Rumput gajah dipanen pada umur 60 hari dan dilayukan semalam. Rumput gajah tersebut dicacah dengan ukuran 5 cm. Rumput gajah yang sudah dicacah ditambahkan 2% (w/v) aditif fermentasi dan 7% (w/v) molasses kemudian dimasukkan ke dalam kantong plastik polyethylene (270 mm x 50 mm x 0.07 mm) sebanyak 2 lapis dan kedap udara. Inokulan dan/atau enzim ditambahkan dengan cara dicampurkan terlebih dahulu pada aditif fermentasi. Dosis inokulan T. reesei
yang ditambahkan pada Hi-fer sebanyak 2.13 x 107 cfu/kg berat segar (Jaelani 2007). Dosis pemberian crude enzyme yang ditambahkan pada Hi-fer sebanyak 11.4 IU/kg berat segar (Hidayat et al. 2005). Tiap kantong berisi 3 kg. Hi-fer
disimpan pada suhu dan di tempat yang terhindar sinar matahari langsung Hi-fer
dibuka setelah 21 hari fermentasi.
Komposisi Nutrien
6
acid detergent fiber (ADF), selulosa dan lignin ditentukan dengan metode Van Soest et al. (1991).
Karakteristik Fisik Hi-fer
Sifat fisik yang diamati meliputi warna, aroma, tektur dan keberadaan jamur. Kriteria warna Hi-fer yaitu baik (hijau terang sampai hijau kecoklatan tergantung materi silase), sedang (hijau kekuningan sampai hijau kecoklatan), dan buruk (hijau kebiruan, abu-abu atau coklat). Kriteria aroma Hi-fer yaitu baik (asam), sedang (sedikit bau tengik), dan buruk (bau amoniak, busuk). Kriteria tektur Hi-fer yaitu baik (kompak, lembut), sedang (lembut, sedikit mudah hancur), dan buruk (berlendir, mudah hancur). Keberadaan jamur dihitung dengan cara membagi bobot silase yang berjamur dengan bobot total silase.
Pengukuran pH Hi-fer
Sampel Hi-fer diambil sebanyak 20 g dan dihomogenkan dengan 200 ml air. Cairan tersebut digunakan untuk pengukuran pH, kandungan asam laktat, kandungan VFA parsial, dan NH3. Nilai pH diukur menggunakan pH meter.
Perhitungan Nilai Fleigh
Nilai fleigh (NF) dihitung berdasarkan rumus perhitungan Idikut et al. (2009).
NF = 220 + (2 x %BK – 15) – (40 x pH)
Perhitungan Koloni Bakteri Asam Laktat (Fardiaz 1992)
Koloni bakteri asam laktat dihitung dengan metode Total Plate Count (TPC). Sampel Hi-fer ditambahkan aquades dengan perbandingan 1:2. Sebanyak 1 ml cairan Hi-fer dimasukkan ke dalam 9 ml aquades kemudian diencerkan dengan mengambil 1 ml lalu dimasukkan ke dalam 9 ml aquades sampai tujuh kali pengenceran. Sebanyak 0.1 ml dari pengenceran ke 6 dan ke 7 diinokulasi pada cawan petri berisi media MRS A. Media agar tersebut kemudian diinkubasi pada suhu ruang selama tiga hari. Koloni yang tumbuh akan berbentuk bulat miring berwarna agak kekuningan. Koloni bakteri asam laktat dihitung dengan rumus:
Populasi fu g⁄ umlah koloni Pengen eran
Pengambilan Cairan Rumen
Cairan rumen diambil dari sapi fistula dua jam setelah ternak makan. Termos vakum yang akan dipakai terlebih dahulu diisi dengan air panas sehingga suhunya mencapai 39 °C kemudian ditutup. Cairan rumen diperas dengan menggunakan kain kasa dan dimasukkan ke dalam termos tersebut. Sebelum digunakan, air panas yang ada di dalam termos dibuang terlebih dahulu. Untuk menjaga agar cairan rumen tetap dalam kondisi anaerob, termos harus segera ditutup rapat dan dialiri gas CO2 sebelum digunakan.
Prosedur Pembuatan Larutan Mc Dougal
7
(3.42 gram), NaCl (2.82 g), MgSO4.7H2O (0.72 g) dan CaCl2 (0.24 g) dimasukkan
dan dilarutkan. Kemudian ditambahkan CaCl2 setelah semua bahan larut. Lalu
leher labu dicuci dengan air destilasi hingga permukaan air mencapai tanda tera. Campuran lalu dikocok dengan gas CO2 secara perlahan-lahan dengan cara
melewatkannya sampai pH larutan mencapai 7.
Prosedur Fermentasi Pakan (Tilley dan Terry 1963)
Tabung fermentor yang telah diisi dengan 0.5 g sampel ransum perlakuan ditambahkan 10 ml cairan rumen dan 40 ml larutan Mc Dougal. Tabung fermentor dikocok dengan cara mengaliri gas CO2 selama 30 detik (pH 6.5-6.9) dan ditutup
dengan karet berventilasi. Tabung dimasukkan ke dalam shaker waterbath dengan suhu 39 °C, dilakukan fermentasi selama 4 jam untuk sampel VFA/NH3, dan
fermentasi 48 jam untuk sampel degradasi bahan kering (DBK)/degradasi bahan organik (DBO) serta koeficien cerna bahan kering (KCBK)/koeficien cerna bahan organik (KCBO). Untuk menghentikan fermentasi tutup karet berventilasi dibuka dan ditetesi 2 tetes HgCl2 jenuh untuk menghentikan aktivitas mikroba.
Pengukuran pH Fermentasi Rumen
Pengukuran pH dilakukan pada sampel inkubasi 4 jam dengan menggunakan pH meter yang telah dikalibrasi. Nilai pH yang diambil yaitu nilai pH yang konsisten.
Pengukuran Konsentrasi Asam Laktat Tertitrasi (AOAC 2005)
Sebanyak 25 ml substrat ditambahkan 2- 3 tetes indikator PP kemudian dititrasi dengan 0.1 M NaOH secara perlahan sampai terlihat warna merah muda. Setiap ml 0.1 NaOH setara dengan 90.08 mg asam laktat.
Total asam laktat tertitrasi (mg/ml) = (ml NaOH x N NaoH x 90.08)/ml sampel
Pengukuran VFA Hi-fer dan Rumen (General Laboratory Procedure 1966) Pengukuran konsentrasi VFA total dan parsial (asetat, propionat dan butirat) menggunakan gas chromatography (GC 8A, Shimadzu Crop., Kyoto, Japan, Capillary, dengan kolom berisi 10% SP-1200, 1% H3PO4 on 80/100 Cromosorb WAW dan nitrogen sebagai gas carrier). Sampel VFA yang digunakan berasal dari cairan Hi-fer dan supernatan yang dihasilkan dari proses fermentasi dengan inkubasi 4 jam yang diambil sebanyak 1.5 ml ke dalam tabung eppendorf dan ditambahkan H2SO4. Sampel dianalisa dengan menginjekkan 0.4 μl sampel pada
gas chromatography.
Pengukuran NH3 Hi-fer dan Rumen (General Laboratory Procedure 1966)
Pengukuran konsentrasi NH3 menggunakan metode Mikrodifusi Conway.
8
Setelah itu cawan Conway digoyang-goyangkan hingga supernatan dan NaCO3
tercampur rata, dan dibiarkan dalam suhu ruang selama 24 jam. Setelah 24 jam asam borat berindikator dititrasi dengan H2SO4 0.005 N sampai terjadi perubahan
warna dari biru menjadi merah. Kemudian konsentrasi NH3 dihitung berdasarkan
rumus:
Konsentrasi NH3 = [ml H2SO4 x N H2SO4 x 1000] mM
Pengukuran Koeficien Cerna Bahan Kering (KCBK) dan Bahan Organik (KCBO) (Tilley & Terry 1963)
Setelah diinkubasi selama 48 jam, ditambahkan 2-3 tetes HgCl2 jenuh untuk
menghentikan aktivitas mikroba. Lalu disentrifuge dengan kecepatan 3000 rpm selama 15 menit dan supernatannya dibuang, kedalam tabung ditambahkan 50 ml larutan pepsin-HCl 0.2%. Pencernaan enzimatis berlangsung aerob selama 48 jam. Hasil pencernaan hidrolisis (residu) disaring menggunakan kertas Whatman no. 41 yang dibantu dengan pompa vakum. Kemudian residu tersebut dimasukkan kedalam cawan porselen dan dikeringkan di dalam oven suhu 105 °C selama 24 jam untuk menentukan BK residu. Selanjutnya residu bahan kering (BK) dimasukan dalam tanur selama 6 jam untuk mendapatkan residu bahan organik (BO). Nilai KCBK dan KCBO dihitung berdasarkan rumus:
%KCBK = BK asal - (BK residu - BK blanko) x 100% BK asal
%KCBO = BO asal - (BO residu – BO blanko) x 100% BO asal
Pengukuran Degradasi Bahan Kering (DBK) dan Bahan Organik (DBO) (Tilley & Terry 1963)
Setelah diinkubasi selama 48 jam, ditambahkan 2-3 tetes HgCl2 jenuh untuk
menghentikan aktivitas mikroba. Lalu disentrifuge dengan kecepatan 3000 rpm selama 15 menit dan supernatannya dibuang. Kemudian residu tersebut dimasukkan kedalam cawan porselen dan dikeringkan di dalam oven suhu 105 °C selama 24 jam untuk menentukan BK residu. Selanjutnya residu bahan kering (BK) dimasukan dalam tanur selama 6 jam untuk mendapatkan residu bahan organik (BO). Nilai KCBK dan KCBO dihitung berdasarkan rumus:
%DBK = BK asal - (BK residu - BK blanko) x 100% BK asal
%DBO = BO asal - (BO residu – BO blanko) x 100% BO asal
Pengukuran Degradasi NDF (Tilley & Terry 1963)
Setelah diinkubasi selama 48 jam, ditambahkan 2-3 tetes HgCl2 jenuh untuk
9
selama 24 jam. Selanjutnya residu tersebut dianalisis kandungan NDF dengan metode Van Soest et al (1991). Nilai degradasi NDF dihitung berdasarkan rumus:
%DNDF = NDF pakan – NDF residu x 100% NDF pakan
Produksi Gas Metan (Moss et al. 2000)
Pengukuran estimasi produksi gas metan dihitung dengan menggunakan formula berdasarkan produksi VFA parsial (asetat, propionat, dan butirat).
CH4(%) = 0.45 C2– 0.275 C3 + 0.40 C4
Rancangan Percobaan
Penelitian ini menggunakan rancangan acak lengkap (RAL) dengan tiga perlakuan dan enam ulangan untuk kualitas fermentasi Hi-fer rumput gajah. Karakteristik fermentasi rumen menggunakan rancangan acak kelompok (RAK) dengan tiga perlakuan dan enam kelompok. Pengambilan cairan rumen sebagai kelompok. Perlakuan untuk kedua rancangan penelitian ini adalah:
T0: Hi-fer rumput gajah
T1: Hi-fer rumput gajah + inokulan T. reesei sebanyak 2.13 x 107 cfu/kg substrat
T2: Hi-fer rumput gajah + crude enzyme T. reesei sebanyak 11.4 IU/kg substrat
Model matematika dari rancangan ini adalah: Rancangan acak lengkap : Yij µ + τi + εij
Rancangan acak kelompok : Yij µ + τi + βj+ εij
Keterangan:
Yij: Hasil pengamatan pengaruh perlakuan ke-i dan ulangan ke-j
µ : Rataan umum
τi : Efek utama perlakuan ke-i
βj : Efek kelompok perlakuan ke-j
εij : galat perlakuan ke-i dan ulangan ke-j
Data karakteristik fisik (warna, bau, tekstur dan kontaminasi jamur) dianalisis secara deskriptif. Data komposisi kimia Hi-fer rumput gajah (proksimat, komposisi serat Van Soest, pH, asam laktat, VFA parsial dan NH3
dari Hi-fer), dan data karakteristik fermentasi rumen in vitro (pH, DBK, DBO, DNDF, KCBK, KCBO, total VFA dan parsial VFA, NH3 serta estimasi CH4)
12
20.19 – 22.74 % dan 85.59 – 87.01 % BK. Kandungan BK Hi-fer ini lebih tinggi dari silase rumput gajah dengan atau tanpa aditif yang dilaporkan Bureenok
et al. (2006) yaitu sekitar 14.5 – 18.1 %. Penurunan kandungan BK dan BO pada Hi-fer yang diberi perlakuan crude enzyme disebabkan karena perlakuan crude enzyme mendorong lebih banyak proses pendegradasian pati dan gula menjadi asam laktat dan produk VFA. Hasil yang sama dilaporkan Dehghani et al. (2012) bahwa beberapa enzim fibrolitik dengan penambahan 500 mg PK dari produk enzim/kg DM menurunkan BK silase maize stover.
Rataan kandungan SK Hi-ferpenelitian ini berkisar 25.65 – 28.95 % BK. Kandungan SK Hi-fer yang diberi crude enzyme lebih rendah dibandingkan perlakuan lainnya, mengindikasikan bahwa crude enzyme lebih efektif untuk mendegradasi komponen dinding sel tanaman dibandingkan penambahan inokulan T. reesei. McDonald et al. (1991) menyatakan bahwa enzim pendegradasi dinding sel mampu mendegradasi komponen dinding sel tanaman menjadi molekul yang lebih sederhana, sehingga menyediakan lebih banyak susbtrat untuk BAL.
Tabel 2 Komposisi nutrien rumput gajah dan Hi-ferrumput gajah setelah 21 hari fermentasi enzyme sebanyak 11.4 unit/kg berat segar, BK: bahan kering, BETN: bahan ekstrak tanpa nitrogen, TDN: total digestible nutrient, NDF: neutral detergent fiber, ADF: acid detergent fiber, SEM:
standard error of means. Rataan dalam baris yang sama dengan perbedaan huruf kecil menunjukkan berbeda nyata (P<0.05). Rataan dalam baris yang sama dengan perbedaan huruf kapital menunjukkan berbeda sangat nyata (P<0.01).
13
bakteri asam laktat (BAL) dan jumlah BAL meningkat yang menyebabkan peningkatan asam laktat, penurunan pH lebih cepat dan menghambat non-BAL dan enzim tanaman untuk proses proteolisis, akibatnya lebih sedikit proses degradasi serat dan protein dalam silase. Hasil penelitian yang sama dilaporkan Li
et al. (2014) bahwa penambahan aditif selulase silase rumput raja meningkatkan kandungan PK silase. Kandungan PK Hi-fer (6.76 – 7.40 % BK) pada penelitian ini lebih tinggi dari silase rumput gajah dengan atau tanpa aditif yang dilaporkan oleh Bureenok et al. (2012) yaitu sekitar 4.5 – 4.3 % BK.
Tidak ada perbedaan yang nyata pada kandungan LK (1.70 – 2.26 % BK) dan BETN (49.27 – 49.59 % BK) Hi-fer di antara semua perlakuan (P>0.05). Hasil yang sama dilaporkan Li et al. (2014) bahwa penambahan enzim selulase tidak berpengaruh terhadap kandungan LK silase rumput raja. Rataan kandungan TDN Hi-fer pada penelitian ini berkisar 50.76 – 54.61 % BK. Perlakuan penambahan crude enzyme pada Hi-fer meningkatkan (P<0.05) kandungan TDN dibandingkan dengan kontrol dan perlakuan inokulan T. reesei. Peningkatan kandungan TDN Hi-fer yang diberi aditif crude enzyme diduga akibat kandungan SK yang lebih rendah dan kandungan PK yang lebih tinggi. Sementara perlakuan penambahan inokulan T. reesei tidak berpengaruh terhadap kandungan TDN Hi-fer(P>0.05).
Perlakuan inokulan T. reesei dan crude enzyme signifikan menurunkan (P<0.01) kandungan NDF Hi-fer. Kandungan NDF Hi-fer yang diberi crude enzyme lebih rendah dibandingkan perlakuan lainnya. Hal ini mengindikasikan bahwa perlakuan crude enzyme lebih efektif dibandingkan perlakuan inokulan T. reesei untuk mendegradasi NDF. Perlakuan aditif T. reesei menurunkan NDF, hal ini diduga dari degradasi kompenen SK yang mudah dihidrolisis oleh enzim yang disekresikan T .reesei selama proses fermentasi. Enzim yang ditambahkan pada silase akan mendegradasi serat secara parsial menjadi water soluble carbohydrate
(WSC) untuk digunakan bakteri asam laktat (Eun & Beauchemin 2007). Hasil yang sama dilaporkan Xing et al. (2009) bahwa penambahan enzim selulase sebanyak 0.033 mg/g menurunkan kandungan NDF silase jerami sorgum sebesar 13.32%. Li et al. (2014) juga melaporkan bahwa penambahan enzim selulase sebanyak 0.02% menurunkan kandungan NDF silase rumput sebanyak 7.5%. Kandungan NDF Hi-fer (62.96 – 67.76 % BK) lebih rendah dari silase rumput gajah tanpa aditif yang dilaporkan Bureenok et al. (2012) yaitu sekitar 71.2 % BK.
Kandungan ADF Hi-fer tidak berbeda nyata (P>0.05) di antara semua perlakuan. Hal ini menunjukkan bahwa fraksi ADF belum dapat dirombak secara efektif oleh crude enzyme. Hasil yang sama dilaporkan Hidayat et al. (2005) bahwa penambahan crude enzyme T. viride sebanyak 3.8 – 11.4 IU/kg tidak berpengaruh nyata dalam menurunkan kandungan ADF silase jerami padi. Kandungan ADF Hi-fer (42.83 – 44.38 % BK) lebih tinggi dari silase rumput gajah tanpa aditif yang dilaporkan Bureenok et al. (2012) yaitu sekitar 40.1 % BK.
14
efektif menghidrolisis selulosa dan hemiselulosa namun mempunyai potensi dalam menghidrolisis komponen serat kasar yang relatif lebih mudah dihidrolis terutama hemiselulosa. Rataan kandungan lignin Hi-fer pada penelitian ini berkisar 6.45 – 7.45 % BK. Perlakuan inokulan T. reesei dan crude enzyme tidak signifikan menurunkan (P>0.05) kandungan NDF Hi-fer. Terdapat kecenderungan penurunan kandungan lignin Hi-fer yang diberi aditif. Penurunan kandungan lignin diduga akibat produksi asam-asam organik dari perlakuan penambahan aditif inokulan T. reesei dan crude enzyme selama proses fermentasi Hi-fer yang dapat mendegradasi lignin. Hidayat et al. (2005) melaporkan bahwa penurunan kandungan lignin sebesar 14 - 25% pada silase jerami padi yang diberi perlakuan crude enzyme T. viride disebabkan oleh produksi asam organik yang dihasilkan selama proses ensilase.
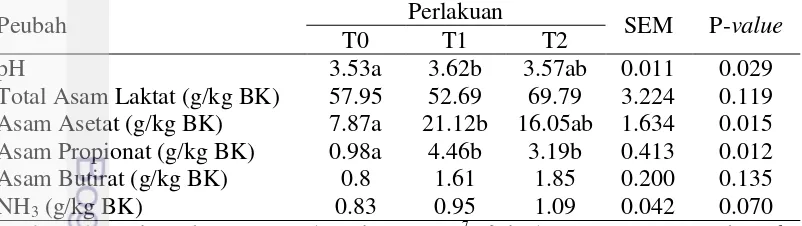
Karakteristik Fermentasi Hi-ferRumput Gajah
Pengaruh perlakuan terhadap karakteristik fermentasi Hi-fer rumput gajah ditunjukkan pada Tabel 3. Nilai pH Hi-fer pada perlakuan aditif crude enzyme
dan kontrol menunjukkan hasil yang sama. Perlakuan aditif T. reesei
meningkatkan pH Hi-fer dibandingkan dengan kontrol (P<0.05). Silase yang baik memiliki nilai pH<4.5 (McDonald et al. 1991). Rataan nilai pH Hi-fer
semua perlakuan berkisar 3.53–3.62 yang menunjukan Hi-fer yang dihasilkan berkualitas baik. Nilai pH Hi-feryang rendah mengakibatkan daya simpan yang baik. Tingginya konsentrasi asam asetat, asam propionat dan NH3 pada Hi-fer
yang diberi perlakuan inokulan T. reesei dan crude enzyme menyebabkan kenaikan pH. Arriola et al. (2011) menyatakan bahwa nilai pH silase yang tinggi berkaitan dengan rendahnya asam laktat serta tingginya asam asetat dan dan propionat. Penambahan inokulan T. reesei diduga mendorong pertumbuhan bakteri heterofermentatif yang menyebabkan nilai pH tinggi, asam laktat rendah dan produksi VFA tinggi. Arriola et al. (2011) melaporkan bahwa fermentasi bakteri heterofermentatif memiliki nilai pH yang tinggi, rendah konsentrasi asam laktat, tinggi konsentrasi asam asetat dan propionat.
Tabel 3 Karakteristik fermentasi Hi-fer rumpu gajah setelah 21 hari fermentasi
15
Rataan konsentrasi total asam laktat tertitrasi Hi-fer dari semua perlakuan sekitar 52.69 – 69.79 g/kg BK sehingga masuk dalam kategori berkualitas baik. Menurut McDonald et al. (1991) silase yang berkualitas baik memiliki konsentrasi asam <30 g/kg DM. Bureenok et al. (2012) mendapatkan konsentrasi asam laktat pada silase rumput gajah tanpa aditif sekitar 49 g/kg DM sementara pada silase rumput gajah dengan aditif sekitar 62 – 94 g/kg DM. Rataan kandungan total asam laktat Hi-fer yang diberi inokulan T. reesei dan crude enzyme tidak berbeda nyata (P>0.05) dengan kontrol. Hal ini diduga penambahan inokulan T. reesei dan crude enzyme asal T. reesei belum mampu meningkatkan pertumbuhan BAL selama proses fermentasi. Hasil penelitian yang sama dilaporkan Dean et al. (2005) bahwa perlakuan enzim fibrolitik pada silase
bermudagrass tidak signifikan meningkatkan konsentrasi asam laktat, namun nilainya lebih tinggi dibandingkan tanpa perlakuan enzim. Namun demikian, konsentrasi total asam laktat Hi-fer yang diberi perlakuan crude enzyme lebih tinggi dibandingkan kontrol. Hal ini diduga penambahan crude enzyme
meningkatkan degradasi NDF sehingga menyediakan lebih banyak substrat untuk
pertumbuhan BAL akibatnya produksi asam laktat meningkat. Konsentrasi asam asetat Hi-fer dipengaruhi oleh perlakuan penambahan
aditif. Hi-fer yang ditambahkan inokukan T. Reesei dan crude enzyme memiliki kandungan asam asetat yang lebih tinggi (P<0.05) dibandingkan kontrol. Hal ini menunjukkan bahwa penambahan inokulan T. reesei menyebabkan fermentasi didominasi oleh bakteri heterofermentatif sehingga terjadi peningkatan asam asetat. Menurut Bureenok et al. (2012) bahwa kandungan asam asetat yang tinggi pada silase akibat pertumbuhan bakteri BAL heterofermentatif. Konsentrasi asam asetat pada silase yang cukup (1 – 3% BK) akan menjaga kondisi stablitas aerob yang baik namun jika konsentrasinya tinggi (>3% BK) menggambarkan fermentasi yang tidak efisien karena kehilangan energi dan BK silase (Kung dan Shaver 2001). Rataan konsentrasi asam asetat Hi-fer yang diberi perlakuan inokulan T. reesei (21.12 g/kg BK) dan crude enzyme (16.05 g/kg BK) sudah cukup untuk menjaga kondisi stabilitas aerobik. Sesuai dengan hasil penelitian Xing et al. (2009) melaporkan bahwa konsentrasi asam asetat silase jerami sorgum berkisar 17.8 – 27.0 g/kg BK sudah cukup untuk menjaga stabilitas aerobik silase. Asam asetat dapat meningkatkan kondisi stabilitas aerobik karena dapat menghambat pertumbuhan yeast dan jamur lebih baik dibandingkan asam laktat (Moon 1983).
Perlakuan inokulan T. reesei dan crude enzyme asal T. reesei signifikan meningkatkan konsentrasi asam propionat (P<0.05). Hal ini diduga fermentasi
Hi-fer yang diberi perlakuan inokulan T. reesei dan crude enzyme didominasi oleh BAL heterofermentatif. Arriola et al. (2011) melaporkan bahwa fermentasi silase yang didominasi bakteri heterolaktat menghasilkan konsentrasi asam laktat yang rendah, konsentrasi asam asetat dan propionat yang tinggi. Hasil yang sama dilaporkan Dehghani et al. (2012) bahwa beberapa enzim fibrolitik meningkatkan kandungan asam propionat silase lucerne dan maize stover. Rataan konsentrasi asam propionat Hi-fer (0.98 – 4.46 g/kg BK) masih termasuk kategori baik. Menurut Kung dan Shaver (2001) melaporkan bahwa konsentrasi asam propionat pada silase rumput yang baik yaitu sekitar <1 g/kg BK. Kandungan asam asetat dan propionat yang cukup tinggi pada perlakuan inokulan dan crude enzyme
16
asam propionate merupakan agen anti jamur yang dapat menghambat pertumbuhan yeast dan jamur pada silase (Moon 1983; McDonald et al. 1991). Filya dan Sucu (2007) melaporkan bahwa tingginya konsentrasi asam asetat (14.9 – 27.4 g/kg BK) dan asam propionat (0.1 – 7.8 g/kg BK) dapat meningkatkan kondisi stabilitas aerobik dari silase tanaman gandum.
Rataan konsentrasi asam butirat Hi-fer pada penelitian ini yaitu berkisar 0.8 – 1.85 g/kg BK. Konsentrasi asam butirat Hi-fer tidak berbeda nyata (P>0.05) di antara semua perlakuan. Hasil penelitian yang sama dilaporkan Dean
et al. (2005) bahwa pada pemberian beberapa enzim fibrolitik tidak memberikan pengaruh terhadap kandungan asam butirat silase bermudagrass. Kandungan asam butirat Hi-fer pada penelitian ini masih cukup rendah sehingga masih termasuk ke dalam kategori silase yang baik. Menurut McDonald et al. (1991) bahwa silase yang mengandung asam butirat lebih dari 10% (% dari total asam) termasuk silase berkategori tidak baik. Konsentrasi asam butirat Hi-fer ini lebih rendah dari silase rumput gajah tanpa aditif yang dilaporkan Bureenok et al. (2012) yaitu sekitar 16.1 g/kg BK.
Konsentrasi NH3 Hi-fer (0.83 – 1.09 g/kg BK) tidak signifikan (P>0.05)
dipengaruhi oleh perlakuan. Hasil ini menunjukkan bahwa penambahan aditif inokulan T. reesei dan crude enzyme tidak berpengaruh terhadap aktivitas proteolisis pada Hi-fer. Sesuai dengan penelitian ini, Xing et al. (2009) melaporkan bahwa kandungan NH3-N silase jerami sorgum pada kontrol dan
perlakuan enzim fibrolitik (0.033 mg/g) menunjukkan hasil yang serupa. Kandungan N-NH3 berhubungan pada degradasi PK dan asam amino, yang
merupakan indikator tingkat proteolysis di silase (Chen et al. 2014). Pada kondisi pH rendah, pertumbuhan clostridia proteolitik yang merombak asam amino menjadi NH3 akan menurun (Bureenok et al. 2006). Kondisi pH Hi-fer yang
rendah ternyata belum mampu menghambat pertumbuhan clostridia proteolitik dan hidrolisis protein secara enzimatik. Hal ini diduga WCS yang diproduksi dari degradasi serat kasar oleh inokulan T. reesei dan crude enzyme belum cukup untuk menurunkan kandungan NH3 Hi-fer. Kandungan NH3 Hi-fer pada
penelitian ini tergolong rendah sehingga kualitas Hi-fer yang dihasilkan termasuk kategori baik. Menurut Xing et al. (2009) bahwa konsentrasi N-NH3
yang rendah (<25g/kg BK) menunjukkan silase yang baik.
Nilai Fleigh dan Populasi Bakteri Asam Laktat Hi-ferRumput Gajah Pengaruh perlakuan aditif terhadap nilai fleigh dan jumlah koloni BAL
Hi-ferrumput gajah ditunjukkan pada Tabel 4. Perlakuan aditif pada Hi-fertidak signifikan mempengaruhi nilai fleigh (P>0.05). Nilai fleigh Hi-fer pada penelitian ini berkisar 102.71 – 105.80. Berdasarkan NF yang dihasilkan maka
Hi-fer pada penelitian ini termasuk kategori sangat baik. Idikut et al. (2009) menyatakan bahwa silase yang termasuk kategori sangat baik memiliki NF>85.
17
pertumbuhan BAL terhambat. Cai et al. (1999) melaporkan bahwa silase yang berkualitas baik ketika populasi BAL setidaknya mencapai 105 (cfu/g).
Tabel 4 Nilai fleigh dan populasi bakteri asam laktat Hi-ferrumput gajah
Peubah Perlakuan SEM P-value
T0 T1 T2
Nilai Fleigh 102.71 105.80 102.71 0.605 0.101 BAL (log cfu/g) 5.40 5.18 5.34 0.221 0.916
T0: kontrol, T1: kontrol+ T. reesei sebanyak 2.13 x 107 cfu/kg berat segar, T2: control+crude enzyme sebanyak 11.4 unit/kg berat segar, BAL: bakteri asam laktat, SEM: standard error of means.
Karakteristik Fermentasi Rumen
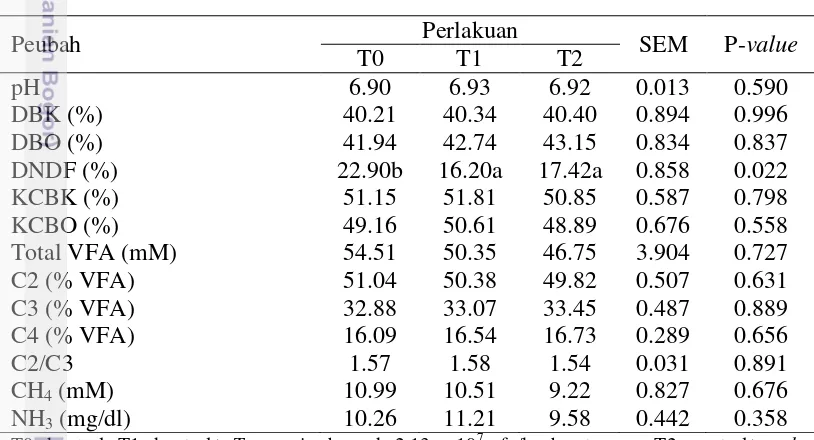
Karakteristik fermentasi rumen Hi-fer dapat dilihat pada Tabel 5. Tidak ada perbedaan signifikan (P>0.05) di antara semua perlakuan terhadap semua peubah karakteristik fermentasi rumen Hi-fer. Rataan nilai pH rumen yang dihasilkan pada penelitian ini berkisar 6.90 – 6.92. Menurut Krause dan Oetzel (2006) kisaran normal nilai pH yaitu 5.5-7.0. Nilai pH rumen yang berada pada kisaran normal menggambarkan bahwa Hi-fer pada setiap perlakuan masih dapat mempertahankan kondisi rumen yang sesuai untuk proses fermentasi pakan.
Tabel 5 Karakteristik Fermentasi Rumen
Peubah Perlakuan SEM P-value
T0 T1 T2
pH 6.90 6.93 6.92 0.013 0.590 DBK (%) 40.21 40.34 40.40 0.894 0.996 DBO (%) 41.94 42.74 43.15 0.834 0.837 DNDF (%) 22.90b 16.20a 17.42a 0.858 0.022 KCBK (%) 51.15 51.81 50.85 0.587 0.798 KCBO (%) 49.16 50.61 48.89 0.676 0.558 Total VFA (mM) 54.51 50.35 46.75 3.904 0.727 C2 (% VFA) 51.04 50.38 49.82 0.507 0.631 C3 (% VFA) 32.88 33.07 33.45 0.487 0.889 C4 (% VFA) 16.09 16.54 16.73 0.289 0.656 C2/C3 1.57 1.58 1.54 0.031 0.891 CH4 (mM) 10.99 10.51 9.22 0.827 0.676
18
Nilai DBK dan DBO tidak dipengaruhi (P>0.05) oleh perlakuan inokulan T. reesei dan crude enzyme. Hal ini menunjukkan bahwa penambahan aditif T. reesei
dan crude enzyme pada Hi-fer belum mampu meningkatkan aktivitas mikroba rumen dalam mendegradasi pakan. Rataan nilai DBK dan DBO pada penelitian ini masing-masing berkisar 40.21 – 40.40% dan 41.94 – 43.15%. Hasil penelitian yang sama dilaporkan oleh Kozelov et al. (2008) bahwa penambahan enzim selulase (5 kg/t) tidak memberikan pengaruh terhadap nilai DBK silase alfalfa. Lynch et al. (2014) juga melaporkan hasil yang sama bahwa penambahan enzim fibrolitik tidak memberikan pengaruh positif terhadap nilai DBK silase alfafa.
Perlakuan penambahan inokulan T. reesei dan crude enzyme signifikan menurunkan (P<0.05) nilai DNDF dibandingkan kontrol. Meskipun kandungan NDF Hi-fer perlakuan inokulan T. reesei dan crude enzyme menurun, perlakuan memberikan respon negatif terhadap nilai DNDF. Penurunan nilai DNDF pada Hi-feryang diberi perlakuan inokulan T. reesei dan crude enzyme diduga karena selama fermentasi terjadi hidrolisis fraksi serat yang mudah dicerna sehingga menurunkan nilai degradasi NDF. Menurut Kim et al. (2014) rendahnya degradabilitas NDF dikaitkan pada rendahnya fraksi NDF yang mudah dicerna. Sesuai dengan penelitian Nadeau et al. (2000) bahwa penurunan kecernaan NDF pada silase yang diberi perlakuan enzim selulase dikaitkan pada degradasi dari bagian NDF yang mudah dicerna oleh selulase selama proses ensilase sehingga tersisa bagian NDF yang sulit dicerna oleh mikroba rumen. Kozelov et al. (2008) dan Lynch et al. (2014) melaporkan bahwa penambahan enzim fibrolitik tidak memberikan pengaruh terhadap nilai DNDF silase.
Nilai kecernaan dapat menunjukkan ketersediaan nutrien pakan yang dapat dimanfaatkan oleh mikroba rumen. Nilai KCBK dan KCBO pada penelitian ini masing-masing berkisar 50.85 – 51.81% dan 48.89 – 50.61%. Penambahan aditif inokulan T. reesei dan crude enzyme pada Hi-fer belum memberikan efek positif terhadap kecernaan in vitro. Sesuai dengan penelitian Dean et al. (2005), hanya satu dari empat jenis enzim fibrolitik yang meningkatkan nilai KCBK silase
bermudagrass yang diinkubasi selama 48 jam. Dehghani et al. (2012) juga melaporkan bahwa meskipun penambahan enzim fibrolitik menurunkan kandungan NDF namun nilai KCBO silase alfafa tidak dipengaruhi oleh penambahan enzim dan bahkan menurunkan KCBO untuk beberapa perlakuan penambahan enzim pada silase maize stover. Adanya penurunan kandungan SK dan NDF pada Hi-fer yang ditambahkan crude enzyme tidak menyebabkan
Hi-fer tersebut lebih mudah didegradasi oleh mikroba rumen. Hal ini diduga enzim selulase pada penelitian ini telah menghidrolisis komponen serat kasar yang mudah dicerna selama proses fermentasi dan komponen tersebut juga yang didegradasi oleh mikroba rumen. Akibatnya komponen serat yang tersisa berupa komponen serat yang sulit dicerna mikroba rumen. Menurut Nadeau et al. (1996) penambahan enzim selulase pada hijauan silase menurunkan degradabilitas rumen karena enzim sudah mendegradasi fraksi serat yang mudah dicerna selama proses ensilase sehingga menyisakan serat yang kurang dapat dicerna oleh mikroba rumen.
Penambahan aditif pada Hi-fer menghasilkan total VFA yang sama dengan kontrol. Hal ini menunjukkan bahwa penambahan aditif pada Hi-fer
19
belum mampu menstimulir pertumbuhan mikroba rumen. Pemberian pakan yang mudah difermentasi oleh mikroba rumen akan meningkatkan kecernaan dan degradasi pakan yang diikuti peningkatan VFA. Rataan produksi total VFA pada penelitian ini (46.75 – 54.51 mM) berada di bawah batas konsentrasi optimum yang dinyatakan McDonald et al. (2010) yaitu 70 – 150 mM. Total VFA penelitian ini sedikit lebih tinggi dibandingkan dengan total VFA rumput gajah (48.54 mM) yang dilaporkan oleh Okoruwa dan Igene (2014). Kisaran rataan konsentrasi VFA yang rendah diduga oleh karena rendahnya kandungan karbohidrat dan protein yang mudah dicerna dalam Hi-fer menyebabkan perbedaan populasi mikroba rumen yang berakibat pada penurunan konsentrasi VFA total. Persentase asetat (C2), propionat (C3), dan butirat (C3) tidak dipengaruhi oleh penambahan aditif. Rataan persentase rasio C2/C3 juga tidak dipengaruhi oleh perlakuan penambahan aditif pada Hi-fer. Semua perlakuan menghasilkan rasio C2/C3 yang rendah (1.54 – 1.57). Rendahnya rasio C2/C3 dapat memberikan keuntungan terhadap produksi ternak. Propionat merupakan prekursor utama untuk proses glukoneogenesis di ruminansia, yang akan dikonversi menjadi glukosa di dalam hati. Glukosa yang terbentuk dapat digunakan ternak untuk aktivitas hidup pokok, produksi dan reproduksi (McDonald et al. 2010).
Menurut Moss et al. (2000) produksi CH4 dipengaruhi oleh produksi asetat,
propionat dan butirat. Proporsi C3 yang tinggi menurunkan produksi CH4 karena
H2 digunakan selama pembentukan propionat. Rasio C2/C3 Hi-fer yang rendah
dan mengarah pada pembentukan propionat umumnya menghasilkan konsentrasi CH4 yang rendah, namun pada penelitian ini produksi CH4 tidak dipengaruhi
(P>0.05) oleh perlakuan. Cao et al. (2010) melaporkan bahwa kandungan asam laktat yang tinggi pada silase menurunkan produksi metan dalam rumen. Rendahnya kandungan NDF Hi-fer belum mampu menghasilkan substrat yang lebih banyak untuk pertumbuhan BAL sehingga kandungan asam laktat Hi-fer
yang tidak berbeda nyata. Akibatnya proporsi molar C3 yang dihasilkan sama dan produksi CH4 tidak berbeda di antara semua perlakuan. Asam laktat yang berasal
dari silase akan dikonversi menjadi asam propionat dalam rumen oleh bakteri
Selenomonas ruminantium dan Megasphaera elsdenii (Cao et al. 2010; Flythe & Aiken 2010).
Perlakuan penambahan aditif pada Hi-fer menghasilkan konsentrasi NH3
rumen yang sama dengan kontrol. Tidak adanya perbedaan pada konsentrasi NH3
menunjukkan bahwa Hi-fer pada setiap perlakuan memberikan efisiensi penggunaan amonia yang sama. Hal ini diduga penambahan aditif Hi-fer belum mampu meningkatkan fermentabilitas Hi-fer sehingga tidak ada perubahan aktivitas mikroba rumen yang mendorong pembentukan NH3. pada Di dalam
rumen NH3 digunakan mikroba rumen sebagai sumber nitrogen untuk sintesis
protein mikroba (McDonald et al. 2010). Rataan konsentrasi NH3 rumen pada
penelitian ini berkisar 9.58 – 11.21 mg/dl atau setara 5.63 – 6.58 mM. Nilai tersebut masih dalam kisaran konsentrasi optimum yang dinyatakan McDonald et al. (2010) berkisar 85 – 300 mg/l atau setara 6 – 21 mM. Menurut Syahrir et al. (2009), konsentrasi NH3 yang rendah dapat menujukkan proses fermentasi yang
20
PK Hi-fer yang diduga menyebabkan rendahnya konsentrasi NH3 yang
terbentuk.
4 SIMPULAN
Semua perlakuan menghasilkan Hi-fer rumput gajah berkualitas baik dengan nilai pH Hi-fer yang rendah, tinggi konsentrasi asam laktat dan rendah konsentrasi asam butirat. Penambahan inokulan Trichoderma reesei atau crude enzyme asal T. reesei dapat meningkatkan aerobic stability Hi-fer dengan meningkatkan konsentrasi asam asetat dan asam propionat Hi-fer. Penambahan
21
DAFTAR PUSTAKA
Adav SS, Chao LT, Sze SK. 2012. Quantitative secretomic analysis of Trichoderma reesei strains reveals enzymatic composition for ligocellulosic biomass degredation. Mol Cell Proteomics. 11(7):1-15.
Amanullah SM, Kim DH, Lee HJ, Joo YH, Kim SB, Kim SC. 2014. Effect of Microbial Additives on Chemical Composition and Fermentaton Characteristics of Barley Silage. Asian-Australas J Anim Sci. 27(4): 511-517.
Anwar N, Widjaja A, Winardi S. 2010. Peningkatan unjuk kerja hidrolisis enzimatik jerami padi menggunakan campuran selulase kasar dari Trichoderma reesei dan Aspergillus niger. Makara Sains. 14(2): 113-116. [AOAC] Association of Official Agricultural Chemists. 2005. Official Methods of
Analysis of the Association of Analytical Chemist. 18th Ed. Arlington (US): Assoc of Offi. Anal. Chem.
Arriola KG, Kim SC, Adesogan AT. 2011. Effect of applying inoculants with heterolactic or homolactic and heterolactic bacteria on the fermentation and quality of corn silage. J Dairy Sci. 94:1511-1516.
Avellaneda JH, Pinos-Rodríguez JM, González SS, Bárcena R, Hernández A, Cobos M, Hernández D, Montanez O. 2009. Effects of exogenous fibrolytic enzymes on ruminal fermentation and digestion of Guinea grass hay. Anim Feed Sci Technol.149: 70-77.
Bailey MJ, Buchert J, Viikari L. 1993. Effect of pH on production of xylanase by
Trichoderma reesei on xylan- and cellulose-based media. Appl Microbiol Biotechnol. 40: 224-229.
Bureenok S, Namihara T, Mizumachi S, Kawamoto Y, Nakada T. 2006. The effect of epiphytic lactic acid bacteria with or without different byproduct from defatted rice bran and green tea waste on napiergrass (Pennisetum purpureum Shumach) silage fermentation. J Sci Food Agric. 86:1073-1077. Bureenok S, Yuangklang C, Vasupen K, Schonewille JT, Kawamoto Y. 2012.
The effects of additives in napier grass silages on chemichal composition, feed intake, nutrient digestibility and rumen fermentation. Asian-Australas J Anim Sci. 25:1248-1254.
Cai Y, Benno Y, Ogawa M, Kumai S. 1999. Effect of applying lactic acid bacteria isolated from forage crops on fermentation characteristics anad aerobic deterioration of silage. J Dairy Sci. 82:520-526.
Cao Y, Takahashi T, Horiguchi K, Yoshida N. 2010. Effect of adding lactic acid bacteria and molasses on fermentation quality and in vitro ruminal digestion of total mixed ratio silage prepared with whole crop rice. Grassl Sci. 56:19-25.
Chen L, Guo G, Yuan X, Shimojo M, Yu C, Shao T. 2014. Effect of applying molasses and propionic acid on fermentation quality and aerobic stability of total mixed ratio silage prepared with whole-plant corn in Tibet. Asian-Australas J Anim Sci. 27(3): 349-356.