i
KARAKTERISASI DAN PEMANFAATAN PLASMA NUTFAH
MELALUI PENDEKATAN PEMULIAAN DAN MOLEKULER
UNTUK PENINGKATAN HASIL BUAH KOPYOR
DAN KUALITAS BENIH KOPYOR
ISMAIL MASKROMO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
iii
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertasi berjudul “Karakterisasi dan Pemanfaatan Plasma Nutfah Melalui Pendekatan Pemuliaan dan Molekuler untuk Peningkatan Hasil Buah Kopyor dan Kualitas Benih Kopyor” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka pada setiap bagian disertasi ini. Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
Ismail Maskromo
iii
RINGKASAN
ISMAIL MASKROMO. Karakterisasi dan Pemanfaatan Plasma Nutfah Melalui Pendekatan Pemuliaan dan Molekuler untuk Peningkatan Hasil Buah Kopyor dan Kualitas Benih Kopyor. Dibimbing oleh SUDARSONO sebagai ketua, HENGKY NOVARIANTO, SUKENDAH dan DEWI SUKMA sebagai anggota komisi pembimbing.
Kelapa kopyor merupakan salah satu komoditi perkebunan yang prospektif untuk meningkatkan pendapatan petani dan pemerintah daerah. Namun demikian dalam pengembangannya masih menghadapi beberapa masalah yaitu rendahnya produksi, keterbatasan bahan tanaman yang berkualitas, teknik budidaya yang belum optimal dan pola pengelolaan tanaman yang kurang tepat. Penelitian ini bertujuan untuk : (1) Melakukan inventarisasi keberadaan dan mendata kuantitas hasil aksesi dan varietas kelapa kopyor dari berbagai lokasi di Indonesia, (2) Mengevaluasi karakteristik pembungaan dan perkembangan buah kopyor pada varietas kelapa Genjah kopyor, (3) Mengidentifikasi pengaruh negatif xenia terhadap hasil buah kopyor pada varietas kelapa Genjah dan kelapa Dalam kopyor, (4) Mengembangkan dan mengevaluasi metode peningkatan hasil buah kopyor di tingkat petani melalui penyerbukan terkontrol, (5) Menentukan mekanisme genetik pengendali sifat kopyor pada aksesi kelapa kopyor yang berasal dari berbagai lokasi di Indonesia, (6) Mengevaluasi keragaman genetik aksesi kelapa kopyor yang berasal dari berbagai lokasi di Indonesia berdasarkan hasil analisis marka molekuler SSR, (7) Mengevaluasi pola ko-segregasi antar lokus dari sejumlah marka molekuler yang telah dikembangkan untuk genom kelapa dan menganalisis keterpautan (linkage) antara sifat kopyor dengan marka
molekuler yang dievaluasi.
Hasil survey menunjukkan bahwa tanaman kelapa kopyor menyebar dalam bentuk populasi pada beberapa sentra kelapa di wilayah Sumenep dan Jember-Jawa Timur, Pati-Jember-Jawa Tengah, Purwakarta-Jember-Jawa Barat, Tangerang-Banten dan Kalianda-Lampung Selatan. Selain itu terdapat tanaman kelapa kopyor secara terbatas di daerah Banyuwangi dan Ponorogo-Jawa Timur, Purbalingga dan Purwokerto-Jawa Tengah serta di Bogor-Jawa Barat. Secara umum produksi buah kopyor semua aksesi kelapa kopyor yang diamati di tingkat petani relatif rendah, yaitu di bawah 25 % untuk kelapa kopyor tipe Dalam dan di bawah 40% untuk kelapa kopyor tipe Genjah. Kuantitas endosperma kelapa Genjah kopyor dapat dikategorikan berdasarkan volume daging buahnya dengan skor 1-6, sedangkan pada kelapa Dalam kopyor memiliki skor 1-9.
iv kelapa Genjah kopyor asal Pati dikategorikan sebagai tanaman menyerbuk sendiri secara langsung atau autogami langsung.
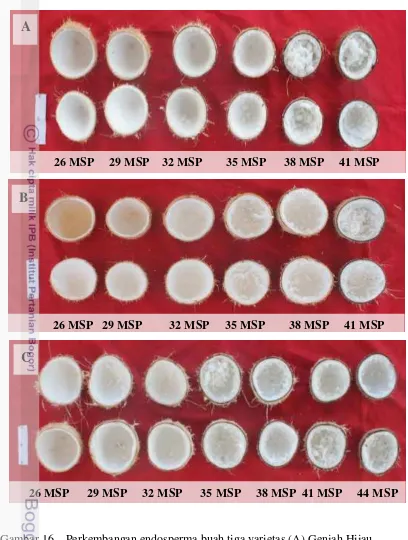
Perkembangan buah pada tanaman kelapa Genjah kopyor tidak berbeda dengan buah kelapa normal. Endosperma tiga varietas kelapa Genjah kopyor mulai dapat dibedakan dengan buah normal pada umur 23 minggu setelah penyerbukan (MSP), dengan struktur remah dan tidak stabil. Proses penebalan endosperma kelapa kopyor mulai berlangsung pada umur 23 MSP dan mencapai maksimal pada umur 41 MSP. Endosperma mulai terlepas dari tempurung pada umur 32-35 MSP. Kuantitas endosperma optimal pada umur 41 MSP, tertinggi pada kelapa Genjah kopyor Coklat (GCK) yaitu 275 g dan terendah pada Genjah Kopyor Hijau (GHK) yaitu 190 g. Penurunan bobot buah utuh dan bobot sabut pada buah kopyor terjadi pada umur 35 MSP, yang berbeda secara signifikan dengan buah normal akibat proses pematangan buah yang lebih cepat. Karakteristik buah kopyor dengan sabut yang lebih dahulu mengalami kematangan dibanding buah normal, menjadi indikator pemilihan buah kopyor oleh petani pemanen kopyor melalui metode pengetukan buah mulai umur 35 MSP. Perubahan warna kulit buah kopyor mulai terjadi pada umur 38 MSP.
Pengamatan terhadap populasi campuran kelapa Dalam kopyor heterosigot Kk dengan kelapa Dalam normal KK dilakukan di Kalianda, Lampung Selatan, sedangkan populasi kelapa Genjah kopyor heterosigot Kk dengan kelapa normal KK dilakukan di Kecamatan Tayu, Pati, Jawa Tengah. Hasil penelitian menunjukkan bahwa Xenia terjadi karena keberadaan kelapa normal di antara tanaman kelapa kopyor. Xenia berpengaruh negatif terhadap hasil buah kopyor yang dipanen dari kelapa Dalam Kopyor Kalianda dan kelapa Genjah Kopyor Pati. Selain menyebabkan jumlah panen buah kopyor yang rendah, keberadaan tanaman kelapa normal KK juga dapat menyebabkan rendahnya kualitas benih dan bibit yang dihasilkan dari pohon kelapa heterosigot Kk. Untuk mencegah pengaruh negatif xenia pada produksi buah kopyor, jumlah pohon kelapa kopyor yang ada di lokasi pertanaman kelapa kopyor heterosigot Kk perlu ditingkatkan atau jumlah pohon kelapa normal KK yang ada disarankan untuk ditebang, kemudian diganti dengan tanaman kelapa kopyor homosigot kk.
Pengembangan teknologi peningkatan produksi buah kopyor dan produksi benih true-to-type dilakukan dengan penyerbukan terkontrol pada tanaman kelapa
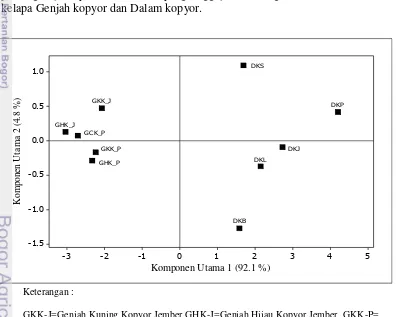
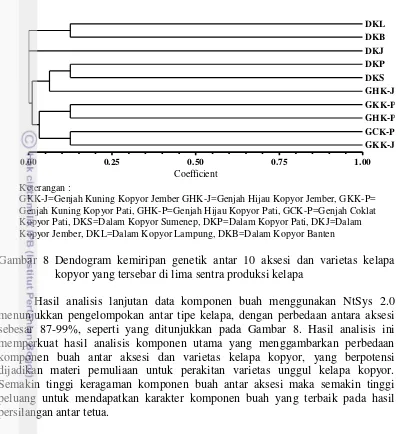
v Keberadaan tanaman kelapa kopyor yang menyebar di berbagai daerah memunculkan pertanyaan, kemungkinan adanya hubungan genetik antar aksesi atau varietas kelapa kopyor. Melalui analisis molekuler dengan marka SSR, sebanyak lima aksesi kelapa Dalam kopyor dan lima aksesi/varietas kelapa Genjah kopyor dipelajari keragaman genetiknya. Selanjutnya untuk mempelajari lokus gen pengendali sifat kopyor pada aksesi dan varietas kelapa kopyor yang berasal dari lima sentra produksi kelapa kopyor, dilakukan persilangan terkontrol antara varietas kelapa Genjah Hijau kopyor heterosigot Kk dengan kelapa Dalam kopyor heterosigot asal Kalianda-Lampung Selatan, Pati-Jawa Tengah, Jember dan Sumenep-Jawa Timur. Secara umum masing-masing aksesi dan varietas yang diuji menunjukkan perbedaan genetik satu dengan lainnya dengan membentuk dua kelompok besar berdasarkan tipe kelapa kopyor, yaitu tipe Genjah dan tipe Dalam. Namun demikian terdapat beberapa individu yang mengelompok dengan aksesi atau varietas lainnya, yang diduga akibat telah terjadi persilangan alami antar provenan yang berbeda aksesi dan varietas. Selain itu diduga akibat terjadinya penyebaran tanaman kelapa berbuah kopyor ke daerah lain yang telah terjadi pada kurun waktu sebelumnya. Melalui persilangan terkontrol antar aksesi dan varietas kelapa kopyor dapat menghasilkan buah kopyor dengan persentase yang berbeda antar kombinasi persilangan. Hal ini menunjukkan bahwa sifat kopyor yang eksis pada beragam genetik kelapa tetapi dikendalikan oleh lokus gen yang sama. Berdasarkan perbandingan jumlah buah normal dan buah kopyor diduga sifat kopyor pada kelapa dikendalikan oleh satu sampai dua lokus gen. Hasil penelitian ini dapat dijadikan dasar penyusunan program pemuliaan untuk mendapatkan varietas unggul baru kelapa kopyor dengan menggabungkan sifat kopyor dari berbagai latar belakang genetik kelapa berbeda.
Ko-segregasi marka SSR dan SNP yang diuji pada 121 progeni populasi F2 kelapa kopyor menghasilkan 14 lokus yang mengikuti pola segregasi Mendel, sedangkan 7 lokus lainnya menyimpang. Sebanyak enam lokus yang terpaut dan tersebar pada dua group pautan pada peta awal keterkaitan marka SSR dan SNP dengan lokus pengendali sifat kopyor pada kelapa. Dua lokus SSR dan dua marka SNP terletak pada group pautan dengan sifat kopyor. Marka SNP WRKY_21 yang sangat dekat posisinya dengan sifat kopyor yaitu jarak 9 cM diikuti marka CnCir_J2 dengan jarak 12.2 cM dan marka SNP CnSUS#3 dengan jarak 14.4 cM Penambahan jumlah marka dan jumlah sampel tanaman yang diuji perlu dilakukan untuk mendapatkan kerapatan marka yang tinggi pada kromosom dan marka yang lebih dekat serta sangat terpaut dengan sifat kopyor pada kelapa. Marka-marka tersebut dapat diuji lebih lanjut untuk dijadikan sebagai kandidat marka, dalam seleksi dini bibit kelapa kopyor.
vii
SUMMARY
ISMAIL MASKROMO Characterization and utilization of germplasm through breeding and molecular approach to improved kopyor fruit yield and kopyor seed quality. Supervised by SUDARSONO as chairman, HENGKY NOVARIANTO, SUKENDAH and DEWI SUKMA as member of advisory committee.
Kopyor coconut is one of the commodities that prospectively to increase the income of farmers and local governments. However, its development still faces some problems ie : low production, quality plant material limitations, which is not optimal cultivation techniques and crop management patterns that are less precise. This study aims to: (1) Conduct an inventory existence and record quantity of accession and kopyor coconut varieties from various locations in Indonesia, (2) Evaluate the characteristics of flowering and fruit development the Dwarf kopyor coconut varieties, (3) Identification negative influence of the xenia on kopyor fruits harvested of Tall kopyor and Dwarf kopyor varieties, (4) Develop and evaluate methods of increasing yields kopyor fruit at the farm level through controlled pollination, (5) Determining the genetic mechanisms controlling the kopyor trait of the accession coconut kopyor originating from various locations in Indonesia, (6) Evaluating the genetic diversity of kopyor coconut accessions originating from various locations in Indonesia based on molecular markers, (7) To evaluate patterns of co-segregation between the locus of a number of molecular markers which have been developed for coconut genome and analyzing the linkage of kopyor trait with molecular markers evaluated.
The survey results showed that coconut kopyor plants spread in population form at some coconut centers in the area of Java, namely Sumenep and Jember in East Java, Pati, Central Java, Purwakarta, West Java, Tangerang, Banten and Kalianda, South Lampung. In addition it was reported that there is limited kopyor palm plant in Banyuwangi and Ponorogo in East Java, Purbalingga and Purwokerto, Central Java, and in Bogor, West Java. In general, the production of kopyor fruit in all accession kopyor observed at the farm level is relatively low, at under 25% for Tall kopyor accessions and under 40% for coconut Dwarf kopyor accessions. Character of fruit components vary between accessions, with a high value of diversity in the fruit weight and endosperm weight. Endosperm quantity of Dwarf kopyor accessions can be categorized by a score of 1-6 based on the volume of the fruit, while in Tall kopyor has a score of 1-9.
viii In general, the development of the fruit in the Dwarf kopyor varieties no different from normal coconut. Dwarf kopyor endosperm kopyor of three varieties began to be distinguished with normal fruit endosperm at the age of 23 weeks after pollination (WAP), with the crumb structure and unstable. Process of endosperm thickening in kopyor coconut start at the age of 23 WAP and reaches a maximum at the age of 41 WAP. Endosperm start regardless of the shell at the age of 32-35 WAP. Endosperm optimal quantity at the age of 41 WAP highest in Kopyor Brown Dwarf (GCK) is 275g and the lowest in Kopyor Green Dwarf (GHK) is 190g. Fruit weight and weight of fiber loss in kopyor fruits occurs at age 35 WAP, which differ significantly from the normal fruit due process of fruit ripening faster. Characteristics of kopyor fruit with husks that previously maturity than normal fruit be an indication for kopyor fruit selection by farmers to harvesting kopyor fruit through tapping method start at age 35 WAP. Kopyor fruit skin discoloration began to occur at the age of 38 WAP.
Observation at the mixture population of kopyor coconut heterozygous Kk and normal coconut tree KK conducted in Kalianda, South Lampung, while for Dwarf kopyor heterozygous Kk population with normal cococnut KK in Tayu District, Pati, Central Java. The results showed that xenia occurs due to the presence of the normal tree on kopyor plantations. The presence of more normal trees (KK) among provenances of Kopyor Tall coconut (Kk) resulted in lower harvested kopyor fruits than the expected one. Moreover, removal of normal coconut trees (KK) from Kopyor Dwarf coconut provenances (Kk) increased yield of kopyor fruits. Those indicated that xenia occurs in both Kopyor Tall and Dwarf coconut types. Xenia negatively affects yield of kopyor fruits and quality of seeds and seedlings are produced by farmers.
Technology development to increase kopyor fruit production and produce of true-to-type kopyor seed was conducted by a controlled pollination in Dwarf coconut kopyor heterozygous use kopyor coconut pollen homozygous that grown through embryo culture technique. Research conducted on two controlled pollination method is : Metode 1 by applying pollen pure on the stigma of the female flower coconut kopyor and method 2 by spraying pollen added with talcum on the female flowers using a plastic bottle as a spray. The results showed that coconut controlled pollination of kopyor heterozygous Kk use kopyor coconut pollen homozygous (kk) can increase yields of kopyor fruit up to 50%. Method of pollination controlled by spraying pollen were added talc to produce the amount of fruit harvest is not significantly different than the method of stocking of pure pollen, but more efficient in the use of pollen. True to type kopyor seed heterozygous (Kk) can be obtained through controlled pollination between coconut kopyor heterozygous (Kk) with coconut kopyor pollen homozygous (kk) while increasing yields of kopyor fruit.
ix Java. In general, each individual accessions and varieties tested showed genetic differences from one another by forming two large groups based on the kopyor Tall type and kopyor Dwarf type. However, there are some individuals who are clustered with other accessions or varieties, that allegedly caused had occurred crossbreeding between provenances of different varieties and accession. Besides allegedly caused by the spread of kopyor plants to other areas which have occurred in the previous period. Through a controlled cross between kopyor varieties and accession can produce fruit kopyor with different percentages between cross combinations. This suggests that kopyor that kopyor mutans existed in different coconut genetic background but controlled by similar locus. It can be used as a basis for the breeding program to get new varieties of kopyor coconut by combining kopyor trait from various genetic backgrounds of different coconut.
Co-segregation of SSR and SNP markers were tested on a population of 121 F2 progeny coconut kopyor produce 14 loci which follows Mendelian segregation patterns, while seven other loci diverge. A total of four loci were adrift and spread on two linkage groups of kopyor coconut tested. Two SSR loci and two SNP markers located on linkage group with kopyor trait. SNP markers `with distance of 12.2 cM and SNP marker CnSUS # 3 with a distance of 14.4 cM. Addition number of markers and the number of samples tested plants needs to be done to obtain a high density on chromosome and markers closer with kopyor trait on coconut. The markers can be used as candidate markers for early selection of kopyor coconut seedlings.
xi
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
xiii
KARAKTERISASI DAN PEMANFAATAN PLASMA
NUTFAH MELALUI PENDEKATAN PEMULIAAN DAN
MOLEKULER UNTUK PENINGKATAN HASIL BUAH
KOPYOR DAN KUALITAS BENIH KOPYOR
ISMAIL MASKROMO
Disertasisebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Pemuliaan dan Bioteknologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
xiv Penguji pada Ujian Tertutup: - Dr. Ir. Asep Setiawan, M.S
- Dr. Ir. Rr. Sri Hartati M.P Penguji pada Ujian Terbuka: - Dr. Ir. Asep Setiawan, M.S
xvii
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala rahmat dan karuniah-Nya sehingga penulis berhasil menyelesaikan disertasi ini dengan judul:
“Karakterisasi dan Pemanfaatan Plasma Nutfah Melalui Pendekatan Pemuliaan dan Molekuler untuk Peningkatan Hasil Buah Kopyor dan Kualitas Benih Kopyor”.
Penulis mengucapkan terima kasih kepada Prof. Dr. Ir. Sudarsono, MSc. sebagai Ketua Komisi Pembimbing, dan kepada Prof. (Riset) Dr. Ir. Hengky Novarianto, MS, Dr. Ir. Sukendah, MSc dan Dr. Dewi Sukma, SP. MSi. sebagai anggota Komisi Pembimbing yang telah banyak memberi saran-saran dan masukan sejak persiapan, pelaksanaan penelitian sampai penyusunan disertasi ini. Penulis juga menyampaikan terima kasih kepada Dr. Ir. Sudrajat MS, Dr. Ir. Yudiwanti Wahyu EK. MS. Dr. Ir. Asep Setiawan MS, Dr. Ir. Rr. Sri Hartati, MP, dan Dr. Ir. Fadjry Djufry, MSi, yang telah bersedia menjadi penguji luar komisi pada ujian pra kualifikasi program Doktor, Ujian Tertutup dan Ujian Terbuka, serta memberikan masukan dan saran perbaikan untuk kesempurnaan disertasi ini.
Terima kasih juga penulis ucapkan kepada Balai Penelitian Tanaman Palma, Pusat Penelitian dan Pengembangan Perkebunan dan Badan Penelitian dan Pengembangan Pertanian, Kementerian Pertanian Republik Indonesia, yang telah memberi kesempatan dan dukungan biaya kepada penulis untuk melangsungkan studi S3 di IPB.
Ucapan terimakasih disampaikan juga kepada Badan Penelitian dan Pengembangan Pertanian, Kementerian Pertanian Republik Indonesia yang telah memberikan dukungan dana penelitian melalui Program KKP3T tahun 2011 dan Program KKP3N 2012-2015, serta Direktorat Jenderal Pendidikan Tinggi, Kementerian Pendidikan dan Kebudayaan melalui Program Hi-link 2012-2014.
Tidak lupa penulis ucapkan terima kasih kepada Kepala Balai Penelitian Tanaman Palma, seluruh teman-teman di Balai Penelitian Tanaman Palma, pimpinan dan staf Dinas Kehutanan dan Perkebunan Kabupaten Pati, Kabupaten Jember, Kabupaten Sumenep dan Provinsi Banten, pimpinan dan staf Dinas Perkebunan Kabupaten Lampung Selatan, serta teman-teman dan teknisi di Laboratorium Biologi Molekuler Tanaman Depertemen AGH IPB, yang telah membantu baik secara fisik maupun psikologis selama berlangsungnya kegiatan penelitian dan dukungannya dalam penulisan disertasi ini.
Ungkapan terima kasih juga penulis sampaikan kepada kedua orang tua,
sanak saudara, istri tercinta Ashaima Nazly Syarif dan anakku tersayang M. Rayyan Ismail Maskromo, atas dukungan dan kesabarannya mendampingi
penulis selama menempuh pendidikan S3 di IPB.
Penulis menyadari masih banyak kekurangan, sehingga diharapkan saran dan kritik yang membangun demi penyempurnaan disertasi. Akhir kata, semoga disertasi ini bermanfaat bagi kemajuan ilmu pemuliaan dan biologi molekuler tanaman, khususnya tanaman kelapa di Indonesia.
xix
DAFTAR ISI
DAFTAR TABEL ... xxii
iDAFTAR GAMBAR ... xxiv
DAFTAR LAMPIRAN ... xxviii
1 PENDAHULUAN 1 Latar Belakang ... 1
Tujuan Penelitian ... 4
Manfaat Penelitian ... 4
Kebaruann Penelitian ... 5
Kerangka Berpikir dan Garis Besar Disertasi ... 5
Diagram Alir Penelitian ... 10
2 TINJAUAN PUSTAKA 11 Kelapa Kopyor………... 11
Penyebaran Kelapa Kopyor ... 11
Karakteristik Tanaman Kelapa Kopyor ... 11
Pola Pembungaan Kelapa Kopyor... 14
Persilangan pada Tanaman Kelapa ... 15
Marka Morfologi dan Molekuler... 16
Peta Keterpautan Genetik ... 18
Daftar Pustaka ... 20
3 HASIL DAN KUANTITAS ENDOSPERMA BUAH KOPYOR ASAL LIMA SENTRA PRODUKSI KELAPA KOPYOR INDONESIA 25 Abstrak ... 25
Abstract ... 26
Pendahuluan ... 27
Bahan dan Metode ... 28
Hasil dan Pembahasan ... 29
Simpulan ... 38
Daftar Pustaka ... 38
4 FENOLOGI BUNGA DAN PERKEMBANGAN BUAH KELAPA GENJAH KOPYOR 41 Abstrak ... 41
Abstract ... 42
Pendahuluan ... 43
Bahan dan Metode ... 44
Hasil dan Pembahasan ... 46
Simpulan ... 58
xx 5 XENIA BERPENGARUH NEGATIF TERHADAP HASIL BUAH
KOPYOR PADA KELAPA DALAM DAN GENJAH KOPYOR
61 Abstrak ... 61 Abstract ... 62 Pendahuluan ... 63 Bahan dan Metode ... 64 Hasil dan Pembahasan ... 66 Simpulan ... 72 Daftar Pustaka ... 72
6 SIFAT KOPYOR EKSIS PADA BERAGAM GENETIK KELAPA TETAPI DIKENDALIKAN OLEH LOKUS GEN YANG SAMA
75 Abstrak ... 75 Abstract ... 76 Pendahuluan ... 77 Bahan dan Metode ... 79 Hasil dan Pembahasan ... 81 Simpulan ... 92 Daftar Pustaka ... 92
7 PENYERBUKAN TERKONTROL MENINGKATKAN HASIL BUAH KOPYOR DAN BENIH KELAPA KOPYOR TRUE TO TYPE
95
Abstrak ... 95 Abstract ... 96 Pendahuluan ... 97 Bahan dan Metode ... 99 Hasil dan Pembahasan ... 102 Simpulan ... 107 Daftar Pustaka ... 108
8 PETA AWAL KETERKAITAN ANTARA MARKA SSR DAN LOKUS PENGENDALI SIFAT KOPYOR PADA KELAPA
109 Abstrak ... 109 Abstract ... 110 Pendahuluan ... 111 Bahan dan Metode ... 112 Hasil dan Pembahasan ... 115 Simpulan ... 122 Daftar Pustaka ... 123
9 PEMBAHASAN UMUM 127
10 SIMPULAN UMUM 129
RIWAYAT HIDUP LAMPIRAN
xxi
DAFTAR TABEL
1 Kemungkinan genotipe dan fenotipe embryo yang terbentuk pada segregasi persilangan antara tanaman kopyor heterosigot Kk 13 2 Kemungkinan genotope endosperma yang terbentuk pada segregasi
persilangan antara tanaman kopyor heterosigot Kk 13 3 Lokasi penyebaran, tipe kelapa, luas dan hasil panen buah kelapa
kopyor pada lima sentra produksi kelapa kopyor di Indonesia 32 4 Karakter komponen buah lima aksesi kelapa Dalam kopyor asal
Lampung, Banten, Pati, Jember dan Sumenep 34
5 Karakter komponen buah tiga varietas kelapa Genjah kopyor asal Pati, Jawa Tengah dan dua aksesi kelapa Genjah kopyor asal Jember, Jawa
Timur 34
6 Rata-rata jumlah bunga betina tiga varietas kelapa Genjah kopyor
di KP. Kima Atas, BalitPalma Manado dan di Kabupaten Pati 46 7 Pola pembungaan tiga varietas Genjah Kopyor Pati yang tumbuh di
Kabupaten Pati dan yang ditanam di KP. Kima Atas, BalitPalma
Manado 48
8 Rataan jumlah buah kelapa total dan buah kopyor dipanen dari masing-masing sub-populasi di tiga lokasi pertanaman kelapa. Data merupakan rataan dari hasil pengamatan selama tiga tahun. 69 9 Jumlah alel, jumlah alel homosigot dan alel heterosigot, nilai
heterosigositas dan PIC 26 lokus SSR yang mengampifikasi 125 sampel
DNA kelapa kopyor 82
10 Jumlah buah jadi hasil persilangan terkontrol beberapa genotipe kelapa Genjah Hijau Kopyor Pati dengan empat aksesi Kelapa Dalam kopyor 90 11 Jumlah buah, segregasi dan prediksi lokus segregasi hasil persilangan
model genetik pada kelapa kopyor Genjah dan Dalam 90 12 Segregasi progeni hasil persilangan tetua kelapa kopyor heterosigot P1
(K1k1) x P2(K2k2) dengan model genetik satu lokus 91 13 Segregasi progeni hasil persilangan tetua kelapa kopyor heterosigot P1
(K1k1K2k2) x P2(K1k1K2k2) dengan model genetik dua lokus 91 14 Perbandingan jumlah buah kopyor hasil penyerbukan alami dan
penyerbukan terkontrol pada varietas GHK (Genjah Hijau Kopyor), GKK (Genjah Kuning Kopyor) dan GCK (Genjah Coklat Kopyor) 104 15 Perbandingan jumlah buah kopyor hasil penyerbukan terkontrol metode
1 dan penyerbukan terkontrol metode 2 pada varietas GHK (Genjah Hijau Kopyor), GKK (Genjah Kuning Kopyor) dan GCK (Genjah
Coklat Kopyor) 105
16 Jumlah total bunga betina, buah panen, buah kopyor dan buah normal hasil penyerbukan terkontrol tiga varietas kelapa Genjah kopyor Kk
dengan serbuk sari kelapa Genjah kopyor kk 106
17 Daftar lokus SSR kelapa hasil seleksi pada individu F1 kelapa kopyor 117 18 Daftar primer SNP kelapa hasil seleksi pada individu F1 kelapa kopyor 118 19 Ko-segregasi 19 marka SSR dan tiga marka SNP pada populasi F2
xxiii
DAFTAR GAMBAR
1 Diagram alir penelitian 10
2 Perbandingan endosperma (A) Buah kelapa normal dan (B) Buah
kelapa kopyor 12
3 Perbandingan endosperma abnormal pada (A) kelapa kopyor (B) kelapa
Lilin dan (C) kelapa Makapuno. 12
4 Peta letak lima sentra produksi kelapa kopyor di Indonesia 30 5 Pertanaman kelapa kopyor di lapang (A) : Sumenep, Jawa Timur, (B)
Populasi Kalianda, Lampung Selatan, (C) Pati, Jawa Tengah dan(D)
Jember, Jawa Timur. 30
6 Keragaman buah kopyor (A) tipe Dalam dan (B) tipe Genjah 33 7 Hasil analisis komponen utama lima aksesi kelapa Dalam kopyor dan
lima varietas aksesi kelapa Genjah kopyor 35
8 Dendogram jarak genetik antar 10 aksesi dan varietas kelapa kopyor
yang tersebar di lima sentra produksi kelapa 36
9 Skor kuantitas endosperma aksesi kelapa Dalam kopyor 37 10 Skor kuantitas endosperma varietas/aksesi kelapa Genjah kopyor 37 11 Tandan bunga (A) Genjah Hijau di Tayu, Pati dan (B) Genjah Hijau
Kopyor di Kima Atas Manado 47
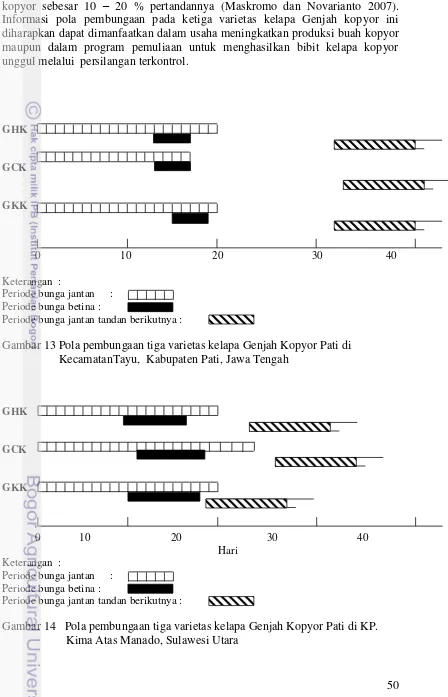
12 Pola pembungaan kelapa Genjah kopyor (A) Awal perode bunga jantan, (B) Masa reseptif bunga betina, (C) Akhir periode bunga jantan 48 13 Pola pembungaan tiga varietas kelapa Genjah Kopyor Pati di
KecamatanTayu, Kabupaten Pati, Jawa Tengah 50
14 Pola pembungaan tiga varietas kelapa Genjah Kopyor Pati di KP. Kima
Atas Manado, Sulawesi Utara 50
15 Perkembangan ukuran, rongga dan endosperma buah tiga varietas
kelapa Genjah kopyor umur 2- 23 MSP 51
16 Perkembangan endosperma buah tiga varietas (A) Genjah Hijau Kopyor-GHK, (B) Genjah Kuning Kopyor – GKK, dan(C) Genjah
Coklat Kopyor - GCK pada umur 26-44 MSP 52
17 Perbandingan bobot endosperma buah kopyor (K) dan buah normal (N) umur 23-41 MSP pada tiga varietas (A) Genjah Hijau Kopyor, (B) Genjah Coklat Kopyor dan (C) Genjah Kuning Kopyor 53 18 Perbandingan bobot buah kopyor (K) dan buah normal (N) umur 2-41
MSP pada tiga varietas (A) Genjah Hijau Kopyor, (B) Genjah Coklat
Kopyor dan (C) Genjah Kuning Kopyor 55
19 Perbandingan bobot sabut buah kopyor (K) dan buah normal (N) umur 23-41 pada tiga varietas (A) Genjah Hijau Kopyor, (B) Genjah Coklat
Kopyor dan (C) Genjah Kuning Kopyor 56
20 Perbandingan warna buah kopyor (K) dan buah normal (N) pada tiga varietas (A) Genjah Hijau Kopyor, (B) Genjah Coklat Kopyor dan (C) Genjah Kuning Kopyor pada tiga varietas kelapa Genjah kopyor umur
32-41 MSP 57
xxiv kategori sub-populasi di lokasi (A) Kecapi (B) Agom Jaya dan (C)
Palembapang 67
22 Hubungan antara curah hujan pada dua tahun sebelum buah dipanen (2009-2011) dan rataan jumlah buah kelapa total yang dipanen per pohon per tahun (2011-2013) di tiga lokasi penelitian 67 23 Hubungan antara curah hujan pada dua tahun sebelum buah dipanen
(2009-2011) dan rataan jumlah buah kelapa kopyor total yang dipanen per pohon per tahun (2011-2013) di tiga lokasi penelitian 68 24 Rataan persentase buah kopyor per tandan dipanen dari masing-masing
kategori sub-populasi di tiga lokasi pertanaman kelapa. Data merupakan rataan dari hasil pengamatan selama tiga tahun. Kategori 1 (N:K=2:1) : sub-populasi dengan rasio jumlah pohon kelapa normal KK dan pohon kopyor Kk = 2:1 dan kategori 2: dengan rasio KK : Kk =
1:1. N = Normal ; K = Kopyor 69
25 Peta tanaman kelapa normal KK dan kelapa Genjah Kopyor Pati Kk di Desa Sambiroto Tayu, Pati, Jawa Tengah. Petak mengindikasikan sub-populasi sampel yang dievaluasi (A) Kondisi sebelum penebangan kelapa normal KK dan (B) Kodisi setelah penebangan sebagian pohon
kelapa normal KK yang ada di lokasi. 71
26 Rataan persentase buah kopyor per tandan yang dipanen dari pohon kelapa Genjah Kopyor Pati Kk di Tayu, Pati. Jawa Tengah sebelum dan setelah penebangan secara bertahap sebagian pohon kelapa normal KK
yang ada di lokasi. 71
27 Hasil seleksi lokus SSR (A) CnCir_147, (B) CnCir_47, (C) CnCir_73, (D) CnCir_109, (E)_CnCir192, (F) CnCir_206 dan (G) CnCir_H4 pada
tujuh sampel DNA kelapa kopyor 82
28 Profil marka SSR pada 15 genotipe kelapa Dalam Kopyor Lampung
(DKL) menggunakan primer CnCir_A9. 84
29 Profil marka SSR pada 25 genotipe Genjah Kopyor asal Pati, jawa
Tengah menggunakan primer CnCir_206. 84
30 Profil marka SSR pada 25 genotipe Genjah Kopyor asal Jember, Jawa
Timur menggunakan primer CnCir_206. 84
31 Dendogram kemiripan genetik 125 individu kelapa kopyor menggunakan 26 primer SSR berdasarkan analisis DARwin 5.0.158. 85 32 Dendogram kemiripan genetik kelapa Genjah dan Dalam kopyor asal
Pati dengan kelapa Genjah kopyor asal Jember menggunakan 26 primer
SSR berdasarkan analisis DARwin 5.0.158. 86
33 Dendogram kemiripan genetik lima aksesi kelapa Dalam kopyor menggunakan 26 primer SSR berdasarkan analisis DARwin 5.0.158. 87 34 Struktur populasi 10 aksesi kelapa kopyor dari lima sentra produksi
kelapa berdasarkan analisis Structure (Pritchard 2000) 88 35 Struktur populasi lima aksesi kelapa Genjah kopyor berdasarkan
analisis Structure (Pritchard 2000) 88
36 Struktur populasi lima aksesi kelapa Dalam kopyor dari lima sentra produksi kelapa berdasarkan analisis Structure (Pritchard 2000) 88 37 Perkembangan jumlah buah jadi hasil penyerbukan alami dan
xxv 38 Representasi perkembangan buah jadi varietas Genjah Kuning Kopyor
hasil penyerbukan terkontrol pada umur (A) 2 minggu setelah penyerbukan (MSP), (B) 4 MSP, (C ) 8 MSP dan (D) 12 MSP 104 39 Perkembangan jumlah buah jadi hasil penyerbukan terkontrol metode 1
dan metode 2 pada varietas (A) Genjah Hijau Kopyor, (B) Genjah
Kuning Kopyor dan (C) Genjah Coklat 105
40 Jumlah buah jadi hasil penyerbukan terkontrol untuk produksi benih
True-to-Type pada tiga varietas kelapa Genjah kopyor 107 41 Skema diagram genotyping marka SNAP yang digunakan untuk
validasi marka putatif SNAP pada fragmen gen WRKY, SUS, SACPD
dan ABI dari tanaman kelapa. 115
42 Seleksi tujuh lokus SSR pada sampel DNA F1 dan F2 kelapa kopyor 116 43 Hasil analisis SSR terhadap 25 genotipe populasi F2 kelapa kopyor
menggunakan lokus (A) CNZ_21, (B) CNZ_51 dan (C) CnCir_A9 118 44 Hasil analisis SSR terhadap 25 genotipe populasi F2 kelapa kopyor
menggunakan marka SNP Sus#3 118
45 Peta pautan empat marka SSR dan dua marka SNP dengan sifat kopyor
xxvii
DAFTAR LAMPIRAN
1 Bahan tanaman yang digunakan dalam sub kegiatan penelitian "Penyerbukan terkontrol untuk meningkatkan hasil buah kopyor dan benih kelapa kopyor True To Type" ... 133 2 Prosedur pelaksanaan pengolahan bunga jantan dan pengujian serbuk
1
PENDAHULUAN
Latar Belakang
Kelapa kopyor adalah kelapa mutan asli Indonesia dengan endosperma (daging buah kelapa) abnormal. Abnormalitas endosperma dengan fenotipe lunak dan terlepas dari cangkangnya tersebut, ternyata memiliki nilai komersial yang tinggi. Harga buah kelapa kopyor di pasaran bisa mencapai 10 kali lipat dibanding buah kelapa biasa.
Kelapa kopyor berbeda fenotipenya dengan kelapa "Makapuno", yang berasal dari Filipina. Abnormalitas endosperma pada kelapa "Makapuno" menyebabkan jaringannya menjadi lunak seperti jeli dan jika terlalu tua sebagian dari endospermanya akan terlarut dalam air kelapa, sehingga air kelapanya menjadi kental seperti minyak pelumas. Dari hasil penelitian dapat diketahui bahwa abnormalitas endosperma kelapa Makapuno terjadi karena defisiensi aktivitas ensim α-D-Galaktosidase dalam perkembangan endospermanya (Mujer et al. 1984, Samonthe et al. 1989). Karakteristik mutan pada kelapa Makapuno tersebut dilaporkan dapat diturunkan secara genetik dari tetua ke progeninya (Santos 1999).
Karakteristik umum abnormalitas endosperma pada kelapa kopyor adalah tekstur endosperma tidak padat namun lembut sampai remah seperti tekstur gabus, terlepas dari tempurungnya, membentuk serpihan-serpihan yang memenuhi seluruh lubang tempurung (Maskromo et al. 2007). Seperti halnya pada makapuno, abnormalitas fenotipe endosperma kelapa kopyor diduga juga terjadi akibat adanya defisiensi enzim penting tertentu selama dalam proses perkembangan endospermanya. Walaupun demikian identitas enzim yang menyebabkan abnormalitas endosperma kelapa kopyor sampai saat ini masih belum diketahui. Karakteristik mutan pada kelapa kopyor juga dapat diturunkan secara genetik dari tetua ke progeninya (Sukendah 2009).
Keberadaan dan potensi kelapa kopyor perlu terus dilestarikan dan dikembangkan lebih lanjut, agar sumberdaya genetik asli Indonesia tersebut dapat dimanfaatkan sepenuhnya untuk kesejahteraan rakyat Indonesia. Produksi buah kelapa kopyor dari beberapa sentra produksi tanaman kelapa kopyor masih terbatas. Pasokan per minggu sebanyak 3,000-5,000 butir buah dari Pati, Jawa Tengah dan 300-500 butir kelapa kopyor dari Kalianda, Lampung Selatan, belum mampu memenuhi permintaan pasar di Jakarta yang terus meningkat.
2 tanaman (benih kelapa kopyor) yang digunakan dan pola pengelolaan (budidaya) tanaman kelapa kopyor yang dipraktekkan di lapangan.
Beberapa penelitian pendahuluan telah dilakukan untuk mendukung pengembangan kelapa kopyor di Indonesia ke depan. Informasi keberadaan tanaman kelapa kopyor di berbagai daerah telah dilaporkan, dan menjadi dasar untuk program penelitian selanjutnya. Di pulau Jawa, kelapa kopyor banyak ditemukan di Provinsi Jawa Tengah (Sukendah 2009), di daerah Tangerang, Provinsi Banten, serta di Sumenep, Provinsi Jawa Timur (Akuba 2002). Mahmud (2000) melaporkan bahwa kelapa kopyor juga terdapat di daerah Kalianda, Kabupaten Lampung Selatan. Namun demikian informasi terkait produksi buah kopyor dan karakteristik daging buah kopyor belum tersedia. Penelitian terkait identifikasi keragaman morfologi dan keragaman genetik berdasarkan 5 marka SSR telah dilakukan pada populasi kelapa kopyor yang tumbuh di Kalianda, Lampung Selatan, Ciomas, Bogor, dan Sumenep, Jawa Timur (Maskromo 2005). Namun demikian, dengan masih terbatasnya aksesi yang diamati dan marka SSR yang digunakan, hasil yang didapat masih belum dapat memberikan informasi yang lengkap tentang keragaman genetik plasma nutfah kelapa kopyor di Indonesia.
Penggunaan marka SSR dalam analisis DNA kelapa, didasarkan pada keunggulannya yaitu memiliki resolusi tinggi, bersifat kodominan dan keterulangan yang tinggi dibandingkan dengan metode lainnya (Teulat et al. 2000). Melalui metode analisis DNA juga sudah dirancang primer-primer terkait enzim dan protein yang diduga berhubungan dengan pembentukan daging buah kopyor (Sukendah 2009). Namun demikian primer-primer tersebut belum diuji kemampuannya, untuk membedakan tanaman yang berbuah kopyor dengan tanaman kelapa normal. Padahal jika informasi tersebut dapat diperoleh, akan sangat bermanfaat penggunaannya pada seleksi dini bibit kelapa kopyor.
Usaha peningkatan produksi buah kopyor telah dilakukan melalui pengembangan bibit dengan metode kultur embrio (Mashud et al. 2004). Tanaman
hasil kultur embrio tersebut berpotensi menghasilkan buah kopyor sampai 100 persen per tandannya. Namun demikian bibit kelapa kopyor berteknologi tinggi ini masih terbatas jumlahnya dan relatif mahal harganya untuk tingkat petani. Oleh sebab itu petani kelapa kopyor masih memilih untuk menggunakan bibit alami, dengan potensi berbuah kopyor hanya berkisar 10 - 40 persen, tergantung pada jenis kelapa yang dikembangkan. Keberhasilan terbentuknya buah kopyor diduga terjadi karena adanya peluang bertemunya gen pengendali sifat kopyor yang dimiliki bunga betina, dengan gen pengendali sifat kopyor yang dibawa oleh serbuk sari dari pohon yang sama atau pohon lainnya. Melalui persilangan terkontrol pada kelapa kopyor heterosigot menggunakan serbuk sari dari tanaman kelapa kopyor homosigot hasil kultur embrio, diharapkan dapat meningkatkan peluang terjadinya buah kopyor pada tanaman kelapa berbuah kopyor tersebut.
3 "k", sedangkan endosperma normal dikendalikan oleh gen dominan "K." Secara genetik, segregasi genotipe tanaman kelapa kopyor pada saat terbentuk buah dalam satu tandan akan menghasilkan tiga macam genotipe yaitu genotipe "kk", "Kk" dan "KK".
Pola tersebut di atas diduga mengikuti pola pewarisan sifat mutan pada kelapa Makapuno yang dilaporkan Santos (1999). Buah kelapa kopyor dengan genotipe embrio sigotik "kk" tidak akan berkecambah secara alami karena abnormal pada endospermanya, namun embrio sigotiknya dapat tumbuh normal seperti layaknya pada buah kelapa normal. Embrio sigotik kk dari buah kopyor dapat ditumbuhkan dengan bantuan teknologi kultur in vitro. Selanjutnya buah
dengan genotipe embrio sigotik "Kk" atau "KK" memiliki fenotipe endosperma normal, sehingga dapat berkecambah secara alami. Campuran bibit kelapa hasil perkecambahan embrio sigotik dengan genotipe "Kk" atau "KK" tersebut yang digunakan oleh petani sebagai bibit kelapa kopyor alami. Fenotipe bibit kelapa
yang berkembang dari embrio sigotik “Kk” atau “KK” tersebut masih sulit dibedakan secara morfologi, sehingga perlu sentuhan teknologi konvensional ataupun non-konvensional untuk mendapatkan solusinya.
Pada penyerbukan antar tanaman kelapa yang memiliki gen pengendali sifat kopyor di lapang, terdapat peluang terbentuknya buah kopyor dengan
genotipe “kk” dan buah normal heterosigot “Kk” atau buah normal homosigot
„KK”. Peluang terbentuknya buah normal heterosigot “Kk” melalui penyerbukan alami tersebut, dapat dijadikan dasar untuk perakitan benih true to type kelapa kopyor dengan metode penyerbukan terkontrol. Metode yang dapat digunakan yaitu dengan cara menyilangkan tetua betina tanaman kelapa kopyor heterosigot
„Kk‟ dengan serbuk sari kelapa kopyor hasil kultur embryo yang bergenotipe homosigot “kk” atau sebaliknya membiarkan serbuk sari kelapa kopyor heterosigot Kk atau kelapa normal KK yang unggul menyerbuki tanaman dalam populasi kelapa kopyor hasil kultur bergenotipe kk. Pemilihan metode didasarkan pada ketersediaan bahan tanaman, kemudahan dan tingkat keberhasilan pelaksanaan penyerbukan terkontrol.
Keberadaan kelapa kopyor di berbagai sentra produksi memunculkan pertanyaan tentang keragaman genetik antar aksesi dan varietas dalam kaitannya dengan kemungkinan adanya keragaman gen pengendali sifat kopyor. Melalui persilangan terkontrol antar aksesi dan varietas kelapa kopyor dapat dipelajari mekanisme genetik pengendali sifat kopyor. Selanjutnya untuk mendapatkan informasi kandidat marka molekuler terpaut sifat kopyor pada kelapa, dapat dimulai dengan menggunakan populasi F2 hasil selfing dari hibrida F1 kelapa kopyor. Tujuannya untuk mengevaluasi pola ko-segregasi antar lokus dari sejumlah marka molekuler yang telah dikembangkan untuk genom kelapa dan menganalisis keterpautan (linkage) antara sifat kopyor dengan marka molekuler
pengembangan peta genetik awal genom kelapa Genjah kopyor berbasis marka molekuler SSR.
Berbagai masalah tentang kelapa kopyor di lapangan saat ini dan kendala-kendala yang ada terkait pengembangannya di Indonesia ke depan, menjadi dasar pelaksanaan kegiatan penelitian ini. Sebagian besar penelitian disertasi ini merupakan bagian dari kegiatan penelitian yang didanai oleh Kementerian Pertanian melalui skema penelitian KKP3T tahun 2011 (Sudarsono et al. 2012)
4 dan oleh Kementerian Pendidikan dan Kebudayaan melalui skema penelitian Hi-LINK dari tahun 2012-2014 (Sudarsono et al. 2015b).
Tujuan Penelitian
Tujuan umum penelitian ini adalah untuk mengembangkan teknologi peningkatan produksi buah kopyor dan produksi benih true to type melalui
persilangan terkontrol serta mendapatkan kandidat marka molekuler untuk seleksi bibit kelapa kopyor. Adapun tujuan khusus penelitian adalah:
1. Menginventarisasi keberadaan dan keragaaan hasil aksesi dan varietas kelapa kopyor dari berbagai lokasi di Indonesia,
2. Mengevaluasi karakteristik pembungaan dan perkembangan buah kopyor pada varietas kelapa Genjah kopyor
3. Mengidentifikasi pengaruh negatif xenia terhadap hasil buah kopyor pada varietas kelapa Genjah dan kelapa Dalam kopyor
4. Mengembangkan dan mengevaluasi metode peningkatan hasil buah kopyor di tingkat petani melalui penyerbukan terkontrol
5. Mengevaluasi keragaman genetik aksesi kelapa kopyor yang berasal dari berbagai lokasi di Indonesia berdasarkan hasil analisis marka molekuler SSR
6. Menentukan mekanisme genetik pengendali sifat kopyor pada aksesi kelapa kopyor yang berasal dari berbagai lokasi di Indonesia
7. Mengevaluasi pola ko-segregasi antar lokus sejumlah marka molekuler yang telah dikembangkan untuk genom kelapa dan menganalisis keterpautan (linkage) antara sifat kopyor dengan marka molekuler yang dievaluasi
Manfaat Penelitian
5 Kebaruan Penelitian
Hasil penelitian yang disajikan pada disertasi ini meliputi aspek genetika, pemuliaan dan molekuler tentang permasalahan dalam pengembangan kelapa kopyor di lapang dan solusi pemecahannya melalui penelitian lapang dan laboratorium. Adapun poin-poin kebaruan yang berhasil diperoleh dari penelitian ini meliputi :
(1) Informasi keberadaan pertanaman, potensi hasil panen buah kopyor dan karakteristik komponen buah dan kuantitas endosperma aksesi/varietas kelapa kopyor yang berkembang di lima sentra kelapa kopyor di Indonesia, sebagai informasi dasar dalam menyusun program pemuliaan selanjutnya. (2) Informasi pola pembungaan dan perkembangan buah kelapa Genjah
kopyor sebagai dasar pemahaman tentang karaktersik bunga dan buah kelapa kopyor yang sangat bermanfaat dalam program pemuliaan kelapa kopyor.
(3) Diketahuinya efek xenia sebagai salah satu penyebab rendahnya produksi pada populasi campuran kelapa kopyor heterosigot Kk dan kelapa normal KK di lapang, diharapkan dapat membantu pola pengelolaan tanaman untuk meningkatkan hasil panen buah kopyor pada tingkat petani.
(4) Diperolehnya metode penyerbukan terkontrol yang efisien untuk meningkatkan hasil panen buah kopyor dan merakit bibit true to type
kelapa kopyor heterzigot Kk. Peningkatan hasil buah panen akan meningkatkan pendapatan petani kopyor, sedangkan penggunaan bibit true to type akan memperbaiki pola pertanaman di lapang dalam mendukung
peningkatan produksi kelapa kopyor.
(5) Informasi keragaman genetik secara lengkap 10 aksesi dan varietas kelapa kopyor dari lima sentra produksi kelapa kopyor berdasarkan marka SSR dan kaitannya dengan lokus pengendali sifat kopyor pada kelapa yang diuji melalui persilangan terkontrol antar varietas atau aksesi kelapa kopyor. Informasi ini bermanfaat dalam menyusun program pemuliaan merakit varietas unggul baru kelapa kopyor.
(6) Diperolehnya dua lokus SSR dan dua marka SNP yang terletak pada satu group pautan dengan sifat kopyor sebagai langkah awal penyusunan peta genetik kelapa kopyor untuk mendapatkan kandidat marka SSR dan SNP terpaut sifat kopyor pada kelapa.
Kerangka Berpikir dan Garis Besar Disertasi
6 Beberapa penelitian pendahuluan telah dilakukan oleh para peneliti BalitPalma Manado meliputi eksplorasi dan identifikasi plasma nutfah, analisis keragaman morfologi dan genetik secara terbatas pada beberapa aksesi kelapa kopyor yang menyebar di beberapa sentra kelapa di wilayah Pulau Jawa dan ujung selatan Pulau Sumatera (Maskromo 2005). Penelitian selanjutnya adalah identifikasi dan evaluasi potensi plasma nutfah kelapa Genjah kopyor yang menyebar di Kabupaten Pati, Jawa Tengah (Maskromo 2010). Salah satu hasilnya adalah telah dirilisnya tiga varietas kelapa Genjah kopyor asal Pati, Jawa Tengah oleh Menteri Pertanian pada tahun 2010 (Maskromo et al. 2015).
Pengembangan tanaman kelapa kopyor oleh petani selama ini masih menggunakan benih alami bergenotipe Kk dari tanaman kelapa kopyor di lapang. Hal ini diduga menjadi penyebab rendahnya produksi buah kopyor. Penelitian terkait penyediaan benih true-to-type (kk) kelapa kopyor melalui kultur embryo
yang memiliki potensi menghasilkan buah kopyor 100 persen pertandannya telah dilakukan oleh Puslit Biotek Perkebunan, Bogor, BalitPalma Manado, IPB, Bogor dan Universitas Muhamadiyah Purwokerto. Tingkat keberhasilan menghasilkan bibit siap tanam di lapang yang masih rendah serta biaya penyediaan yang relatif mahal, menyebabkan harganya yang tidak terjangkau oleh petani. Keberadaan kelapa kopyor di lapang, potensi tanaman, rendahnya hasil buah kopyor dan keterbatasan bahan tanaman berkualitas serta berbagai permasalahan dalam pengembangan kelapa kopyor tersebut menjadi dasar pemikiran dilakukannya serangkaian penelitian ini.
Melalui dana penelitian dari Badan Litbang Pertanian, Kementerian Pertanian dengan Program KKP3T tahun 2011, KKP3N tahun 2013-2015, serta dari Direktorat Pendidikan Tinggi, Kementerian Pendidikan melalui Program Hi-Link tahun 2012-2014, disusun rencana dan dilakukan serangkaian kegiatan penelitian dari aspek genetika, pemuliaan tanaman sampai molekuler untuk menjawab permasalahan pada kelapa kopyor yang disajikan pada disertasi ini. Masing-masing bab menyajikan hasil penelitian dengan topik yang berkesinambungan dan berisi dasar pemikiran, tujuan, metode serta hasil penelitian, mulai dari pengamatan di lapang sampai penerapannya pada tahapan penelitian selanjutnya di lapang maupun di laboratorium. Pada Bab 1 disertasi ini diuraikan tentang latar belakang, tujuan, manfaaat, kebaruan penelitian dan kerangka berpikir serta garis besar isi disertasi. Bab 2 berisi kajian literatur tentang potensi kelapa kopyor di Indonesia dan perkembangan pemanfaatan plasma nutfah kelapa kopyor. Pada bab ini diuraikan juga hasil penelitian tentang kelapa kopyor dan hasil-hasil penelitian terkait semua aspek yang diteliti pada disertasi ini.
7 Sumenep dan Jember-Jawa Timur, Pati-Jawa Tengah, Purwakarta-Jawa Barat, Tangerang-Banten dan Kalianda-Lampung Selatan. Selain itu dilaporkan bahwa terdapat tanaman kelapa kopyor secara terbatas di daerah Banyuwangi dan Ponorogo Jawa Timur, Purbalingga dan Purwokerto, Jawa Tengah serta di Bogor, Jawa Barat. Secara umum produksi buah kopyor semua aksesi kelapa kopyor yang diamati di tingkat petani relatif rendah yaitu di bawah 25 % untuk kelapa kopyor tipe Genjah dan di bawah 40% untuk kelapa kopyor tipe Dalam. Karakter komponen buah antar aksesi kelapa Dalam kopyor menunjukkan perbedaan yang jauh dengan dan Genjah kopyor, dan memberikan peluang mendapatkan efek heterosis apabila dilakukan persilangan antar aksesi untuk merakit varietas baru kelapa kopyor. Kuantitas endosperma kelapa Genjah kopyor dapat dikategorikan dengan skor 1-6 berdasarkan volume daging buahnya, sedangkan pada kelapa Dalam kopyor memiliki skor 1-9 (Maskromo et al. 2014)
Masalah rendahnya hasil panen buah kelapa kopyor yang diamati di berbagai sentra produksi perlu dicarikan solusinya. Beberapa tindakan perbaikan dapat dilakukan untuk meningkatkan produksi buah kopyor antara lain melalui penyediaan bahan tanaman unggul, perbaikan teknik budidaya dan pengelolaan tanaman. Bahan tanaman unggul dapat diperoleh melalui program pemuliaan dengan merakit varietas unggul yang diikuti dengan penyediaan benih berkualitas. Salah satu metode pemuliaan yang dapat digunakan untuk merakit varietas unggul dan memproduksi benih bermutu kelapa kopyor adalah dengan persilangan terkontrol. Metode ini dilakukan dengan cara menyilangkan antar aksesi atau varietas unggul kelapa kopyor yang telah dievaluasi dan diseleksi berdasarkan karakter yang diinginkan. Sebelum persilangan terkontrol dilakukan diperlukan informasi tentang pola pembungaan tanaman yang akan dijadikan tetua. Potensi jumlah bunga betina sangat menentukan jumlah buah jadi yang dapat dihasilkan pada persilangan terkontrol. Informasi pola pembungaan tanaman tiga varietas kelapa Genjah kopyor diperlukan pada saat digunakan sebagai tetua dalam pelaksanaan persilangan terkontrol, sedangkan informasi perkembangan buah kelapa kopyor perlu dipelajari dalam mendukung pengamatan buah hasil persilangan terkontrol dan verifikasi metode identifikasi panen buah kopyor. Topik yang dibahas pada Bab 4 tersebut juga bertujuan untuk mendukung pelaksanaan topik penelitian peningkatan produksi buah kopyor dan produksi benih true-to-type dengan persilangan terkontrol yang dibahas pada Bab 6 dan studi mekanisme genetik sifat kopyor melalui persilangan terkontrol pada Bab 7.
8 produksi buah kopyor akibat xenia, yaitu pengaruh serbuk sari kelapa normal yang menyebabkan tidak terbentuknya buah kopyor pada tanaman kelapa kopyor heterosigot. Pengamatan terhadap populasi campuran kelapa Genjah kopyor heterosigot (Kk) dengan kelapa normal KK di Kecamatan Tayu, Pati, Jawa Tengah, menunjukkan bahwa terdapat peningkatan hasil panen buah kopyor sebelum dan setelah tanaman kelapa normal KK ditebang. Selain menyebabkan jumlah panen buah kopyor yang rendah, keberadaan tanaman kelapa normal KK juga dapat menyebabkan rendahnya kualitas benih dan bibit yang dihasilkan dari pohon kelapa heterosigot Kk. Pengaruh negatif xenia pohon normal KK akan menurunkan peluang terbentuknya buah dengan genotipe Kk yang dijadikan benih alami oleh petani.
Berdasarkan hasil pengamatan potensi hasil buah kopyor yang dibahas pada Bab 3, pola pembungaan dan perkembangan buah pada pab 4 dan pengaruh fenomena xenia terhadap hasil panen buah kopyor dan kualitas bibit kopyor pada Bab 5, maka dilakukan penelitian untuk meningkatkan jumlah panen buah kopyor dan merakit bibit true-to type kelapa kopyor melalui persilangan terkontrol yang
disajikan pada Bab 6. Penelitian ini dilakukan dengan melakukan penyerbukan terkontrol pada tanaman kelapa Genjah kopyor heterosigot menggunakan serbuk sari kelapa kopyor homosigot hasil kultur embryo. Pengujian dilakukan terhadap dua metode penyerbukan terkontrol yaitu metode1 dengan mengoleskan serbuk sari murni pada putik bunga betina kelapa kopyor dan metode 2 dengan cara menyemprotkan serbuh sari yang ditambahkan talkum pada bunga betina menggunakan botol plastik sebagai alat semprot. Hasil yang diperoleh menunjukkan bahwa penyerbukan terkontrol kelapa kopyor heterosigot Kk menggunakan serbuk sari kelapa kopyor homosigot (kk) mampu meningkatkan hasil panen buah kopyor sampai dengan 50%. Metode penyerbukan terkontrol dengan penyemprotan serbuk sari yang ditambahkan talkum menghasilkan jumlah buah panen yang tidak berbeda nyata dibandingkan dengan metode penebaran serbuk sari murni, namun lebih efisien dalam penggunaan serbuk sari. Benih kopyor heterosigot True-to-Type (Kk) dapat diperoleh melalui penyerbukan
terkontrol antara kelapa kopyor heterosigot (Kk) dengan serbuk sari kelapa kopyor homosigot (kk) sekaligus meningkatkan hasil panen buah kopyor
9 penyebaran tanaman kelapa berbuah kopyor ke daerah lain yang telah terjadi pada kurun waktu sebelumnya. Melalui persilangan terkontrol antar aksesi dan varietas kelapa kopyor dapat menghasilkan buah kopyor dengan persentase yang berbeda antar kombinasi persilangan. Hal ini menunjukkan bahwa sifat kopyor yang eksis pada beragam genetik kelapa tetapi dikendalikan oleg lokus gen yang sama. Hal ini dapat dijadikan dasar penyusunan program pemuliaan mendapatkan varietas unggul baru kelapa kopyor dengan menggabungkan sifat kopyor dari berbagai latar belakang genetik kelapa berbeda.
Pada bagian terakhir disertasi ini yaitu pada Bab 8 dibahas tentang evaluasi pola ko-segregasi antar lokus dari sejumlah marka molekuler yang telah dikembangkan untuk genom kelapa dan menganalisis keterpautan (linkage) antara sifat kopyor dengan marka molekuler yang dievaluasi. Hasil yang diperoleh bahwa Ko-segregasi marka SSR dan SNP yang diuji pada 121 progeni populasi F2 kelapa kopyor menghasilkan 13 lokus yang mengikuti pola segregasi Mendel, sedangkan 7 lokus lainnya menyimpang. Sebanyak empat lokus yang terpaut dan tersebar pada dua group pautan atau kromosom kelapa kopyor yang diuji. Dua lokus SSR dan dua marka SNP teletak pada group pautan dengan sifat kopyor. Marka SNP WRKY_21 yang sangat dekat posisinya dengan sifat kopyor yaitu 9 cM diukuti marka CnCir_J2 dengan jarak 12.2 cM dan marka SNP CnSUS#3 dengan jarak 14.4 cM. Semua lokus yang terdapat pada linkage group dengan sifat kopyor tersebut berpeluang untuk diuji lanjut pada popupasi bersegregasi lainnya seperti popupasi backcross, pseudotestcros dalam mencari marka yang terpaut dengan sifat kopyor. Tujuan akhir dari penelitian pada Bab 8 ini adalah untuk mendapatkan marka molekuler yang dapat digunakan untuk membantu seleksi dini bibit kelapa kopyor.
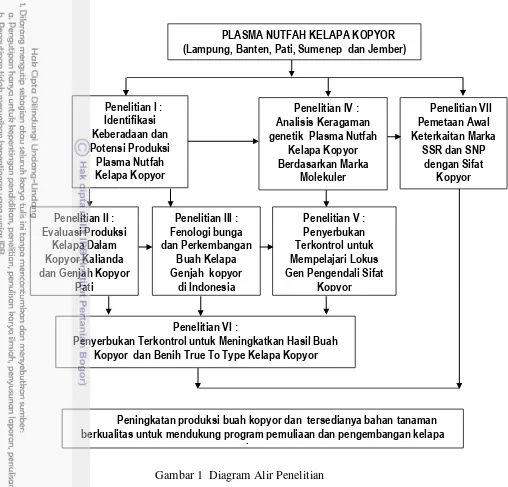
10 Gambar 1 Diagram Alir Penelitian
Penelitian I : Identifikasi Keberadaan dan Potensi Produksi
Plasma Nutfah
Kelapa Kopyor
PLASMA NUTFAH KELAPA KOPYOR (Lampung, Banten, Pati, Sumenep dan Jember)
Penelitian III : Fenologi bunga dan Perkembangan
Buah Kelapa Genjah kopyor
di Indonesia Penelitian II :
Evaluasi Produksi Kelapa Dalam Kopyor Kalianda dan Genjah Kopyor
Pati
Penelitian IV : Analisis Keragaman genetik Plasma Nutfah
Kelapa Kopyor Berdasarkan Marka
Molekuler
Penelitian V : Penyerbukan Terkontrol untuk Mempelajari Lokus Gen Pengendali Sifat
Kopyor
Penelitian VI : Penyerbukan Terkontrol untuk Meningkatkan Hasil Buah
Kopyor dan Benih True To Type Kelapa Kopyor
di Indonesia
Penelitian VII Pemetaan Awal Keterkaitan Marka
SSR dan SNP dengan Sifat
Kopyor
Peningkatan produksi buah kopyor dan tersedianya bahan tanaman berkualitas untuk mendukung program pemuliaan dan pengembangan kelapa
11
2
TINJAUAN PUSTAKA
Kelapa Kopyor
Kelapa kopyor merupakan salah satu jenis kelapa unik, karena karakteristik daging buahnya yang lunak dan berbeda dengan kelapa normal pada umumnya. Jenis kelapa yang memiliki endosperma seperti ini diduga merupakan hasil mutasi alamiah tipe liar kelapa normal seperti kelapa Makapuno di Philipina yang dilaporkan Samonthe et al. (1989). Populasinya di alam sangat sedikit,
hanya ditemui di beberapa daerah sentra kelapa saja.
Penyebaran Kelapa Kopyor
Sampai saat ini belum ada informasi tentang asal tanaman kelapa berbuah kopyor ini, meskipun diketahui tanaman kelapa berbuah kopyor tersebar di Pulau Jawa, dan Sumatera. Di pulau Jawa, kelapa kopyor dapat ditemukan di Jawa Tengah (Novarianto dan Miftahorrachaman, 2000), Tanggerang Jawa Barat (Asmah, 1999), dan Sumenep Jawa Timur (Akuba et al. 2000), sedangkan di Sumatera terdapat Kec. Kalianda, Lampung Selatan (Mahmud, 2000). Kelapa kopyor juga ditanam dan dibudidayakan di daerah tertentu, seperti Banyumas, Klampok, Rembang dan Kebumen.
Kabupaten Sumenep Pulau Madura merupakan daerah sentra kelapa kopyor. Penduduk setempat menyebutnya dengan nama Kelapa Puan. Dalam suatu populasi tanaman, jumlah kelapa puan sebanyak 2 %. Kelapa Puan ini sebagian besar tersebar secara sporadis di Kecamatan Batang-batang, Dungkek dan Gapura. Produksi Kelapa Dalam Puan Sumenep di tiga Kecamatan tersebut diperkirakan rata-rata 1000 butir per minggu (Akuba et al. 2000).
Karakteristik Tanaman Kelapa Kopyor
Secara morfologi fenotipe pohon kelapa berbuah kopyor sulit dibedakan dari kelapa normal di sekitarnya. Pengamatan morfologi secara menyeluruh belum pernah dilakukan, sehingga belum ada laporan tentang karakteristik morfologi tanaman kelapa berbuah kopyor, kecuali sifat endosperma yang berbeda dengan kelapa normal. Kepastian tanaman kelapa berbuah kopyor diketahui setelah buahnya dipanen.
Salah satu cara sederhana untuk mengetahui buah kelapa kopyor dengan mengguncang buah tersebut. Pada saat diguncang buah kopyor akan berbunyi seperti kaleng berisi pasir yang diguncang (bunyi gemericik). Daging buah telah hancur dan seringkali telah terlepas dari tempurung sehingga bercampur dengan air kelapa. Embrio kelapa juga telah terlepas dari daging buah. Kelapa kopyor yang ditemukan di Sumenep memiliki tiga tipe berdasarkan karakteristik daging buah. Tipe pertama, daging buah telah hancur disebut buah puan atau kopyor.
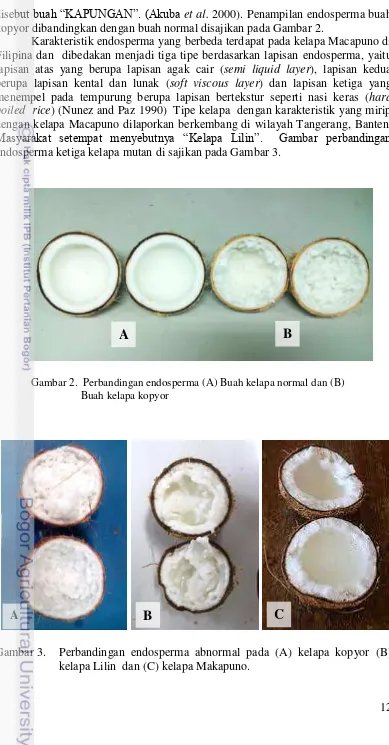
12 disebut buah “KAPUNGAN”. (Akuba et al. 2000). Penampilan endosperma buah kopyor dibandingkan dengan buah normal disajikan pada Gambar 2.
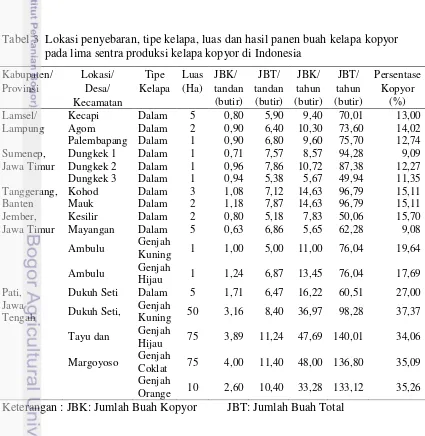
Karakteristik endosperma yang berbeda terdapat pada kelapa Macapuno di Filipina dan dibedakan menjadi tiga tipe berdasarkan lapisan endosperma, yaitu lapisan atas yang berupa lapisan agak cair (semi liquid layer), lapisan kedua berupa lapisan kental dan lunak (soft viscous layer) dan lapisan ketiga yang
menempel pada tempurung berupa lapisan bertekstur seperti nasi keras (hard boiled rice) (Nunez and Paz 1990) Tipe kelapa dengan karakteristik yang mirip dengan kelapa Macapuno dilaporkan berkembang di wilayah Tangerang, Banten.
Masyarakat setempat menyebutnya “Kelapa Lilin”. Gambar perbandingan
[image:42.595.90.479.78.823.2]endosperma ketiga kelapa mutan di sajikan pada Gambar 3.
Gambar 2. Perbandingan endosperma (A) Buah kelapa normal dan (B) Buah kelapa kopyor
Gambar 3. Perbandingan endosperma abnormal pada (A) kelapa kopyor (B) kelapa Lilin dan (C) kelapa Makapuno.
A
B
13 Pohon kelapa berbuah kopyor yang terdapat di lapang atau yang sekarang dikembangkan petani diduga memiliki genotipe heterosigot. Buah kelapa normal dari pohon tersebut jika ditanam berpeluang tumbuh menjadi tanaman kelapa berbuah kopyor, dengan persentase menghasilkan buah kopyor sekitar 1 - 5%, bergantung pada genotipe tepung sari yang menyerbuki bunga betina. Buah kopyor memiliki gen resesif kopyor homosigot sehingga tidak tumbuh menjadi tanaman baru (Toruan 1998).
Bersatunya gen kopyor (k) menyebabkan endosperma berkembang tidak normal, sehingga tidak mampu mendukung perkecambahan embrio (Santos 1999). Untuk mendapatkan tanaman yang memiliki sifat kopyor bergenotipe homosigot, maka embrio dari buah homosigot tersebut diselamatkan lewat kultur embrio. Sifat kopyor secara genetika dikendalikan oleh genotipe tetua betina dan tepung sari (tetua jantan) seperti diitunjukkan pada Tabel 1 dan Tabel 2.
Apabila buah diharapkan memiliki endosperma gen resesif kopyor (kkk), maka bunga betina yang mengandung sepasang alel k (kk), harus diserbuki dengan tepung sari yang memiliki gamet k. Pada tanaman kelapa kopyor hasil
kultur embrio yang memiliki genotipe homosigot resesif seharusnya menghasilkan 100% kopyor, apabila dapat menyerbuk sendiri atau terjadi penyerbukan antar tanaman homosigot. Apabila dibiarkan serbuk sari berasal dari tanaman yang bukan kopyor, maka kemungkinan buah kopyor hanya 0% - 67%.
Tabel 1 Kemungkinan genotipe dan fenotipe embryo yang terbentuk pada segregasi persilangan antara tanaman kopyor heterosigot Kk
Jantan
Betina Inti sperma (inti generatif -1n)
Inti sel telur K k
K KK
( Normal Homosigot)
Kk
(Normal Heterosigot)
k Kk
( Normal Heterosigot)
kk
(Kopyor Homosigot)
Tabel 2 Kemungkinan genotipe dan fenotipe endosperma yang terbentuk pada segregasi persilangan antara tanaman kopyor heterosigot Kk
Jantan
Betina Inti sperma(inti generatif - 2n)
Inti polar K k
KK KKK
(Normal Homosigot)
KKk
(Normal Heterosigot)
kk Kkk
(Normal Heterosigot)
kkk
14 Dalam rangka pengembangan kelapa kopyor, BalitPalma Manado, Puslitbiotek Bogor, IPB Bogor dan Universita Muhamadiayah Purwokerto sudah melakukan penelitian perbanyakan bibit kelapa kopyor melalui kultur embrio. Balai Penelitian Perkebunan Ciomas Bogor telah melakukan metode perbanyakan tersebut sejak beberapa tahun yang lalu. Saat ini cukup banyak koleksi tanaman kelapa kopyor yang sudah mencapai usia produktif. Bibit kelapa kopyor hasil kultur embryo ini dijual dengan harga Rp. 500.000 - Rp.1.000.000,-/bibit.
Pola Pembungaan Kelapa
Tanaman kelapa memiliki tipe bunga monoeceus dan terbuka yaitu bunga
jantan dan betina teletak pada satu tanaman dengan posisi terpisah. Bunga betina terletak pada pangkal spikelet, sedangkan bunga jantan pada bagian atas spikelet. Hal tersebut memungkinkan terjadinya penyerbukan silang antar tanaman kelapa (Perera et al. 2010). Berdasarkan pola pembungaan kelapa Sangare et al. (1978)
membagi dalam beberapa tipe penyerbukan.
Berdasarkan morfologi bunganya, kelapa dikatagorikan sebagai tanaman berumah satu (monoecieus) karena bunga jantan dan bunga betina berada satu
pohon. Pada satu tandan bunga terdapat bunga betina yang terletak pada pangkal
spikelet, sedangkan bunga jantan terletak pada ujung spikelet.Spikelet adalah
jari-jari tempat melekatnya bunga jantan dan bunga betina. Jumlah bunga jantan sangat banyak (ribuan jumlahnya), sedangkan bunga betina berjumlah 20 – 100 butir.
Rangkaian bunga jantan dan betina yang bisanya disebut mayang diselaputi oleh seludang bunga. Bunga kelapa muncul setiap bulan pada setiap ketiak daun yang terbentuk, sehingga jumlah bunga setahun berkisar antara 12 - 16 tandan, tergantung jenis dan kesuburan tanamannya. Bunga jantan lebih dahulu terbuka, cenderung bersamaan dengan pecahnya seludang/tandan bunga. Terbukanya bunga jantan dimulai dari bagian yang terletak pada ujung kemudian ke bagian pangkal spikelet. Setelah bunga jantan terbuka, serbuk sarinya terlepas
dan bunga jantannya jatuh terlepas dari spikelet. Proses terbukanya bunga jantan,
sampai terlepasnya dari spikelet ini berlangsung satu hari saja. Waktu mulai pecahnya seludang bunga sampai jatuhnya bunga jantan terakhir dari spikelet
disebut fase bunga jantan. Fase bunga jantan ini berlangsung kurang lebih 20 hari,
dan bervariasi menurut jenis kelapa dan dipengaruhi oleh musim. Sedangkan bunga betina mulai reseptif beberapa hari setelah seludang bunga terbuka. Masa
reseptif ditandai dengan keluarnya cairan nectar atau madu pada stigma (ujung
atas bunga betina). Waktu mulai reseptif bunga betina pertama sampai
berakhirnya masa reseptif bunga betina terakhir disebut fase bunga betina. Lama fase bunga betina dipengaruhi oleh kondisi lingkungan dan tergantung pada jenis kelapa. Pada jenis kelapa Dalam fase bunga betina berlangsung 3 – 5 hari, sedangkan untuk kelapa Genjah 8 – 15 hari Santos et al. (1997).
Selanjutnya menurut Sangare et al. (1978), berdasarkan pola fase bunga jantan dan betina dikenal 4 klasifikasi pola penyerbukan pada tanaman kelapa yaitu :
15
2. Indirect autogami yaitu tidak adanya overlaping antara fase bunga jantan dan bunga betina pada satu tandan, tapi terjadi overlaping antara bunga betina pada tandan tersebut dengan bunga jantan tandan berikutnya.
3. Direct autogami yaitu adanya overlaping secara penuh antara fase bunga jantan dan fase bunga betina dalam satu tandan. Hal ini yang menyebabkan terjadinya penyerbukan sendiri pada satu tandan yang sama, karena fase bunga jantan dan betina terjadi bersamaan.
4. Semi direct autogami yaitu terjadinya overlaping antara fase bunga betina dengan fase bunga jantan pada tandan yang sama, juga pada bunga jantan tandan berikutnya.
Terjadinya overlaping bunga betina dengan bunga jantan berikutnya tergantung pada jumlah tandan yang terbentuk per tahun, yang dipengaruhi oleh kesuburan tanaman dan kondisi lingkungan seperti jumlah bulan kering (musim panas) (Sangare et al. 1978). Selain pada berpengaruh pada jumlah tandan yang
terbentuk, pola curah hujan pada lokasi pertanaman sangat mempengaruhi jumlah bunga betina yang terbentuk (Perera et al. 2010).
Beberapa penelitian tentang biologi bunga dan pola pembungaan kelapa telah dilakukan. Hasil pengamatan biologi bunga beberapa kelapa Genjah di Parungkuda Jawa Barat, menunjukkan bahwa kelapa Genjah Kuning Nias memiliki jumlah bunga betina paling banyak dibandingkan kelapa Genjah lainnya. Genjah Kuning Bali memiliki ukuran bunga betina lebih besar sedangkan Genjah Hijau Nias memiliki fase reseptif bunga betina yang cepat dan fase bunga jantan lebih lama Marfuati et al. (1988). Menurut Palilu dan Rompas (1988), masa reseptif (subur) bunga betina dipengaruhi oleh iklim, kondisi kesuburan tanaman,
jumlah bunga betina dan jenis kelapa. Hasil pegamatannya menunjukkan bahwa awal fase bunga betina kelapa Genjah adalah 2-7 hari sesudah seludang terbuka dan masa reseptif (fase bunga betina) 10 – 14 hari, sehingga penyerbukan dapat dilakukan pada selang waktu tersebut.
Persilangan pada Tanaman Kelapa
Hibridisasi merupakan salah satu upaya untuk meningkatkan produksi kelapa. Tahapan pada proses hibridisasi meliputi penyediaan serbuk sari, emaskulasi, dan polinasi/penyerbukan (Randriani dan Saefudin 1993). Persilangan secara alami dilakukan oleh serangga atau angin, sedangkan persilangan buatan dilakukan dengan mengembuskan serbuk sari pada bunga betina yang sedang reseptif.
Metode hibridisasi kelapa ditujukan untuk merakit kelapa hibrida. Persilangan dua tetua yang memiliki gen-gen dominan yang diinginkan, turunannya akan memiliki ketegaran hibrida (heterosis tinggi). Ketegaran hibrida (heterosis) didefinisikan sebagai meningkatnya ketegaran (vigor) dari turunan (F1) melebihi kedua tetuanya .
16 berikut: kandungan minyak tinggi, resisten terhadap penyakit busuk pucuk dan gugur buah, hasil kopra tinggi per unit areal dengan pemeliharaan sederhana, toleran terhadap lahan pasang surut, toleran terhadap kekeringan, kandungan asam laurat tinggi di dalam minyak, dan kandungan protein tinggi di dalam daging buah Persilangan kelapa hibrida terdapat beberapa pola, yaitu Genjah x Dalam, Dalam x Genjah, Dalam x Dalam dan Genjah x Genjah. Di Indonesia yang sudah dilakukan adalah perakitan kelapa hibrida Dalam x Dalam dan Genjah x Dalam. Varietas kelapa hibrida yang sudah dilepas sejak tahun 1984 sampai tahun 2009 adalah empat hibrida Dalam x Dalam, dan lima kelapa hibrida Genjah x Dalam. (Novarianto et al.1984; Novarianto et al. 1992; Tenda,et al. 1996). Di samping
itu, pelepasan varietas kelapa unggul melalui seleksi dan pemumian telah berhasil pada empat kelapa tipe Genjah dan 10 kelapa tipe Dalam.
Pada kelapa kopyor persilangan yang dapat dilakukan bertujuan untuk meningkatkan produksi buah kopyor, merakit tanaman kelapa kopyor unggul dan mempelajari pola pewarisan sifat serta gen pengendali sifat kopyor pada kelapa.
Marka Morfologi dan Molekuler
Keanekaragaman genetika tanaman merupakan modal dasar dalam program pemuliaan tanaman. Keanekaragaman genetika tanaman secara alami dapat terjadi karena terjadi persilangan seksual, mutasi, dan isolasi geografi. Untuk mendapatkan materi genetik tanaman, perlu dilakukan eksplorasi, karakterisasi, koleksi, seleksi, dan evaluasi. Dalam kegiatan evaluasi, karakterisasi dan seleksi diperlukan dasar atau marka yang dapat dijadikan ciri pembeda antar individu