KONFORMASI ENZIM LIPASE T1 PADA PELARUT
ALKOHOL DIKAJI DENGAN TEKNIK SIMULASI
DINAMIKA MOLEKUL
AYU MELISA PUTRI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Konformasi Enzim
Lipase T1 Pada Pelarut Alkohol Dikaji Dengan Teknik Simulasi Dinamika
Molekul adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, November 2016
RINGKASAN
AYU MELISA PUTRI. Konformasi Enzim Lipase T1 Pada Pelarut Alkohol Dikaji Dengan Teknik Simulasi Dinamika Molekul Enzim. Dibimbing oleh TONY IBNU SUMARYADA dan SETYANTO TRI WAHYUDI.
Lipase T1 (Geobacilus zalihae) secara ekstensif dipelajari dalam produksi biodiesel. Secara umum enzim lipase dapat dimanfaatkan dalam pembuatan deterjen, surfaktan, pangan, produk farmasi, kosmetik, industri kertas, dan nutrisi. Tujuan penelitian ini adalah untuk mempelajari kestabilan konformasi secara komputasi enzim Lipase T1 pada pelarut metanol dan etanol. Simulasi dinamika molekul dilakukan dengan mengamati energi konformasi enzim lipase T1 pada pelarut yang digunakan. Simulasi dinamika molekul lipase T1 dilakukan menggunakan perangkat lunak Ambertools12 pada suhu 300 K.
Parameter yang digunakan untuk membandingkan stabilitas enzim lipase T1 dalam berbagai pelarut adalah dengan melihat nilai RMSD (Root Mean Square Deviation), SASA (Solvent Accessible Surface Area), Rg (Radius of Gyration), RMSF (Root Mean Square Fluctuation), Energi Konformasi dan Struktur Sekunder. Simulasi dinamika molekul yang dilakukan pada ketiga pelarut menunjukkan bahwa lipase pada etanol dan metanol lebih kaku atau kurang fleksibel karena lebih banyak saltbridge yang terikat kuat di etanol dan metanol dibanding air. Lipase harus fleksibel pada saat aktif. Sehingga dapat disimpulkan keadaan yang terlalu kaku akan mengurangi fungsi lipase. Enzim lipase Geobacilus zalihae butuh fleksibilitas dalam berbagai pelarut.
Untuk analisis struktur sekunder lipase T1 pada pelarut etanol dan metanol tidak terjadi banyak perubahan struktur beta-sheet dan alpa-helix yang signifikan atau tidak terjadi kerusakan struktur enzim lipase T1 pada pelarut yang digunakan. Pelarut organik tidak begitu mempengaruhi komposisi alpahelix dan beta-sheet nya. Diharapkan hasil penelitian ini dapat memberikan saran rancangan penelitian yang dapat diimplementasikan pada laboratorium basah (wet experiment).
IBNU SUMARYADA and SETYANTO TRI WAHYUDI.
T1 lipase (Geobacilus zalihae) extensively studied in the production of biodiesel. In general, lipase enzymes can be utilized in the manufacture of detergents, surfactants, food, pharmaceutical, cosmetic, paper industry, and nutrition. The purpose of this research is to study conformational stability T1 Lipase enzyme in methanol and ethanol via computational approach. Molecular dynamics simulations were performed observing the energy conformation of the T1 lipase enzyme in various alcohol solvent. Molecular dynamics simulations of T1 lipase enzyme were performed using the software Ambertools12 at 300 K for 10 ns
For the stability analysis of T1 lipase enzyme, the output such as RMSD (Root Mean Square Deviation), SASA (Solvent Accessible Surface Area), Rg (Radius of Gyration), RMSF (Root Mean Square Fluctuation), Energy conformation and Secondary structure were analyzed. The results have shown that in ethanol and methanol lipase enzyme was more rigid due to more fastened saltbridge as compared to the water solvent. The rigidity will affect the functionality of the enzyme as too rigid will reduce the function of the enzyme.
For the analysis of the secondary structure of T1 lipase in ethanol and methanol does not happen a lot of changes in the structure of the beta-sheet and alpha-helix significant or structural failure lipase T1 on the solvent used. Organic solvents are not so affect the composition alpahelix and its beta-sheet. We hope this research can provide advice study designs that can be implemented on a wet lab (wet experiment).
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biofisika
DIKAJI DENGAN TEKNIK SIMULASI
DINAMIKA MOLEKUL
AYU MELISA PUTRI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
KONFORMASI ENZIM LIPASE T1 PADA PELARUT
ALKOHOL DIKAJI DENGAN TEKNIK SIMULASI
PRAKATA
Puji dan syukur penulis panjatkan atas kehadirat Allah SWT yang telah melimpahkan Rahmat dan Karunia-Nya, sehingga penulis dapat menyelesaikan tesis ini dengan judul “Konformasi Enzim Lipase T1 Pada Pelarut Alkohol Dikaji
Dengan Teknik Simulasi Dinamika Molekul”. Tesis ini merupakan salah satu syarat untuk mendapatkan gelar Magister Sains pada Program Studi Biofisika, Sekolah Pascasarjana, Institut Pertanian Bogor.
Penulis mengikuti pendidikan di Sekolah Pascasarjana IPB ini tidak lepas dari dukungan berbagai pihak. Penulis menyampaikan banyak terima kasih dan penghargaan yang tinggi kepada
1. Dr Tony Ibnu Sumaryada dan Dr Setyanto Tri Wahyudi SSi MSi selaku komisi pembimbing atas kesediaan waktu untuk membimbing, memberikan arahan dan masukan selama penyusunan tesis ini.
2. Dr Mersi Kurniati SSi MSi selaku ketua Program Studi Biofisika.
3. Ayahanda Zulkifli. R dan Ibunda Yetty Susi, beserta seluruh keluarga atas doa, motivasi, finansial dan semangat selama penulis menempuh studi.
4. Bapak dan Ibu staf pengajar, staf administrasi Program Studi Biofisika yang telah banyak membantu dan kerjasamanya yang baik selama penulis menempuh studi.
5. Rekan rekan Forum Mahasiswa Pasca Sarjana yang telah banyak berbagi dalam suka dan duka serta selalu memberikan dukungan kepada penulis saat proses penelitian serta penulisan tesis ini .
Penulis menyadari bahwa tesis ini masih jauh dari sempurna. Semoga karya ilmiah ini membawa manfaat bagi seluruh civitas IPB khususnya dan masyarakat Indonesia umumnya.
Bogor, November 2016
DAFTAR ISI
Lipase Geobacilus zalihae 3
Faktor yang Mempengaruhi Aktivitas Lipase 4
Simulasi Dinamika Molekul 5
Gaya Intamolekul 7
3 METODOLOGI 7
Bahan dan Alat 7
Waktu dan Tempat Penelitian 8
DAFTAR GAMBAR
8 Grafik energi elektrostatik pasangan jembatan garam Glu132-Lys84 169 Posisi residu Glu132-Lys84 17
10 Grafik energi elektrostatik pasangan jembatan garam Glu149-Arg134 18
11 Posisi residu Glu149-Arg134 19
12 Grafik energi elektrostatik pasangan jembatan garam Asp205-Arg92 20
13 Posisi residu Asp205-Arg92 21
14 Grafik energi elektrostatik pasangan jembatan garam Glu250-Arg330 22
15 Posisi residu Glu250-Arg330 23
16 Analisis struktur sekunder selama simulasi pada suhu 300 K selama 10 ns pada (a) air (b) metanol (c) etanol ; α-helix (Ungu), β-sheet (Kuning), Turn (Biru) dan Coil (Hitam) ; α-helix (Ungu), β-sheet
(Kuning) 3-10helix (Biru), Turn (Hijau) dan Coil (Putih) 24
DAFTAR LAMPIRAN
1 Diagram Alir Penelitian 31
2 Pasangan jembatan garam yang muncul selama simulasi dalam pelarut
air, metanol, dan etanol selama 10 ns dan suhu 300 K 32 3 Karakteristik Residu yang terdapat pada protein 2DSN 33 4 Residu penyusun struktur sekunder protein 2DSN 43 5 Nilai energi rata-rata enzim 2DSN pada ketiga pelarut 45
6 Daftar 14 pasang jembatan garam yang mucul pada semua pelarut 46 7 Pasangan jembatan garam yang terdapat pada pelarut air, etanol dan
1 PENDAHULUAN
Latar Belakang
Enzim adalah golongan protein yang disintesis oleh sel hidup dan mempunyai fungsi penting sebagai katalisator dalam setiap reaksi metabolisme yang terjadi pada organisasi hidup. Enzim juga merupakan biokatalisator yang menunjang berbagai proses industri. Hal ini disebabkan enzim mempunyai efisiensi dan efektifitas yang tinggi, reaksinya tidak menimbulkan produk samping, serta dapat digunakan berulangkali dengan teknik amobilisasi (Lehninger 1995).
Salah satu jenis enzim yang mempunyai peran penting dalam pertumbuhan bioteknologi adalah enzim lipase. Enzim ini memiliki sifat khusus dapat memecahkan ikatan ester pada lemak dan gliserol. Selain itu, lipase mempunyai kemampuan mengkatalis reaksi organik baik didalam media berair maupun dalam media non air (Sumarsih 2004). Enzim lipase sangat berperan dalam pemisahan asam lemak dan pelarutan noda minyak pada alat industri agar minyak dapat dilarutkan dalam air. Beberapa reaksi yang dikatalisis oleh enzim lipase diantaranya adalah reaksi hidrolisis, alkoholisis, esterifikasi dan interesterifikasi (Dosanjh dan Kaur 2002).
Beberapa lipase yang dihasilkan oleh bakteri dari genus geobacillus seperti G. stearothermophilus, G. thermocatenulatus, G. thermoleovorans, G. kaustophilus, G. thermoglucosidasius, G. thermodenitrificans, G. subterraneus, G. uzenensis, G. caldoxylosilyticus, G. toebii, G. vulcani, G. lituanicus, G. tepidamans, G. gargensis , G. jurassicus, G. caldoproteolyticus, G. pallidus dan G. debilis dengan suhu pertumbuhan berkisar 35-78 ° C. Anggota dari genus ini
tersebar luas di berbagai wilayah geografis termofilik dan mesofilik di bumi seperti ladang minyak, jerami kompos, lubang hidrotermal atau tanah (Nazina et al. 2001).
Lipase banyak digunakan dalam pengolahan lemak dan minyak, detergen, pengolahan makanan, sintesis bahan kimia, farmasi, sintesis kertas, produksi kosmetik dan juga industri biodiesel. Biodiesel adalah salah satu bahan bakar alternatif yang dapat diperbaruhi, ramah terhadap lingkungan, tidak mempunyai efek terhadap kesehatan dan dapat dipakai sebagai bahan bakar kendaraan bermotor. Lipase dapat mengkatalisis reaksi transesterifikasi pada sintesis biodiesel dengan reagent yang sesuai dan kehadiran air yang terbatas. Reaksi transesterifikasi yang dikatalisis dengan menggunakan bantuan enzim lipase memiliki beberapa kelebihan dalam peningkatan kuantitas dan kualitas konversi minyak nabati ke biodiesel, yakni tanpa busa, hasil konversi tinggi, produk yang dihasilkan mudah dimurnikan, bahkan dapat dilakukan tanpa pemurnian, gliserol mudah dipisahkan (Fukuda et al. 2001).
minyak nabati berasal dari sumberdaya yang dapat diperbaruhi. Beberapa bahan baku untuk pembuatan biodiesel antara lain kelapa sawit, kedelai, bunga matahari, dan jarak pagar (Haryahto 2002).
Biodiesel telah diproduksi secara komersial melalui reaksi transesterifikasi minyak nabati dengan metanol menggunakan katalis alkali. Tetapi katalis alkali ini mempunyai beberapa kelemahan, seperti terjadinya reaksi pembentukan sabun akibat bereaksinya katalis (logam alkali) dengan mudah. Alkohol yang paling umum digunakan adalah metanol dan etanol, terutama metanol, karena harganya murah dan reaktifitasnya paling tinggi (sehingga reaksinya disebut metanolisis). Produk yang dihasilkan (jika menggunakan metanol) lebih sering disebut sebagai metil ester asam lemak (fatty acid methyl ester/FAME) daripada biodiesel (Knothe et al. 2005), sedangkan jika etanol yang digunakan sebagai reaktan, maka akan diperoleh campuran etil ester asam lemak (fatty acid ethyl ester/FAEE) (Lam et al. 2010). Dengan minyak berbasis bio (minyak nabati) maka hubungan stoikiometrinya memerlukan 3 mol alkohol per mol TAG (3:1), tetapi reaksi biasanya membutuhkan alkohol berlebih berkisar 6:1 hingga 20:1, tergantung pada reaksi kimia untuk transesterifikasi katalis basa dan 50:1 untuk transesterifikasi katalis asam (Zhang et al. 2003).
Pada dasarnya teknologi produksi biodiesel yang diterapkan dalam skala industri melalui reaksi transesterifikasi dari trigliserida (yang terdapat pada minyak nabati) dengan alkohol (umumnya metanol) menggunakan katalis basa (alkali). Teknologi ini banyak dikembangkan dikarenakan proses ini relatif lebih murah. Namun, penggunaan katalis alkali ini mempunyai beberapa kelemahan diantaranya proses pemurnian produk yang bercampur homogen sehingga relatif sulit. Selain itu, katalis alkali tersebut akan bereaksi dengan trigliserida sehingga terjadi reaksi samping yaitu reaksi saponifikasi (penyabunan). Reaksi saponifikasi ini akan mengakibatkan proses pemisahan produk semakin sulit. Kelemahan lain dari teknologi ini adalah perlunya sejumlah asam untuk penetralan katalis basa yang ikut dalam aliran produk sehingga akan berdampak terhadap lingkungan (Furuta 2006).
Untuk mengatasi masalah tersebut, diperlukan katalis yang tidak bercampur homogen dan mampu mengarahkan reaksi secara spesifik guna menghasilkan produk yang diinginkan tanpa reaksi samping. Belakangan ini, riset sintesis biodiesel menggunakan enzim lipase semakin banyak dilakukan. Enzim lipase yang bisa menjadi biokatalis dalam sintesis biodiesel tersebut mampu memperbaiki kelemahan katalis alkali, yakni tidak bercampur homogen sehingga pemisahannya lebih mudah. Akan tetapi penggunaan lipase sebagai biokatalis menyisakan satu persoalan. Lingkungan beralkohol seperti metanol menyebabkan lipase terdeaktivasi secara cepat dan stabilitas enzim tersebut dalam mengatalisis reaksi menjadi buruk. Oleh karena itu, diperlukan pelarut yang dapat membuat enzim lipase teraktivasi secara cepat dan dapat menjaga stabilitas enzim.
Analisis hasil simulasi tersebut dapat dimanfaatkan oleh peneliti secara in-vitro sebagai bahan pertimbangan untuk menentukan residu asam amino yang akan dimutasi pada rekayasa genetik (Ashutosh et al. 2014). Penelitian ini bertujuan untuk menganalisis kestabilan enzim lipase G. zalihae wild type (Kode PDB : 2DSN) pada pelarut organik dengan mengamati peristiwa unfolding yang terjadi untuk mendapatkan enzim yang memiliki stabilitas lebih tinggi terhadap pelarut organik dan aktivitas yang lebih tinggi pada suhu tinggi. Selanjutnya membandingkan hasil analisis kestabilan enzim dengan membandingkan parameter Root Mean Squared Deviation (RMSD), Radius of Gyration (Rg), Solvent Accessible Surface Area (SASA), Root Mean Squared Fluctuation (RMSF) , Struktur Sekunder, dan analisis energi konformasi.
Tujuan penelitian
Untuk mempelajari kestabilan konformasi Lipase T1 pada pelarut metanol dan etanol dengan metode komputasi.
Ruang Lingkup Penelitian
Penelitian ini berupa simulasi dinamika molekuler pada pelarut alkohol untuk menganalisis energi konformasi dari enzim lipase Geobacilus zalihae. Simulasi dinamika molekuler untuk mengamati kestabilan enzim dalam berbagai pelarutdilakukan pada temperatur 300 K selama 10 ns
Manfaat Penelitian
Memberikan informasi ilmiah mengenai pemanfaatan simulasi Dinamika Molekul untuk melihat konformasi enzim Lipase T1 pada pelarut etanol dan metanol.
2 TINJAUAN PUSTAKA
Lipase Geobacilus zalihae
Geobacillus zalihae (za.li'ha.e. NL n. Zalihae dari Zaliha gen.). Spesies baru
Gambar 1 Representasi struktur tersier enzim lipase Enzim G. zalihae
Enzim lipase T1 memiliki jumlah asam amino 774 residu dan memiliki 2 chain yakni chain A dan chain B. Enzim lipase T1 memiliki struktur sekunder utama 13 α-helix (warna ungu), 7 β-sheet (warna kuning). Representasi struktur sekunder secara new cartoon menggunakan program VMD dapat dilihat pada Gambar 1.
Distribusi geografis geobacillus cukup banyak seperti ladang minyak, jerami kompos, lubang hidrotermal atau tanah. Para anggota dari genus geobacillus memiliki suhu pertumbuhan berkisar 35 hingga 78 ° C dan berisi asam lemak jenuh iso-bercabang (iso-15: 0, iso-16: 0 dan iso-17: 0) sebagai asam lemak utama. Para anggota Geobacillus memiliki kesamaan dalam urutan gen 16S rRNA (96,5 hingga 99,2 persen). Enzim termopilus yang stabil cocok untuk aplikasi industri (Nazina TN et al. 2001).
Dua puluh sembilan produsen lipase disaring dan diisolasi dari limbah pabrik kelapa sawit di Malaysia. Dari jumlah tersebut, dipilih T1 untuk studi lebih lanjut sebagai aktivitas lipase yang relatif lebih tinggi terdeteksi secara kuantitatif. T1 lipase mentah menunjukkan suhu optimum tinggi 70°C dan juga stabil sampai 60°C tanpa kehilangan aktivitas enzim kasar (Perry JJ et al. 1997).
Faktor yang Mempengaruhi Aktivitas Lipase
Aktivitas enzim lipase dipengaruhi oleh beberapa faktor, diantaranya adalah temperatur, air, pelarut organik, konsentrasi enzim dan substrat. Peningkatan temperatur akan menambah kecepatan reaksi kimia akibat peningkatan jumlah energi bagi molekul reaktan, sehingga tumbukan antara molekul persatuan waktu lebih produktif. Temperatur yang terlalu tinggi akan menyebabkan enzim sangat mudah terinaktifasi karena sifat enzim yang mudah terdenaturasi. Inaktivasi enzim pada temperatur tinggi disebabkan oleh dua hal yaitu adanya pembukaan partial struktural molekul enzim dan perubahan struktur primer enzim karena adanya perubahan atau kerusakan molekul-molekul asamasam amino tertentu (Saktiwansyah 2001).
kondisi air terbatas, lipase masih menunjukkan aktifitasnya pada temperatur 100oC. Hal ini berarti struktur lipatan protein enzim dalam media organik masih terjaga (August 2000).
Pelarut organik berhubungan dengan kenaikan “rigiditas” molekul enzim yang disebabkan oleh konstanta dielektrika yang rendah yang akan memperkuat dan menstabilkan struktur enzim secara keseluruhan (August 2000). Penggunaan enzim untuk sintesis dalam pelarut organik memberikan beberapa keuntungan antara lain kelarutan substrat organik dan enzim dalam pelarut organik lebih tinggi dibandingkan dengan air, kestabilan enzim meningkat dan mudah mengisolasi produk. Penggunaan pelarut organik juga mempunyai kelemahan seperti residu pada produk akhir, toksisitas bagi makhluk hidup, mahal dan mudah terbakar (Saktiwansyah 2001).
Studi mengenai aktivitas katalitik enzim dalam pelarut organik menunjukkan bahwa pelarut yang baik yang dapat mempertahankan aktivitas dan stabilitas enzim adalah pelarut yang bersifat hidrofobik. Pelarut ini pada kondisi bebas air sangat baik untuk menjaga aktivitas enzim pada temperatur tinggi.
Faktor yang perlu diperhatikan dalam pemilihan pelarut organik adalah kelarutan substrat dan produk dalam pelarut, reaktivitas pelarut, densitas, viskositas dan tegangan permukaan, toksisitas pelarut, mudah tidaknya terbakar, harga serta masalah limbah bahan tersebut dan pembuangan sisa pelarut tersebut ke alam (August 2000).
Simulasi Dinamika Molekul
Dinamika molekuler merupakan suatu metode untuk menyelidiki struktur dari zat padat, cair dan gas. Umumnya dinamika molekuler menggunakan teknik persamaan hukum newton dan mekanika klasik. Dinamika molekuler pertama kali diperkenalkan oleh Alder dan Wainwright pada akhir tahun 1950-an, metode ini digunakan untuk mempelajari interaksi pada bola keras. Dari studi tersebut mereka mempelajari mengenai sifat sebuah cairan sederhana. Pada tahun 1964,
Rahman melakukan simulasi pertama menggunakan energi potensial terhadap cairan argon. Dan di tahun 1974, Rahman dan Stillinger melakukan simulasi dinamika molekuler pertama menggunakan sistem yang realistic yaitu simulasi dengan menggunakan air. Kemudian pada tahun 1977, muncul pertama kali simulasi terhadap protein yaitu simulasi sebuah inhibitor enzim tripsin bovine z
pancreas (BPTI) (Astuti dan Mutiara 2009). Tujuan utama dari simulasi dinamika molekuler yang pertama adalah untuk memberi prediksi apa yang akan terjadi apabila parameter eksternal diubah, menghasilkan trajektori molekul dalam jangka waktu terhingga, menjadi jembatan antara teori dan hasil eksperimen, dan untuk memudahkan para ahli kimia melakukan simulasi yang tidak bisa dilakukan dalam laboratorium.
Dinamika Molekul adalah metode untuk mengevaluasi gerakan yang dihasilkan dari interaksi atom dengan mengintegrasikan persamaan gerak. Secara umum molecular dynamics dapat digunakan untuk mengevaluasi gerak dari setiap sistem partikel. Pendekatan mudah adalah dengan mengintegrasikan persamaan gerak yang diperoleh dalam hukum kedua Newton :
dimana (t) dan menunjukkan posisi dan gaya yang bekerja pada atom i dalam arah x pada waktu t dan mi adalah massa atom i (Wolf 2009).
Nilai energi berhubungan dengan temperatur melalui momentum partikel
(2)
dimana �� adalah jumlah konstrain dan 3N-Nc = N df adalah jumlah derajat kebebasan. Temperatur rata-rata � identik dengan temperatur makroskopik (Ruhle 2007)
Simulasi dilakukan menggunakan NVT ensemble (Jumlah molekul, Volume dan Temperatur tetap). Distribusi Boltzman untuk ensembel NVT kanonik dalam NAMD dihitung menggunakan persamaan Langevin
(3)
dimana m adalah massa, � kecepatan, � gaya, � jarak, � koefisien gesek. �b konstanta boltzman, � temperatur dan (�) adalah proses acak Gaussian (Philips 2005).
Prinsip simulasi dinamika molekul adalah menjadikan perubahan koordinat atom sebagai fungsi dari waktu dan menghitung pergerakan atom-atom yang ada di dalam molekul. Yang dijadikan sebagai titik awal dari perhitungan ini adalah struktur tunggal yang biasanya dijadikan sebagai struktur ekuilibrium yang didapatkan setelah meminimalkan energi potensial. Kecepatan pergerakan atom akan meningkat secara perlahan dari nol hingga mencapai nilai yang sesuai dengan suhu tertentu. Suhu ini disebut sebagai suhu ekuilibrasi yang dibutuhkan untuk memastikan sistem menuju ke suhu yang diinginkan (Levitt 1995)
Fungsi energi potensial yang mendasari simulasi dinamika molekul merupakan penjumlahan antara energi potensial yang terjadi akibat interaksi ikatan (bonded interaction) dengan energi potensial akibat interaksi non-ikatan (nonbonded interaction). Interaksi ikatan pada ikatan kovalen berupa stretching, bending, dan torsion dapat dijelaskan oleh hukum Hooke
Energi stretching dari semua ikatan kovalen dalam protein direpresentasikan oleh suku penjumlahan pertama, dimana Kb adalah konstanta regangan
(stretching), b adalah panjang ikatan, beq adalah nilai keseimbangan panjang
Dalam simulasi dinamika molekul, fungsi energi potensial diberikan oleh medan gaya, yaitu fungsi yang mendefinisikan gaya-gaya yang bekerja pada suatu atom individual pada keadaan energi rendah (kesetimbangan termal) (Ponder dan Case 2003). Ada dua kelompok interaksi yang dapat memberikan pengaruh terhadap energi potensial, yaitu interaksi internal dan eksternal. Interaksi internal didefinisikan sebagai interaksi kovalen antar atom yang disebut sebagai interaksi ikatan (bonded interaction), meliputi uluran ikatan (bonded streching) yaitu interaksi kovalen antara dua atom pada jarak kesetimbangan tertentu, sudut ikatan dan sudut dihedral. Pada simulasi dinamika molekul, ikatan kovalen didefinisikan sebagai pegas dengan pergerakan harmonis (Becker 2001). Sedangkan interaksi eksternal mengekspresikan interaksi non-ikatan. Interaksi non-ikatan mempresentasikan interaksi fleksibel diantara pasangan atom atau partikel. Dua jenis interaksi non-ikatan paling umum yang dapat mengakibatkan perubahan energi potensial adalah interaksi elektrostatik (Potential Coulomb) dan interaksi van der Waals (Potential Lennard Jones) (Becker et al. 2001).
3 METODOLOGI
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah data eksperimen hasil Xray Diffraction berupa data koordinat lipase G. Zalihae yang dapat diunduh pada Protein Data Bank (http://www.rscb.org/pdb/) (Berman et al. 2000) dengan kode PDB 2DSN.
Waktu dan Tempat Penelitian
Penelitian ini telah dilakukan pada bulan Desember 2015 sampai bulan Juli 2016 di Laboratorium Fisika Teori dan Komputasi, Departemen Fisika, Fakultas Matematika dan Ilmu Pengethaun Alam, Institut Pertanian Bogor, Bogor.
Tahapan Penelitian Preparasi Sistem Simulasi
Struktur kristal Lipase G. Zalihae (Kode PDB : 2DSN) (Matsumura et al. 2007) yang digunakan pada simulasi diperoleh dari bank data Protein Data Bank (PDB), (http://www.rscb.org/pdb/) (Berman et al. 2000). Penentuan struktur kristal 2DSN dilakukan dengan metode sinar X dengan jumlah residu penyusunnya 774 yang mana enzim lipase T1 memiliki 2 chain yaitu chain A dan chain B yang masing-masing chain memiliki residu penyusun 387 dan terdapat 1 frame sebagai model konformasinya.
Solvasi model molekul menggunakan TIP3P sebagai molekul pelarut (Jorgensen et al. 1983). Pelarut yang digunakan pada penelitian ini adalah ; etanol, methanol, dan air sebagai pembanding dengan menggunakan program packmol.Pada tahap preparasi molekul, program yang digunakan adalah VMD. Data 2DSN.pdb yang telah diunduh kemudian dihilangkan Na dan Cl. Kemudian dilakukan pemisahan chain A dan chain B. Pada penelitian ini menggunakan chain A yang berjumlah 387 residu asam amino.
Data 2DSN.pdb tersebut tidak memuat informasi spesifik bagaimana atom bisa saling berinteraksi, sehingga dibutuhkan data agar dapat digunakan untuk menerapkan medan gaya tertentu ke dalam sistem molekul. Medan gaya yang digunakan untuk mendefinisikan energi potensial struktur kristal adalah ff12sb. Tahap preparasi berikutnya adalah melarutkan molekul kedalam pelarut (air,metanol,etanol) dalam kotak berukuran 10 Å x 10 Å x 10 Å sebagai wadah pelarut molekul 2DSN. Jumlah molekul air yang ditambahkan pada kotak adalah 14658molekul air, 4542 molekul etanol, dan 6514 molekul metanol.
Kondisi Simulasi
Keseluruhan simulasi menggunakan parameter integrasi waktu (time step) setiap 2 femto sekon (fs). Simulasi dilakukan dengan menggunakan metode periodic boundary condition (PBC) untuk menghilangkan efek tegangan permukaan dan untuk mencapai kondisi dengan kerapatan dan tekanan yang lebih seragam (Allen 1989). Algoritma SHAKE dengan toleransi 10-5 diterapkan kedalam sistem untuk mengekang seluruh ikatan yang mengandung atom hidrogen (Ryckaert et al. 1977). Energi elektrostatik sistem dihitung secara menyeluruh menggunakan metode particle mesh Ewald (PME) (Darden et al. 1993 dan Essmann et al. 1995) sedangkan Interaksi van der Waals dihitung menggunakan potensial Lennard-Jones dengan cutoff masing-masing adalah 12 Å (Bui et al. 2009). Untuk mengontrol tahapan minimisasi, pemanasan dan production run dilakukan dengan menggunakan NVT ensemble (constant number, volume and temperature) pada temperatur yang diinginkan dimana sistem dikopling terhadap termostat velocity rescaling pada konstanta kopling 0,1 (Bussi et al. 2007). Untuk mengontrol tahapan ekuilibrasi dijaga pada NPT Ensemble (constant number, pressure and temperature) oleh termostat Nose-Hoover (Martyna et al. 1994 dan Feller et al. 1995).
Minimisasi
Molekul yang telah disolvasi kemudian diminimisasi untuk menghindari kontak van der Waals yang tidak sesuai (bad contact) dan untuk meminimalkan efek-efek sterik yang berenergi tinggi. Setiap simulasi dimulai dengan minimisasi selama 40 ps untuk membuat protein berada di energi terendahnya (keadaan stabil) dengan menggunakan program sander.
Pemanasan dan Ekuilibrasi
Adapun proses pemanasan molekul yang telah diminimisasi menggunakan perangkat lunak Ambertools12 dengan program pmemd yang dilakukan secara bertahap selama 40 ps dengan suhu 300 K. Selanjutnya dilakukan ekuilibrasi yang bertujuan untuk menahan protein stabil di dalam sistem simulasi. Ekuilibrasi dilakukan dengan protokol Langevin selama 50 ps.
Production Run
Setelah molekul enzim terekuilibrasi dengan baik maka tahap selanjutnya adalah tahap produksi (Production Run). Konstrain yang diterapkan pada proses ekuilibrasi kemudian dihilangkan pada proses production run sehingga molekul protein bebas bergerak. Production run dilakukan secara bertahap selama 10 ns dengan temperatur 300 K pada pelarut air, metanol, dan etanol.
Analisis Simulasi Dinamika Molekul
Root-mean-square deviation (RMSD) adalah akar kuadrat rata-rata penyimpangan koordinat atom dari posisi referensi. RMSD merupakan ukuran perbedaan struktur protein selama proses simulasi terhadap struktur awal protein. Hasil analisis RMSD tersebut akan di plot dalam bentuk grafik antara RMSD terhadap waktu simulasi (Becker et al. 2001 dan Coutsias et al. 2004).
Analisis jari-jari girasi (radius of gyration) juga dapat digunakan untuk menggambarkan kekompakan (compactness) serta kepadatan/ densitas molekul protein. Nilai jari-jari girasi dihitung menggunakan fungsi posisi atom terhadap pusat massa protein. Root-mean-square fluctuation (RMSF) adalah akar kuadrat rata-rata fluktuasi koordinat atom terhadap struktur referensinya. RMSF merupakan analisis fleksibilitas residu asam amino penyusun protein.
Solvent accessible surface area (SASA) merupakan luas permukaan biomolekul (protein misalnya) yang dapat di akses oleh pelarut (Lee 1971). Nilai SASA memberikan gambaran terhadap struktur tersier protein. Pada umumnya, protein mengemas struktur tersiernya sedemikian rupa sehingga gugus-gugus yang bersifat hidrofobik akan berada di bagian dalam sedangkan gugus-gugus asam amino yang bersifat hidrofil akan terdapat di bagian luar. Ketika proses simulasi termal berlangsung, kestabilan struktur tersier protein dan derajat keeksposuran dapat diamati dengan melakukan perhitungan SASA.
Struktur sekunder protein tersusun atas interaksi lokal inter-residu yang pada umumnya dimediasi oleh ikatan hidrogen. Struktur sekunder yang paling umum adalah α-helix dan β-sheet. Analisis struktur sekunder dilakukan untuk melihat kestabilan struktur sekunder selama proses simulasi berlangsung yang dikarakterisasi dengan perubahan komposisi α-helix, β-sheet, turn dan coil. Perhitungan struktur sekunder dilakukan dengan menggunakan plug-in yang terdapat pada perangkat lunak VMD yang didasarkan atas algoritma (Frishman 1995).
Analisis energi konformasi merupakan energi total yang diperoleh dari penjumlahan antara energi ikatan, energi sudut, energi dihedral, dan energi akibat interaksi van der Waals antara atom-atom pada protein. Energi konformasi yang tinggi menandakan suatu keadaan kurangnya stabilitas suatu protein.
4 HASIL DAN PEMBAHASAN
Penelitian pada enzim atau protein termostabil dapat memberikan pengetahuan baru tentang faktor-faktor yang menentukan stabilitas enzim, maupun aplikasi praktis pada bidang industri. Pemahaman tentang faktor penentu stabilitas suatu protein atau enzim dapat dijadikan sebagai acuan melakukan penelitian secara in-vitro.
Root Mean Square Deviation (RMSD)
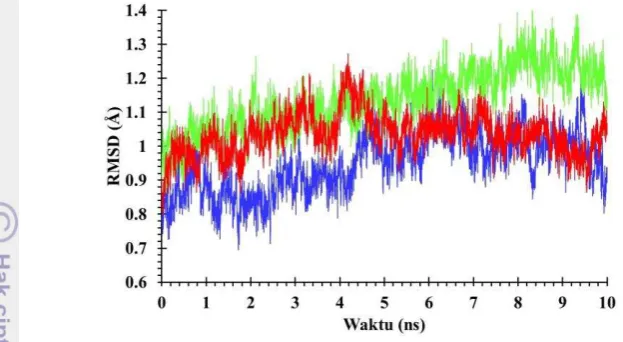
tinggi dibanding pada pelarut air dan etanol yaitu sebesar 1.1387Å dengan nilai RMSD awal sebesar 0.8956Å (Gambar 3).
Gambar 3 Root Mean Square Deviation (RMSD) selama simulasi pada suhu 300K selama 10 ns. Biru (air), merah (etanol), hijau (metanol).
Nilai RMSD pada pelarut air, metanol, dan etanol cukup konstan dan tidak mengalami peningkatan nilai yang signifikan. Adanya perbedaan pelarut pada sistem tersebut tidak mempengaruhi kestabilan protein. Pada pelarut metanol terjadi peningkatan nilai RMSD pada rentang waktu 8 hingga 9 ns yang mengindikasikan enzim cukup aktif bergerak dan tidak kaku dalam pelarut metanol. Enzim dikatakan rusak dari struktur awalnya jika nilai RMSD diatas 7 hingga 9 Å.
Pada gambar 3 dapat dilihat pada waktu 5 ns gerakan molekul didalam air relatif lebih rendah dari pada dalam pelarut etanol dan metanol. Namun setelah itu, pergerakan molekul dalam etanol relatif sama dengan pergerakan molekul didalam air. Pergerakan molekul hingga waktu 10 ns dalam pelarut metanol lebih tinggi dibandingkan pergerakan molekul dalam pelarut air dan etanol. Dinamika RMSD dalam pelarut metanol lebih tinggi daripada di air dan etanol, diduga karena polaritas relatif metanol (0.762) lebih dekat dengan polaritas air (1) dibandingkan dengan polaritas relatif etanol (0.654). Enzim Lipase T1 pada pelarut metanol diduga memiliki residu hidrofilik dan hidrofobik yang membuat enzim tetap bergerak. Gerakan tinggi pelarut etanol pada 5 ns diduga karena gerakan residu kutub pada permukaan ditekan saat bertemu pelarut non-polar.
Jari-jari Girasi (Rg)
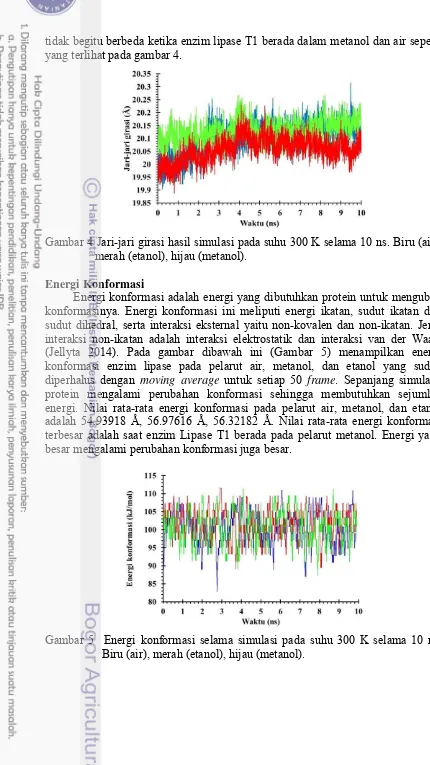
tidak begitu berbeda ketika enzim lipase T1 berada dalam metanol dan air seperti yang terlihat pada gambar 4.
Gambar 4 Jari-jari girasi hasil simulasi pada suhu 300 K selama 10 ns. Biru (air), merah (etanol), hijau (metanol).
Energi Konformasi
Energi konformasi adalah energi yang dibutuhkan protein untuk mengubah konformasinya. Energi konformasi ini meliputi energi ikatan, sudut ikatan dan sudut dihedral, serta interaksi eksternal yaitu non-kovalen dan non-ikatan. Jenis interaksi non-ikatan adalah interaksi elektrostatik dan interaksi van der Waals (Jellyta 2014). Pada gambar dibawah ini (Gambar 5) menampilkan energi konformasi enzim lipase pada pelarut air, metanol, dan etanol yang sudah diperhalus dengan moving average untuk setiap 50 frame. Sepanjang simulasi, protein mengalami perubahan konformasi sehingga membutuhkan sejumlah energi. Nilai rata-rata energi konformasi pada pelarut air, metanol, dan etanol adalah 54.93918 Å, 56.97616 Å, 56.32182 Å. Nilai rata-rata energi konformasi terbesar adalah saat enzim Lipase T1 berada pada pelarut metanol. Energi yang besar mengalami perubahan konformasi juga besar.
Solvent Accessible Surface Area (SASA)
Analisis SASA dapat digunakan untuk menghitung luas area permukaan protein yang dapat dimasuki oleh molekul pelarut. Dalam penelitian ini molekul pelarut yang digunakan adalah air, metanol, dan etanol. Semakin besar nilai SASA menunjukkan semakin besar molekul protein berekspansi. Peningkatan nilai SASA total ini karena sebagian struktur protein menjadi terbuka sehingga permukaan protein dapat dimasuki molekul pelarut. Grafik pada Gambar 6 diperhalus dengan moving average untuk setiap 50 frame.
Pada simulasi 4 ns, SASA total molekul lipase T1 dalam pelarut metanol lebih tinggi daripada dalam pelarut air dan etanol. Setelah waktu 4 ns hingga akhir simulasi, SASA total dalam pelarut metanol dan air relatif seimbang yang ditunjukkan pada gambar 6 (c). Hasil yang berbeda dari SASA total ditunjukkan pada gambar 6 (c) ketika molekul dalam pelarut etanol. Nilai SASA total pada pelarut etanol cenderung menurun dan lebih rendah dibanding dalam pelarut air dan metanol seperti yang terlihat pada gambar 6 (c). Nilai rata-rata SASA total tertinggi pada saat molekul dalam pelarut metanol sebesar 16600 Å2, Nilai SASA total dalam air sebesar 16343 Å2 , dan dalam pelarut etanol sebesar 16185 Å2. Nilai rata-rata SASA total tertinggi ketika dilarutkan dalam metanol disebabkan oleh nilai SASA non-polar paling tinggi ketika berada dalam pelarut metanol dibandingkan pelarut air dan etanol seperti yang terlihat pada gambar 6 (a). Nilai rata-rata SASA non-polar masing-masing adalah 5264 Å2, 5112 Å2 dan 4729 Å2 dalam metanol, etanol dan air. Perhitungan SASA polar untuk lipase T1 ketika dalam metanol diperoleh nilai rata-rata yaitu sebesar 11336 Å2, sementara nilai rata-rata SASA polar dalam air adalah 11614 Å2 dan dalam etanol adalah 11074 Å2 yang ditunjukkan pada gambar 6 (b). Hasil analisis SASA sesuai dengan data RMSD pada gambar 3, dimana dinamika enzim lipase T1 yang lebih tinggi dalam pelarut metanol karena adanya dinamika gerakan residu non-polar dan memiliki gerakan residu polar yang cukup stabil. Penurunan nilai RMSD enzim lipase T1 ketika dalam pelarut etanol ditunjukkan pada gambar 3, diduga bahwa pada SASA polar terjadi penyusutan nilai SASA dan pada SASA non polar tidak banyak berubah setelah 4 ns. Peningkatan yang tinggi pada analisis SASA polar dalam pelarut air serta nilai SASA yang tinggi pada pelarut non polar yaitu metanol menandakan bahwasannya sistem berjalan dengan baik.
(a)
(b)
(c)
Gambar 6 Nilai SASA (a) non-polar (b) polar (c) total selama simulasi pada suhu 300 K selama 10 ns. Biru (air), merah (etanol), hijau (metanol).
Root-mean-square fluctuation (RMSF)
Gambar 7 Nilai RMSF selama simulasi pada suhu 300 K selama 10 ns. Biru (air), merah (etanol), hijau (metanol).
Parameter RMSF dari gambar 7 terlihat bahwa simulasi pada pelarut air, metanol, dan etanol nilai RMSF tidak mengalami peningkatan yang signifikan. Pola yang diperlihatkan juga cenderung sama pada ketiga variasi pelarut. Ini menandakan bahwa protein masih cenderung stabil, walaupun ada beberapa puncak yang muncul menjelaskan bahwa residu fleksibel dalam pelarut, tetapi jumlahnya tidak terlalu banyak. Sedangkan pada pelarut air dan metanol nilai RMSF meningkat cukup signifikan dan pola nilainya juga sedikit berbeda dibandingkan pada pelarut etanol. Dari grafik dapat dilihat bahwa residu-residu yang semula rigid menjadi fleksibel, yang ditandai oleh banyaknya puncak yang muncul.
Pada grafik RMSF, puncak tertinggi dari residu asam amino menunjukkan fleksibilitas residu. Residu asam amino dalam pelarut air dan metanol menunjukkan fleksibilitas yang lebih tinggi daripada dalam pelarut etanol (Gambar 7). Residu Thr74, Thr93, Gly104, Ser201, Arg230, dan Ser333 adalah residu polar, yang terletak di permukaan dan memiliki fleksibilitas yang lebih tinggi saat berada dalam pelarut air dibanding dalam pelarut metanol dan etanol. Namun, ada dua residu hidrofobik Pro217 dan Val294 yang memiliki fleksibilitas tinggi dalam pelarut air dibanding dalam pelarut metanol dan etanol. Residu Pro217 adalah residu hidrofobik tapi posisinya berada diantara 3 residu polar, sehingga nilai RMSF menjadi lebih tinggi dalam pelarut air dibanding dalam pelarut metanol dan etanol. Residu hidrofobik Val294 terletak pada helix ke 9 yang berdekatan dengan pusat tutupnya aktif dan posisi Val294 didorong oleh residu Arg179 yang merupakan residu polar. Hasil RMSF menunjukkan bahwa fluktuasi asam amino dalam pelarut etanol umumnya lebih kaku daripada dalam pelarut air dan metanol serta lebih fleksibel pada pelarut metanol dan air (Gambar 7).
Interaksi Elektrostatik/ Jembatan Garam
(1978) yang menyatakan bahwa interaksi ini memiliki kontribusi signifikan untuk menstabilkan protein.
Pada hasil simulasi terdapat 19 pasangan jembatan garam pada temperatur 300 K selama 10 ns pada simulasi dengan masing-masing pelarut dan terdapat 14 pasangan jembatan garam yang muncul pada ketiga pelarut. Selanjutnya jembatan garam diplot antara jarak (Å) dengan waktu (ns). Pada simulasi yang telah dilakukan terdapat 8 pasangan jembatan garam yang statis yaitu Asp166-Lys329, Asp178-Lys229, Asp36-Arg21, Asp76-Arg89, Glu23-Lys207, Glu284-Arg271, Glu360-Arg271, Glu38-Arg21 (lampiran 6). Pasangan jembatan garam ini mengindikasikan bahwa enzim Lipase T1 stabil dalam semua pelarut ini menunjukkan saltbridge terikat kuat baik dalam air, etanol, dan metanol dapat dikatakan 8 pasang jembatan garam ini sebagai penstabil struktur. Pada simulasi terdapat 6 pasangan jembatan garam yang memiliki dinamika yaitu Asp205-Arg92, Asp209-Asp205-Arg92, Glu132-Lys84, Glu149-Arg134, Glu250-Arg330 (lampiran 6). Pada grafik dapat dilihat bahwasannya terjadi dinamika enzim Lipase T1 pada pelarut air,etanol, dan metanol ini berarti saltbridge bersifat fluktuatif sehingga membuat struktur enzim menjadi lebih dinamis.
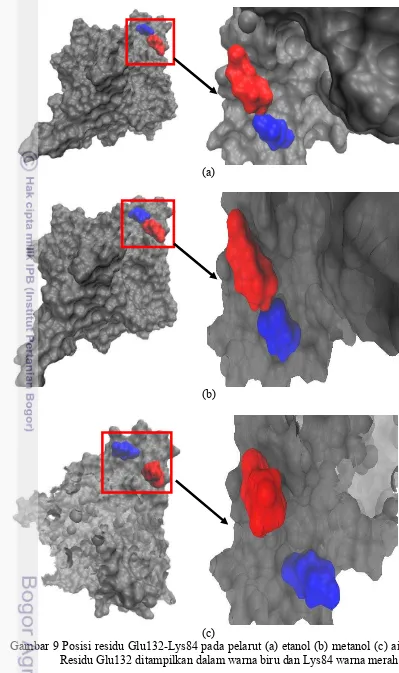
Pada simulasi yang telah dilakukan terdapat 3 pasangan saltbridge yang muncul pada pelarut air yaitu Asp43-Arg373, Glu381-Arg4, dan Glu100-Arg47, 2 saltbridge pada pelarut etanol yaitu Asp36-Lys28 dan Glu360-Arg34, serta 2 saltbridge yang muncul pada pelarut metanol yaitu Asp371-Arg34 dan Glu149-Lys138 (lampiran 7). Selain itu juga terdapat 4 pasangan saltbridge yang berperan dalam meningkatkan kekakuan struktur konformasi dari enzim Lipase T1 pada pelarut etanol dan metanol yaitu pasangan Glu132-Lys84, Glu149-Arg134, Asp205-Arg92, dan Glu250-Arg330 dapat dilihat pada gambar berikut.
(a)
(b)
(c)
Pasangan jembatan garam Glu132-Lys84 seperti yang terlihat pada gambar 8 pasangan jembatan garam terlihat stabil dalam pelarut etanol dan metanol. Pasangan jembatan garam ini menjauh saat waktu 2 ns hingga 4 ns dalam pelarut etanol tetapi muncul lagi hingga akhir simulasi. Ini menunjukkan bahwa jembatan garam ini kuat dan berperan penting terhadap stabilitas enzim lipase T1 dalam etanol dan metanol. Hal ini dapat dilihat gada gambar 9 posisi residu Glu132-Lys84 pada pelarut etanol dan metanol posisinya berdekatan daripada saat berada didalam air. Hal ini menjelaskan bahwasannya pasangan jembatan garam Glu132-Lys84 pada pelarut etanol dan metanol lebih terikat kuat daripada saat berada dalam air.
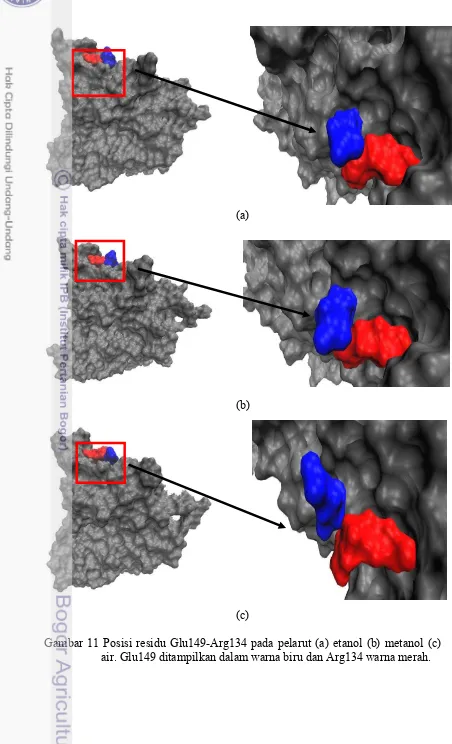
Glu149-Arg134 merupakan pasangan jembatan garam yang cukup memiliki dinamika sejak awal simulasi hingga akhir simulasi pada pelarut air yang ditunjukkan pada gambar 10. Jembatan garam ini juga memiliki jarak ikatan cukup lemah dalam etanol namun cukup kuat setelah 6 ns, seperti yang ditunjukkan pada gambar 10. Ketika molekul ini berada dalam pelarut metanol, pasangan jembatan garam ini memiliki energi sekitar -40 kJ / mol sejak awal simulasi sampai 8ns. Pasangan jembatan garam ini memiliki karakteristik yang sama ketika berada dalam etanol, seperti yang ditunjukkan pada gambar 10. Ini menunjukkan bahwa pasangan jembatan garam ini cukup kuat untuk membuat struktur molekul stabil dalam metanol dan bahkan lebih kaku dalam etanol.
Pada gambar 11 menunjukkan posisi residu Glu149-Lys134 pada pelarut etanol dan metanol posisinya berdekatan dibandingkan pada saat berada didalam air. Hal ini menjelaskan bahwasannya pasangan jembatan garam Glu149-Lys134 pada pelarut etanol dan metanol lebih terikat kuat daripada saat berada dalam air terlihat pada gambar 10, pada grafik dapat dilihat pada pelarut air residu memiliki banyak dinamika daripada saat berada pada pelarut metanol dan etanol.
Gambar 10 Grafik energi elektrostatik pasangan jembatan garam Glu149-Arg134. Biru (air), merah (etanol), hijau (metanol).
(a)
(b)
(c)
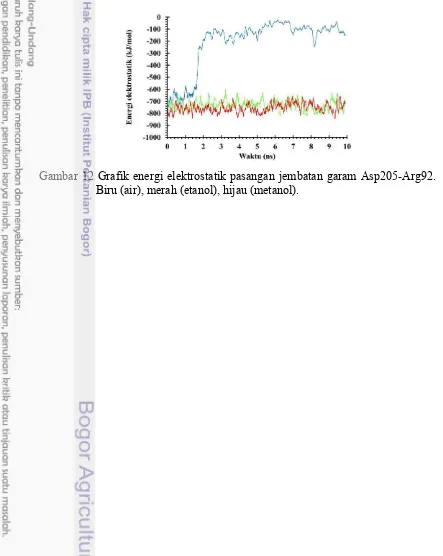
Pasangan jembatan garam Asp205-Arg92 memiliki energi yang kuat untuk mengikat dalam pelarut etanol dan metanol dan juga menunjukkan energi yang lemah setelah rusak pada waktu di 1,6 ns dalam pelarut air seperti yang terlihat pada gambar 12. Ini menunjukkan bahwa jembatan garam ini sangat kuat untuk membuat struktur lebih kaku dalam etanol dan metanol.
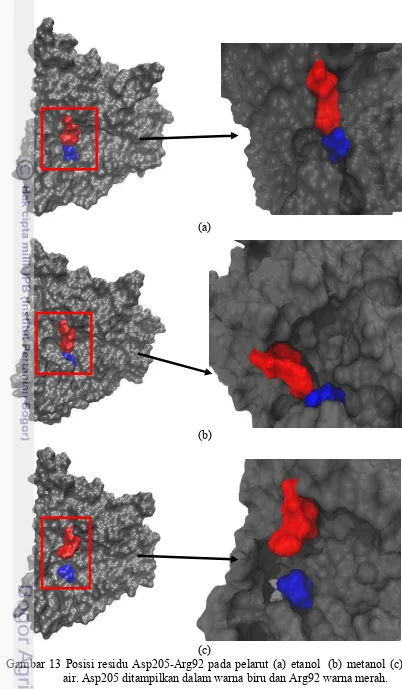
Pada gambar 13 menjelaskan bahwa pasangan residu Asp205-Arg92 saat berada dalam pelarut etanol dan metanol residu stabil dan terikat kuat. Hal ini dapat dilihat dari posisi residu yang berada berdekatan pada pelarut etanol dan metanol dibandingkan pada saat didalam air. Posisi residu Asp205-Arg92 saat didalam air setelah 2ns terjadi dinamika (gambar 12) serta letaknya yang berjauhan menandakan bahwasannya ikatan antara kedua residu cukup lemah (gambar 13) .
Gambar 12 Grafik energi elektrostatik pasangan jembatan garam Asp205-Arg92. Biru (air), merah (etanol), hijau (metanol).
(a)
(b)
(c)
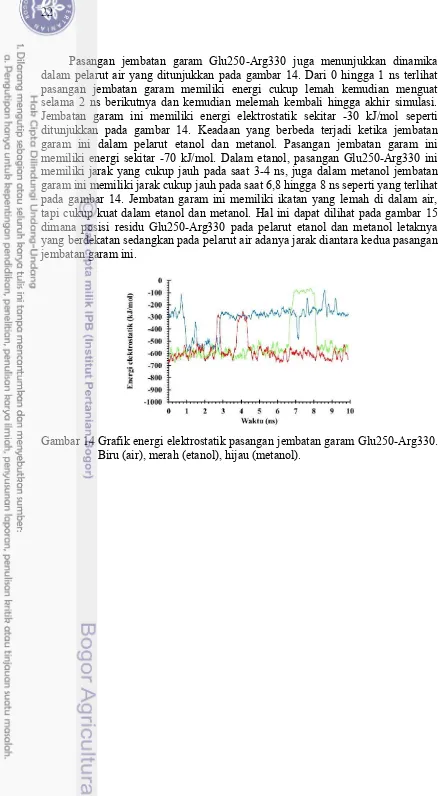
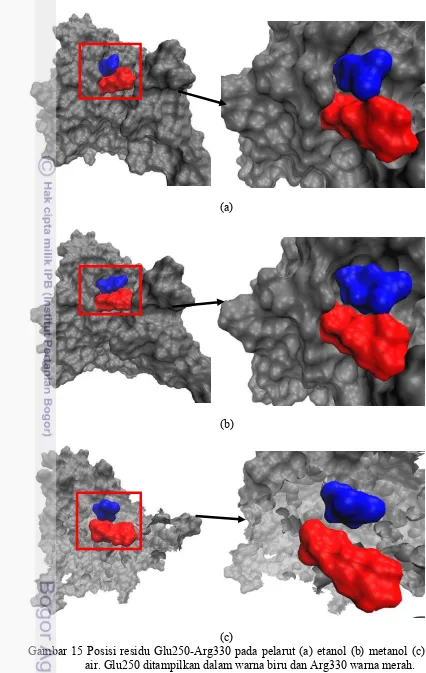
Pasangan jembatan garam Glu250-Arg330 juga menunjukkan dinamika dalam pelarut air yang ditunjukkan pada gambar 14. Dari 0 hingga 1 ns terlihat pasangan jembatan garam memiliki energi cukup lemah kemudian menguat selama 2 ns berikutnya dan kemudian melemah kembali hingga akhir simulasi. Jembatan garam ini memiliki energi elektrostatik sekitar -30 kJ/mol seperti ditunjukkan pada gambar 14. Keadaan yang berbeda terjadi ketika jembatan garam ini dalam pelarut etanol dan metanol. Pasangan jembatan garam ini memiliki energi sekitar -70 kJ/mol. Dalam etanol, pasangan Glu250-Arg330 ini memiliki jarak yang cukup jauh pada saat 3-4 ns, juga dalam metanol jembatan garam ini memiliki jarak cukup jauh pada saat 6,8 hingga 8 ns seperti yang terlihat pada gambar 14. Jembatan garam ini memiliki ikatan yang lemah di dalam air, tapi cukup kuat dalam etanol dan metanol. Hal ini dapat dilihat pada gambar 15 dimana posisi residu Glu250-Arg330 pada pelarut etanol dan metanol letaknya yang berdekatan sedangkan pada pelarut air adanya jarak diantara kedua pasangan jembatan garam ini.
Gambar 14 Grafik energi elektrostatik pasangan jembatan garam Glu250-Arg330. Biru (air), merah (etanol), hijau (metanol).
(a)
(b)
(c)
Analisis Struktur Sekunder
Waktu (ns) Waktu (ns) Waktu (ns) (a) (b) (c)
Gambar 16 Analisis struktur sekunder selama simulasi pada suhu 300 K selama 10 ns pada (a) air (b) metanol (c) etanol ; α-helix (Ungu), β-sheet (Kuning), Turn (Biru) dan Coil (Hitam) ; α-helix (Ungu), β-sheet (Kuning) 3-10helix (Biru), Turn (Hijau) dan Coil (Putih)
Analisis struktur sekunder menunjukkan bahwa selama simulasi, α-helix di lingkungan etanol hanya mengalami kerusakan relatif sangat kecil dibandingkan dengan pelarut lain seperti yang terlihat pada gambar. Struktur β-sheet relatif stabil dan konstan di semua pelarut yang ditunjukkan pada Gambar. Pada titik ini, masih terlalu cepat untuk mengatakan bahwa protein telah terdenaturasi saat dilakukan simulasi karena perlu waktu simulasi lagi (di kisaran milidetik), tetapi indikasi awal cenderung mendukung model zipper yang dimulai dengan kerusakan α-helix. Pada simulasi dengan menggunakan pelarut alkohol dapat dilihat struktur α-helix dan β-sheet enzim lipase T1 tidak mengalami kerusakan. Struktur sekunder dikatakan rusak jika struktur α-helix dan β-sheet berubah menjadi struktur turn dan coil.
Str
uktur
sek
5 SIMPULAN DAN SARAN
Simpulan
Simulasi dinamika molekul telah menunjukkan bahwa enzim lipase T1 mampu menjaga integritas struktur dalam berbagai pelarut misalkan pada 100% murni etanol, metanol, dan air. Hasil ini penting karena asumsi sebelumnya menyatakan bahwa enzim lipase T1 akan rusak dengan mudah di lingkungan alkohol murni. Dalam lingkungan alkohol, hasil penelitian ini menunjukkan bahwa enzim lebih dinamis dalam pelarut metanol dan lebih kaku dalam pelarut etanol. Ketika konformasi terlalu kaku, fungsi enzim akan berkurang, sehingga sangat penting untuk mempertahankan struktur enzim lipase T1 agar tetap memiliki dinamika. Hasil penelitian ini juga menunjukkan bahwa penggunaan pelarut metanol dalam reaksi biodiesel akan diharapkan baik daripada pelarut etanol.
Saran
DAFTAR PUSTAKA
Allen MP, Tildesley DJ. 1989. Computer Simulation of Liquids, Oxford University Press, USA.
Aunstrup KO, Andressen, Falch EA, Nielsen. 1979. Production of Microbial Enzymes. Microbial Technology. Vol. 1. Academic Press Inc. New York. Becker OM, Mackerell ADJr, Roux B, Watanabe M. 2001. Computational
Biochemistry and Biophysics. New York: Marcel Dekker.
Beckerman M. Molecular and Cellular Signaling. Tannessee (US): Springer Science. 2005.
Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, Shindyalov IN, Bourne PE. 2000. The Protein Data Bank. Nucleic Acids Res. 28: rescaling. J. Chem. Phys. 126: 014101.
Caflisch A, Paci E. 2005. Molecular Dynamics Simulations to Study Protein Folding and Unfolding. Protein Folding Handbook. Part I. Edited by J. Buchner and T. Kiefhaber. Copyright 8 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim. ISBN: 3-527-30784-2.
Corzo G, Revah S. 1999. Production and characteristics of the lipase from Yarrowia lipolytica 681. Bioresource Technology. 70: 173–180.
Coutsias EA, Seok C, dan Dill KA. 2004. Using quaternions to calculate RMSD, J.Comput. Chem. 25(15) : 1849-1857.
Darden T, York D, Pedersen L. 1993. Particle mesh Ewald: An N. log(N) method for Ewald sums in large systems. J. Chem. Phys. 98(12) : 10089- 10092. Dosanjh, N.S., dan Kaur, J. 2002. Immobilization, Stability and esterification
Studies of A Lipase From Bacillus sp. Journal Biotechnology and Applied Biochemistry. Vol. 36. Hlm 7-12. Punjab University. Chandigarh.
Essmann U, Perera L, Berkowitz ML, Darden T, Lee H, dan Pedersen, L.G. (1995) : A smooth particle mesh Ewald method, J. Chem. Phys., 103(19), 8577-8593.
Feller SE, Zhang Y, Pastor RW, Brooks BR. 1995. Constant pressure molecular dynamics simulation: The Langevin piston method. J. Chem. Phys. 103 (11): 4613-4621.
Frishman D, Argos P. 1995. Knowledge-based protein secondary structure assignment.Proteins: Structure, Function, and Genetics. 23: 566-579. Fodiloglu S, Erkmen O. 1999. Lipase production by Rhizopus oryzae growing
on different carbon and nitrogen sources. Journal of the Science of Food and Agriculture. 79 : 1936–1938.
Furuta, S., Hiromi, M., Kazushi, R. (2006), “Green Disel Fuel Production with Solid Amorphous-Zirconia Catalyst in Fixed Bed Reactor”, J Biomass And Bioenergy, Vol. 30, hal. 870-873.
Haryahto, B. 2002. Bahan Bakar Alternatif Biodiesel. Sumatera Utara: Jurusan Teknik Kimia Universitas Sumetera Utara.
Hati, Jellyta. Analisis kestabilan protein 1GB1 menggunakan simulasi dinamika molekul. [skripsi]. Bogor : Institut Pertanian Bogor. 2014.
Hasan, F., Shah, A. A., dan Hameed, A. (2006) : Industrial Aplication of Microbial Lipases, Microbial research Lab., Department of Biological, Quid-i-Azam University, Islamabad Pakistan.
Humphrey W, Dalke A, Schulten K. 1996. VMD: visual molecular dynamics. J. Mol. Graph, 14(1) : 27 – 38.
Jorgensen WL, Chandrasekhar J, Madura JD, Impey RW, Klein ML. 1983. Comparison of simple potential functions for simulating liquid water.J. Chem. Phys. 79: 926-935.
Knothe, G., Van Gerpen, J. H. and Krahl, J. (2005). The biodiesel handbook, AOCS Press, Champaign, Ill.
Kulkarni N, Gadre RV. 2002. Production and properties of an alkaline, thermophilic lipase from Pseudomonas fluorescens NS2W. Journal of Industrial Microbiology and Biotechnology. 28: 344–348.
Lee B, Richards FM. 1971. The interpretation of protein structures: estimation of static accessibility. J. Mol. Biol. 55(3) : 379-400.
Levitt M, Hirshberg M, Sharon R, Daggett V. Potential energy function and parameters for simulations of the molecular dynamics of proteins and nucleic acids in solution. Computer Physics Communication. 1995; 91:215-231.
Malau, Nya Daniaty. Kestabilan termal asam amino enzim lipase bacillus subtilis menggunakan simulasi dinamika molekul. [tesis]. Bogor : Institut Pertanian Bogor. 2015.
Martyna GJ, Tobias DJ, Klei ML. 1994. Constant pressure molecular dynamics algoritm. J. Chem. Phys. 101 (5): 4177-4189.
Matsumura H, et al. 2007. Novel cation-pi interaction revealed by crystal of thermoalkalophilic.Interscience.
Micaelo, Nuno. 2010. Analysis of molecular simulation experiment [bahan presentasi] Portugal Universidade do Minho.
Perutz M. 1978. Electrostatic effects in proteins. Science. 201(4362) : 1187-1191. Phillips, J. C., Braun, S., Wang, W., Gumbart, J., Tajkhorshid, E., Villa, E.,
Chipot, C., Skeel, R. D., Kale, L., Schulten, K., Scalable Molecular Dynamics with NAMD. Journal of Computational Chemistry. 2005; 26: 1781-1802.
Prasad. M.P . 2015. Studies on lipase enzyme production from bacillus subtilis under different culture conditions. EJBB. 2 : 15-19.
Ruhle, Victor. Berendsen and Nose-Hoover thermostats. 2007.
Ryckaert J, Ciccotti G, dan Berendsen H. 1977. Numerical Integration of the Cartesian Equations of Motion of a System with Constraints : Molecular Dynamics of n-Alkanes. J Comput. Phys.23: 327.
Supadi dan Nurmanaf, A. R. (2006) : Pemberdayaan Petani Kelapa Dalam Upaya Peningkatan Pendapatan Petani, Jurnal Penelitian dan Pengembangan Pertanian, 25(1), 31-36.
Sommer P, Bormann C, Gotz F. 1997. Genetic and biochemical characterization of a new extracellular lipase from Streptomyces cinnamomeus. Applied and Environmental Microbiology. 63: 3553–3560.
Van Gerpen, John (2004 - 07). Business Management for Biodiesel Producers, August 2002 - January 2004. National Renewable Energy Laboratory. Retrieved 2008-01-07.
Vieille C, Zeikus GJ. 2001. Hyperthermophilic enzymes: sources, uses, dan molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev.65(1) : 1-43.
Lampiran 1 Diagram alir penelitian
File PDB Protein Lipase T1 Mulai
Solvasi dan Netralisasi protein dengan box air
File protein T1 (tersolvasi)
Simulasi MD: 1.Minimisasi 2.Pemanasan (300K)
3.Ekuilibrasi 4.Dinamika molekul
(production run)
Simulasi dalam
air Simulasi dalam metanol Simulasi dalam etanol
Analisis Data
Penyusunan laporan
Lampiran 2 Pasangan jembatan garam yang muncul selama simulasi dalam pelarut air, metanol, dan etanol selama 10 ns dan suhu 300 K
No Air Metanol Etanol
1 Asp166-Lys329 Asp166-Lys329 Asp166-Lys329 2 Asp178-Lys229 Asp178-Lys229 Asp178-Lys229
3 Asp205-Arg92 Asp205-Arg92 Asp205-Arg92
4 Asp209-Arg92 Asp209-Arg92 Asp209-Arg92
5 Asp222-Lys185 Asp222-Lys185 Asp222-Lys185 6 Asp311-Arg274 Asp311-Arg274 Asp334-Arg120 7 Asp357-Arg271 Asp334-Arg120 Asp357-Arg271
8 Asp36-Arg21 Asp36-Arg21 Asp36-Arg21
9 Asp43-Arg373 Asp371-Arg34 Asp36-Lys28
10 Asp76-Arg89 Asp76-Arg89 Asp76-Arg89
11 Glu132-Lys84 Glu132-Lys84 Glu132-Lys84 12 Glu149-Arg134 Glu149-Arg134 Glu149-Arg134 13 Glu23-Lys207 Glu149-Lys138 Glu23-Lys207 14 Glu250-Arg330 Glu23-Lys207 Glu250-Arg330 15 Glu360-Arg271 Glu250-Arg330 Glu284-Arg271 16 Glu38-Arg21 Glu284-Arg271 Glu315-Arg274 17 Glu381-Arg4 Glu315-Arg274 Glu360-Arg271 18 Glu100-Arg47 Glu360-Arg271 Glu360-Arg34
Lampiran 3 Karakteristik Residu yang terdapat pada protein 2DSN
Struktur
Sekunder Indeks Residu Kode Residu Jenis Residu Asam Amino
Lampiran 4 Residu penyusun struktur sekunder protein 2DSN
No. Struktur Sekunder Index Residu Kode Residu
Lampiran 5 Nilai energi rata-rata enzim 2DSN pada ketiga pelarut
Air (Å) Metanol (Å) Etanol (Å)
Bond 28.63231 36.13036 32.79824
Angle 45.90275 48.93602 45.39155
Dihedral 32.66561 29.02724 22.54778
Impr 0 0 0
Elektrostatik 116.0409 71.80574 76.87536
Van der waals 34.89678 28.44984 31.03495
Konformasi 54.93918 56.97616 56.32182
Nonbonding 129.0462 63.08786 76.57268
Lampiran 6 Daftar 14 pasang jembatan garam yang mucul pada semua pelarut 8 pasangan jembatan garam yang statis dalam ketiga pelarut
Asp166-Lys329 Asp178-Lys229
Asp36-Arg21 Asp76-Arg89
Glu23-Lys207 Glu284-Arg271
6 Pasangan jembatan garam yang memiliki dinamika pada ketiga pelarut
Asp205-Arg92 Asp222-Lys185
Asp209-Arg92 Glu132-Lys84
Glu149-Arg133 Glu249-Arg329
Lampiran 7 Pasangan jembatan garam yang terdapat pada pelarut air
Asp43-Arg373 Glu100-Arg47
Glu381-Arg4
Pasangan jembatan garam yang terdapat pada pelarut etanol
Pasangan jembatan garam yang terdapat pada pelarut metanol
Glu149-Lys137 Asp370-Arg33
RIWAYAT HIDUP
Penulis dilahirkan di Bukittinggi pada tanggal 30 Juni 1990 dari Ayah Zulkifli.R dan Ibu Yetty Susi. Penulis adalah seorang anak tunggal. Pada tahun 2008 penulis berhasil menyelesaikan studi di SMA Negeri 4 Bukittinggi dan pada tahun 2009 penulis diterima pada Program Studi Pendidikan Fisika, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Negeri Padang. Pada tahun 2013 penulis lulus pada Program Studi Pendidikan Fisika.