SEJARAH PEMBENTUKAN BANGSA SAPI MADURA
BERDASARKAN GEN BRANCHED-CHAIN
α
-KETOACID
DEHYDROGENASE E1
α
(BCKDHA)
ASRI FEBRIANA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Sejarah Pembentukan Bangsa Sapi Madura Berdasarkan Gen Branched-Chain α-Ketoacid Dehydrogenase E1α (BCKDHA) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
ASRI FEBRIANA. Sejarah Pembentukan Bangsa Sapi Madura berdasarkan Gen Branched-Chain α-Ketoacid Dehydrogenase E1α (BCKDHA). Dibimbing oleh ACHMAD FARAJALLAH dan RR DYAH PERWITASARI.
Sapi madura merupakan salah satu bangsa sapi lokal Indonesia yang berasal dari persilangan sapi zebu (Bos indicus) dan banteng (Bos javanicus). Branched-chain α-ketoacid dehydrogenase (BCKDH) adalah salah satu dari kompleks enzim utama yang ada di membran dalam mitokondria yang memetabolisme branched-chain amino acid (BCAA), yaitu valin, leusin, dan isoleusin. Penelitian ini bertujuan mempelajari diferensiasi peruntukan sapi madura berdasarkan gen BCKDHA. Penelitian ini dilakukan terhadap tiga peruntukan sapi madura, yaitu sapi karapan, sonok, dan pedaging. Analisis urutan gen BCKDHA dilakukan dengan teknik sekuensing DNA.
Berdasarkan hasil pensejajaran urutan nukleotida menunjukkan panjang urutan sembilan ekson gen BCKDHA sapi madura yang diperoleh yaitu 1347 pb. Total 18 variasi ditemukan pada ruas ekson gen BCKDHA sapi madura. Selain variasi pada ruas ekson, terdapat 32 variasi yang ditemukan pada ruas intron berdasarkan tujuh pasang primer yang digunakan. Hasil pensejajaran urutan nukleotida gen BCKDHA pada sapi madura menunjukkan tidak ada variasi khusus yang ditemukan, baik pada ruas ekson maupun ruas intron, pada masing-masing peruntukan sapi madura. Diferensiasi yang ada tidak sampai mengarahkan pada pembentukan bangsa sapi baru. Hal ini menunjukkan tidak ada diferensiasi peruntukan sapi madura dari sisi tekanan seleksi terhadap gen BCKDHA.
SUMMARY
ASRI FEBRIANA. Natural History of Madura Cattle based on Branched-Chain α-Ketoacid Dehydrogenase E1α (BCKDHA). Supervised by ACHMAD FARAJALLAH and RR DYAH PERWITASARI.
Madura cattle is one of the Indonesian local cattle breeds derived from crossing between zebu cattle (Bos indicus) and banteng (Bos javanicus). Branched-chain α-ketoacid dehydrogenase (BCKDH) is one of the main enzyme complexes in the inner mitochondrial membrane that metabolizes branched-chain amino acid (BCAA), i.e., valine, leucine, and isoleucine. This study aimed to analyze the differentiation on madura cattle based on BCKDHA gene. This study was conducted on three madura cattle that used as karapan (bull race), sonok (art contest), and beef cattle by DNA sequencing of BCKDHA gene.
Based on the sequence analysis it was revealed that nine exons of BCKDHA gene in madura cattle consists of 1347 bp in length. A total of 18 variations found in exon region. In addition, there are 32 variations found in intron region based on seven pairs of primers used. The alignment results of BCKDHA gene sequence on madura cattle showed that there was no specific variation found, neither in the exon and intron, on karapan, sonok, and beef cattle. The differentiation was not lead to the formation of a new breed. This indicated the absence of differentiation madura cattle are used as karapan, sonok, and beef cattle of selection pressure of BCKDHA gene.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biosains Hewan
SEJARAH PEMBENTUKAN BANGSA SAPI MADURA
BERDASARKAN GEN BRANCHED-CHAIN
α
-KETOACID
DEHYDROGENASE E1
α
(BCKDHA)
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala rahmat, hidayah, dan karunia-Nya sehingga penulis dapat menyelesaikan karya ilmiah dengan judul Sejarah Pembentukan Bangsa Sapi Madura berdasarkan Gen Branched-Chain α-Ketoacid Dehydrogenase E1α (BCKDHA) pada Sapi Madura.
Terima kasih penulis ucapkan kepada Bapak Dr Ir Achmad Farajallah, MSi dan Ibu Dr Ir Rd. Roro Dyah Perwitasari, MSc selaku pembimbing, serta Bapak Prof Dr Ir Cece Sumantri, MSc selaku penguji yang telah memberikan saran dan masukan bagi perbaikan karya ilmiah ini. Di samping itu, ungkapan terima kasih juga penulis sampaikan kepada Bapak Dr Ir Jakaria, MSi dari Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, IPB atas sampel darah sapi bali dan PO yang diberikan, Kepala dan staf Rumah Pemotongan Hewan (RPH) terpadu Kota Bogor atas izin pengambilan sampel yang diberikan, Bapak Moh Romli dari Kecamatan Pakong, Kabupaten Pamekasan, yang telah mendampingi selama pengambilan sampel sapi sonok, Ibu Tini Wahyuni dan Bapak Adi Surachman, Wildan Najmul Muttaqin, MSi, beserta staf dan teman-teman di Bagian Fungsi Hayati dan Perilaku Hewan, Departemen Biologi, Fakultas Matematika dan IPA, IPB yang telah membantu selama pelaksanaan penelitian. Ungkapan terima kasih juga disampaikan kepada papa, mama, kakak dan adik tercinta serta seluruh keluarga, atas segala doa, dukungan, dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
2 TINJAUAN PUSTAKA 2
Sapi Lokal Indonesia 2
Sapi Madura 5
Sejarah Sapi Karapan (Balapan) 7
Sejarah Sapi Sonok (Pajangan) 8
Gen Branched Chain α-Ketoacid Dehydrogenase E1α (BCKDHA) 9
Desain Primer 12
Polymerase Chain Reaction (PCR) 15
Perunutan DNA (Sekuensing) 15
3 METODE 16
Waktu dan Tempat Penelitian 16
Sampel Darah dan Ekstraksi DNA 16
Amplifikasi DNA 17
DNA Sekuensing dan Analisis 19
4 HASIL DAN PEMBAHASAN 19
Struktur Daerah Penyandi Gen BCKDHA 19
Variasi Gen BCKDHA pada Sapi Madura 20
Hubungan antara Variasi Gen BCKDHA dengan Sejarah Sapi Madura 27
Indel Simultan pada Intron 7 Gen BCKDHA 29
Variasi pada Sapi Madura Pedaging 30
5 SIMPULAN DAN SARAN 30
Simpulan 30
Saran 30
DAFTAR PUSTAKA 30
LAMPIRAN 35
DAFTAR TABEL
1 Sampel darah sapi yang digunakan sebagai cetakan DNA berdasarkan
jenis sapi 17
2 Pooled samples darah sapi yang digunakan sebagai cetakan DNA
berdasarkan jenis sapi 17
3 Urutan primer untuk amplifikasi DNA 18
4 Panjang urutan ekson 1-9 gen BCKDHA sapi madura 20 5 Variasi yang ditemukan pada ruas ekson gen BCKDHA sapi madura 23 6 Variasi yang ditemukan pada ruas ekson gen BCKDHA pooled samples 23 7 Variasi yang ditemukan pada ruas intron gen BCKDHA 24 8 Variasi yang ditemukan pada ruas intron gen BCKDHA pooled samples 26 9 Variasi basa nukleotida (ruas ekson) yang hanya ditemukan pada sapi
13 Variasi nukleotida pada intron 7 gen BCKDHA 29
DAFTAR GAMBAR
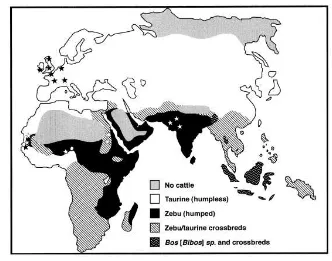
1 Tipe-tipe sapi domestikasi yang terdapat di Asia, Afrika, dan Eropa
(MacHugh 1996) 3
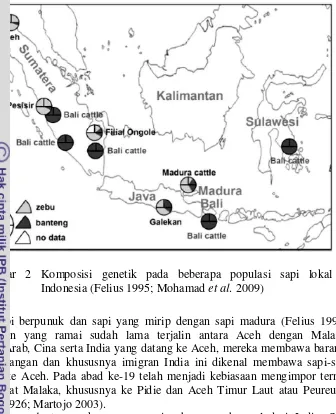
2 Komposisi genetik pada beberapa populasi sapi lokal Indonesia (Felius
1995; Mohamad et al. 2009) 4
3 Komponen genomik sapi indonesia berdasarkan penanda genetik mtDNA, kromosom Y, dan mikrosatelit (Mohamad et al. 2009) 6 4 Sapi madura yang terdapat di Kecamatan Pakong, Kabupaten
Pamekasan (foto oleh: Achmad Farajallah) 7
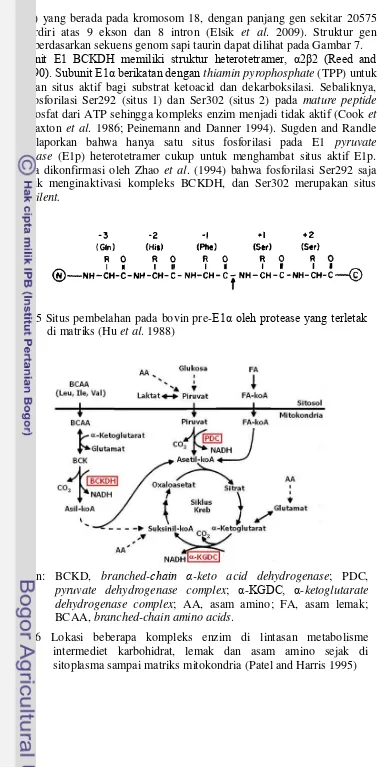
5 Situs pembelahan pada bovin pre-E1α oleh protease yang terletak di
matriks (Hu et al. 1988) 10
6 Lokasi beberapa kompleks enzim di lintasan metabolisme intermediet karbohidrat, lemak dan asam amino sejak di sitoplasma sampai matriks
mitokondria (Patel and Harris 1995) 10
7 Struktur gen BCKDHA berdasarkan urutan genom B. taurus (Elsik et
al. 2009; Zimin et al. 2009) 11
8 Posisi transit peptida pada gen BCKDHA 12
9 Tampilan menu awal program Primer3 13
10 Tampilan kondisi pemilihan primer secara umum 14 11 Skema posisi penempelan tujuh pasang primer pada ruas gen
BCKDHA 18
12 Urutan nukleotida dan asam amino subunit E1α gen BCKDHA 21 13 Pensejajaran urutan nukleotida dan asam amino pada daerah ujung 5’
14 Hasil kromatogram variasi alel 1 dan alel 2 pada ekson 1 gen
BCKDHA 22
DAFTAR LAMPIRAN
1
1
1
PENDAHULUAN
Latar Belakang
Sapi madura merupakan salah satu bangsa sapi lokal Indonesia yang berasal dari persilangan sapi zebu (Bos indicus) dan sapi bali atau banteng (Bos javanicus). Sapi madura yang ada saat ini merupakan hasil seleksi secara terus-menerus oleh petani di Pulau Madura sehingga menghasilkan suatu populasi bangsa yang seragam. Sapi madura memiliki nilai budaya penting bagi masyarakat madura. Berdasarkan kearifan lokal Madura, sapi madura dapat dikelompokkan menjadi tiga peruntukan, yaitu sapi karapan (balapan), sapi sonok (pajangan), dan sapi sayur (pedaging). Sapi karapan membutuhkan metabolisme energi yang tinggi untuk mendapatkan kekuatan fisik, kerja keras otot kerangka dan emosional (aggressive). Sebaliknya, sapi sonok membutuhkan metabolisme energi yang tinggi untuk menahan peregangan otot kerangka dan emosi terkendali (tamed). Sapi yang tidak memiliki sifat-sifat tersebut atau yang ‘pensiun’ dari sapi karapan dan sonok termasuk dalam sapi pedaging.
Lomba sapi karapan dan sapi sonok merupakan acara budaya penting di Pulau Madura (De Jonge 1990). Sapi karapan dan sapi sonok yang diikutsertakan dalam perlombaan merupakan sapi pilihan dengan performan dan kondisi yang sangat baik. Performan dan kondisi tersebut dapat dipengaruhi oleh faktor genetik dan lingkungan, termasuk lingkungan pakan dan kesehatan. Sapi madura termasuk sapi potong lokal yang memiliki keunggulan antara lain, yaitu kemampuan adaptasi yang baik terhadap lingkungan tropis, manajemen tradisional, kualitas pakan yang kurang baik, tahan terhadap serangan caplak, dan kualitas karkas yang tinggi.
Pemanfaatan sapi dalam tradisi budidaya masyarakat Madura mengarahkan seleksi sapi berdasarkan peruntukannya yang memiliki performan yang sesuai dengan selera masyarakat. Seleksi dengan cara tersebut kemungkinan mempengaruhi variasi gen yang terlibat dalam metabolisme energi. Variasi yang ditemukan kemungkinan terdiferensiasi mengikuti pola berdasarkan sapi karapan, sapi sonok, dan sapi pedaging. Selain sapi madura, pada penelitian ini juga digunakan sampel pembanding, yaitu sapi bali dan sapi Peranakan Ongole (PO) untuk melihat sejarah pembentukan bangsa sapi madura berdasarkan gen yang terlibat dalam metabolisme energi. Karakterisasi terhadap gen yang terlibat dalam metabolisme energi perlu dilakukan untuk meningkatkan kualitas sapi madura dalam skema pemuliaan ternak.
Branched-chain α-ketoacid dehydrogenase (BCKDH) adalah salah satu dari kompleks enzim utama yang ada di membran dalam mitokondria yang memetabolisme branched-chain amino acid (BCAA), yaitu valin, leusin, dan isoleusin. Kompleks BCKDH terdiri atas subunit α dan -ketoacid dehydrogenase (E1αdan E1β) dan dua subunit lainnya (Patel and Harris 1995). Subunit E1α pada sapi disandikan oleh gen branched-chain α-ketoacid dehydrogenase E1α (BCKDHA) yang berada pada kromosom 18, dengan panjang gen sekitar 20575 pb, dan terdiri atas 9 ekson dan 8 intron (Elsik et al. 2009).
2
proses metabolisme BCAA. Analisis variasi gen BCKDHA dilakukan dengan menggunakan teknik DNA sekuensing. Teknik ini digunakan untuk menentukan urutan nukleotida pada suatu molekul DNA. Tidak terdapat data mengenai urutan nukleotida gen BCKDHA pada sapi madura, sehingga urutan yang diperoleh diharapkan dapat digunakan sebagai referensi untuk penelitian lebih lanjut mengenai gen BCKDHA pada sapi lokal Indonesia lainnya.
Tujuan Penelitian
Penelitian ini bertujuan mempelajari diferensiasi peruntukan sapi madura berdasarkan gen BCKDHA.
2
TINJAUAN PUSTAKA
Sapi Lokal Indonesia
Sapi merupakan hewan ternak dengan keanekaragaman jenis yang tinggi dan ditemukan hampir di semua negara termasuk Indonesia. Sapi (Bos sp.) merupakan anggota famili Bovidae, subfamili Bovinae, genus Bos (Geraads 1992). Menurut Perkins (1969), domestikasi sapi pertama kali ditemukan di Turki dengan dua tipe sapi, yaitu tipe berpunuk (zebu) dan tidak berpunuk (taurin). Sebagian besar sapi di wilayah Asia merupakan keturunan tipe B. indicus atau dikenal sebagai sapi zebu, sedangkan sapi yang tersebar di wilayah Eropa termasuk dalam spesies B. taurus atau sapi taurin (Bradley and Cunningham 1999). Tipe sapi domestikasi yang terdapat di Indonesia, berbeda dari tipe B. indicus (sapi zebu) dan B. taurus (sapi taurin), dan berasal dari domestikasi yang terpisah (Gambar 1).
Sapi bali merupakan sapi asli Indonesia yang diduga berasal dari hasil domestikasi banteng liar, dan mempunyai ciri-ciri fisik yang hanya mengalami perubahan kecil dibandingkan dengan moyangnya (Handiwirawan dan Subandriyo 2004). Proses domestikasi sapi bali ini terjadi sebelum 3500 SM di Indonesia (Rollinson 1984). Mereka diklasifikasikan sebagai B. javanicus (Bradley and Cunningham 1999). Menurut Payne and Rollinson (1973), sapi bali memiliki karakteristik berukuran sedang, tidak berpunuk (humpless), memiliki warna putih pada bagian belakang paha, bagian perut, dan keempat kaki bawah sampai di atas kuku (white stocking).
3
diduga terjadi pada bangsa-bangsa sapi lokal Indonesia baik secara spontan maupun melalui pemuliaan (Felius 1995).
Gambar 2 menunjukkan bahwa berdasarkan penanda molekuler genetik DNA mitokondria, kromosom Y (analisis paternal), dan mikrosatelit (analisis autosomal) menunjukkan sapi bali memiliki komposisi genetik yang sama dengan banteng. Berdasarkan penanda genetik molekuler DNA mitokondria (analisis maternal) terdapat komposisi genetik sapi zebu pada sapi aceh dan pesisir, sedangkan pada sapi PO dan madura terdapat komposisi genetik sapi zebu dan sapi bali atau banteng, hal ini menunjukkan bahwa sapi PO dan madura berasal dari dua moyang betina yang berbeda. Pada sapi aceh, pesisir, dan madura komposisi genetik berdasarkan penanda molekuler kromosom Y (analisis paternal) menunjukkan ketiga bangsa tersebut berasal dari pejantan zebu. Berdasarkan penanda genetik mikrosatelit (analisis autosomal) sapi aceh, pesisir, PO, dan madura memiliki komposisi genetik sapi zebu dan sapi bali atau banteng (Felius 1995; Mohamad et al. 2009). Hal ini menunjukkan bahwa komposisi spesies bangsa sapi zebu Indonesia unik dan bervariasi dari sebagian besar zebu hingga sepenuhnya banteng. Sejarah dan pembiakan sapi Indonesia telah menghasilkan sumberdaya genetik yang unik yang mengombinasikan karakter toleran terhadap iklim tropis dari zebu dan iklim yang kering dari banteng pada kondisi peternakan di Indonesia.
Persilangan sapi-sapi di Indonesia dengan sapi zebu berhubungan dengan masuknya agama Hindu ke Indonesia. Tidak terdapat catatan yang pasti kapan masuknya sapi zebu ke Indonesia. Sapi zebu kemungkinan dibawa masuk ke Indonesia oleh imigran Singalese sekitar 1500 tahun yang lalu. Selain itu, terdapat patung sapi berpunuk pada candi-candi Hindu di Jawa bukti adanya sapi zebu pada abad ke-10. Foto-foto sapi asli jawa dari awal abad ke-20 juga menunjukkan
4
gambar sapi berpunuk dan sapi yang mirip dengan sapi madura (Felius 1995). Perdagangan yang ramai sudah lama terjalin antara Aceh dengan Malaka. Pedagang Arab, Cina serta India yang datang ke Aceh, mereka membawa barang-barang dagangan dan khususnya imigran India ini dikenal membawa sapi-sapi dari India ke Aceh. Pada abad ke-19 telah menjadi kebiasaan mengimpor ternak melalui Selat Malaka, khususnya ke Pidie dan Aceh Timur Laut atau Peureulak (Merkens 1926; Martojo 2003).
Sapi ongole termasuk rumpun sapi zebu yang berasal dari India. Sapi ongole mulai disilangkan dengan bangsa sapi lokal Indonesia pada abad ke-19, impor sapi ongole zebu terus digunakan untuk pembiakan di Pulau Jawa dan pulau-pulau lainnya di Indonesia, kecuali Pulau Madura dan Bali (Rollinson 1984; Felius 1995; Mohammad et al. 2009). Sapi ongole berperan penting dalam upaya pengembangan peternakan sapi di Indonesia. Impor sapi ongole dari India ke Pulau Sumba dalam jumlah yang banyak berlangsung sampai tahun 1920, sehingga pada tahun 1923 terdapat sekitar 1500 sapi ongole di Pulau Sumba yang kemudian dikenal dengan sapi sumba ongole (SO). Selanjutnya sapi SO disebarkan ke wilayah lain di Indonesia. Penyebaran tersebut menyebabkan terbentuknya suatu bangsa sapi yaitu peranakan ongole (PO) yang merupakan hasil persilangan antara sapi SO dengan sapi asli jawa yang saat ini telah punah. Selain sapi ongole, sapi hissar dari India juga diimpor dan digunakan untuk pembiakan di Sumatra dan Sulawesi Utara (Martojo 2003).
5 Sapi Madura
Sapi madura merupakan salah satu sapi potong lokal (indigenous) yang berkembang di Indonesia dan hasil persilangan antara banteng atau sapi bali (B. javanicus) dengan sapi zebu (B. indicus) sekitar 1500 tahun yang lalu (Payne and Rollinson 1973; Bradley and Cunningham 1999). Selain sapi zebu, sapi taurin (B. taurus) yang pernah dimasukkan ke Pulau Madura sebagai pejantan antara lain yaitu, Danish Red dan persilangan antara Shorthorn dengan Brahman atau dikenal sebagai Santa Gertrudis, yang semuanya memiliki warna merah coklat yang mirip dengan warna sapi madura, sapi Limousin juga diperkenalkan sebagai pejantan unggul dalam upaya meningkatkan produktivitas sapi madura.
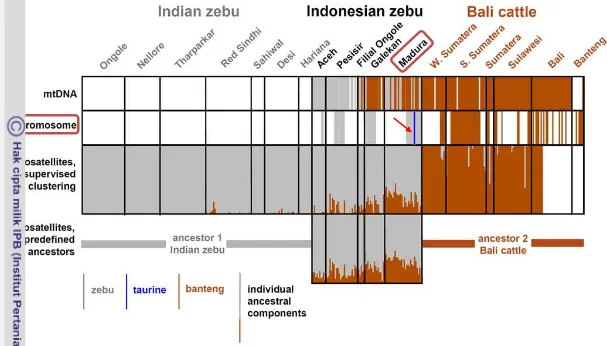
Pada Gambar 3, tanda panah pada baris kedua menunjukkan bahwa berdasarkan penanda genetik molekuler kromosom Y (analisis paternal) terdapat komponen genomik sapi taurin yaitu sapi Limousin pada sapi madura. Komunitas sapi madura yang berkembang saat ini merupakan hasil isolasi dan seleksi alamiah yang ketat, sehingga menghasilkan sapi yang relatif memberikan keseragaman genotip yang mantap (Soehadji 1993; Wijono dan Setiadi 2004; Mohamad et al. 2009).
Sapi madura memiliki karakteristik tubuh berwarna kuning kecoklatan pada sapi betina, sedangkan sapi jantan berwarna merah bata atau merah coklat bercampur putih dengan batas yang tidak jelas pada bagian pantat (Gambar 4). Warna putih seperti pada kaki (white stocking) juga sering ditemukan di daerah abdomen dan bagian paha dalam. White stocking yang ditemukan pada kaki sapi Madura tidak sejelas yang ditemukan pada sapi bali. Ukuran tubuhnya kecil sampai sedang, berpunuk kecil, bertulang dan berotot bagus, terutama sapi jantan (karapan) dan mempunyai kaki yang cukup kuat untuk bertahan terhadap kerja yang berat. Pada bagian wajah di sekitar mata berwarna hitam, pinggir telinga berwarna hitam, serta tanduk yang berwarna hitam, berukuran kecil dan pendek mengarah ke bagian luar. Bagian bawah (tarsal/metatarsal) berwarna putih, ekor berwarna coklat sampai hitam. Sapi jantan memiliki gelambir lebih besar dibandingkan dengan sapi betina (Setiadi dan Diwyanto 1997).
Berdasarkan karakter morfologi, sapi madura memiliki karakteristik yang hampir sama dengan sapi bali, kecuali ukuran tubuh dan tanduknya yang lebih kecil (Setiadi 2010). Sapi madura jantan memiliki tanduk yang pendek dan beragam lebih kurang 15-20 cm, sedangkan pada betina, tanduk lebih kecil dan pendek lebih kurang 10 cm. Panjang badan hampir sama dengan sapi bali, namun berpunuk kecil sedangkan sapi bali tidak berpunuk. Tinggi badan sekitar 118 cm dengan berat badan rata-rata 350 kg, sedangkan pada sapi bali yang berumur sekitar 2 tahun memiliki berat 400 kg dan umur 4 tahun dengan berat badan sekitar 600-800 kg (Martojo 2003). Menurut Soehadji (1993), sapi madura sebagai sapi potong tipe kecil memiliki variasi berat badan sekitar 300 kg. Pada sapi madura yang menang kontes, berat badan mampu mencapai lebih dari 500 kg yang diperoleh dari hasil pemeliharaan yang baik dengan pemenuhan kebutuhan pakan yang baik.
6
6 Gambar 3 Komponen genomik sapi indonesia berdasarkan penanda genetik mtDNA, kromosom Y, dan mikrosatelit (Mohamad et al.
2009)
7
7 infestasi serangan caplak, dan kualitas karkas yang tinggi. Namun, pengembangbiakkan yang dilakukan di luar Pulau Madura, tidak menunjukkan adanya perkembangan yang signifikan walaupun kondisi lingkungan di wilayah tersebut hampir sama. Sejak awal abad ke-20 bangsa sapi lain dilarang masuk ke Pulau Madura untuk menjaga kemurniannya (Martojo 2003). Hal ini dapat menyebabkan terjadinya penurunan produktivitas yang dapat diakibatkan oleh seleksi negatif, yaitu pemotongan sapi produktif dan faktor inbreeding akibat Pulau Madura merupakan wilayah tertutup bagi sapi potong lain (Wijono dan Setiadi 2004).
Pengaruh nilai sosiobudaya masyarakat Madura terhadap ternak sapi madura memiliki nilai tersendiri terutama terhadap tradisi sapi betina pajangan yang dikenal sebagai sapi sonok dan lomba balapn sapi jantan yang dikenal sebagai karapan. Aspek sosiobudaya masyarakat Madura yang mengembangkan kesenangan memelihara sapi yang baik untuk dilombakan, merupakan faktor pendukung didalam upaya pelestarian dan perbaikan mutu ternak. Namun, upaya pelestarian sapi madura yang telah lama dilaksanakan sejak pemerintahan Hindia Belanda ini juga memiliki kelemahan, yaitu kegiatan lomba karapan sapi dan sapi sonok menyebabkan peternak cenderung lebih mengutamakan sapinya daripada kebutuhan keluarga (Setiadi dan Diwyanto 1997). Selain itu, peranan pemeliharaan sapi madura seperti pemeliharaan sapi potong lainnya yaitu sebagai sumber penghasil daging, tenaga kerja, dan kebutuhan ekonomi (Wijono dan Setiadi 2004).
Sejarah Sapi Karapan (Balapan)
Karapan memiliki sejarah yang panjang. Noer dan Maduratna (1975) menyatakan bahwa di Pulau Sapudi yang terletak di Kabupaten Sumenep, selama abad ke-12 sampai 13, Pangeran Katandur ingin membuat tanah gersang dan tidak subur di pulau tersebut menjadi lebih subur sehingga dapat dibuat untuk bercocok tanam. Pangeran Katandur merupakan seorang tokoh penyebar agama islam, beliau memiliki nama asli Habib Ahmad Baidawi putra dari Panembahan Pakaos atau Habib Sholeh putra dari Sunan Kudus. Katandur atau Sinandur sendiri memiliki arti yang menanam. Pangeran memperkenalkan cara membajak sawah
8
dengan menggunakan tenaga sepasang sapi jantan. Pangeran menciptakan permainan lomba balapan sapi, atau dikenal dengan karapan, dimana lomba mengadu sepasang sapi jantan melawan satu sama lain di lapangan untuk mendorong masyarakat menjaga sapi mereka. Selain itu, tujuan awal lomba karapan sapi hanya untuk menghibur masyarakat yang bergembira atas keberhasilan panen mereka. Seiring waktu, karapan menjadi permainan favorit bagi masyarakat madura, yang diselenggarakan setelah musim panen. Peralatan yang digunakan dalam lomba mirip dengan peralatan yang digunakan dalam membajak atau disebut dengan kaleles. Kaleles merupakan tempat atau alat yang digunakan oleh seorang joki untuk menaiki sepasang sapi dengan panjang lintasan sekitar 100-140 m (Widi et al. 2013).
Sejarah Sapi Sonok (Pajangan)
Sejarah sapi sonok berasal dari kegiatan yang pada umumnya dilakukan oleh masyarakat madura yaitu menambatkan sapi mereka di antara dua pilar, dengan nama lokal tacek, di beranda depan rumah. Kaki depan sapi berdiri di atas platform, sepotong kayu dengan tinggi sekitar 15 cm, sementara mereka ditambatkan. Kegiatan ini memberikan petani kesempatan untuk menunjukkan kebanggaan mereka dalam merawat sapi mereka. Pada tahun 1963, seorang Kepala Desa sangat tertarik pada kegiatan para peternak ini. Lalu, Kepala Desa menunjukkan sapi dengan bentuk tubuh yang baik pada saat ditambatkan di depan rumah petani, kepada pejabat tinggi yang sedang mengunjungi desanya. Ide ini kemudian menjadi kegiatan rutin di daerah tersebut (Widi et al. 2013).
Pada tahun 1927, Sommerfeld (1927) menjelaskan tentang pajengan dimana petani membawa sapi mereka, terutama sapi betina, untuk ditambatkan bersama di lapangan. Kebiasaan ini juga dilatarbelakangi oleh kebiasaan petani memajang sapi mereka dalam posisi tegap, memiliki keindahan tubuh, dan warna kulit yang bagus. Pada tahun 1967, seorang pejabat pemerintahan lokal menyelenggarakan kegiatan pajengan dua kali seminggu selama musim panen tembakau. Hal ini menjadi ajang bagi para petani untuk bertemu, jual-beli sapi, berbagi informasi dengan penyuluh, serta mampu meningkatkan prestise dan status sosial pemiliknya.
9 Gen Branched Chain α-Ketoacid Dehydrogenase E1α (BCKDHA)
Proses metabolisme melibatkan banyak gen dan keseluruhan gen yang terlibat dalam jalannya proses metabolisme tersebut dinamakan genom. Metabolisme merupakan rangkaian reaksi kimia yang mengubah satu molekul menjadi molekul lainnya. Setiap tahap reaksi kimia dalam sel memerlukan satu jenis enzim sebagai katalisator dan setiap jenis enzim dibentuk di bawah kendali satu jenis gen. Gen adalah bagian dari genom yang berperan dalam proses ekspresi gen, yaitu sebagai ruas model atau ruas penyandi dalam proses transkripsi. Dari keseluruhan genom organisme, tidak semuanya menjadi ruas penyandi, hanya bagian yang diapit oleh promotor dan terminator (Jusuf 2002).
Daerah penyandi merupakan bagian dari gen yang ditranskripsi dan diterjemahkan menjadi protein, sehingga perubahan pada daerah tersebut dapat
mempengaruhi fungsi gen dengan mempengaruhi pola splicing mRNA atau
dengan mengubah fungsi protein (Cartegni et al. 2002). Struktur suatu gen penyandi terdiri atas 5’ UTR (Untranslated Region), ekson, intron, dan 3’ UTR pada sel eukariotik. Wilayah yang tak tertranslasikan pada ujung 5’ dan 3’ mRNA atau disebut sebagai 5’ UTR dan 3’ UTR merupakan bagian yang tidak akan ditranslasikan menjadi protein, namun memiliki fungsi-fungsi lain, misalnya pengikatan ribosom. Ekson adalah sekuens nukleotida yang diekspresikan atau ditranslasikan menjadi sekuens asam amino. Sedangkan intron merupakan sekuens nukleotida DNA yang tidak ikut ditranslasikan. Sebagian intron mengandung sekuens yang meregulasi aktivitas gen. Salah satu konsekuensi dari keberadaan intron-intron dalam gen adalah satu gen dapat mengodekan lebih dari satu jenis polipeptida (Allison 2007).
10
(BCKDHA) yang berada pada kromosom 18, dengan panjang gen sekitar 20575 pb, dan terdiri atas 9 ekson dan 8 intron (Elsik et al. 2009). Struktur gen BCKDHA berdasarkan sekuens genom sapi taurin dapat dilihat pada Gambar 7.
Subunit E1 BCKDH memiliki struktur heterotetramer, α2β2 (Reed and Hackert 1990). Subunit E1α berikatan dengan thiamin pyrophosphate (TPP) untuk menghasilkan situs aktif bagi substrat ketoacid dan dekarboksilasi. Sebaliknya, dua situs fosforilasi Ser292 (situs 1) dan Ser302 (situs 2) pada mature peptide menerima fosfat dari ATP sehingga kompleks enzim menjadi tidak aktif (Cook et al. 1984; Paxton et al. 1986; Peinemann and Danner 1994). Sugden and Randle (1978) melaporkan bahwa hanya satu situs fosforilasi pada E1 pyruvate dehydrogenase (E1p) heterotetramer cukup untuk menghambat situs aktif E1p. Hal ini juga dikonfirmasi oleh Zhao et al. (1994) bahwa fosforilasi Ser292 saja cukup untuk menginaktivasi kompleks BCKDH, dan Ser302 merupakan situs fosforilasi silent.
Keterangan: BCKD, branched-chain α-keto acid dehydrogenase; PDC, pyruvate dehydrogenase complex; α-KGDC, α-ketoglutarate dehydrogenase complex; AA, asam amino; FA, asam lemak; BCAA, branched-chain amino acids.
Gambar 6 Lokasi beberapa kompleks enzim di lintasan metabolisme intermediet karbohidrat, lemak dan asam amino sejak di sitoplasma sampai matriks mitokondria (Patel and Harris 1995) Gambar 5 Situs pembelahan pada bovin pre-E1α oleh protease yang terletak
11
Keragaman urutan nukleotida suatu gen menentukan efisiensi dari enzim yang disandikannya. Pada gen BCKDHA, terdapat ruas transit peptide yang disandikan oleh bagian ujung 5’ ekson 1 sampai ujung 5’ ekson 2 dan mature peptide yang menyandikan polipeptida untuk membentuk struktur kuartener dengan subunit lainnya (bagian ujung 3’ ekson 2 sampai ekson 9) (Gambar 8). Transit peptide terdiri atas 165 pb yang menyandikan 55 aa, sedangkan mature peptide terdiri atas 1200 pb yang menyandikan 400 aa, sehingga Open Reading Frame menyandikan 455 aa (Hu et al. 1988; Zimin et al. 2009). Mutasi nukleotida pada bagian ruas ekson kemungkinan akan mempengaruhi kualitas transit peptide, situs pelekatan dengan subunit lainnya dan situs katalitik.
Variasi dapat ditemukan baik di daerah penyandi maupun bukan penyandi. Variasi tersebut dapat mempengaruhi fungsi atau ekspresi gen sehingga dapat mengakibatkan kondisi yang tidak diinginkan, seperti penyakit. Ibeagha-Awemu et al. (2008) menyebutkan bahwa dalam sebagian besar laporan mutasi patologis yang ditemukan terjadi pada daerah fungsional protein dan bersifat lestari (conserved). Gen BCKDH telah digunakan sebagai marka untuk mendeteksi suatu penyakit, yaitu Maple Syrup Urine Disease (MSUD). Penyakit ini merupakan penyakit metabolisme resesif autosomal yang mempengaruhi metabolisme BCAA (valin, leusin, dan isoleusin). Pada keadaan normal, kelebihan protein akan memicu BCAA untuk menghasilkan energi. BCAA akan didegradasi oleh kompleks enzim branched chain α-ketoacid dehydrogenase (BCKD). Pada MSUD, jika satu atau beberapa gen yang menyandikan komponen-komponen kompleks enzim ini mengalami mutasi maka BCAA tidak bisa masuk kedalam rangkaian metabolisme mitokondria sehingga terjadi akumulasi dalam darah dan urin. Akumulasi ini menyebabkan urin penderita memiliki bau seperti sirup maple (Fisher et al. 1991). Furney et al. (2006) menemukan bahwa gen penyakit yang dipengaruhi oleh mutasi dominan lebih conserved dibandingkan gen yang
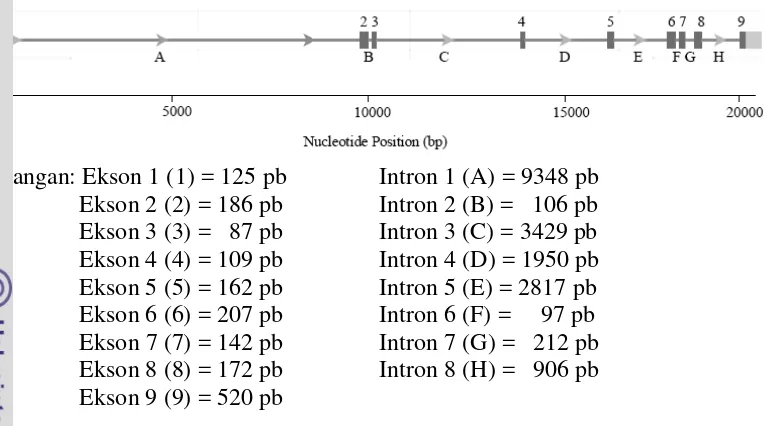
Keterangan: Ekson 1 (1) = 125 pb Intron 1 (A) = 9348 pb
12
dipengaruhi oleh mutasi resesif. Hal ini disebabkan mutasi resesif tetap tersembunyi dari seleksi karena heterozigot.
MSUD telah diidentifikasi baik pada manusia maupun sapi (Fisher et al. 1991; Chuang et al. 1993; Dennis and Healy 1999). Mutasi gen BCKDHA ekson 2 pada sapi Polled Shorthorn terjadi karena substitusi 248C/T (CAG menjadi TAG) pada kodon 6 dan menyebabkan stop kodon yang menghasilkan terminasi prematur pada saat translasi (Zhang et al. 1990), sedangkan pada sapi Polled Hereford mutasi disebabkan oleh 1380C/T (Pro372Leu) pada gen yang sama (Dennis and Healy 1999). Sari (2013) menemukan variasi 36G>T pada ekson 4 gen BCKDHA pada sapi PO yang mengubah asam glutamat (GAA) menjadi stop kodon (TAA).
Desain Primer
Primer merupakan salah satu bahan yang digunakan dalam proses perbanyakan utas DNA dengan menggunakan metode Polymerase Chain Reaction (PCR). Primer PCR adalah DNA utas tunggal (single-stranded DNA) berukuran pendek yang spesifik, dikenal sebagai oligodeoxyribonucleotides atau oligomer. Desain primer dilakukan untuk membuat untaian basa nukleotida yang akan menempel dan mengapit daerah untaian DNA tertentu (DNA target) dari total genom yang ada pada kromosom. Pengapitan daerah untaian DNA diperlukan untuk melakukan perbanyakan (amplifikasi) hanya pada daerah untaian DNA tertentu sehingga lebih mudah dalam melakukan analisis terhadap daerah untai DNA tersebut. Keberhasilan melakukan amplifikasi daerah untai DNA sangat dipengaruhi oleh kecocokan primer terhadap kondisi untai DNA target (Muladno 2002).
Desain primer untuk mempelajari ruas-ruas ekson gen BCKDHA dapat dilakukan dengan menggunakan Primer3 (http://bioinfo.ut.ee/primer3/) secara online berdasarkan hasil pensejajaran gen BCKDHA pada B. taurus (NW_001493616), Capra hircus (NW_005100869), dan Ovis aries (NW_004080177). Gambar 9 menunjukkan tampilan situs Primer3. Desain primer dilakukan dengan mempertimbangkan kondisi optimum PCR sesuai dengan kondisi target DNA. Selain itu, desain primer juga mengikuti konsensus intron secara umum, yaitu intron mRNA mengandung tiga unsur konsensus yaitu GU pada ujung 5’ dan AG pada ujung 3’. Seluruh intron mengandung GT pada ujung
13
5’ dan AT pada ujung 3’, sehingga disebut aturan GT-AG atau GU-AG pada RNA (Breathnach et al. 1978).
Desain primer yang baik sangat penting bagi keberhasilan reaksi PCR. Berikut beberapa hal penting yang perlu dipertimbangkan dalam mendesain primer untuk memperoleh amplifikasi spesifik dengan hasil tinggi antara lain, yaitu panjang primer, primer melting temperature (Tm), primer annealing temperature (Ta), jumlah basa G dan C (GC content), dan jumlah basa G dan C yang terdapat pada 5 basa terakhir (3’) (GC clamp). Selain itu, struktur sekunder primer juga mempengaruhi kualitas dan kuantitas produk PCR, karena akan mengurangi kemampuan primer menempel pada template. Struktur sekunder primer dapat disebabkan oleh interaksi intra dan inter-molekuler primer. Berikut ini beberapa jenis struktur sekunder primer antara lain, yaitu hairpins, self-dimer, dan cross dimer. Hairpins terbentuk akibat interaksi intramolekuler primer sehingga membentuk semacam lipatan (loop). Self-dimer terbentuk oleh interaksi intermolekuler antara dua molekul primer yang sama, dimana primer homolog terhadap dirinya sendiri. Penambahan primer dalam jumlah besar dibanding jumlah DNA template dapat menyebabkan primer lebih suka membentuk dimer intermolekuler dibanding berhibridisasi dengan DNA template, sehingga produk (yield) akan berkurang. Primer cross dimer terbentuk karena interaksi intermolekuler antara primer sense dan antisense, dimana keduanya memiliki homologi (Rustam 2010).
Panjang optimal primer PCR umumnya adalah 18-23 nukleotida. Namun, menurut Viljoen et al. (2005) panjang primer yang digunakan dalam PCR sebaiknya memiliki ukuran 18-28 nukleotida agar dapat mengamplifikasi target DNA dengan spesifisitas yang bagus. Semakin pendek ukuran primer akan menyebabkan terjadinya mispriming (penempelan primer di tempat lain yang tidak dikehendaki) tinggi sehingga menyebabkan spesifitas dari primer tersebut berkurang dan berakibat pada efektifitas dan efisiensi proses PCR. Sedangkan
14
panjang primer lebih dari 28 pb tidak menyebabkan bertambahnya spesifisitas dan akan berpengaruh pada Tm primer (Handoyo dan Rudiretna 2001).
Suhu leleh atau melting temperature (Tm) merupakan temperatur dimana setengah dari dupleks DNA akan terpisah menjadi utas tunggal dan Tm juga mengindikasikan stabilitas dupleks. Primer dengan Tm berkisar antara 52-58oC sangat ideal, sedangkan Tm di atas 65oC akan mengurangi efektifitas penempelan primer sehingga proses amplifikasi DNA kurang berjalan baik. Nilai Tm sangat ditentukan oleh jumlah basa GC (GC content) (Rustam 2010).
Primer annealing temperature (Ta) merupakan suhu dimana primer dapat berikatan dengan template DNA dengan stabil (DNA hybrid stability). Suhu leleh (Tm) primer antara forward dan reverse primer tidak boleh terlalu jauh karena digunakan sebagai acuan untuk penentuan atau optimasi Ta. Jika suhu penempelan primer (Ta) terlalu tinggi akan menyulitkan terjadinya ikatan primer dengan template DNA sehingga akan menghasilkan produk PCR yang rendah (kurang efisien). Namun jika Ta terlalu rendah akan menyebabkan terjadinya penempelan primer pada template DNA yang tidak spesifik yang disebabkan oleh tingginya kemungkinan kesalahan penempelan primer pada cetakan DNA (Newton and Graham 1997).
Jumlah basa G dan C (GC content) yang ideal di dalam primer sekitar 40-60%. Pada untai DNA pasangan basa G-C melibatkan tiga ikatan hidrogen, sedangkan pasangan basa A-T dua ikatan hydrogen, sehingga keseimbangan persentase GC memungkinkan ikatan yang terbentuk lebih spesifik dan stabil. Spesifitas dan stabilitas ini juga dipengaruhi oleh Tm primer. Perbedaan Tm sepasang primer sebaiknya tidak lebih dari 5oC. Jumlah basa G dan C yang terdapat pada 5 basa terakhir (3’) disebut dengan GC clamp. GC clamp yang baik sekitar 3 basa G/C dan tidak melebihi 5 basa G/C. Keberadaan G/C di ujung 3’ primer sangat membantu terjadinya stabilitas ikatan antara primer dengan template DNA yang diperlukan untuk inisiasi polymerase DNA pada proses PCR (Rustam 2010).
Program Primer3 menyediakan tampilan pilihan yang bisa diatur oleh peneliti sesuai dengan kriteria yang diinginkan (Gambar 10). Namun, pilihan alternatif tersebut dapat diabaikan (tidak dilakukan perubahan, sesuai dengan pengaturan yang sudah tersedia pada tampilan program Primer3). Berikut tahapan desain primer menggunakan Primer3. Sekuen basa nukleotida (DNA target)
15 dimasukkan ke dalam kotak menu dalam program Primer3 lalu klik Pick Primers untuk mendapatkan primer yang akan mengapit DNA target yang akan dianalisa oleh peneliti. Jika program Primer3 tidak dapat memberikan keluaran (output) primer maka modifikasi dapat dilakukan dengan mengganti pengaturan atau menurunkan interval jumlah sekuen pada kotak menu product size sehingga program Primer3 dapat mencari daerah penempelan primer yang baru.
Polymerase Chain Reaction (PCR)
PCR adalah suatu reaksi in vitro untuk menggandakan jumlah molekul DNA pada bagian tertentu dengan cara mensintesis molekul DNA baru yang berkomplemen dengan molekul DNA target dengan bantuan enzim dan fragmen oligonukleotida (primer) dalam suatu thermocycler. Komponen penting yang dibutuhkan dalam reaksi PCR adalah DNA template yaitu molekul DNA yang digunakan sebagai cetakan untuk pembentukan molekul DNA baru; sepasang primer oligonukleotida (forward dan reverse) yang akan membatasi fragmen DNA target yang akan diamplifikasi; deoxyribonucleoside triphosphat (dNTP) yang mengandung basa nukleotida (A, C, G, T,); Buffer dan MgCl2 berfungsi untuk menjaga pH medium dan sebagai kofaktor untuk menstimulasi aktivitas DNA Polimerase; dan enzim DNA Polimerase berfungsi sebagai katalisator dalam reaksi polimerisasi DNA (Viljoen et al. 2005).
Satu siklus dalam reaksi PCR terdiri atas tiga tahap dasar yang terjadi pada suhu yang berbeda. Tahap pertama adalah denaturasi yaitu pemisahan molekul
utas ganda DNA dengan proses pemanasan pada suhu tinggi (94-95oC) sehingga
menjadi untai tunggal dan primer dapat menempel pada utas tunggal DNA cetakan. Setelah tahap pemanasan, tahap selanjutnya adalah annealing yaitu
penurunan suhu (sekitar 50-60oC) sehingga primer yang urutan nukleotidanya
berkomplemen dengan salah satu untai tunggal akan menempel pada posisi komplemennya. Setelah kedua primer tersebut menempel pada posisinya masing-masing maka pada suhu 72oC terjadi proses elongasi (pemanjangan) dimana enzim DNA Polimerase, buffer PCR, dNTP, dan MgCl2 memulai proses replikasi atau mensintesis molekul DNA baru sehingga satu molekul DNA ganda akan berlipat jumlahnya menjadi dua molekul DNA. Ketiga tahap dasar tersebut (denaturasi, annealing, dan elongasi) diulang kembali hingga 25-30 siklus (Muladno 2002; Viljoen et al. 2005).
Perunutan sekuens DNA (DNA Sequencing)
16
menghasilkan tangga pita (ladder) berlabel dengan tiap pita mewakili tersekuensnya satu basa (Sanger et al. 1977; Maxam and Gilbert 1977).
Metode Sanger lebih banyak digunakan karena lebih aman, mudah, praktis dan efisien. Larutan utama yang digunakan di dalam reaksi metode Sanger adalah dNTPs (deoxynucleotides triphosphates) untuk mensintesis molekul DNA baru dan ddNTPs (dideoxynucleotides triphosphates) yang akan menghentikan pemanjangan molekul DNA pada basa tertentu. Hasil akhir dari reaksi tersebut adalah sejumlah potongan DNA yang panjangnya bervariasi tetapi semuanya berakhir dengan nukleotida A (jika dNTP dicampur dengan ddATP), nukleotida C (jika dNTP dicampur dengan ddCTP), nukleotida G (jika dNTP dicampur dengan ddGTP), dan nukleotida T (jika dNTP dicampur dengan ddTTP). Untuk mendeskripsikan hasil elektroforesis dari metode ini adalah dengan menggunakan label yang berbeda (deoxynucleotides yang mengandung radioaktif atau label fluorescent pada primer, dNTP atau ddNTP) atau dengan pendekatan staining (silver staining) (Nicholas 1993; Albert et al. 2008).
Teknologi DNA Sequencing yang umum digunakan saat ini adalah Dye-terminator sequencing, dimana output alat sequencer-nya adalah peak-peak (puncak) yang terdiri atas 4 warna yang mewakili masing-masing nukleotida yaitu hijau untuk Adenin (A), merah untuk Timin (T), biru untuk Sitosin (C) dan hitam untuk Guanin (G). Peak inilah yang dinamakan electropherogram yang sudah diinterpretasikan secara otomatis oleh program komputer DNA Sequencer menjadi urutan-urutan basa nukleotida (A-C-G-T). Namun program komputer juga dapat melakukan kesalahan interpretasi pada electropherogram yang bermasalah sehingga urutan basa nukleotida yang dihasilkannya pun bisa salah. Hal ini dapat mempengaruhi hasil penelitian dan juga analisa lanjutan yang akan dilakukan terhadap hasil sekuensing tersebut.
3
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2014 sampai dengan Februari 2015 di Bagian Fungsi Hayati dan Perilaku Hewan, Departemen Biologi, Fakultas Matematika dan IPA, Institut Pertanian Bogor.
Sampel Darah dan Ekstraksi DNA
17 Jumlah sampel darah yang digunakan yaitu sebanyak 25 sampel sapi madura, 18 sampel sapi bali, dan 16 sampel sapi PO (Tabel 1 dan 2). Sampel darah diawetkan dalam alkohol absolut yang mengandung EDTA 100 mM. Ekstraksi DNA dilakukan dengan mengikuti protokol Genomic DNA Mini Kit for Fresh Blood (Geneaid). Variasi ruas-ruas ekson gen BCKDHA diperkirakan akan sangat rendah. Oleh karena itu, amplifikasi PCR pertama kali dilakukan dengan menggunakan pooled samples, yaitu sampel DNA sapi madura, bali, dan PO dijadikan menjadi satu tabung sampel dengan konsentrasi yang berimbang berdasarkan peruntukan sapinya (Tabel 2). Teknik DNA pooling merupakan gabungan dari beberapa sampel dijadikan satu dalam satu tabung sehingga dapat mendeteksi varian pada suatu lokus secara cepat (fast screening).
Amplifikasi DNA
Primer untuk mengamplifikasi DNA didesain menggunakan Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/primer3/) secara online berdasarkan hasil pensejajaran gen BCKDHA pada Bos taurus (NW_001493616), Capra hircus (NW_005100869), dan Ovis aries (NW_004080177). Desain primer dilakukan mulai dari ujung 5’ (forward) dan 3’ (reverse) setiap ekson dengan mengikuti konsensus 5’ GT-AG 3’ (Breathnach et al. 1978). Opsi-opsi yang digunakan dalam mendesain primer adalah yang optimal untuk ruas-ruas DNA berukuran pendek dengan suhu Tm sekitar 60 oC. Pada Tabel 3 terdapat tujuh pasang primer
18
yang didesain untuk mengamplifikasi sembilan ekson gen BCKDHA dan sebagian ruas pengapitnya (intron). Gambar 11 menunjukkan skema posisi penempelan masing-masing primer pada ruas ekson gen BCKDHA.
Amplifikasi DNA dilakukan dalam volume total 25 μL berisi 50-100 ng DNA template, primer forward dan reverse masing-masing 1 μM, dan GoTaq® Green Master Mix konsentrasi akhir 1X. Amplifikasi dilakukan dengan menggunakan mesin ESCO Swift Maxi Thermal Cycler dengan kondisi: predenaturasi 95 °C selama 2 menit, dilanjutkan dengan 30 siklus yang terdiri atas denaturasi 95 °C selama 45 detik, penempelan primer 60 °C selama 1 menit, pemanjangan DNA 72 °C selama 1 menit, dan diakhiri dengan pemanjangan DNA 72 °C selama 5 menit. Amplikon yang menunjukkan pita tunggal di atas gel poliakrilamida setelah elektroforesis gel poliakrilamida (PAGE) 6% dan pewarnaan perak (Byun et al. 2009), dijadikan cetakan dalam reaksi perunutan DNA (DNA sekuensing).
Gambar 11 Skema posisi penempelan tujuh pasang primer pada ruas gen BCKDHA Tabel 3 Urutan primer untuk amplifikasi DNA
Ekson Primer Urutan (5' → 3') Ukuran Produk
1 AF501F TGGCATTTGAGTCAGGCTTA 363 pb
AF502R ATTCCCTTGCATGTCCTGTC
2 & 3 AF503F GGCTGCATCACCAGCAAY 666 pb AF504R GATCCGGTTCAACCTTCTGC
4 AF505F CCCAGCCTCASTGAACCC 229 pb
AF506R GAGCCTGACCTGTCTCCAC
5 AF507F GRGGTAGCCTTCAGGGGTTG 381 pb
AF508R CCAYGAGGGTCYCAGAAAGA
6 & 7 AF509F CCAGAGGCCYGGGTYGG 640 pb AF510R GAATGGAAYGGAGGCCAGAG
8 AF511F CGTTGGYCACTGTCYCCACA 409 pb
AF512R CCCRGCCTCTCCMTTCTT
9 AF513F AGTGCCTGGTTCCACTAAGG 864 pb
19 DNA Sekuensing dan Analisis
Perunutan DNA menggunakan metode sequencing big dye terminator dilakukan oleh lembaga komersial jasa sekuensing First Base Laboratories, Malaysia. Perunutan DNA dilakukan dari dua arah, yaitu forward dan reverse. Urutan nukleotida yang diperoleh diedit dengan menggunakan program BioEdit versi 7.0.8.0 (Hall 1999) dan disejajarkan dengan urutan nukleotida gen BCKDHA pada B. taurus (NW_001493616). Proses pensejajaran dilakukan dengan menggunakan program ClustalW yang kemudian diedit ulang secara manual dan dibantu dengan program ClustalW yang ada dalam program MEGA6 (Tamura et al. 2013).
4
HASIL DAN PEMBAHASAN
Tujuh pasang primer yang digunakan berhasil mengamplifikasi urutan target, yaitu sembilan ekson gen BCKDHA dan daerah pengapitnya (intron) dengan teknik PCR. Hasil pensejajaran urutan nukleotida sapi madura pada penelitian ini dibandingkan dengan urutan gen BCKDHA B. taurus (NW_001493616) yang diperoleh dari GenBank sebagai referensinya.
Struktur Daerah Penyandi Gen BCKDHA
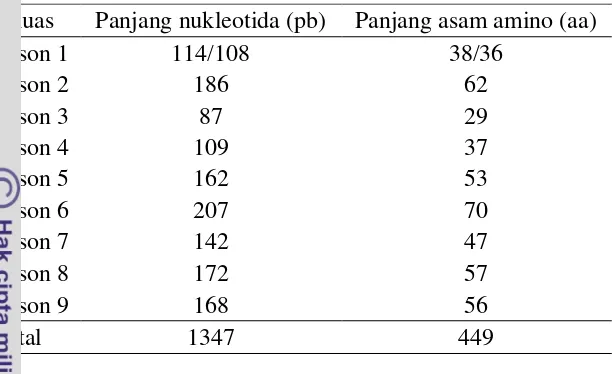
Hasil analisis urutan nukleotida ruas-ruas ekson gen BCKDHA yang diperoleh dengan teknik sekuensing disajikan pada Gambar 12. Analisis urutan nukleotida menunjukkan panjang urutan sembilan ekson gen BCKDHA yang diperoleh yaitu 1347 pb. Panjang ukuran masing-masing ekson terdapat pada Tabel 4. Ukuran masing-masing ekson yang dianalisis pada sapi madura memiliki panjang urutan yang hampir sama dengan urutan referensinya (Gambar 7). Akan tetapi, ruas ekson 1 yang ditemukan pada sapi madura dan sapi taurin (NW_001493616) memiliki urutan nukleotida yang berbeda dengan sapi taurin (J03759 dan NM_174506). Perbedaan ini disebabkan oleh perbedaan urutan nukleotida pada bagian ujung 5’, penentuan posisi kodon pertama dan delesi 6 pb pada ekson 1 (Gambar 13). Delesi tersebut mempengaruhi ukuran panjang ekson 1, sehingga menghasilkan urutan yang lebih pendek dibandingkan dengan urutan sapi taurin yaitu 108 pb dan 36 aa (Tabel 4).
20
Hal ini sesuai dengan studi yang dilakukan oleh Hu et al. (1988) yang menyatakan bahwa daerah situs fosforilasi protein E1α pada mamalia conserved.
Variasi Gen BCKDHA pada Sapi Madura
Variasi ruas-ruas ekson gen BCKDHA diperkirakan akan sangat rendah seperti pada gen-gen vital metabolisme lainnya, karena perubahan pada daerah tersebut dapat mempengaruhi fungsi suatu gen atau protein (Ibeagha-Awemu et al. 2008). Hasil pensejajaran urutan nukleotida menunjukkan bahwa daerah ekson lebih conserved dibandingkan dengan intron. Tidak ditemukan variasi pada daerah ekson 2, 4, dan 7 (conserved). Total 18 variasi ditemukan pada ruas ekson gen BCKDHA sapi madura (Tabel 5). Variasi juga ditemukan pada ruas ekson gen BCKDHA pooled samples (Tabel 6). Variasi yang ditemukan pada umumnya dalam bentuk delesi dan substitusi basa nukleotida, serta puncak kromatogram yang saling tumpang-tindih. Enam dari total variasi yang ditemukan merupakan delesi dan sisanya merupakan substitusi basa nukleotida. Ekson 1 memiliki variasi yang paling tinggi dibandingkan dengan ekson lainnya. Terdapat 12 variasi yang ditemukan pada ekson 1 yang terdiri atas enam delesi dan enam substitusi basa nukleotida. Variasi yang terdeteksi pada gen BCKDHA menyebabkan terjadinya dua tipe mutasi, yaitu mutasi sense dan missense. Mutasi sense pada penelitian ini umumnya ditemukan pada basa ke-tiga kodon, sehingga tidak menyebabkan terjadinya perubahan asam amino, sedangkan mutasi missense disebabkan oleh variasi yang terjadi pada basa pertama atau ke-dua dari setiap kodon, sehingga menyebabkan terjadinya perubahan asam amino (Lampiran 1).
Variasi yang ditemukan pada ruas intron lebih tinggi dibandingkan dengan ruas ekson. Total 32 variasi ditemukan pada delapan ruas intron gen BCKDHA berdasarkan tujuh pasang primer yang digunakan (Tabel 7 dan 8). Bentuk variasi yang ditemukan pada ruas intron pada umumnya sama dengan ruas ekson. Lima dari total variasi yang ditemukan merupakan delesi lima basa nukleotida pada intron 6 dan variasi lainnya merupakan substitusi basa nukleotida. Akan tetapi, terdapat satu variasi yang tidak ditemukan pada ruas ekson, yaitu insersi/delesi (indel) yang ditemukan secara bersamaan (simultan) pada daerah intron 7. Selain itu, intron 7 memiliki variasi yang paling tinggi dibandingkan dengan intron
Tabel 4 Panjang urutan ekson 1-9 gen BCKDHA sapi madura Ruas Panjang nukleotida (pb) Panjang asam amino (aa)
22
Gambar 14 Hasil kromatogram variasi alel 1 dan alel 2 pada ekson 1 gen BCKDHA
Gambar 13 Pensejajaran urutan nukleotida dan asam amino pada daerah ujung 5’ ekson 1
290 * 300 *
GluAlaMETThrTyrArgIleGlyHisHisSerThrSerAspAspSerSerAlaTyrArgSerValAspG
GAGGCCATGACCTACAGGATCGGGCACCACAGCACCAGTGACGACAGCTCGGCGTACCGCTCAGTGGACG 1120
310 320
luValAsnTyrTrpAspLysGlnAspHisProIleSerArgLeuArgHisHisLeuGlnSerArgGlyTr
AGGTCAACTACTGGGACAAGCAGGACCACCCCATCTCCCGGCTGCGGCATCACCTGCAGAGCCGCGGCTG 1190
330 340 350
pTrpAspAspGluGlnGluLysAlaTrpArgLysGlnSerArgLysLysValMETGluAlaPheGluGln
GTGGGACGACGAGCAGGAGAAGGCCTGGAGGAAGCAGTCCCGCAAGAAGGTAATGGAGGCCTTTGAGCAG 1260
360 370
GluGlnGluLysAlaTrpArgLysGlnSerArgLysLysValMETGluAlaPheGluGlnProAlaGlnL
GCTGAGCGGAAGCTGAAGCCCAACCCCAGCTTGATCTTCTCGGACGTGTATCAGGAGATGCCTGCCCAGC 1330
380 390
euArgLysGlnGlnGluSerLeuAlaArgHisLeuGlnThrTyrGlyGluHisTyrProLeuAspHisPh
TCCGCAAGCAGCAGGAGTCTCTGGCACGTCACCTCCAGACCTATGGTGAACACTACCCGCTGGACCACTT 1400
400 eGluLys***
CGAGAAGTGAGGCCCCATAGCCCTTGGGGTAGTGCCTCAACTACCCCAAGAGGCAGCCCCAGTGTGAGGG 1470
GCAGATGGGACTGGCAGGGTGCCAGCTTCCTCAGACAGCTGCCTCCAACACACTCAGGAGCCAGGCAGTA 1540
CTTGAGGCGGCTCCTGTTCCGCTAGGCTGTTACACTGGTCAGCCCCTTCTCCACCCTGAGTTACACGGTC 1610
TTCTCCCAGGGGCTGGGCGAGGGCACCTCTGGGACTAGACACCCCTGAGGGCTGGGGCTAGACGTGGTGG 1680
GTCAGTCTGTGGAACTTGACTCAGAATTAGAGGTCAGCACGTAGTGAATAAACTATGTCTCTGTGTTTGG 1750
CTCCCTGCC 1759
23 lainnya. Tidak ditemukan variasi pada daerah ujung 5’ dan 3’ intron karena situs nukleotida yang paling conserved berada di bagian ujung 5’ dan 3’ intron yang berdekatan dengan ekson (Hare and Palumbi 2003; Lynch 2006). Variasi yang ditemukan pada daerah intron gen BCKDHA pada penelitian ini tidak menyebabkan perubahan asam amino.
Tabel 6 Variasi yang ditemukan pada ruas ekson gen BCKDHA pooled samples
Jenis Sampel Jumlah Sampel Mutasi Posisi Tipe Mutasi
Tabel 5 Variasi yang ditemukan pada ruas ekson gen BCKDHA sapi madura
Jenis Sampel Jumlah Sampel Mutasi Posisi Tipe Mutasi
165G>A Arg34Gln Ekson 1 Missense
Sapi sonok 1
Sapi pedaging 3 Sapi karapan 3
195T>C Phe44Ser Ekson 1 Missense
Sapi sonok 1
Sapi pedaging 1 109A>G Arg240Gly Ekson 6 Missense
Sapi pedaging 1 240G>A Ekson 6 Sense
Sapi karapan 1
266C>T Arg381Cys Ekson 8 Missense Sapi pedaging 1
Sapi sonok 1
24
Tabel 7 Variasi yang ditemukan pada ruas intron gen BCKDHA
Jenis Sampel Jumlah Sampel Mutasi Nukleotida Posisi Primer
Sapi karapan 2
Tabel 6 Variasi yang ditemukan pada ruas ekson gen BCKDHA pooled samples (lanjutan) Jenis Sampel Jumlah Sampel Mutasi Posisi Tipe Mutasi
165G>A Arg34Gln Ekson 1 Missense Sapi bali (RPH Bogor) 8
Sapi madura (RPH Bogor) 3 Sapi bali (BPTO Bali) 10
25 Tabel 7 Variasi yang ditemukan pada ruas intron gen BCKDHA (lanjutan) Jenis Sampel Jumlah Sampel Mutasi Nukleotida Posisi Primer sapi PO (Kebumen) 1
26
Tabel 8 Variasi yang ditemukan pada ruas intron gen BCKDHA pooled samples Jenis Sampel Jumlah Sampel Mutasi Nukleotida Posisi Primer
Sapi PO (Kebumen) 10
Sapi madura (Bangkalan) 8 598C>T Intron 3 AF503-504 Sapi bali (RPH Bogor) 8
27 Variasi yang ditemukan pada daerah penyandi kemungkinan mempengaruhi fungsi dan efisiensi kompleks enzim BCKDH dan transpor polipeptida dari sitosol ke mitokondria. Selain itu, variasi yang ditemukan pada ruas mature peptide subunit E1α kemungkinan akan mempengaruhi stabilitas mature peptide dan kemampuan untuk membentuk struktur tetramer (Davie et al. 1992). Berdasarkan variasi yang ditemukan pada ruas-ruas ekson gen BCKDHA pada sapi madura (Tabel 5), daerah transit peptide (bagian ujung 5’ ekson 1 sampai ujung 5’ ekson 2) memiliki variasi yang lebih tinggi dibandingkan dengan mature peptide (bagian ujung 3’ ekson 2 sampai ekson 9). Hal ini menunjukkan bahwa ruas transit peptide dan mature peptide pada sapi madura tidak conserved. Terdapat tiga variasi yang ditemukan pada daerah presekuen pada ruas transit peptide, ketiga variasi tersebut adalah 112G>C, 148G>T, dan 151C>G. Ketiga variasi tersebut merupakan mutasi sense, sehingga kemungkinan tidak mempengaruhi target pre-E1α ke mitokondria. Selain ketiga variasi tersebut, baik mutasi sense maupun missense lainnya ditemukan pada residu asam amino yaitu pada situs non aktif subunit E1α.
Hubungan antara Variasi Gen BCKDHA dengan Sejarah Sapi Madura
Variasi gen BCKDHA yang ditemukan pada sapi madura menunjukkan bahwa sapi madura merupakan hasil persilangan sapi lokal (sapi bali atau banteng) dengan sapi eksotis (sapi zebu dan taurin). Salah satu contohnya adalah variasi delesi 6 pb yang ditemukan pada ekson 1 menghasilkan alel yang berbeda (heterozigot). Alel 1 sapi madura memiliki urutan nukleotida yang sama dengan sapi bali, yaitu mengalami delesi 6 pb atau dua kodon (Met dan Ala), sedangkan alel 2 memiliki urutan nukleotida yang sama dengan sapi PO dan taurin (Gambar 13). Hal ini juga ditunjukkan oleh kromatogram (parsial) alel 1 dan 2 yang terdapat pada Gambar 14. Alel heterozigot ini ditemukan pada ketiga peruntukan sapi madura, yaitu karapan, sonok, serta pedaging, namun tidak semua sampel memiliki alel heterozigot (Tabel 5 dan 6). Nijman et al. (2003) menemukan bahwa sapi madura merupakan hasil hibridisasi antara banteng dengan sapi zebu berdasarkan penanda DNA mitokondria, DNA satelit, AFLP, dan mikrosatelit. Menurut Mohamad et al. (2009), berdasarkan penanda genetik molekuler kromosom Y (analisis paternal) terdapat komposisi genetik sapi zebu dan taurin pada sapi madura, sedangkan berdasarkan penanda genetik molekuler DNA mitokondria (analisis maternal) dan mikrosatelit (analisis autosomal) sapi madura memiliki komposisi genetik sapi zebu dan sapi bali atau banteng. Hal ini juga sesuai dengan hasil penelitian yang dilakukan oleh Firdhausi (2010) dan Febriana (2011) yang menunjukkan bahwa sapi madura berasal dari dua moyang betina yang berbeda.
28
kemudian bermigrasi ke Pulau Madura. Adaptasi iklim lokal dan kondisi geografis menyebabkan sapi jawa di Pulau Madura terisolasi dan membentuk suatu bangsa tersendiri, yaitu sapi madura. Faktor iklim dan kondisi lingkungan, seperti pakan, kemungkinan berpengaruh terhadap variasi yang hanya ditemukan pada sapi madura.
Tabel 10 Variasi basa nukleotida (ruas ekson) yang hanya ditemukan pada sapi madura (pooled samples)
Jenis Sampel Lokasi Sampel Jumlah Sampel Mutasi Nukleotida Posisi
Sapi pedaging Bangkalan 8 148G>T T Ekson 1
Sapi pedaging Bangkalan 8 196T>A A Ekson 1
Sapi pedaging Bogor 3 392A>G G Ekson 3
Sapi pedaging Bogor 3 92G>A A Ekson 5
Tabel 9 Variasi basa nukleotida (ruas ekson) yang hanya ditemukan pada sapi madura Jenis Sampel Lokasi Sampel Jumlah Sampel Mutasi Nukleotida Posisi
Sapi karapan Sampang 1 148G>T T Ekson 1
Tabel 11 Variasi basa nukleotida (ruas intron) yang hanya ditemukan pada sapi madura
Jenis Sampel Lokasi (Jumlah) Sampel Mutasi Nukleotida Posisi
Sapi pedaging Sampang (1) 8G>A A Intron 1
Sapi karapan Sampang (1) 598C>T T Intron 3
Sapi pedaging Bangkalan (1), Sampang (4)
Sapi karapan Sampang (1) 370A>G A Intron 5
Sapi pedaging Bangkalan (1), Sampang (3), Pamekasan (2)
Sapi karapan Sampang (1) 101T>C C Intron 7
Sapi sonok Pamekasan (1)
Sapi pedaging Sampang (2), Pamekasan (1)
Sapi karapan Sampang (2) 111T>C C Intron 7
Sapi pedaging Bangkalan (1), Sampang (3), Pamekasan (2)
Sapi karapan Sampang (1) 120G>A A Intron 7
Sapi sonok Pamekasan (1)
Sapi pedaging Sampang (2), Pamekasan (1)
29
Indel Simultan pada Intron 7 Gen BCKDHA
Tabel 13 menunjukkan sembilan variasi yang ditemukan pada intron 7. Posisi indel simultan pada intron 7 ditemukan pada posisi basa ke-34 dan 68. Berdasarkan variasi indel yang terjadi pada posisi tersebut, intron 7 pada penelitian ini dapat dikelompokkan menjadi tiga kelompok. Kelompok pertama adalah kelompok yang mengalami insersi basa G pada posisi ke-34 dan delesi basa C pada posisi ke 68 atau memiliki urutan yang sama dengan B. taurus (NW_001493616). Variasi insersi basa G dan delesi basa C pada posisi basa ke-34 dan 68 menunjukkan sapi PO, bali, dan madura (sonok, dan karapan 2) memiliki alel heterozigot. Perbedaan bentuk alel ini dilihat berdasarkan urutan nukleotida yang berbeda pada puncak kromatogram. Sebaliknya kelompok kedua adalah kelompok yang mengalami delesi basa G pada posisi ke-34 dan insersi basa C pada posisi ke-68. Variasi ini homozigot pada sapi madura karapan 1, pedaging 4, dan pedaging 5. Pada kelompok ketiga tidak terjadi delesi, sehingga kelompok ketiga memiliki basa G pada posisi ke-34 dan basa C pada posisi ke-68. Akan tetapi, variasi heterozigot hanya ditemukan pada posisi basa G ke-34. Variasi ini terdapat pada sapi pedaging 1, pedaging 2, pedaging 3, pedaging 6, dan karapan 3. Seperti halnya variasi delesi 6 pb pada ekson 1, variasi indel simultan ini tidak ditemukan khusus pada salah satu peruntukan sapi madura.
Tabel 13 Variasi nukleotida pada intron 7 gen BCKDHA Jenis Tabel 12 Variasi basa nukleotida (ruas intron) yang hanya ditemukan pada
sapi madura (pooled samples)
30
Variasi pada Sapi Madura Pedaging
Sebagian besar variasi yang ditemukan terdapat pada sapi pedaging menyebabkan sapi pedaging memiliki variasi yang paling tinggi dibandingkan dengan sapi karapan dan sonok. Tingginya variasi pada sapi pedaging kemungkinan disebabkan oleh persilangan yang tidak terkontrol. Berdasarkan hasil wawancara dengan peternak di Pulau Madura, sapi karapan berasal dari Pulau Sapudi yang terletak di bagian timur Pulau Madura. Pulau Sapudi merupakan sentra sapi karapan di Pulau Madura dan terpilih sebagai wilayah konservasi bagi pemurnian plasma nutfah sapi madura (Kutsiyah 2012). Sapi yang diperuntukkan sebagai sapi karapan dan sonok biasanya tidak dikawinkan. Akan tetapi, jika sapi tersebut sudah tua atau kalah dalam pertandingan, maka sapi tersebut akan dijadikan sebagai sapi pedaging dan dikawinkan. Hal ini menyebabkan komposisi nukleotida yang terdapat pada sapi pedaging merupakan campuran dari ketiga peruntukan sapi madura.
5
SIMPULAN DAN SARAN
Simpulan
Berdasarkan hasil pensejajaran urutan nukleotida gen BCKDHA pada sapi madura menunjukkan tidak ada diferensiasi basa nukleotida maupun fungsi gen. Diferensiasi yang ada tidak sampai mengarahkan pada pembentukan bangsa baru. Mutasi-mutasi gen yang ditemukan tidak terdiferensiasi antar peruntukan sapi. Tekanan seleksi berbasis peruntukan sapi karapan, sonok, dan pedaging tidak terbukti mengarahkan mutasi-mutasi pada gen BCKDHA.
Saran
Perlu dilakukan penelitian lebih lanjut mengenai sapi madura dan bangsa sapi lokal Indonesia lainnya dengan sampel yang lebih banyak dan asal daerah yang berbeda sehingga diperoleh hasil dan informasi yang lebih lengkap untuk mengetahui keragaman dan diferensiasinya berdasarkan gen BCKDHA. Informasi yang lengkap diperlukan untuk studi lebih lanjut mengenai fungsi dan efisiensi BCKDHA dalam proses metabolisme BCAA.
DAFTAR PUSTAKA
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. 2008. Molecular Biology of the Cell. 5th ed. New York (US): Garland Science.
Allison LA. 2007. Fundamental Molecular Biology. Oxford (UK): Blackwell Publishing Ltd.
31 Breathnach R, Benoist C, O'Hare K, Gannon F, Chambon P. 1978. Ovalbumin gene: evidence for a leader sequence in mRNA and DNA sequences at the exon-intron boundaries. Proc Natl Acad Sci USA. 75:4853-4857.
Byun SO, Fang Q, Zhou H, Hickford JGH. 2009. An effective method for silver staining DNA in large numbers of polyacrylamide gels. Anal Biochem. 385:174-175.
Cartegni L, Chew SL, Krainer AR. 2002. Listening to silence and understanding nonsense: exonic mutations that affect splicing. Nat Rev Genet. 3:285-298. Chuang JL, Cox RP, Chuang DT. 1993. Characterization of the
promoter-regulatory region and structural organization of E1α gene (BCKDHA) of human branched-chain α-ketoacid dehydrogenase complex. J Biol Chem. 268:8309-8316.
Cook KG, Bradford AP, Yeaman SJ, Aitken A, Fearnley IM, Walker JE. 1984. Regulation of bovine kidney branched-chain 2-oxoacid dehydrogenase complex by reversible phosphorylation. Eur J Biochem. 145:587-591.
Davie JR, Wynn RM, Cox RP, Chuang DT. 1992. Expression and assembly of a functional E1 component (α2β2) of mammalian branched chain α-ketoacid dehydrogenase complex in Escherichia coli. J Biol Chem. 267:16601-16606. De Jonge H. 1990. Of bull and men: the madurese aduan sapi. Bijdr Taal-, Land-
Volkenkd. 146:423-447.
Dennis JA, Healy PJ. 1999. Definition of the mutation responsible for maple syrup urine disease in poll shorthorns and genotyping poll shorthorns and poll herefords for maple syrup urine disease alleles. Res Vet Scie. 67:1-6.
Elsik CG, Tellam RL, Worley KC, and 306 co-authors [Bovine Genome Sequencing and Analysis Consortium]. 2009. The genome sequence of taurine cattle: a window to ruminant biology and evolution. Science. 324:522-528. Febriana A. 2011. Filogeni berdasarkan sekuens DNA mitokondria gen
cytochrome oxidase I (Gen COI) pada beberapa bangsa sapi lokal Indonesia [Skripsi]. Bogor (ID): Institut Pertanian Bogor.
Felius M. 1995. Cattle Breeds: An Encyclopedia Misset. Doetinchem, The Netherlands.
Fisher CR, Fisher CW, Chuang DT, Cox RP. 1991. Occurrence of a Tyr393-Asn (Y393N) mutation in the E1 alpha gene of the branched-chain α-keto acid dehydrogenase complex in maple syrup urine disease patients from a Mennonite population. Am J Hum Genet. 49:429-434.
Firdhausi NF. 2010. Asal usul sapi madura berdasarkan penanda DNA mitokondria [Tesis]. Bogor (ID): Institut Pertanian Bogor.
Furney SJ, Alba MM, Lopez-Bigas N. 2006. Differences in the evolutionary history of disease genes affected by dominant or recessive mutations. BMC Genomics. 7:165.
Geraads D. 1992. Phylogenetic analysis of the tribe bovine (mammalia: Artiodactyla). Zool J Linn soc. 104:193-207.
Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser. 41:95-98. Handiwirawan E, Subandriyo. 2004. Potensi keragaman sumberdaya genetik sapi
Bali. Wartazoa. 14:107-115.