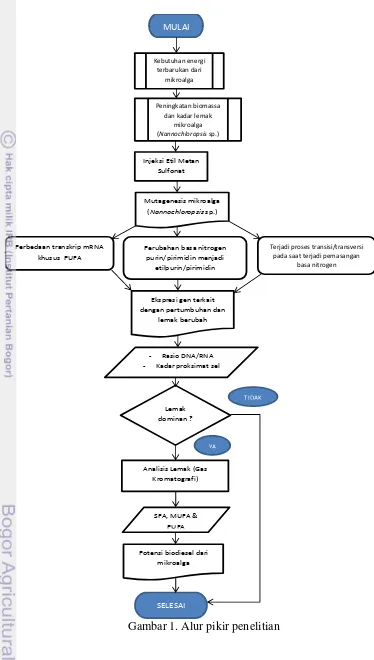
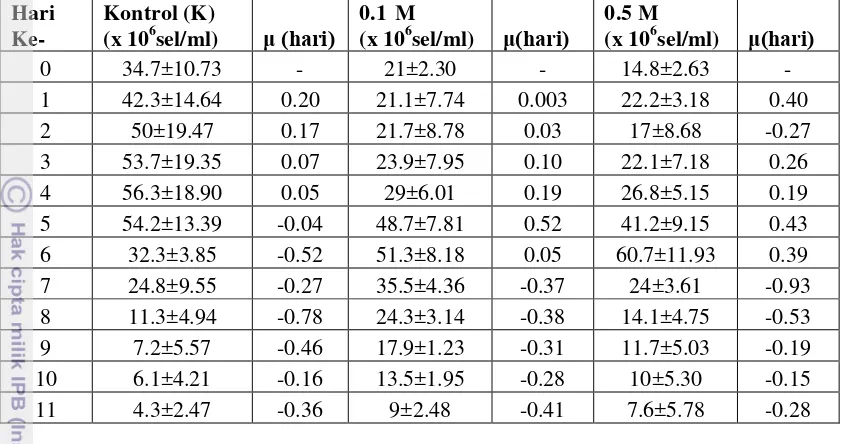
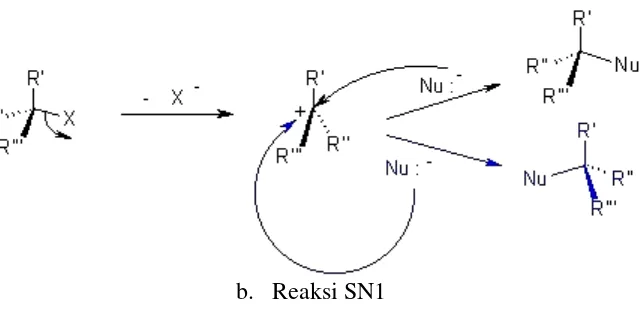
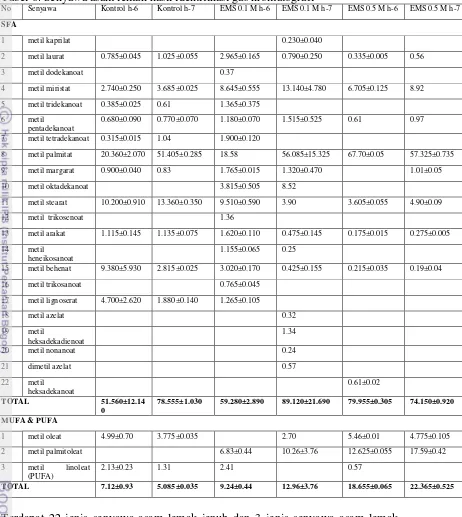
Pengembangan Mutagenesis Spesies Mikroalga Penghasil Biofuel dengan Metode EMS (Etil Metan Sulfonat)
Bebas
52
0
0
Teks penuh
Gambar

+2
Dokumen terkait
