BIOLOGI DAN POTENSI PREDASI TUNGAU PREDATOR Neoseiulus longispinosus Evans (ACARI: PHYTOSEIIDAE) PADA TUNGAU HAMA
Tetranychus kanzawai Kishida (ACARI: TETRANYCHIDAE)
MIA NURATNI YANTI RACHMAN A351080051
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN
Dengan ini saya menyatakan bahwa tesis “Biologi dan Potensi Predasi Tungau Predator Neoseiulus longispinosus Evans (Acari: Phytoseiidae) pada Tungau Hama Tetranychus kanzawai Kishida (Acari: Tetranychidae) ” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juli 2011
Mia Nuratni YR
ABSTRACT
MIA NURATNI YANTI RACHMAN. The Biology and Predation Rate of Predatory Mite, Neoseiulus longispinosus Evans (Acari: Phytoseiidae), on Its Prey
Tetranychus kanzawai Kishida (Acari: Tetranychidae). Supervised by SUGENG SANTOSO and PUDJIANTO
Predatory mite, Neoseiulus longispinosus Evans (Acari: Phytoseiidae), is a potensial predator of a polyphagous spider mite, Tetranychus kanzawai Kishida (Acari: Tetranychidae). Research was conducted to study the biological characteristics of N. longispinosus, including the life table, life cycle, fecundity,functional response, numerical response, prey prefference, and the ability of individual predatory mite in suppressing the prey population. The study revealed that the life cycle of N. longispinosus was short, i. e. 3.23 days. The oviposition period was 8.76 days with maximum daily egg production was 4 eggs/female/day when the predatory mite was 6 days old. When fed with T. kanzawai, the intrinsic rate of increase (rm) of N. longispinosus was 0.44, mean
generation time (T) was 4.05 days, and ner reproduction rate (R0) was 24.96
RINGKASAN
MIA NURATNI YANTI RACHMAN. Biologi dan Potensi Predasi Tungau Predator Neoseiulus longispinosus Evans (Acari: Phytoseiidae) pada Tungau Hama Tetranychus kanzawai Kishida (Acari: Tetranychidae). Dibimbing oleh SUGENG SANTOSO dan PUDJIANTO.
Tetranychus kanzawai Kishida termasuk dalam famili Tetranychidae dan merupakan spesies tungau hama yang sering menimbulkan kerugian secara ekonomis. Salah satu pengendalian tungau hama Tetranychidae adalah pemanfaatan tungau predator. Neoseiulus longispinosus merupakan agen hayati potensial untuk pengendalian tungau Tetranychidae. Tungau predator tersebut adalah predator lokal Indonesia yang belum dimanfaatkan secara maksimal. Penelitian ini dilakukan untuk melihat biologi dan potensi tungau predator N. longispinosus pada tungau hama T. kanzawai.
Neraca hayati merupakan salah satu percobaan yang dilakukan untuk mengetahui biologi tungau predator N. longispinosus. Sedangkan potensi predasi diketahui dengan melakukan percobaan tanggap fungsional, tanggap numerik, preferensi mangsa, kemampuan menekan populasi mangsa oleh individu betina, dan potensi pemangsaan predator selama masa perkembangan pradewasa. Data neraca hayati dianalisis dengan rumus parameter demografi. Tanggap fungsional diuji dengan fungsi polinom yang akan membedakan tanggap fungsional tipe II dan III. Sementara parameter lainnya dianalisis menggunakan excel dan SPSS.
Hasil analisis data neraca hayati menunjukkan bahwa N. longispinosus
memiliki nilai waktu generasi (T) sebesar 4.05 hari dengan laju reproduksi bersih (Ro) sebesar 24.96 butir telur per generasi. Hal tersebut mengisyaratkan bahwa
populasi N. longispinosus dapat berkembang sebanyak 24.96 kali dalam satu generasi selama 4.05 hari. Nilai laju pertambahan intrinsik rm adalah 0.44
betina/betina/hari dan laju pertambahan terbatas λ sebesar 1.55 betina/betina/hari. Distribusi sebaran umur N. longispinosus memperlihatkan bahwa sebagian besar populasi sekitar 83.78% merupakan populasi pradewasa dan siasnya 16.22% adalah populasi dewasa.
Waktu perkembangan N. longispinosus dari pradewasa menjadi dewasa realtif singkat yaitu 3.23 hari. Waktu tersebut merupakan waktu yang paling singkat karena pada umumnya tungau predator Phytoseiidae memiliki waktu perkembangan selama 4 hari. Masa oviposisi lebih singkat yaitu 8.76 hari namun keperidian yang diperoleh lebih tinggi. Nilai keperidian N. longispinosus yang dibiakkan pada T. kanzawai adalah 32.78 butir telur.
N. longispinosus memiliki kurva sintasan tipe I. Hal ini menandakan bahwa tingkat kematian N. longispinosus yang tinggi dialami saat N. longispinosus berumur tua. Mortalitas mulai terjadi saat imago N. longispinosus
berumur 3 hari. Tingkat produksi telur tertinggi dicapai saat tungau predator betina N. longispinosus berumur 6 hari dengan rata-rata produksi telur 4 butir.
Preferensi N. longispinosus terhadap mangsa tertentu akan meningkatkan perilaku pemangsaan yang tinggi. Stadia dan jenis mangsa mempengaruhi perilaku pemangsaan predator terhadap mangsa. Sebagian besar imago N. longispinosus lebih banyak dijumpai pada daun yang berisi telur tungau T. kanzawai. Hal ini menunjukkan bahwa N. longispinosus memiliki ketertarikan yang cukup tinggi terhadap mangsa dalam stadia telur.
N. longispinosus yang diberikan mangsa T. kanzawai memiliki tanggap fungsional tipe III. Tanggap fungsional tipe III ditunjukkan oleh koefisien linear yang bernilai positif. Hal ini menunjukkan bahwa pada awalnya proporsi mangsa yang dipredasi meningkat kemudian menurun seiring dengan bertambahnya kepadatan mangsa. Tanggap fungsional tipe III merupakan satu-satunya tipe tanggap fungsional yang memiliki kontribusi nyata dalam regulasi populasi hama sehingga N. longispinosus berpotensi sebagai agen biokontrol yang efisien.
Tanggap numerik N. longispinosus yang diberikan mangsa T. kanzawai
menunjukkan model respons langsung. Model respons langsung merupakan model yang menunjukkan dimana populasi predator akan meningkat seiring dengan peningkatan jumlah mangsa.
Kemampuan individu imago betina tungau predator N. longispinosus
© Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
BIOLOGI DAN POTENSI PREDASI TUNGAU PREDATOR Neoseiulus longispinosus Evans (ACARI: PHYTOSEIIDAE) PADA TUNGAU HAMA
Tetranychus kanzawai Kishida (ACARI: TETRANYCHIDAE)
MIA NURATNI YANTI RACHMAN
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Entomologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Biologi dan Potensi Predasi Tungau Predator Neoseiulus longispinosus (Acari: Phytoseiidae)
pada Tungau Hama Tetranychus kanzawai Kishida (Acari: Tetranychidae)
Nama : Mia Nuratni Yanti Rachman
NRP : A 351080051
Program Studi : Entomologi
Disetujui
Komisi Pembimbing
Dr. Ir. Sugeng Santoso, MAgr. Dr. Ir. Pudjianto, MSi
Ketua anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karuniaNya sehingga tesis ini berhasil diselesaikan. Terima kasih penulis ucapkan kepada Dr. Ir. Sugeng Santoso, MAgr dan Dr. Ir. Pudjianto, MSi selaku pembimbing. Selain itu, penulis sampaikan terima kasih kepada Bapak Wawan, teman-teman Entomologi 2008 (Yani, bang Dedi, kak Kiki, pak Umbu, mbak Nella, mbak Rika, pak Gatot, pak Aser), Nia, Putri, dan teman-teman Wisma Bintang (Nahrin dkk) yang membantu penulisan tesis baik secara langsung maupun tidak langsung. Ungkapan terima kasih juga penulis haturkan kepada Papa, Mama, suami serta seluruh keluarga atas segala doa dan kasih sayangnya. Penulis juga mengucapkan terima kasih kepada Departemen Pendidikan Tinggi Republik Indonesia atas beasiswa pascasarjana yang diperoleh.
Bogor, Juli 2011
Mia Nuratni YR
RIWAYAT HIDUP
Penulis dilahirkan di Bekasi pada tanggal 23 September 1982 dari ayah H. Taufik Rachman dan ibu Hj. Fatmah AF. Penulis merupakan putri pertama dari empat bersaudara.
Tahun 2001 penulis lulus dari SMA Negeri 1 Bekasi dan pada tahun yang sama lulus seleksi masuk IPB melalui Ujian Masuk Perguruan Tinggi Negeri. Penulis memilih program studi Hama dan Penyakit Tumbuhan, Fakultas Pertanian. Pada tahun 2006, penulis menamatkan sarjana.
DAFTAR ISI
Halaman
DAFTAR TABEL………... i
DAFTAR GAMBAR……….. ii
PENDAHULUAN………... 1
Latar belakang ... 1
Tujuan ... 2
TINJAUAN PUSTAKA ... 3
Tungau Merah Tetranychus kanzawai ... 3
Karakter Morfologi ... 3
Bioekologi ... 3
Tungau Predator Famili Phytoseiidae ... 4
Tungau Predator Neoseiulus longispinosus ... 6
Karakter Morfologi ... 6
Bioekologi ... 8
Neraca Hayati……… .. 9
Tanggap Fungsional………. 10
Tanggap Fungsional Tipe I ... 11
Tanggap Fungsional Tipe II ... 12
Tanggap Fungsional Tipe III ... 13
Tanggap Numerik……… 13
Preferensi Mangsa……… 14
BAHAN DAN METODE ... 17
Waktu dan Tempat ... 17
Metode Penelitian ... 17
Pemeliharaan Tungau ... 17
Neraca Hayati Tungau Predator N.longispinosus ... 18
Preferensi Mangsa ... 19
Tanggap Fungsional……… 20
Tanggap Numerik……… 21
Kemampuan Menekan Populasi Mangsa oleh Individu Predator……….. . 22
HASIL DAN PEMBAHASAN ... 23
Neraca Hayati ... 23
Siklus Hidup dan Perkembangan ... 25
Sintasan dan Jumlah Telur Predator Harian ... 28
Preferensi Stadia Mangsa ... 30
Tanggap Fungsional ... 31
Kemampuan Individu Imago Betina Tungau Betina Predator
N.longispinosus Menekan Populasi Tungau Hama T. kanzawai 36
PEMBAHASAN UMUM ... 41
KESIMPULAN DAN SARAN ... 43
Kesimpulan ... 43
Saran ... 43
DAFTAR PUSTAKA ... 45
DAFTAR TABEL
Halaman 1. Statistik demografi N. longispinosus... 24 2. Proporsi berbagai fase perkembangan N. longispinosus
pada persebaran umur stabil ... 25 3. Berbagai parameter kehidupan N. longispinosus yang
dibiakkan pada T. kanzawai ... 27 4. Hasil analisis regresi logistik proporsi mangsa
T. kanzawai yang mangsa N. longispinosus ... 31 5. Estimasi masa penanganan dan rataan jumlah mangsa
yang dimangsa N.longispinosus………. 33
6. Rataan predasi, produksi telur predator
dan ratio predator:mangsa………. 35 7. Predator dan mangsa yang tersisa pada hari terakhir
pengamatandi laboratorium……… 38 8. Jumlah kumulatif stadia predator yang ditemukan
DAFTAR GAMBAR
Halaman
1. Tetranychus kanzawai ... 3
2. Bagian dorsal Phytoseiidae ... 5
3. Bentuk dan jumlah seta pada bagian dorsal N. longispinosus ... 7
4. Tipe pola kawin N.longispinosus……… 8
5. N. longispinosus………. 9
6. Grafik tanggap fungsional tipe I……… 12
7. Grafik tanggap fungsional tipe II……….. 12
8. Grafik tanggap fungsional tipe III………. 13
9. Potongan daun ubi kayu………. 17
10. Sintasan dan rata-rata jumlah telur harian tungau predator……… 29
11. Keberadaan imago N. longispinosus selama 3 jam………. 31
12. Kurva tanggap fungsional………... 33
13. Proporsi pemangsaan pada tanggap fungsional tungau predator N. longispinosus………... 34
14. Laju rataan produksi telur predator pada kepadatan mangsa berbeda………. 36
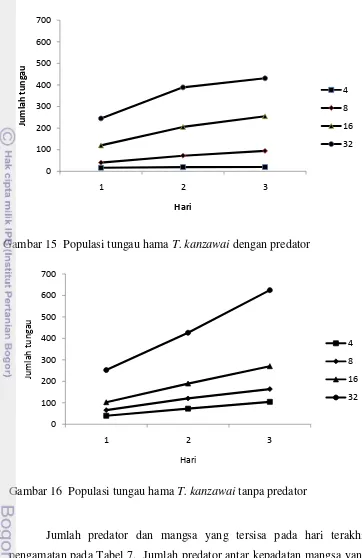
15. Populasi tungau hama T. kanzawai dengan predator……….. 37
PENDAHULUAN
Latar belakang
Salah satu famili tungau yang menjadi pusat perhatian dalam pertanian
adalah Tetranychidae. Sebagian besar spesies famili Tetranychidae merupakan
hama yang bersifat polifag (Kalshoven 1981; Walter & Proctor 1999; Zhang
2003). Tetranychus kanzawai Kishida merupakan salah satu spesies famili
Tetranychidae yang sangat merusak dan bersifat kosmopolit (Walter & Proctor
1999; Zhang 2003; Kasap 2005). Tungau ini dapat dijumpai di berbagai belahan
dunia dan menyerang lebih dari 100 spesies tanaman termasuk tanaman pertanian
dan tanaman hias (Zhang 2003). Tanaman yang terinfestasi T. kanzawai akan
memiliki gejala seperti berikut: permukaan bawah daun berwarna perak
kekuningan sampai kecoklatan, jaring-jaring menutupi daun dan batang serta
kering dan mati bila populasi tungau sangat padat (Zhang 2003).
Pengendalian hayati merupakan salah satu pengendalian yang dilakukan
untuk mengatasi masalah yang disebabkan oleh T. kanzawai. Pengendalian hayati
menggunakan musuh alami T. kanzawai seperti tungau predator dari famili
Phytoseiidae, sudah dilakukan di beberapa Negara. Beberapa tungau predator
bahkan telah dibiakkan secara komersial (Driesche & Bellows 1996; Zhang 2003).
Phytoseiulus persimilis dan Amblyseius californicus merupakan contoh spesies
yang telah diproduksi secara komersial (Luz 2003; Cakmak et al. 2004). Di
Indonesia, Yulianah (2008) melaporkan adanya tungau predator yang berasosiasi
dengan T. kanzawai pada tanaman stroberi di Cisarua dan Lembang. Tungau
predator yang ditemukan oleh Yulianah (2008) termasuk famili Phytoseiidae,
spesies Amblyseius longispinosus.
Neoseiulus (Amblyseius) longispinosus merupakan tungau predator lokal
Indonesia. Tungau predator ini pertama kali ditemukan di Indonesia pada tahun
1952, dan dilaporkan berasosiasi dengan Tetranychus bimaculatus pada tanaman
ubi kayu (Evans 1952). N. longispinosus mudah dijumpai pada tanaman yang
terinfestasi tungau Tetranychidae. Hal tersebut berkaitan dengan preferensi
mangsa dan tipe predator N. longispinosus yangtermasuk dalam tipe predator II,
Kongchuensin et al. 2006). Kelimpahan N. longispinosus relatif tinggi berkaitan
dengan karakter spesies ini yang dapat digunakan sebagai agens hayati tungau
Tetranychidae. Oleh karena itu, potensi N. longispinosus sebagai agens hayati
tungau Tetranychidae di Indonesia relatif tinggi.
Beberapa negara di Asia seperti Thailand, Philipina dan Taiwan telah
mempelajari dan memanfaatkan N. longispinosus sebagai agens hayati
pengendalian tungau. Sebaliknya, pengetahuan biologi dan ekologi tentang N.
longispinosus di Indonesia masih minim, padahal peluang penggunaan N.
longispinosus sebagai agens hayati cukup tinggi. Oleh karena itu, faktor-faktor
yang berkaitan dengan bioekologi predator ini perlu dipelajari untuk mengetahui
potensi tungau predator N. longispinosus dalam pengendalian tungau T. kanzawai
di Indonesia.
Tujuan
Penelitian ini bertujuan sebagai berikut: (1) mengetahui pertumbuhan dan
perkembangan tungau predator N. longispinosus yang meliputi neraca hayati,
lama hidup, dan fekunditas; (2) preferensi mangsa; (3) tanggap fungsional; (4)
tanggap numerik tungau predator; dan (5) kemampuan individu imago betina N.
TINJAUAN PUSTAKA
Tungau Merah Tetranychus kanzawai Karakter Morfologi
Siklus hidup T. kanzawai terdiri dari telur, larva, nimfa (protonimfa dan
deutonimfa) dan dewasa. Telur umumnya diletakkan pada permukaan bawah
daun tapi terkadang juga pada permukaan atas daun bila populasi T. kanzawai
berlimpah. Telur berbentuk bulat seperti bola dan saat baru diletakkan berwarna
putih bening. Larva dan nimfa berwarna hijau kekuningan dengan bintik gelap
pada bagian dorsolateral idiosoma seperti pada gambar 1 (Ehara 2002). Tungau
dewasa umumnya berwarna merah atau merah kekuningan (Ehara 2002). Warna
tubuh imago T. kanzawai terkadang dipengaruhi oleh tanaman inangnya. Tungkai
berwarna kekuningan. Betina dewasa berukuran sekitar 400-500 µm dan jantan
dewasa lebih kecil dengan hysterosoma yang meruncing. Imago T. kanzawai
jantan memiliki knob yang besar pada aedeagus (Zhang 2003).
Gambar 1 Tetranychus kanzawai (a, pradewasa; b, dewasa; sumber Ehara, 2002)
Bioekologi
T. kanzawai pertama kali ditemukan pada tanaman murbei di Jepang
(Kishida 1927). Walter & Proctor (1999) menyatakan bahwa sebelum perang
dunia II, tungau ini merupakan hama sekunder. Penggunaan pestisida kimiawi
secara intensif menyebabkan perubahan status pada spesies ini.
Spesies tungau ini bersifat kosmopolit dan dapat dijumpai hampir di
seluruh belahan dunia. T. kanzawai merupakan spesies tungau hama yang cukup
terkenal di Asia. Tungau ini mudah dijumpai pada pertanaman teh sehingga
dikenal juga sebagai tungau merah teh. Selain itu, T. kanzawai dapat menyerang
lebih dari 100 spesies tanaman. Pada umumnya tungau ini mudah dijumpai di
lapangan, namun juga menjadi hama pada pertanaman dalam rumah kaca seperti
anggur, stroberi, dan lain-lain.
Gejala kerusakan yang diakibatkan oleh tungau hama ini bervariasi
tergantung jenis tanamannya. Nekrotik merupakan gejala yang pasti terjadi pada
daun yang terserang tungau hama ini, kemudian daun tersebut mengering.
Populasi tungau yang sangat tinggi dapat menyebabkan kematian tanaman.
Populasi T. kanzawai dapat meningkat dalam waktu yang cepat. Hal ini
berkaitan dengan waktu perkembangan T. kanzawai yang singkat, yaitu berkisar
12-19 hari pada suhu 20-25°C (Zhang 2003). Keberhasilan hidup sampai tahap
imago dapat mencapai 80 %. Nisbah kelamin bersifat female biased dengan nilai
1:3. Imago betina memiliki lama hidup yang lebih panjang dibandingkan imago
jantan. Tingkat fekunditas bervariasi dan dipengaruhi oleh suhu. Satu imago
betina dapat bertelur sebanyak 28-76 butir pada kisaran suhu 15-30°C (Zhang
2003).
Tungau Predator Famili Phytoseiidae
Kelompok tungau predator yang banyak digunakan sebagai agens
pengendali hama tanaman berasal dari famili Phytoseiidae. Selain memakan
tungau fitofag, tungau predator famili Phytoseiidae juga memakan serangga kecil
yang berada di tanaman. Beberapa spesies juga memakan nematoda, spora
cendawan, polen, dan eksudat tanaman.
Famili Phytoseiidae memiliki tiga subfamili yaitu Amblyseiinae,
Phytoseiinae, dan Typhlodrominae. Spesies tungau yang telah dikembangkan
secara komersial adalah genera Neoseiulus dan Phytoseiulus yang termasuk dalam
subfamili Amblyseiinae dan Phytoseiinae (Zhang 2003).
Siklus hidup tungau predator terdiri dari telur, larva, protonimfa,
deutonimfa, dan imago. Telur memiliki bentuk oval memanjang dan berwarna
bening. Kelembapan yang tinggi yaitu berkisar 90-100%, dibutuhkan untuk
penetasan telur. Perilaku makan larva berbeda untuk beberapa spesies. Beberapa
beberapa spesies membutuhkan makanan untuk perkembangannya. Pada
umumnya perkembangan tungau predator lebih cepat dibandingkan dengan
tungau Tetranychus sp. Sebagian besar tungau predator membutuhkan waktu
sekitar satu minggu untuk perkembangannya. Beberapa spesies Phytoseiulus
bahkan dapat menyelesaikan siklus hidupnya dalam waktu 4 hari.
Famili Phytoseiidae bersifat pseudo-arrhenotokous, yaitu menghasilkan
keturunan jantan haploid dari telur yang dibuahi yang akan kehilangan genom
induk pada awal perkembangan (Walter & Proctor 1999). Oleh karena itu,
kopulasi sangat penting dalam reproduksi. Nisbah kelamin jantan:betina adalah
1:3 (Zhang 2003). Watson (2008) menjelaskan bahwa secara morfologi
perbedaan antara tungau betina dan jantan terletak pada bagian lapisan pelindung
ventral. Tungau jantan hanya memiliki satu lapisan ventral sedangkan tungau
betina memiliki tiga lapis pelindung, yaitu sternal, genital dan anal.
Seta merupakan salah satu unsur dalam klasifikasi tungau. Beberapa
peneliti memiliki penamaan letak seta pada idiosoma dorsal tungau. Pada gambar
2, Zhang (2003) mendeskripsikan ciri khas pada famili Phytoseiidae yaitu
idiosoma bagian dorsal memiliki tidak lebih dari 24 pasang seta dan pada bagian
J1, J3, serta J4 tidak terdapat seta.
Salah satu ciri khas tungau predator adalah pergerakannya yang cepat. Hal
tersebut disebabkan oleh tungkai tungau predator yang relatif panjang. Olfaktori
sangat berguna dalam pencarian mangsa sehingga tungau predator dapat
mengetahui tanaman yang terinfestasi oleh tungau fitofag (Boom et al. 2002;
Zhang 2003, Nachappa 2008 ).
Tungau Predator Neoseiulus longispinosus Karakter Morfologi
N. longispinosus termasuk dalam famili Phytoseiidae, ordo Mesostigmata.
Gerson et al. (2003) menyatakan bahwa N. longispinosus sangat berhubungan
dekat dengan N. womersleyi secara biosistematika. Penampakan morfologi secara
kasat mata hampir sama untuk kedua predator ini. Bentuk tungau betina N.
longispinosus lebih besar dibandingkan tungau betina N. womersleyi (Gerson et
al. 2003). Zhang (2003) menambahkan bahwa N. longispinosus memiliki tekstur
seta lebih halus dan panjang pada seta S5.
Siklus hidup N. longispinosus terdiri dari telur, larva, protonimfa,
deutonimfa dan dewasa. Telur berbentuk oval dan transparan serta berwarna
putih bening. Perubahan warna telur menjadi putih agak keruh terjadi saat
menjelang penetasan. Telur diletakkan secara individu pada permukaan bawah
daun. Stadia telur berlangsung selama 1-2 hari (Puspitarini 2005; Yulianah 2008).
Larva N. longispinosus berwarna putih dengan 3 pasang tungkai. Pada
stadia larva, predator tidak mengkonsumsi mangsa. Mobilitas larva terbilang
pasif karena cenderung lebih banyak diam. Masa stadia larva pada umumnya
relatif singkat dan biasanya hanya dalam hitungan jam.
Stadia nimfa terdiri dari protonimfa dan deutonimfa. Pada stadia ini,
predator lebih aktif dalam mobilitas dan memangsa. Nimfa berwarna putih agak
keruh dan memiliki 4 pasang tungkai. Setelah memangsa, warna nimfa berubah
menjadi putih kekuningan atau kemerahan pada bagian dorsal. Lama stadia nimfa
biasanya berlangsung selama satu hari.
Tungau dewasa memiliki banyak seta pada bagian dorsal. Lapisan dorsal
memiliki 17 pasang seta. Gambar 3 menunjukkan bahwa seluruh seta pada bagian
dorsal berukuran panjang dan berduri kecuali seta pada J1 dan S5 (Zhang 2003).
Tungau dewasa memiliki warna yang sama seperti pada stadia nimfa. Tungau
betina dewasa memiliki ukuran rata-rata sekitar 350 µm. Ukuran tungau dewasa
jantan lebih kecil dibandingkan tungau dewasa betina. Perbedaan tungau jantan
dan betina terletak pada bagian genitalia. Tungau jantan memiliki kaliks
spermateka berbentuk seperti botol. Lama hidup tungau jantan lebih pendek
Gambar 3 Bentuk dan jumlah seta pada bagian dorsal N. longispinosus (Zhang 2003)
Kopulasi terjadi ketika tungau betina menjadi dewasa. Tungau dewasa
jantan akan menunggu deutonimfa betina. Saat penantian tersebut, tungau jantan
akan menjaga area di sekeliling deutonimfa berada. Vantornhout (2006)
menyatakan bahwa detonimfa memiliki feromon seks yang dapat menarik tungau
jantan. Apabila tungau jantan lain memasuki area tersebut maka akan terjadi
pertarungan. Perilaku kawin tungau jantan N. longispinosus cukup unik (Gambar
4). N. longispinosus memiliki pola kawin tipe Phytoseiulus. Tipe Phytoseiulus
memiliki karakter saling berhadapan lalu tungau jantan akan merayap secara
Gambar 4 Tipe pola kawin N. longispinosus (Vantornhout 2006)
Bioekologi
Neoseiulus longispinosus dilaporkan berada di Indonesia pertama kali
dengan nama Typhlodromus longispinosus (Evans 1952). Kongchuesin et al.
(2005) menyatakan bahwa populasi N. longispinosus akan melimpah pada
tanaman yang terinfestasi tungau merah dengan produksi jaring yang banyak pada
permukaan bawah daun. Predator ini banyak dijumpai pada tanaman ubi kayu
yang terinfestasi tungau Tetranychidae terutama T. kanzawai di Indonesia
(Santoso, komunikasi pribadi). Selain itu, predator ini juga ditemui pada tanaman
stroberi dan jeruk di lapangan (Puspitarini 2005; Yulianah 2008). N.
longispinosus ditemukan pada 33 spesies tanaman di Thailand (Kongchuesin et al.
Gambar 5 N. longispinosus (a, telur; b, tungau dewasa; Koleksi Pribadi)
N. longispinosus banyak ditemui di beberapa negara seperti di India, Cina
bagian timur, Philiphina, Indonesia, Thailand, Malaysia, Taiwan, Hawaii,
Pakistan, Papua Nugini, Australia dan New Zealand (Gerson et al. 2003;
Kongchuensin et al. 2005; Raza 2008).
Masa siklus hidup N. longispinosus dipengaruhi oleh kondisi lingkungan
terutama suhu. Zhang (2003) melaporkan bahwa perkembangan N. longispinosus
berlangsung selama 5 hari pada suhu 28 °C. Penelitian Puspitarini (2005) dan
Yulianah (2008) menunjukkan hasil yang sama bahwa siklus hidup N.
longispinosus berlangsung selama 4-5 hari dalam kondisi laboratorium. Hal ini
memperlihatkan bahwa siklus hidup N. longispinosus lebih cepat dibandingkan
siklus hidup tungau Tetranychidae.
Neraca Hayati
Neraca hayati merupakan ringkasan pernyataan tentang kehidupan
individu dalam populasi atau kelompok (Price 1997). Lincoln et al. (1982)
mendefinisikan neraca hayati sebagai tabulasi data mortalitas lengkap dari
populasi terhadap umur. Neraca hayati merupakan riwayat perkembangan cohort
yang bersifat dinamis (Tarumingkeng 1992). Neraca hayati berisi informasi dasar
tentang mortalitas dan kelangsungan hidup suatu populasi dalam penjelasan
statistik. Informasi tersebut diperlukan untuk mengetahui dinamika populasi
Pertumbuhan populasi suatu organisme akan dipengaruhi oleh tiga faktor
yaitu kelahiran, kematian dan migrasi. Pertumbuhan populasi positif terjadi bila
angka kelahiran lebih besar daripada angka kematian dan migrasi bernilai 0
(angka emigrasi = angka imigrasi). Apabila terjadi sebaliknya maka akan terjadi
pertumbuhan populasi negatif (Oka 1995).
Neraca hayati digolongkan menjadi dua tipe yaitu neraca hayati horizontal
yang lebih bersifat spesifik umur dan neraca hayati vertikal yang bersifat spesifik
waktu (Bellows & Van Driesche 1992). Neraca hayati horizontal meliputi
penghitungan berulang terhadap suatu kelompok (cohort) tunggal yang terdiri dari
umur individu yang sama. Data yang berasal dari suatu kejadian tunggal yang
diasumsikan bahwa semua generasinya saling lingkup dengan sempurna karena
kelas umur yang secara simultan sama, merupakan neraca kehidupan vertikal.
Parameter-parameter yang terdapat dalam neraca hayati meliputi laju
reproduktif kotor (GRR), laju reproduktif bersih (Ro), waktu generasi (T), laju
pertambahan intrinsik (r), laju pertambahan terbatas (λ) dan doubling time (DT) (Rauf dan Hidayat 1987). Parameter tersebut berisi informasi dasar seperti
keperidian, kemampuan hidup harian, nisbah kelamin dan laju pertambahan suatu
organisme dalam analisa dinamika populasi. Laju reproduktif kotor (GRR) adalah
rata-rata jumlah keturunan betina per generasi (Σ mx). Laju reproduktif bersih
(Ro) menunjukkan jumlah keturunan betina yang berhasil menjadi imago. Waktu
generasi (T) merupakan waktu yang dibutuhkan untuk menyelesaikan siklus hidup
per generasi. Laju pertambahan intrinsik (r) menggambarkan laju pertambahan
populasi pada keadaan lingkungan konstan, sumber daya tak terbatas serta
kematian yang terjadi hanya disebabkan oleh faktor fisiologi (Birch 1948). Laju pertambahan terbatas (λ) menunjukkan nilai kelipatan populasi organisme per hari. Doubling time (DT) merupakan kemampuan organisme berkembang dalam
satu generasi. Pada umumnya tungau predator famili Phytoseiidae memiliki nilai
laju pertambahan intrinsik yang berkisar dari 0.1 - 0.4 (Escudero LA & Ferragut
F. 2005; Vasconcelos et al. 2008).
Tanggap Fungsional
Keberhasilan pengendalian hayati ditentukan oleh dinamika interaksi
predator-mangsa. Perubahan jumlah mangsa dapat direspons oleh predator.
Peningkatan jumlah generasi predator (tanggap numerik) dan tingkat predasi
predator secara individu (tanggap fungsional) merupakan respon predator
terhadap perubahan jumlah mangsa (Taylor 1984).
Tanggap fungsional merupakan respon perilaku predator terhadap
perubahan jumlah mangsa dalam waktu yang relatif singkat. Keefektifan predator
atau parasitoid dapat dilihat dari tanggap fungsionalnya. Salah satu ciri predator
yang baik adalah memiliki tanggap fungsional yang tinggi.
Tanggap fungsional merupakan komponen yang sangat esensial dari
dinamika interaksi antara predator/parasitoid dan mangsa/inang serta sangat
penting untuk determinasi stabilitas dari sistem yang dikelola (Oaten & Murdoch
1975 dalam Wang & Ferro 1998). Tanggap fungsional menggambarkan
hubungan antara jumlah mangsa/inang yang dikonsumsi/diparasit per
predator/parasitoid dan kepadatan mangsa/inang (Wang & Ferro 1998; Speight
1999). Holling 1959 dalam Hassel 2000 menggolongkan tanggap fungsional
menjadi tiga tipe: linier (Tipe I), hiperbolik (Tipe II), dan sigmoid (Tipe III).
Tanggap Fungsional Tipe I
Tanggap fungsional tipe I memiliki grafik bersifat linier. Hal ini
menunjukkan hubungan yang bersifat konstan. Tingkat predasi meningkat secara
linier dengan peningkatan kepadatan mangsa, kemudian tingkat predasi menjadi
konstan setelah predator berada dalam kondisi kenyang.
Tipe I berasal dari modifikasi sederhana tanggap fungsional linier dari
persamaan Lotka-Volterra. Tipe I dijumpai pada interaksi yang stabil. Tanggap
fungsional tipe I biasa ditemukan pada predator yang bersifat pasif seperti
Gambar 6 Grafik tanggap fungsional tipe I. Hubungan antara mangsa yang dimakan (Ne) dengan kepadatan mangsa yang tersedia (N) pada gambar A; hubungan antara proporsi mangsa yang dimakan (Ne/N) dan kepadatan mangsa yang tersedia (N) pada gambar B (Vantornhout 2006)
Tanggap Fungsional Tipe II
Tanggap fungsional tipe II memiliki grafik yang bersifat hiperbolik.
Model tipe II berasal dari persamaan cakram Holling. Tingkat predasi meningkat
seiring dengan peningkatan kepadatan mangsa secara konstan pada awalnya
hingga kepadatan mangsa maksimum. Penurunan tingkat predasi akan terjadi
secara cepat seiring meningkatnya mangsa sehingga terjadi bentuk grafik yang
hiperbolik.
Pada tanggap fungsional tipe II terdapat waktu penanganan dan laju
pemangsaan. Tanggap fungsional tipe II juga mudah ditemukan dalam kondisi
lingkungan yang stabil. Grafik tipe II umumnya ditemukan pada predator atau
parasitoid.
Tanggap Fungsional Tipe III
Tanggap fungsional tipe III memiliki grafik sigmoid. Tingkat predasi
bersifat cekung pada kepadatan mangsa rendah, tapi akan bersifat cembung pada
kepadatan mangsa tinggi. Tanggap fungsional tipe III dapat terjadi karena
pembelajaran hal baru, perubahan kemampuan, atau hal lain yang belum diketahui
yang terkadang disebut sebagai ekspresi preferensi. Sebagian besar proses
menyertai perubahan nutrisi dari satu tipe mangsa ke tipe mangsa lainnya.
Tanggap fungsional tipe III biasanya terjadi pada lingkungan sekitar kepadatan
mangsa yang seimbang. Ketika kepadatan mangsa bertambah banyak, predator
pun meningkat dan pengaruh stabilisasi lain mengakibatkan perilaku predator
hilang. Stabilitas pada sistem tanggap fungsional tipe III dipengaruhi seluruh
komponen dari biologi spesies dan interaksi antar spesies tersebut (Taylor 1984).
Grafik tipe III umumnya terdapat pada predator yang memangsa beberapa spesies
(Sharov 1996 dalam Hidrayani 2002).
Model tanggap fungsional tipe III menggambarkan bentuk grafik secara
sigmoid. Pada awalnya predasi terjadi secara lambat kemudian meningkat cepat
seiring bertambahnya kepadatan mangsa lalu tingkat predasi akan menurun pada
kepadatan mangsa yang lebih tinggi lagi hingga mencapai kejenuhan.
Tanggap Numerik
Setiap makhluk hidup membutuhkan nutrisi untuk pertumbuhan dan
perkembangannya. Sumber daya makanan yang berlimpah akan memberikan
keuntungan bagi makhluk hidup tersebut. Nutrisi makanan yang berlimpah akan
mempengaruhi tingkat reproduksi makhluk hidup dan secara tidak langsung
berkontribusi terhadap generasi berikutnya.
Peningkatan populasi mangsa dapat menyebabkan perubahan laju
penyerangan per individu predator. Selain itu, peningkatan populasi mangsa juga
dapat mengakibatkan perubahan kepadatan populasi predator. Perubahan populasi
predator ini merupakan respon atau tanggap terhadap peningkatan populasi
mangsa. Respon atau tanggap ini disebut sebagai tanggap numerik.
Tarumingkeng (1992) menguraikan mekanisme terjadinya tanggap
numerik sebagai berikut. Pertama, peningkatan populasi predator karena imigrasi
yang berasal dari daerah sekeliling. Hal ini berkaitan dengan perilaku predator
yang berkelompok dan menempati daerah-daerah dengan tingkat kerapatan
populasi predator yang tinggi. Sekelompok burung yang bergerombol di tempat
dengan kepadatan populasi belalang yang tinggi. Kedua, peningkatan populasi
predator karena peningkatan reproduksi (Ro). Waktu generasi (Ro) predator
umumnya lebih lama daripada waktu generasi (Ro) mangsa. Hal tersebut
menimbulkan penundaan dalam perubahan keterpautan kepadatan atau senjang
waktu (lag). Senjang waktu (lag) menyebabkan terjadinya peningkatan
reproduksi predator.
Tanggap numerik dibatasi oleh waktu generasi makhluk hidup tersebut.
Makhluk hidup yang memiliki siklus hidup yang relatif pendek cenderung
memiliki respon lebih cepat dengan tingkat fluktuasi terhadap kelimpahan sumber
Preferensi Mangsa
Makanan merupakan salah satu komponen yang mempengaruhi suatu
makhluk hidup dalam kehidupan seperti bertahan dan berkembang. Kualitas dan
kuantitas makanan adalah aspek penting untuk diperhatikan dalam pertumbuhan
dan perkembangan makhluk hidup. Kualitas makanan akan berkaitan langsung
dengan fisiologi makhluk hidup. Keberadaan jumlah makanan akan
mempengaruhi kelimpahan populasi suatu makhluk hidup.
Mangsa merupakan sumber daya nutrisi penting bagi predator. Mangsa
yang berbeda dapat mempengaruhi pertumbuhan dan perkembangan predator.
Oleh karena itu, tungau predator memiliki preferensi mangsa. Berdasarkan
preferensi mangsa, tungau predator dapat digolongkan dalam 4 tipe (Zhang 2003),
yaitu: 1) predator spesialis, hanya memakan spesies Tetranychus yang
menghasilkan sarang yang besar contoh spesies dari genera Phytoseiulus; 2)
tungau Phytoseiidae yang memiliki preferensi makan tungau Tetranychinae,
terkadang tungau kecil, dan polen contoh spesies Neoseiulus californicus
(McGregor); 3) predator generalis yang memakan berbagai jenis tungau, polen,
dan serangga tapi tidak dapat mengendalikan spesies Tetranychus yang
menghasilkan sarang yang besar contoh Iphiseius degenerans Berlese; dan 4)
predator generalis tungau dan serangga tapi bersifat spesialis terhadap polen
contoh spesies dari genera Euseius. N. longispinosus termasuk predator tipe 2,
yang memiliki preferensi mangsa pada tungau Tetranychidae dan juga dapat
memakan polen (Gerson et al. 2003; Zhang 2003).
BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilaksanakan pada bulan April-September 2010 di Laboratorium
Ekologi, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian
Bogor.
Metode Penelitian Asal Tungau
Tungau hama T. kanzawai dan tungau predator N. longispinosus diperoleh
dari tanaman ubi kayu di daerah Dramaga, dan dibiakkan di laboratorium dalam
suatu arena.
Arena Percobaan
Arena percobaan berupa petri dish berdiameter 8 cm. Busa (diameter ± 7
cm) diletakkan dalam arena lalu kapas diletakkan di atas busa dan diberikan air
hingga jenuh. Setelah itu, potongan daun ubi kayu yang berukuran 3 cm x 3 cm
diletakkan di atas kapas. Air ditambahkan pada arena percobaan apabila kapas
atau busa mulai terlihat kering. Potongan daun ubi kayu dalam arena percobaan
diganti setiap 3 hari.
Gambar 9 Potongan daun ubi kayu
Pemeliharaan Tungau
Tungau hama T. kanzawai dan tungau predator N. longispinosus dipelihara
dalam laboratorium dengan suhu 25-29°C dan RH 60-70%, menggunakan
Neraca Hayati Tungau Predator N. longispinosus
Percobaan neraca hayati bertujuan untuk mengetahui parameter demografi,
siklus hidup, perkembangan, sintasan dan sebaran umur tungau predator N.
longispinosus. Seratus telur predator ditempatkan secara individual pada arena
percobaan yang baru. Semua telur predator yang digunakan berasal dari umur
yang sama. Satu hari sebelum perlakuan, tiga ekor tungau betina T. kanzawai
juga ditempatkan dalam setiap arena percobaan dan dibiarkan bertelur.
Telur-telur yang dihasilkan dijadikan mangsa bagi predator dalam arena percobaan.
Metode pengamatan terbagi menjadi dua tahap yaitu stadia pradewasa dan
dewasa. Pada stadia pradewasa, pengamatan dilakukan setiap 6 jam untuk melihat
lama pergantian stadia dan jumlah predator yang masih hidup. Pengamatan pada
stadia dewasa dilakukan setiap 24 jam. Tungau dewasa jantan dan betina
dipasangkan. Setelah itu, pengamatan dilakukan dengan melihat parameter seperti
fekunditas, lama hidup, masa praoviposisi, oviposisi, dan pascaoviposisi. Selama
pengamatan berlangsung, tungau predator diberi mangsa berbagai stadia tungau
hama yang berlimpah. Penggantian daun dilakukan setiap 3 hari.
Analisis data neraca hayati tipe kohort menggunakan tabel kehidupan dan
grafik klasifikasi kurva keberhasilan hidup. Data biologi dianalisis menggunakan
excel.
Perumusan neraca hayati merupakan langkah pertama dalam menghitung
laju pertambahan intrinsik (r). Perhitungan parameter r didasarkan hanya pada
populasi betina, dan diasumsikan bahwa jantan cukup tersedia di sekitarnya.
Beberapa parameter yang dibutuhkan dalam perhitungan tersebut adalah sebagai
berikut (Tarumingkeng 1992):
1. x adalah kelas umur kohort (hari);
2. ax adalah jumlah individu yang hidup pada setiap umur pengamatan;
3. lx adalah proporsi individu yang hidup pada umur x (l : living, lx = ax/a0);
4. dx adalah jumlah individu yang mati di setiap kelas umur (d. : death, dx = lx–
lx+1);
5. qx adalah proporsi mortalitas pada masing-masing umur (qx = dx/ax);
6. Lx merupakan jumlah rata-rata individu pada kelas umur x dan kelas umur
7. Tx adalah jumlah individu yang hidup pada kelas umur x = 0 …w (x = w
adalah kelas umur terakhir) ( ); , T1 = T0 – L0, T2 = T1
– L1, Tx = Tx-1– Lx-1;
8. ex adalah harapan hidup individu pada setiap kelas umur x (ex = Tx/lx);
9. mx adalah keperidian spesifik individu-individu pada kelas umur x atau
jumlah anak betina perkapita yang lahir pada kelas x;
10. lxmx adalah banyaknya anak yang dilahirkan pada kelas umur x, lxmx
merupakan proporsi banyaknya anak (betina) dilahirkan oleh semua individu
(betina) sepanjang generasi kohort dan disebut laju reproduksi bersih (R0);
11. xlxmx adalah perkalian x, lx, dan mx untuk setiap kelas umur x yang
digunakan untuk mengaproksimasi lamanya generasi (Tc);
12. px (peluang survival) adalah proporsi individu yang hidup pada kelas umur x
dan mencapai kelas umur x + 1 (px = Lx+1/Lx). Parameter ini digunakan
dalam matriks proyeksi Leslie untuk memprediksi pertumbuhan populasi
secara diskrit.
Dari data neraca hayati tersebut perhitungan dilanjutkan untuk
menentukan parameter-parameter demografi lainnya (Price 1997) seperti:
1. Laju reproduksi kotor (GRR) = ∑ mx
2. Laju reproduksi bersih (Ro) = ∑ lxmx
3. Waktu generasi (Tc) = ∑ x lxmx / ∑ lxmx
4. Laju pertambahan intrinsik = Log e R0/Tc
Preferensi Tungau Predator terhadap Mangsa
Percobaan ini dilakukan untuk mengetahui preferensi mangsa tungau
predator N. longispinosus pada tingkat stadia (telur, nimfa, dan imago) tungau
hama T. kanzawai. Stadia mangsa yang digunakan dalam percobaan adalah telur,
nimfa, dan imago T. kanzawai. Jumlah telur, nimfa, dan imago yang digunakan
berbeda berturut-turut 25 butir, 10 ekor dan 5 ekor. Perbedaan jumlah tersebut
didasarkan pada jumlah maksimum pada uji tanggap fungsional. Perlakuan
diulang sebanyak 10 kali.
Tungau predator yang digunakan berumur sama yaitu imago berumur 2
percobaan dimodifikasi dalam percobaan preferensi mangsa. Tiga potongan daun
ubi kayu berukuran 1.5 cm x 1.5 cm diletakkan dalam satu arena percobaan.
Setiap helai daun berisi mangsa (telur, nimfa dan imago) yang berbeda dengan
jumlah mangsa yang sama. Jarak antar daun berkisar 0.5-1.0 cm. Penghubung
antar daun digunakan jembatan parafilm selebar 2 mm dan berbentuk T. Tungau
predator diletakkan di tengah jembatan. Pengamatan dilakukan setiap 30 menit
selama 3 jam dengan menghitung jumlah mangsa dan keberadaan tungau predator
pada tiap helai daun dalam arena percobaan.
Data percobaan preferensi mangsa dianalisis menggunakan excel dengan
melihat keberadaaan imago N. longispinosus pada arena percobaan setiap 30
menit selama 3 jam.
Tanggap Fungsional
Percobaan tanggap fungsional merupakan percobaan yang dilakukan untuk
mengetahui hubungan antara tingkat predasi tungau predator N. longispinosus
dengan tingkat kepadatan tungau hama T. kanzawai. Mangsa yang digunakan
dalam percobaan ini adalah stadia telur. Kepadatan telur yang digunakan adalah
5, 10, 20, 40 dan 80 butir telur. Masing-masing perlakuan diulang 10 kali.
Percobaan tanggap fungsional menggunakan arena percobaan seperti pada
percobaan neraca hayati. Imago T. kanzawai dimasukkan dalam arena percobaan
tersebut dan dibiarkan selama 24 jam agar bertelur dan telur-telur tersebut yang
digunakan dalam uji tanggap fungsional.
Predator yang digunakan berasal dari stadia imago yang berumur 2 hari.
Predator-predator tersebut dipuasakan selama 8 jam secara individu. Pemuasaan
tersebut bertujuan agar pencernaan setiap predator dalam kondisi yang sama saat
dilakukan perlakuan. Setelah 8 jam, predator tersebut dimasukkan dalam arena
percobaan yang berisi telur dan dibiarkan selama 24 jam. Setelah itu, telur-telur
yang tersisa dihitung.
Tipe tanggap fungsional diketahui dengan menggunakan regresi logistik.
Regresi logistik berasal dari proporsi mangsa yang diserang (Ne/No) sebagai
diuji sesuai pada fungsi polinom yang menggambarkan hubungan Ne/No dan No
sebagai berikut:
=
Keterangan: P0 = titik potong
P1 = koefisien linear
P2 = koefisien kuadratik
P3 = koefisien kubik
Pendugaan parameter (P) dilakukan dengan prosedur PROC CATMOD
SAS (SAS Institute 1989). Tanggap fungsional tipe II akan digambarkan dengan
nilai P1 yang lebih kecil dari 0 atau negatif (P1 < 0). Tanggap fungsional tipe III
akan ditunjukkan dengan nilai P1 yang positif (P1 > 0) namun P2 bernilai negatif
(P2 < 0). Pada tanggap fungsional tipe II dan III terdapat waktu penanganan
mangsa (Th) dan laju pencarian mangsa (a). Pendugaan Th dan a didapatkan dari
persamaan cakram Holing untuk tanggap fungsional tipe II dan persamaan Hassel
untuk tanggap fungsional tipe III (Hassel 1978).
Tanggap Numerik
Percobaan tanggap numerik dilakukan hampir sama dengan percobaan
tanggap fungsional, bertujuan untuk mengetahui hubungan antara peningkatan
populasi predator dengan tingkat kepadatan hama. Kepadatan telur yang
digunakan adalah 5, 10, 20, 40 dan 80 butir. Masing-masing perlakuan diulang 10
kali. Beberapa imago T. kanzawai dimasukkan dalam setiap arena percobaan dan
dibiarkan selama 24 jam. Telur-telur yang dihasilkan dihitung untuk digunakan
dalam percobaan tanggap numerik dan imago dipindahkan dari arena percobaan.
Predator yang digunakan adalah stadia imago yang berumur 2 hari. Setelah 24
jam, pengamatan dilakukan terhadap jumlah telur predator yang dihasilkan dalam
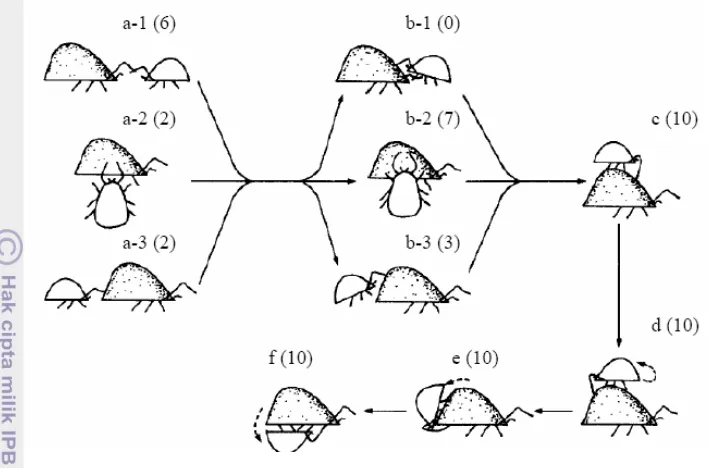
Kemampuan Menekan Populasi Mangsa oleh Individu Predator
Percobaan ini bertujuan mengetahui kemampuan individu tungau predator
menekan populasi mangsa dengan kepadatan mangsa yang berbeda. Kepadatan
imago mangsa yang digunakan sebagai berikut: 4, 8, 16 dan 32 ekor.
Imago-imago mangsa dibiarkan selama 24 jam. Setelah itu, seekor N. longispinosus
betina berumur 5-6 hari dimasukkan dalam arena percobaan yang telah berisi
mangsa. Pengamatan dilakukan setiap hari dengan mencatat jumlah semua stadia
predator dan mangsa yang ada dalam arena percobaan. Pengamatan berlangsung
selama 3 hari. Setiap perlakuan diulang sebanyak 5 kali. Perlakuan kontrol
(tanpa predator) hanya diulang sebanyak 3 kali dan pengamatan berlansung
selama 96 jam.
HASIL DAN PEMBAHASAN
Neraca Hayati
N. longispinosus memiliki nilai waktu generasi (T) sebesar 4.05 hari
dengan laju reproduksi bersih (Ro) sebesar 24.96 butir telur per generasi pada
tabel 1. Hal ini menunjukkan bahwa populasi N. longispinosus dapat berkembang
sebanyak 24.96 kali dalam satu generasi selama 4.05 hari. Nilai laju pertambahan
intrinsik rm adalah 0.44 betina/betina/hari dan laju pertambahan terbatas (λ)
sebesar 1.55 betina/betina/hari.
Nilai laju pertambahan intrinsik rm tungau predator N. longispinosus
adalah 0.44 betina/betina/hari. Nilai rm tersebut lebih tinggi dibandingkan rm
Tetranychidae. Nilai rm Tetranychidae memiliki kisaran 0.1 - 0.3 (Wrensch 1979;
Razmjou et al. 2009). Hal ini menunjukkan bahwa populasi tungau predator N.
longispinosus berkembang lebih cepat dibandingkan tungau Tetranychidae. Oleh
karena itu, N. longispinosus cenderung memiliki potensi tinggi sebagai musuh
alami dalam pengendalian tungau Tetranychidae. Nilai rm diperoleh dari
persamaan Σe-rx
lx mx = 1 (Carey 1993).
GRR dan Ro menunjukkan tingkat reproduksi N. longispinosus yang diberi
mangsa T. kanzawai. Nilai GRR adalah 32.78 individu, yang menunjukkan
rata-rata jumlah keturunan betina per generasi. Ro bernilai 24.96 individu,
menunjukkan jumlah keturunan betina yang berhasil menjadi imago. Nilai GRR
dan Ro N. longispinosus yang diberi mangsa T. kanzawai lebih tinggi
dibandingkan dengan mangsa lain (Puspitarini 2005; Thongtab et al 2002).
Jumlah keturunan betina yang relatif tinggi berimplikasi pada jumlah telur
yang dihasilkan dalam suatu populasi. Birch (1948) menyatakan bahwa makin
besar jumlah telur yang dihasilkan maka makin besar nilai laju pertambahan
intrinsik suatu organisme. Krebs (1978) menambahkan bahwa makin cepat
tercapainya puncak reproduksi maka makin besar nilai r suatu spesies. Nilai r
dipengaruhi oleh tiga faktor yaitu reproduksi pada umur muda, jumlah telur yang
dihasilkan setiap bertelur, dan meningkatnya ulangan peneluran yang berarti
Perkembangan populasi tungau predator N. longispinosus setiap hari (λ)
dapat dilihat pada tabel 1. λ sering juga disebut sebagai laju pertambahan terbatas, yang memiliki arti nilai perkembangan tungau predator pada lingkungan yang terbatas. Nilai λ = 1.55 per satuan hari, yang berarti bahwa perkembangan setiap individu pada setiap generasi menjadi satu individu pada keadaan lingkungan yang terbatas (Tarumingkeng 1992). Bila nilai λ > 1 maka terjadi pertambahan populasi tungau predator N. longispinosus setiap hari.
T dan DT merupakan parameter statistik demografi yang berkaitan dengan
waktu. Waktu yang dibutuhkan untuk menyelesaikan siklus hidup N.
longispinosus per generasi (T) adalah 4.05 hari. Nilai T yang hampir sama juga
ditunjukkan pada penelitian Puspitarini (2005) dan Thongtab et al. (2002).
Doubling time (DT) memiliki nilai sebesar 1.586 hari, yang mengisyaratkan
kemampuan N. longispinosus berkembang dalam satu generasi.
Tabel 1 Statistik demografi N. longispinosus
Parameter demografi Nilai Satuan
GRR 32.78 Individu
Ro 24.96 Individu/induk/generasi
rm 0.44 Individu/induk/hari
T 4.05 Hari
λ 1.55 Hari
DT 1.59 Hari
Pendugaan kelimpahan populasi suatu serangga merupakan implikasi
praktis dari sebaran umur stabil terutama serangga yang memiliki stadia
pradewasa yang tidak terlihat atau tersembunyi (Birch 1948). Selain itu, sebaran
umur stabil suatu populasi juga dapat digunakan untuk memperkirakan jumlah
pertambahan populasi.
Distribusi sebaran umur N. longispinosus pada percobaan ini menunjukkan
bahwa 83.78% populasi pradewasa dan sisanya 16.22% merupakan populasi
dewasa (tabel 2). Proporsi telur memiliki nilai paling tinggi di antara populasi
pradewasa. Nilai proporsi telur adalah 36.26 %. Deutonimfa memiliki nilai
Asumsi yang dihasilkan dari data distribusi sebaran umur N. longispinosus
menunjukkan bahwa sebagian besar populasi N. longispinosus didominasi oleh
populasi pradewasa. Implikasinya pada penarikan contoh di lapangan adalah
jumlah imago N. longispinosus di lapangan yang relatif rendah tidak
menggambarkan jumlah populasi N. longispinosus yang sebenarnya. Populasi
pradewasa juga mempengaruhi jumlah populasi yang sebenarnya. Populasi telur
N. longispinosus yang terkadang tersembunyi memiliki proporsi yang relatif besar
dalam menentukan keadaan populasi N. longispinosus. Percobaan ini dilakukan
dalam skala laboratorium maka faktor abiotik dan biotik yang mempengaruhi
[image:38.595.104.514.91.800.2]populasi dalam lingkungan yang alami tidak diperhitungkan.
Tabel 2 Proporsi berbagai fase perkembangan N. longispinosus pada persebaran umur stabil
Fase perkembangan (stadia) Proporsi (%)
Pradewasa
Telur
83.78
36.26
Larva 23.29
Protonimfa 14.86
Deutonimfa 9.37
Dewasa 16.22
Siklus Hidup dan Perkembangan
Siklus hidup N longispinosus terdiri dari telur, larva, protonimfa,
deutonimfa, dan dewasa yang ditunjukkan pada Tabel 3. N. longispinosus
meletakkan telur secara acak pada permukaan bawah daun ubi kayu. Telur
berbentuk oval dan berwarna putih transparan. Croft et al. (1999) menyatakan
bahwa ukuran telur 13 tungau Phytoseiidae yang diteliti, memiliki kisaran panjang
bervariasi dari 184.5–243.5 µm dan Phytoseiulus persimilis memiliki ukuran telur yang paling besar. Puspitarini (2005) menunjukkan bahwa telur N. longispinosus
yang dibiakkan pada Panonychus citri memiliki panjang berkisar 330-370 µm dan
longispinosus lebih besar dibandingkan ukuran telur P. persimilis. Telur
membutuhkan waktu perkembangan yang paling lama yaitu 1.40 hari.
Stadia larva merupakan stadia yang mudah diamati dan dibedakan dari
stadia lainnya. Ciri khas stadia larva adalah 3 pasang tungkai yang dimilikinya.
Pada stadia ini, larva berwarna putih dan masih bersifat pasif bergerak. Larva N.
longispinosus memiliki panjang rata-rata 230 µm dan lebar rata-rata 220 µm
(Puspitarini 2005). Larva sebagian besar tungau predator tidak makan sehingga
pada stadia ini tidak membutuhkan mangsa (Thongtab et al.2001; Zhang 2003).
Larva memiliki waktu perkembangan yang paling singkat yakni hanya 0.49 hari.
Waktu tersebut tidak berbeda nyata dengan penelitian Puspitarini (2005) dan
Thongtab et al. (2001).
Perubahan stadia larva menjadi protonimfa dapat dilihat pada jumlah
tungkai. Protonimfa memiliki 4 pasang tungkai dan berwarna putih keruh. Pada
stadia protonimfa, N. longispinosus mulai aktif untuk mencari mangsa.
Protonimfa N. longispinosus akan berubah warna setelah memakan mangsa,
menjadi merah kecoklatan. Masa protonimfa berlangsung selama 0.64 hari.
Setelah itu, N. longispinosus akan memasuki stadia deutonimfa. Lama masa
deutonimfa berkisar 0.70 hari. Masa protonimfa dan deutonimfa N. longispinosus
yang dibiakkan pada T. kanzawai memiliki waktu paling singkat dibandingkan N.
longispinosus yang dibiakkan pada P. citri (Puspitarini 2005) dan T. urticae
(Thongtab et al. 2001). Deutonimfa memiliki morfologi badan yang lebih besar
dan idiosoma yang pipih dibandingkan protonimfa. N. longispinosus bergerak
aktif mencari mangsa pada stadia deutonimfa. Protonimfa dan deutonimfa dapat
memangsa semua stadia mangsa yang terdapat dalam arena.
Tungau Phytoseiidae dewasa memiliki ukuran berkisar 200-490 µm,
Jarang sekali tungau Phytoseiidae dewasa memiliki ukuran lebih dari 500 µm.
Tungau dewasa N. longispinosus jantan dan betina memiliki ukuran yang berbeda.
N. longispinosus betina memiliki panjang dan lebar rata-rata berturut-turut: 400
µm dan 290 µm. Sementara panjang dan lebar rata-rata N. longispinosus jantan
lebih kecil yaitu 230 µm dan 170 µm (Puspitarini 2005). N. longispinosus
memiliki warna tubuh putih kekuningan keruh dan mengkilat serta seta-seta yang
mengikuti warna mangsanya. Ketika N. longispinosus diberikan mangsa imago T.
kanzawai yang berwarna merah, maka warna N. longispinosus akan berwarna
merah kecoklatan.
Waktu perkembangan N. longispinosus dari pradewasa menjadi dewasa
relatif singkat, yaitu 3.23 (Tabel 3). Pada umumnya tungau predator famili
Phytoseiidae memiliki waktu perkembangan yang relatif lebih cepat dan singkat
dari Tetranychidae (Zhang 2003). Penelitian Puspitarini (2005) dan Thongtab et
al. (2001) menunjukkan bahwa lama perkembangan N. longispinosus dari telur
hingga menjadi dewasa adalah 4.78 hari dan 4.23 hari. Waktu perkembangan N.
longispinosus yang dibiakkan pada T. kanzawai lebih cepat. Hal ini dapat
disebabkan oleh pengalaman predator terhadap mangsa karena selama pembiakan
N. longispinosus diberikan mangsa T. kanzawai.
Tabel 3 Berbagai parameter kehidupan N. longispinosus yang dibiakkan pada
T.kanzawai
Parameter Durasi (hari) (mean ± SD)a
Lama perkembangan pradewasa 3.23 ± 0.41
Telur 1.40 ± 0.32
Larva 0.49 ± 0.01
Protonimfa
Deutonimfa
0.64 ± 0.21
0.70 ± 0.31
Lama hidup betina 16.78 ± 7.44
Periode praoviposisi
Periode oviposisi
Periode pascaoviposisi
1.55 ± 0.57
8.76 ± 4.68
1.92 ± 2.91
Lama hidup jantan 22.28 ± 7.02
Keperidian (butir/betina) 32.78 ± 1.31
Nisbah kelamin (♂:♀) 1: 2.13
Tabel 3 juga memberikan informasi berbagai parameter kehidupan tungau
dewasa N. longispinosus. Dewasa N. longispinosus betina memiliki masa
[image:40.595.99.515.117.782.2]lebih lama dibandingkan masa praoviposisi dan pascaoviposisi. Masa oviposisi N.
longispinosus yang dibiakkan pada T. kanzawai adalah 8.76 hari. Masa oviposisi
tersebut lebih singkat dibandingkan dengan masa oviposisi N. longispinosus yang
dibiakkan pada tungau merah jeruk P. citri (Puspitarini 2005) dan Eotetranychus
cendanai (Thongtab et al. 2001). Namun, keperidian yang diperoleh lebih tinggi
dibandingkan penelitian Puspitarini (2005) dan Thongtab et al. (2001).
Keperidian N. longispinosus yang dibiakkan pada T. kanzawai adalah 32.78 butir.
Hasil penelitian menunjukkan bahwa rata-rata jumlah telur yang diletakkan per
hari adalah 3.39 butir. Angka tersebut relatif tinggi dibandingkan penelitian N.
longispinosus yang dilakukan oleh Puspitarini (2005) dan Yulianah (2008).
Jumlah telur rata-rata N. longispinosus yang dibiakkan pada T. kanzawai memiliki
nilai yang hampir sama dengan N. longispinosus yang dibiakkan pada T. urticae
yaitu 3.36 butir (Kongchuensin et al. 1989 dalam Thongtab et al. 2001). Hal
tersebut menunjukkan bahwa T. kanzawai merupakan mangsa yang sesuai bagi
perkembangan N. longsipinosus.
Lama hidup tungau jantan lebih lama dibandingkan lama hidup tungau
betina. Tungau jantan memiliki lama hidup yaitu 22.28 hari sedangkan lama
hidup tungau betina hanya berkisar 16.78 hari. Nisbah kelamin N. longispinosus
yang dibiakkan pada T. kanzawai adalah 1:2.13. Penelitian Escudero & Ferragut
(2005) menunjukkan nisbah kelamin 1:2 untuk P. persimilis yang diberikan
mangsa T. urticae, T. turkestani, dan T. ludeni.
Sintasan dan Jumlah Telur Predator Harian
Sintasan menunjukkan tingkat keberhasilan hidup dari suatu populasi
dalam bentuk persen. Sintasan tungau predator N. longispinosus dari
pengamatan harian ditunjukkan pada Gambar 9. Kurva sintasan tungau predator
N. longispinosus memperlihatkan bahwa mortalitas mulai terjadi pada saat tungau
predator N. longispinosus berumur 2 hari. Penurunan sintasan terus terjadi sampai
semua tungau predator betina mati yang terjadi pada saat tungau predator berumur
32 hari. Kurva sintasan tungau predator N. longispinosus termasuk dalam kurva
sintasan tipe I (Tarumingkeng 1992). Kurva sintasan tipe I menunjukkan bahwa
tungau predator N. longispinosus berumur tua. Penelitian Puspitarini (2005) juga
menunjukkan kurva sintasan tipe 1 untuk N. longispinosus yang diberikan mangsa
P. citri. Namun tingkat mortalitas pada N. longispinosus yang diberikan mangsa
P. citri terjadi pada umur 4 hari dan lama hidup yang lebih singkat yaitu 27 hari
(Puspitarini 2005). Perbedaan tersebut dapat disebabkan oleh mangsa yang
diberikan pada N. longispinosus.
Gambar 10 Kurva sintasan (lx) dan rata-rata jumlah telur harian (mx) tungau
predator N. longispinosus
Selain informasi sintasan tungau predator N. longispinosus, gambar 9 juga
menunjukkan informasi tentang rata-rata produksi telur per hari yang diletakkan
oleh individu betina yang hidup pada umur tertentu (mx). Kurva mx
memperlihatkan bahwa tungau predator N. longispinosus mulai mengalami
oviposisi saat berumur 3 hari. Penelitian Thongtab et al. (2001) juga menyatakan
masa oviposisi N. longispinosus yang diberikan E. cendanai yaitu 3.57 hari.
Produksi telur meningkat pada awal umur predator dan menurun seiring
pertambahan umur predator. Tingkat produksi telur tertinggi dicapai saat tungau
predator betina N. longispinosus berumur 6 hari dengan rata-rata produksi telur 4
butir. Gambar 9 juga memberikan informasi tentang masa praoviposisi, oviposisi,
dan pascaoviposisi. Masa praoviposisi relatif singkat yaitu 1.55 hari.
N.longispinosus memasuki stadia dewasa saat hari ke-3. Pada saat tungau
predator betina N. longispinosus berumur 4 hari mulai terlihat peningkatan
produksi telur sehingga ini merupakan awal masa oviposisi N. longispinosus.
Masa oviposisi berlangsung selama 21 hari, berawal saat N. longispinosus 0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 0.00 0.20 0.40 0.60 0.80 1.00 1.20
0 3 6 9 12 15 18 21 24 27 30 33 Rat
berumur 4 hari hingga berumur 24 hari. Rata-rata jumlah telur harian tertinggi
terjadi pada tungau predator N. longispinosus berumur 6 hari. Oleh karena itu,
puncak mx terjadi pada tungau predator betina N. longispinosus umur 6 hari.
Kurva mx mengalami fluktuasi setelah tungau predator N. longispinosus berumur
6 hari. Pada saat berumur 24 hari, N. longispinosus mulai memasuki masa
pascaoviposisi. Masa pascaoviposisi terjadi saat tungau predator N. longispinosus
berumur 25 hari. Hal ini mengindikasikan bahwa N. longispinosus memiliki
potensi relatif besar sebagai agen pengendali tungau Tetranychidae karena
peletakkan telur terjadi pada awal sehingga cukup menguntungkan dari segi
pengendalian tungau Tetranychidae.
Preferensi Stadia Mangsa
Preferensi N. longispinosus terhadap mangsa (telur, nimfa, dan imago T.
kanzawai) berkaitan dengan perilaku pencarian dan pengenalan mangsa oleh
predator. Perilaku N. longispinosus tersebut didukung oleh suatu stimuli dari
mangsa. Stimuli tersebut dapat berupa gerakan atau tanda yang ditinggalkan oleh
mangsa. Keberadaan stimuli yang cukup tinggi dapat meningkatkan peluang
predator untuk menemukan mangsa (Dickens 1999).
Preferensi N. longispinosus terhadap mangsa tertentu akan meningkatkan
perilaku pemangsaan yang tinggi. Stadia dan jenis mangsa mempengaruhi
perilaku pemangsaan predator terhadap mangsa (Gullan & Cranston 1994).
Preferensi keberadaan imago N. longispinosus pada arena percobaan
selama 3 jam ditunjukkan pada gambar 11. Sebagian besar imago N.
longispinosus lebih banyak dijumpai pada daun yang berisi telur tungau T.
kanzawai. Hal ini menunjukkan bahwa N. longispinosus memiliki ketertarikan
yang cukup tinggi terhadap mangsa dalam stadia telur. Bahkan mangsa dalam
stadia imago tidak dikunjungi oleh N. longispinosus pada waktu pengamatan ke-2,
3, dan 6. Jenis, stadia dan ukuran tubuh mangsa yang bervariasi memiliki
pengaruh terhadap preferensi predator (Yang & Li 2002). Semakin besar instar
nimfa mangsa maka semakin kecil preferensi terhadap mangsa. Penyebab
penurunan preferensi predator tersebut mungkin dikarenakan bentuk dan ukuran
untuk memangsa. Mekanisme pertahanan mangsa dalam bentuk karakteristik
mangsa merupakan bagian dari proses interaksi predator dan mangsa (Holling
1961).
Gambar 11 Keberadaan imago N. longispinosus selama 3 jam
Tanggap Fungsional
Hasil analisis regresi logistik predasi N. longispinosus pada mangsa telur
Tetranychus kanzawai diperlihatkan pada tabel 4. Hubungan kepadatan mangsa
dan tingkat predasi memperlihatkan tanggap fungsional tipe III. Hal tersebut
ditunjukkan oleh hasil analisis regresi logistik. Tanggap fungsional tipe III
ditunjukkan oleh koefisien linear yang bernilai positif. Hal ini menunjukkan
bahwa pada awalnya proporsi mangsa yang dipredasi meningkat kemudian
menurun seiring dengan bertambahnya kepadatan mangsa (Sabelis 1985).
Tabel 4 Hasil analisis regresi logistik proporsi mangsa T. kanzawai yang dimangsa N. longispinosus
Parameter Nilai Penduga Standar Error Pa
Titik potong -6.0075 430.0 0.9889
Linear 2.0273 129.0 0.9875
Kuadratik -0.1347 8.5996 0.9875
a P=probability 0 1 2 3 4 5 6 7 8 9 10
1 2 3 4 5 6
Ju m lah i mago N . lo n g is p ino su s
Waktu pengamatan (per 30 menit)
Telur
Nimfa
Tanggap fungsional tipe III merupakan satu-satunya tipe tanggap
fungsional yang memiliki kontribusi nyata dalam regulasi populasi hama (Hassel
1976; Hassel 1978). Fernandez & Corley (2003) menambahkan bahwa predator
atau parasitoid yang memiliki tanggap fungsional tipe III berpotensi sebagai agen
biokontrol yang efisien.
Tipe tanggap fungsional ditentukan oleh perilaku memburu dan menangani
mangsa. Laju pencarian mangsa akan meningkat pada kepadatan mangsa yang
tinggi hingga mencapai kejenuhan. Penambahan populasi mangsa akan
menurunkan laju pencarian predator. Laju pemangsaan diduga berkaitan dengan
waktu penanganan mangsa (mengenal dan memburu) dimana preferensi predator
akan menentukan laju pencarian mangsa (Tarumingkeng 1992).
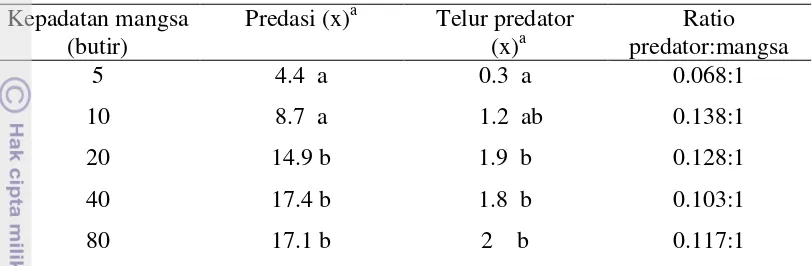
Waktu penanganan mangsa (Th) merupakan salah satu karakter penting
dalam interaksi mangsa-predator. Waktu penanganan ini meliputi mengenal,
memburu dan menangani mangsa. Kepadatan populasi mangsa sangat berkaitan
dengan waktu penanganan mangsa. Hal ini dapat dilihat pada tabel 5. Kepadatan
populasi mangsa yang rendah membutuhkan waktu yang cukup panjang.
Kepadatan mangsa sebanyak 5 butir telur membutuhkan waktu yang paling lama
yaitu 4.06 jam. Sebaliknya pada kepadatan populasi mangsa yang tinggi, predator
membutuhkan waktu yang relatif pendek. Waktu penanganan mangsa paling
pendek yaitu 1.21 jam ditunjukkan pada kepadatan mangsa 40 butir telur.