PENGARUH EMPAT STRAIN BAKTERI PERAKARAN
PEMACU PERTUMBUHAN TANAMAN DAN WAKTU
INOKULASI VIRUS TERHADAP PERTUMBUHAN
TANAMAN SERTA KEPARAHAN PENYAKIT KUNING
PADA TANAMAN CABAI
HARI PRIWIRATAMA A44102014
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
ABSTRAK
HARI PRIWIRATAMA. Pengaruh Empat Strain Bakteri Perakaran Pemacu Pertumbuhan Tanaman dan Waktu Inokulasi Virus terhadap Pertumbuhan Tanaman serta Keparahan Penyakit Kuning pada Tanaman Cabai. Dibimbing oleh SRI HENDRASTUTI HIDAYAT dan WIDODO.
Bakteri perakaran pemacu pertumbuhan tanaman (plant growth promoting rhizobacteria [PGPR]) telah menunjukkan efektivitas dalam menekan serangan patogen. Bacillus dan Pseudomonas mampu menekan keparahan penyakit yang disebabkan oleh cucumber mosaic virus (CMV), tobacco mosaic virus (TMV), tobacco necrosis virus (TNV) dan tomato mottle virus (ToMoV). Efektivitasnya dalam menekan penyakit kuning yang disebabkan oleh geminivirus pada tanaman cabai belum banyak diketahui.
Empat strain PGPR Bacillus polymixa BG25, B. subtilis SB3 , Pseudomonas fluorescens PG01 dan P. fluorescens ES32 serta campuran suspensi keempat bakteri diaplikasikan melalui perendaman benih. Inokulasi geminivirus dilakukan di rumah kaca pada 3, 5 dan 6 minggu setelah tanam dengan bantuan vektor Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae). Tanaman tanpa perlakuan perendaman benih dalam suspensi bakteri digunakan sebagai pembanding.
PENGARUH EMPAT STRAIN BAKTERI PERAKARAN
PEMACU PERTUMBUHAN TANAMAN DAN WAKTU
INOKULASI VIRUS TERHADAP PERTUMBUHAN
TANAMAN SERTA KEPARAHAN PENYAKIT KUNING
PADA TANAMAN CABAI
HARI PRIWIRATAMA A44102014
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian Institut Pertanian Bogor
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
Judul Skripsi : Pengaruh Empat Strain Bakteri Perakaran Pemacu Pertumbuhan Tanaman dan Waktu Inokulasi Virus terhadap Pertumbuhan Tanaman serta Keparahan Penyakit Kuning pada Tanaman Cabai
Nama Mahasiswa : Hari Priwiratama NRP : A44102014
Program Studi : Hama dan Penyakit Tumbuhan
Menyetujui
Pembimbing I
Dr. Ir. Sri Hendrastuti Hidayat, MSc. NIP 131 610 286
Pembimbing II
Dr. Ir. Widodo, MS. NIP 131 476 605
Mengetahui
Dekan Fakultas Pertanian
Prof. Dr. Ir. Supiandi Sabiham, MAgr. NIP 130 422 698
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta, Jawa Barat pada tanggal 2 Januari 1985 sebagai anak kedua dari tiga bersaudara dari pasangan Bapak Djadja ABD Rodjak dan Ibu Mimin Djarnudji.
Penulis memperoleh pendidikan sekolah lanjutan tingkat atas di SMU Negeri 6 Bogor dan menyelesaikannya pada tahun 2002. Pada tahun yang sama penulis diterima di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor melalui Undangan Seleksi Mahasiswa IPB.
Aktifitas kemahasiswaan yang pernah diikuti penulis yaitu menjadi anggota Biro Infokom Himpunan Mahasiswa Proteksi Tanaman (HIMASITA) tahun 2002-2003 dan kepala Departemen Keprofesian HIMASITA pada tahun 2004-2005. Penulis juga pernah menjadi asisten dosen pada mata kuliah Dasar-dasar Perlindungan Tanaman, Nematologi Tumbuhan, Metode Statistika I (tahun ajaran 2004/2005), Ilmu Penyakit Tumbuhan Umum serta Hama dan Penyakit Tanaman Pangan (tahun ajaran 2005/2006).
PRAKATA
Puji dan syukur ke hadirat Illahi Rabbi yang telah memberikan rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan penyusunan skripsi sebagai syarat memperoleh gelar Sarjana Pertanian pada Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Skripsi penulis berjudul Pengaruh Empat Strain Bakteri Perakaran Pemacu Pertumbuhan Tanaman dan Waktu Inokulasi Virus terhadap Pertumbuhan Tanaman serta Keparahan Penyakit Kuning pada Tanaman Cabai dilaksanakan di Rumah Kaca Kebun Percobaan Cikabayan, Laboratorium Mikologi dan Klinik Tanaman Departemen Proteksi Tanaman mulai bulan Januari 2006 hingga Juni 2006. Penelitian ini merupakan bagian dari penelitian dosen pembimbing yang didanai melalui Hibah Penelitian Program Hibah Kompetisi-B (PHK-B) Departemen Proteksi Tanaman, Institut Pertanian Bogor.
Pada kesempatan ini penulis menyampaikan terima kasih kepada: 1. Dr. Ir. Sri Hendrastuti Hidayat, MSc. sebagai pembimbing I tugas akhir. 2. Dr. Ir. Widodo, MS. sebagai pembimbing II tugas akhir.
3. Dr. Ir. Kikin H Mutakin, MSc. yang telah membantu dalam analisis data. 4. Kedua orang tua untuk doa dan dukungannya.
5. Mia Saumiati atas waktu, tenaga dan dukungan yang telah diberikan.
6. Romadhona, Hendrayana, Ari, Lusi, Astri dan Sri Endang yang telah membantu dalam pelaksanaan penelitian penulis.
Penulis berharap hasil penelitian ini dapat bermanfaat sebagai acuan dalam melaksanakan penelitian yang berkaitan.
Bogor, Agustus 2006
DAFTAR ISI
PGPR dalam Teknik Pengendalian Penyakit Tanaman ... 6
Bacillus spp. dan Pseudomonas spp. sebagai Agensi Hayati Pengendalian Virus tanaman ... 7
Induksi Ketahanan Sistemik oleh PGPR ... 9
BAHAN DAN METODE
Perbanyakan Serangga Vektor Bebas Virus ... 11
Pemurnian dan Perbanyakan Isolat Virus ... 11
vii
Halaman HASIL DAN PEMBAHASAN
Pengaruh PGPR dan Waktu Inokulasi Terhadap Morfologi Tanaman Pengaruh PGPR terhadap Perkembangan Tinggi Tanaman, Jumlah
Cabang, Daun dan Bunga ... 14
Pengaruh Waktu Inokulasi Virus terhadap Perkembangan Tinggi Tanaman, Jumlah Cabang, Daun dan Bunga ... 16
Pengaruh Interaksi antara Perlakuan Bakteri dan Waktu Inokulasi Virus terhadap Perkembangan Tinggi Tanaman, Jumlah Cabang, Daun dan Bunga ... 18
Pengaruh PGPR dan Waktu Inokulasi Virus terhadap Perkembangan Penyakit Kuning Pengaruh PGPR terhadap Masa Inkubasi dan Perkembangan Keparahan Penyakit ... 23
Pengaruh Waktu Inokulasi terhadap Masa Inkubasi dan Perkembangan Keparahan Penyakit ... 26
Pengaruh Interaksi Perlakuan Bakteri dan Waktu Inokulasi terhadap Masa Inkubasi dan Tingkat Keparahan Penyakit ... 28
KESIMPULAN DAN SARAN Kesimpulan ... 31
Saran ... 31
DAFTAR PUSTAKA ... 32
DAFTAR TABEL
Halaman Tabel 1 Pengaruh Perlakuan Bakteri terhadap Tinggi Tanaman Cabai
pada Beberapa Waktu Pengamatan ... 14 Tabel 2 Pengaruh Perlakuan Bakteri terhadap Jumlah Cabang Utama,
Cabang Sekunder, Daun dan Bunga ... 15 Tabel 3 Pengaruh Waktu Inokulasi terhadap Jumlah Cabang Utama,
Cabang Sekunder, Daun dan Bunga ... 18 Tabel 4 Pengaruh Interaksi antara Perlakuan Bakteri dan Waktu
Inokulasi Virus terhadap Perkembangan Tinggi Tanaman
Cabai ... 19 Tabel 4 (lanjutan) Pengaruh Interaksi antara Perlakuan Bakteri dan
Waktu Inokulasi Virus terhadap Perkembangan Tinggi
Tanaman Cabai ... 20 Tabel 5 Pengaruh Interaksi Perlakuan Bakteri dan Waktu Inokulasi
Virus Terhadap Jumlah Cabang Utama, Cabang Sekunder,
Daun dan Bunga ... 22 Tabel 6 Pengaruh Perlakuan Bakteri terhadap Keparahan Penyakit
Kuning ... 25 Tabel 7 Pengaruh Waktu Inokulasi Virus Terhadap Keparahan
Penyakit ... 27 Tabel 8 Pengaruh Interaksi Perlakuan Bakteri dan Waktu Inokulasi
DAFTAR GAMBAR
Halaman Gambar 1 Pengaruh Waktu Inokulasi Virus terhadap Perkembangan
Tinggi Tanaman ... 17 Gambar 2 Pengaruh Perlakuan Bakteri terhadap Masa Inkubasi
Penyakit ... 24 Gambar 3 Pengaruh Waktu Inokulasi Virus terhadap Masa Inkubasi
Penyakit ... 26 Gambar 4 Pengaruh Interaksi antara Perlakuan Bakteri dan Waktu
PENGARUH EMPAT STRAIN BAKTERI PERAKARAN
PEMACU PERTUMBUHAN TANAMAN DAN WAKTU
INOKULASI VIRUS TERHADAP PERTUMBUHAN
TANAMAN SERTA KEPARAHAN PENYAKIT KUNING
PADA TANAMAN CABAI
HARI PRIWIRATAMA A44102014
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
ABSTRAK
HARI PRIWIRATAMA. Pengaruh Empat Strain Bakteri Perakaran Pemacu Pertumbuhan Tanaman dan Waktu Inokulasi Virus terhadap Pertumbuhan Tanaman serta Keparahan Penyakit Kuning pada Tanaman Cabai. Dibimbing oleh SRI HENDRASTUTI HIDAYAT dan WIDODO.
Bakteri perakaran pemacu pertumbuhan tanaman (plant growth promoting rhizobacteria [PGPR]) telah menunjukkan efektivitas dalam menekan serangan patogen. Bacillus dan Pseudomonas mampu menekan keparahan penyakit yang disebabkan oleh cucumber mosaic virus (CMV), tobacco mosaic virus (TMV), tobacco necrosis virus (TNV) dan tomato mottle virus (ToMoV). Efektivitasnya dalam menekan penyakit kuning yang disebabkan oleh geminivirus pada tanaman cabai belum banyak diketahui.
Empat strain PGPR Bacillus polymixa BG25, B. subtilis SB3 , Pseudomonas fluorescens PG01 dan P. fluorescens ES32 serta campuran suspensi keempat bakteri diaplikasikan melalui perendaman benih. Inokulasi geminivirus dilakukan di rumah kaca pada 3, 5 dan 6 minggu setelah tanam dengan bantuan vektor Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae). Tanaman tanpa perlakuan perendaman benih dalam suspensi bakteri digunakan sebagai pembanding.
PENGARUH EMPAT STRAIN BAKTERI PERAKARAN
PEMACU PERTUMBUHAN TANAMAN DAN WAKTU
INOKULASI VIRUS TERHADAP PERTUMBUHAN
TANAMAN SERTA KEPARAHAN PENYAKIT KUNING
PADA TANAMAN CABAI
HARI PRIWIRATAMA A44102014
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian Institut Pertanian Bogor
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
Judul Skripsi : Pengaruh Empat Strain Bakteri Perakaran Pemacu Pertumbuhan Tanaman dan Waktu Inokulasi Virus terhadap Pertumbuhan Tanaman serta Keparahan Penyakit Kuning pada Tanaman Cabai
Nama Mahasiswa : Hari Priwiratama NRP : A44102014
Program Studi : Hama dan Penyakit Tumbuhan
Menyetujui
Pembimbing I
Dr. Ir. Sri Hendrastuti Hidayat, MSc. NIP 131 610 286
Pembimbing II
Dr. Ir. Widodo, MS. NIP 131 476 605
Mengetahui
Dekan Fakultas Pertanian
Prof. Dr. Ir. Supiandi Sabiham, MAgr. NIP 130 422 698
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta, Jawa Barat pada tanggal 2 Januari 1985 sebagai anak kedua dari tiga bersaudara dari pasangan Bapak Djadja ABD Rodjak dan Ibu Mimin Djarnudji.
Penulis memperoleh pendidikan sekolah lanjutan tingkat atas di SMU Negeri 6 Bogor dan menyelesaikannya pada tahun 2002. Pada tahun yang sama penulis diterima di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor melalui Undangan Seleksi Mahasiswa IPB.
Aktifitas kemahasiswaan yang pernah diikuti penulis yaitu menjadi anggota Biro Infokom Himpunan Mahasiswa Proteksi Tanaman (HIMASITA) tahun 2002-2003 dan kepala Departemen Keprofesian HIMASITA pada tahun 2004-2005. Penulis juga pernah menjadi asisten dosen pada mata kuliah Dasar-dasar Perlindungan Tanaman, Nematologi Tumbuhan, Metode Statistika I (tahun ajaran 2004/2005), Ilmu Penyakit Tumbuhan Umum serta Hama dan Penyakit Tanaman Pangan (tahun ajaran 2005/2006).
PRAKATA
Puji dan syukur ke hadirat Illahi Rabbi yang telah memberikan rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan penyusunan skripsi sebagai syarat memperoleh gelar Sarjana Pertanian pada Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Skripsi penulis berjudul Pengaruh Empat Strain Bakteri Perakaran Pemacu Pertumbuhan Tanaman dan Waktu Inokulasi Virus terhadap Pertumbuhan Tanaman serta Keparahan Penyakit Kuning pada Tanaman Cabai dilaksanakan di Rumah Kaca Kebun Percobaan Cikabayan, Laboratorium Mikologi dan Klinik Tanaman Departemen Proteksi Tanaman mulai bulan Januari 2006 hingga Juni 2006. Penelitian ini merupakan bagian dari penelitian dosen pembimbing yang didanai melalui Hibah Penelitian Program Hibah Kompetisi-B (PHK-B) Departemen Proteksi Tanaman, Institut Pertanian Bogor.
Pada kesempatan ini penulis menyampaikan terima kasih kepada: 1. Dr. Ir. Sri Hendrastuti Hidayat, MSc. sebagai pembimbing I tugas akhir. 2. Dr. Ir. Widodo, MS. sebagai pembimbing II tugas akhir.
3. Dr. Ir. Kikin H Mutakin, MSc. yang telah membantu dalam analisis data. 4. Kedua orang tua untuk doa dan dukungannya.
5. Mia Saumiati atas waktu, tenaga dan dukungan yang telah diberikan.
6. Romadhona, Hendrayana, Ari, Lusi, Astri dan Sri Endang yang telah membantu dalam pelaksanaan penelitian penulis.
Penulis berharap hasil penelitian ini dapat bermanfaat sebagai acuan dalam melaksanakan penelitian yang berkaitan.
Bogor, Agustus 2006
DAFTAR ISI
PGPR dalam Teknik Pengendalian Penyakit Tanaman ... 6
Bacillus spp. dan Pseudomonas spp. sebagai Agensi Hayati Pengendalian Virus tanaman ... 7
Induksi Ketahanan Sistemik oleh PGPR ... 9
BAHAN DAN METODE
Perbanyakan Serangga Vektor Bebas Virus ... 11
Pemurnian dan Perbanyakan Isolat Virus ... 11
vii
Halaman HASIL DAN PEMBAHASAN
Pengaruh PGPR dan Waktu Inokulasi Terhadap Morfologi Tanaman Pengaruh PGPR terhadap Perkembangan Tinggi Tanaman, Jumlah
Cabang, Daun dan Bunga ... 14
Pengaruh Waktu Inokulasi Virus terhadap Perkembangan Tinggi Tanaman, Jumlah Cabang, Daun dan Bunga ... 16
Pengaruh Interaksi antara Perlakuan Bakteri dan Waktu Inokulasi Virus terhadap Perkembangan Tinggi Tanaman, Jumlah Cabang, Daun dan Bunga ... 18
Pengaruh PGPR dan Waktu Inokulasi Virus terhadap Perkembangan Penyakit Kuning Pengaruh PGPR terhadap Masa Inkubasi dan Perkembangan Keparahan Penyakit ... 23
Pengaruh Waktu Inokulasi terhadap Masa Inkubasi dan Perkembangan Keparahan Penyakit ... 26
Pengaruh Interaksi Perlakuan Bakteri dan Waktu Inokulasi terhadap Masa Inkubasi dan Tingkat Keparahan Penyakit ... 28
KESIMPULAN DAN SARAN Kesimpulan ... 31
Saran ... 31
DAFTAR PUSTAKA ... 32
DAFTAR TABEL
Halaman Tabel 1 Pengaruh Perlakuan Bakteri terhadap Tinggi Tanaman Cabai
pada Beberapa Waktu Pengamatan ... 14 Tabel 2 Pengaruh Perlakuan Bakteri terhadap Jumlah Cabang Utama,
Cabang Sekunder, Daun dan Bunga ... 15 Tabel 3 Pengaruh Waktu Inokulasi terhadap Jumlah Cabang Utama,
Cabang Sekunder, Daun dan Bunga ... 18 Tabel 4 Pengaruh Interaksi antara Perlakuan Bakteri dan Waktu
Inokulasi Virus terhadap Perkembangan Tinggi Tanaman
Cabai ... 19 Tabel 4 (lanjutan) Pengaruh Interaksi antara Perlakuan Bakteri dan
Waktu Inokulasi Virus terhadap Perkembangan Tinggi
Tanaman Cabai ... 20 Tabel 5 Pengaruh Interaksi Perlakuan Bakteri dan Waktu Inokulasi
Virus Terhadap Jumlah Cabang Utama, Cabang Sekunder,
Daun dan Bunga ... 22 Tabel 6 Pengaruh Perlakuan Bakteri terhadap Keparahan Penyakit
Kuning ... 25 Tabel 7 Pengaruh Waktu Inokulasi Virus Terhadap Keparahan
Penyakit ... 27 Tabel 8 Pengaruh Interaksi Perlakuan Bakteri dan Waktu Inokulasi
DAFTAR GAMBAR
Halaman Gambar 1 Pengaruh Waktu Inokulasi Virus terhadap Perkembangan
Tinggi Tanaman ... 17 Gambar 2 Pengaruh Perlakuan Bakteri terhadap Masa Inkubasi
Penyakit ... 24 Gambar 3 Pengaruh Waktu Inokulasi Virus terhadap Masa Inkubasi
Penyakit ... 26 Gambar 4 Pengaruh Interaksi antara Perlakuan Bakteri dan Waktu
DAFTAR LAMPIRAN
Halaman Lampiran 1 Gambar Gejala Tanaman yang Terinfeksi Geminivirus
dengan Berbagai Tingkat Keparahan ... 36 Lampiran 2 Tabel Pengaruh Perlakuan Bakteri terhadap Perkembangan
Tinggi Tanaman Cabai ... 37 Lampiran 3 Tabel Pengaruh Waktu Inokulasi terhadap Perkembangan
Tinggi Tanaman Cabai ... 37 Lampiran 4 Tabel Pengaruh Perlakuan Bakteri terhadap Masa
Inkubasi Penyakit ... 38 Lampiran 5 Tabel Pengaruh Waktu Inokulasi Virus terhadap Masa
Inkubasi Penyakit ... 38 Lampiran 6 Tabel Pengaruh Perlakuan Bakteri dan Waktu Inokulasi
PENDAHULUAN
Latar Belakang
Penyakit kuning yang disebabkan geminivirus merupakan penyakit penting pada tanaman cabai di Indonesia. Penyakit ini mampu menyerang setiap tahap perkembangan tanaman cabai, sejak pesemaian hingga masa pembuahan. Serangan akan lebih merugikan jika terjadi pada fase vegetatif muda karena dapat menyebabkan penurunan kemampuan berbuah sedangkan serangan pada fase generatif menyebabkan penurunan kualitas buah (Sukamto 2005).
Tanaman cabai yang terinfeksi geminivirus akan menunjukkan gejala berupa penguningan disertai penggulungan terutama pada daun muda. Serangan geminivirus juga menyebabkan penghambatan pertumbuhan tanaman. Infestasi geminivirus pada tanaman dapat menyebabkan penurunan hasil. Ahmed et al. (2001) melaporkan bahwa kehilangan hasil karena penyakit ini dapat mencapai 100% di Sudan, sedangkan di Gezira kehilangan hasil mencapai 70%. Di Indonesia kehilangan hasil berkisar 20% sampai 100% serta menyebabkan kerugian hingga Rp. 7,31 milyar dengan luas serangan mencapai 984,6 hektar (Anonim 2004; Sukamto 2005).
Geminivirus tidak dapat ditularkan secara mekanis, tetapi hanya dapat ditularkan dengan bantuan serangga vektor sehingga persebarannya di lapangan bergantung pada kemampuan pemencaran vektornya. Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae) merupakan vektor yang efektif dalam menularkan virus ini. Persentase infeksi melalui penularan oleh B. tabaci mencapai 80% pada varietas cabai besar Capsicum annum (L) (Rusli et al. 1999). Walaupun bersifat persisten, tetapi geminivirus tidak terbawa pada keturunan vektor yang berikutnya. Virus juga tidak terbawa benih sehingga benih yang dihasilkan dari tanaman sakit tidak secara langsung terinfeksi virus dan masih aman untuk digunakan (Hull 2002).
2
varietas tahan dapat mengurangi kejadian penyakit di lapangan secara efektif, tetapi pengembangan varietas tanaman ini relatif lama dan dinilai tidak ekonomis (Hadidi et al. 1998). Disamping itu, munculnya strain virus baru dengan tingkat virulensi yang lebih tinggi menyebabkan efektivitas penggunaan varietas tahan ini semakin menurun.
Pengendalian secara tidak langsung dapat dilakukan melalui pengelolaan vektor virus. Hal ini didasarkan pada kenyataan bahwa geminivirus hanya dapat ditularkan oleh serangga vektor. Melalui penggunan insektisida, populasi vektor dapat ditekan sejak awal budidaya sehingga mencegah penularan penyakit secara luas. Ahmed et al. (2001) memaparkan bahwa penggunaan insektisida dapat menekan kejadian penyakit geminivirus 2,2% hingga 17%. Walaupun demikian, penggunaannya dapat menimbulkan dampak negatif terhadap lingkungan, diantaranya resistensi pada serangga vektor serta matinya musuh alami.
Usaha pengendalian penyakit yang mulai dikembangkan dan relatif aman terhadap lingkungan yaitu penggunaan Bakteri Perakaran Pemacu Pertumbuhan Tanaman (Plant Growth Promoting Rhizobacteria [PGPR]). Usaha ini sering disebut dengan bakterisasi, yaitu perlakuan benih atau akar perkecambahan dengan suspensi bakteri sehingga dapat memperbaiki pertumbuhan tanaman (Baker 1974 dalam Widodo 1993). Selain memperbaiki pertumbuhan, PGPR juga berperan dalam pengendalian penyakit tanaman.
3
Tujuan
Penelitian ini bertujuan untuk mengetahui pengaruh penggunaan PGPR serta variasi waktu inokulasi virus terhadap pertumbuhan tanaman dan perkembangan penyakit kuning pada tanaman cabai besar.
Manfaat
Hasil penelitian ini diharapkan dapat menjadi masukan dalam pengendalian penyakit kuning bagi petani dan pihak yang terkait serta dapat menjadi inspirasi bagi penelitian-penelitian selanjutnya.
Hipotesis
TINJAUAN PUSTAKA
Kemaknaan Geminivirus pada Tanaman Cabai
Geminivirus termasuk kategori lima belas jenis organisme pengganggu tanaman utama yang menyerang tanaman cabai di lapangan. Penyakit yang disebabkannya dikenal dengan sebutan penyakit kuning karena dapat menyebabkan daun-daun tanaman menjadi kuning. Serangan virus juga menyebabkan pemucatan pada tulang daun utama disertai penggulungan daun (cupping) dan kekerdilan pada tanaman sehingga tanaman tidak dapat berproduksi (Brown 2003).
Serangan virus ini dapat menyebabkan kehilangan hasil yang sangat tinggi, berkisar antara 20% dan 100% (Brown & Bird 1992). Di Indonesia, serangan geminivirus menyebabkan tanaman cabai menjadi kerdil dan tidak menghasilkan buah. Kehilangan hasil yang disebabkannya berkisar antara 70% dan 100% pada cabai rawit, sedangkan pada tanaman cabai besar menyebabkan kehilangan hasil antara 20% dan 100% (Anonim 2004). Perbedaan respon tersebut diduga karena perbedaan ketahanan genetik dan teknik budidaya.
Karakteristik Geminivirus
Geminivirus termasuk dalam virus dengan genom berupa DNA utas tunggal, berpartikel kembar (geminate), dan memiliki tiga subgrup utama. Subgrup I dan subgrup II memiliki genom yang monopartit dan ditularkan oleh wereng daun, menginfeksi tanaman monokotil (subgrup I) dan tanaman dikotil (subgrup II). Subgrup yang ke III memiliki genom yang bipartit, menginfeksi tanaman dikotil dan ditularkan oleh kutu kebul (Hull 2002). Di antara tiga subgrup tersebut, subgrup ketiga memiliki anggota yang lebih banyak dan beragam.
5
dengan geminivirus yang telah dilaporkan menyerang cabai sebelumnya (Sukamto 2005). Geminivirus ini kemudian dinamakan pepper yellow leaf curl indonesia virus (PepYLCIDV) yang menyebabkan gejala berupa penguningan disertai penggulungan dan pengeritingan daun pada tanaman yang diinfeksinya, pada beberapa kasus menyebabkan kekerdilan pada tanaman.
Di lapangan, geminivirus hanya ditularkan oleh serangga vektornya. Jika inang utama tidak tersedia, virus ini mampu bertahan dengan baik pada babadotan (Ageratum conyzoides) dan gulma bunga kancing (Gomphrena globosa). Oleh karena itu, serangannya dapat terjadi terus-menerus selama inang alternatif bagi virus tersedia. Selain melalui vektor, virus ini dapat ditularkan melalui penyambungan, tetapi virus tidak tertularkan melalui biji maupun melalui sap tanaman sakit (mekanis) (Hull 2002).
Virulensi geminivirus akan lebih baik pada musim kemarau, yang ditandai dengan kecepatan pemunculan gejala sejak inokulasi oleh vektornya (Hidayat 12 April 2005, komunikasi pribadi). Hal ini didukung dengan lebih tingginya populasi vektor pada musim tersebut, akan tetapi belum ada laporan yang secara khusus menjelaskan mengapa hal tersebut terjadi. Dugaan sementara yaitu adanya tekanan lingkungan yang secara langsung mempengaruhi keadaan tanaman inang sehingga menurunkan ketahanannya terhadap serangan virus.
Kemaknaan B. tabaci sebagai Vektor Geminivirus
B. tabaci memiliki kisaran inang yang luas mencakup 600 spesies dari 67 famili tanaman yang berbeda (Brown 2003). Selain inang yang luas, B. tabaci sebagai vektor virus mampu menularkan 60 jenis virus, di antaranya geminivirus, closterovirus, nepovirus, carlavirus, potyvirus dan rod-shape DNA virus (Smith 2003). Sebagian besar virus yang ditularkannya merupakan penyebab penyakit penting pada tanaman budidaya sehingga keberadaan B. tabaci berpotensi menyebabkan epidemi penyakit.
6
virus di lapangan, terutama pada pertanaman cabai. Anonim (2004) melaporkan efektivitas penularan virus oleh imago B. tabaci di rumah kaca dengan periode makan akuisisi 30 menit mencapai 40%. Hal ini tentu saja sangat berperan dalam penyebaran penyakit karena dalam waktu yang cukup singkat, seekor imago B. tabaci mampu menularkan virus pada beberapa tanaman. Melalui hasil penelitian yang lain, Rusli et al (1999) melaporkan efektivitas penularan oleh imago B. tabaci mencapai 80%. Hasil penelitian tersebut menunjukkan B. tabaci memiliki keefektivan yang cukup tinggi dalam menularkan geminivirus, oleh karena itu intensitas serangan di lapangan juga menunjukkan hasil yang cukup tinggi.
B. tabaci merupakan faktor pembatas dalam penyebaran virus di lapangan. Walaupun sumber inokulum melimpah di lapangan, jika serangga ini tidak ada maka ledakan kejadian penyakit tidak akan terjadi. Karenanya, di samping karakteristik virus itu sendiri, karakteristik vektor merupakan faktor utama yang menentukan keberhasilan penularan dan penyebaran virus secara alami.
PGPR dalam Teknik Pengendalian Penyakit Tanaman
7
PGPR memiliki spektrum yang luas terhadap organisme pengganggu tanaman sehingga menjadikannya lebih efektif, ekonomis dan praktis untuk diterapkan sebagai model dalam teknik pengendalian penyakit tanaman (Ramamoorthy et al. 2001). Hingga saat ini, pemanfaatan PGPR telah banyak dilakukan dan menunjukkan potensi yang cukup tinggi sebagai agens pengendali hayati penyakit tanaman yang disebabkan oleh cendawan, bakteri, virus dan nematoda. Beberapa strain PGPR yang digunakan dilaporkan efektif dalam mengendalikan patogen tanaman seperti: 1) golongan cendawan, diantaranya: Botrytis cinerea, Colletotrichum gloeosporioides, C. orbiculare, Phytium Sclerospora graminicola, Puccinia psidii, Fusarium oxysporum, Sklerotium rolfsii, Rhizoctonia solan, Phytophthora infestans dan Peronospora tabacina; 2) golongan bakteri, diantaranya: Erwinia tracheiphila, E. carotovora, Pseudomonas syringiae dan Ralstonia solanacearum; 3) golongan virus, diantaranya: CMV, TMV, TNV dan ToMoV; 4) golongan nematoda, diantaranya: Heterodera sp., Meloidogyne spp. dan Pratylenchus spp. (De Meyer & Hofte 1997; Jetiyanon et al. 2003; Jetiyanon & Kloepper 2002; Kloepper et al. 2004; Maurhofer et al. 1994; Murphy et al. 2000; Nirajan Raj et al. 2003; Oostendorp & Sikora 1990; Pieterse et al. 1996; Raupach & Kloepper 1998; Siddiqui & Shaukat 2004; Yan et al. 2002; Zehnder et al. 2000). Melalui penelitian-penelitian yang telah dilakukan tersebut dapat diketahui bahwa peran PGPR dalam menekan penyakit tanaman sangat besar dan memiliki peluang yang cukup tinggi untuk dijadikan sebagai teknik pengendalian penyakit tanaman yang utama di masa mendatang.
Bacillus spp. dan Pseudomonas spp. sebagai Agensi Hayati Pengendalian
Virus Tanaman
8
terhadap kedua genus bakteri ini banyak diarahkan pada penyakit-penyakit yang disebabkan cendawan, bakteri dan nematoda seperti yang telah disebutkan diatas dan telah dilaporkan efektif. Walaupun belum banyak dilaporkan, namun kedua genus bakteri juga memiliki pengaruh positif terhadap virus tanaman.
Beberapa strain Bacillus spp. telah dilaporkan efektivitasnya terhadap virus tanaman. Murphy et al. (2000) melaporkan adanya penurunan kejadian dan keparahan penyakit yang disebabkan ToMoV secara nyata pada tanaman tomat yang diberi perlakuan bakteri dibandingkan dengan kontrol, hal ini juga berkorelasi positif dengan keberadaan vektornya (B. tabaci). Pada tanaman mentimun, penggunaan Bacillus spp. di lapangan mampu menekan perkembangan penyakit CMV secara nyata (Jetiyanon et al. 2002). Penelitian serupa yang dilakukan berikutnya menunjukkan penggunaan Bacillus spp. mampu menekan kejadian penyakit hingga 80% (Jetiyanon et al. 2003). Melalui percobaan rumah kaca, penggunaan Bacillus spp. mampu menekan kejadian penyakit CMV hingga 64%, disamping itu juga menurunkan tingkat keparahan penyakitnya (Murphy et al. 2003; Zehnder et al 2000).
9
Induksi Ketahanan Sistemik oleh PGPR
Melalui penelitian-penelitian yang telah disebutkan di atas, dapat disimpulkan adanya induksi ketahanan sistemik tanaman oleh bakteri sehingga tanaman menjadi lebih toleran terhadap patogen yang menyerang, bahkan diantaranya menjadi tahan terhadap patogen tertentu. Ketahanan Sistemik Terinduksi (induced systemic resistance [ISR]) pada dasarnya memiliki kesamaan dengan Ketahanan Sistemik yang Diterima (systemic acquired resistance [SAR]). Mekanisme ini terjadi sebagai akibat adanya infeksi oleh patogen sehingga tanaman memberikan respon berupa reaksi-reaksi pertahanan seperti reaksi hipersensitif yang menyebabkan terjadinya lesio nekrotik pada daerah terserang. Berbeda dengan SAR, ISR tidak menyebabkan adanya gejala tampak seperti lesio nekrotik (Compant et al. 2005).
Ramamoorthy et al. (2001) memaparkan bahwa mekanisme ISR terjadi sebagai akibat perubahan fisiologi tanaman yang kemudian menstimulasi terbentuknya senyawa kimia yang berguna dalam pertahanan terhadap serangan patogen. Perubahan fisiologi tersebut dapat berupa modifikasi struktural dinding sel atau perubahan reaksi biokimia pada tanaman inang. Beberapa faktor yang dapat menyebabkan adanya induksi ketahanan sistemik oleh bakteri yaitu: 1) adanya sumbangan lipopolisakarida oleh bakteri; 2) produksi siderofor oleh bakteri; dan 3) produksi asam salisilat, yang dapat terjadi secara langsung oleh bakteri ataupun secara tidak langsung (van Loon et al. 1998).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan di Laboratorium Mikologi dan Klinik Tanaman Departemen Proteksi Tanaman serta di Rumah Kaca Kebun Percobaan Cikabayan Institut Pertanian Bogor sejak bulan Januari 2006 sampai Juni 2006.
Bahan
Bakteri yang digunakan dalam penelitian ini yaitu B. polymixa BG25, B. subtilis SB3, P. fluorescens PG01 dan P. fluorescens ES32 yang merupakan koleksi Klinik Tanaman Departemen Proteksi Tanaman. Bahan tanaman uji yang digunakan adalah cabai besar (C. annuum) varietas Hot Chili yang didapat dari toko pertanian. Sumber inokulum berasal dari Koleksi Laboratorium Virologi Departemen Proteksi Tanaman, yaitu tanaman tomat yang terinfeksi geminivirus isolat Segunung.
Metode Pembuatan Suspensi Bakteri
Bakteri yang berasal dari media Nutrient Broth (NB) diremajakan pada media Triptic Soy Agar (TSA [kelompok Bacillus]) dan King’s B (kelompok Pseudomonas) dengan menggoreskan masing-masing 1 loop suspensi pada media tersebut. Bakteri yang telah diremajakan diinkubasikan pada suhu ruangan selama 48 jam. Sebanyak 15 loop biakan murni yang didapat diencerkan dalam 100 ml NaCl 0,85% sehingga didapat suspensi bakteri (stok) dengan kepadatan masing-masing 1012 cfu/ml untuk Bacillus dan 1013 cfu/ml untuk Pseudomonas (Jamaliyah 2005).
Perlakuan Benih dan Penanaman Tanaman Uji
11
suspensi kombinasi keempat bakteri tersebut (Campuran) dengan kepadatan yang sama. Benih cabai yang sebelumnya telah dicuci kemudian dimasukkan ke dalam suspensi bakteri (BG25, SB3, PG01, ES32 dan Campuran) dan dibiarkan selama 10 jam pada suhu ruang. Sebagai kontrol, benih cabai direndam pada 8 ml NaCl 0,85% selama 10 jam.
Benih yang telah direndam kemudian ditanam pada baki semai yang berisi media berupa campuran tanah dan pupuk kandang dengan perbandingan 2:1 (v/v). Setelah berumur empat minggu setelah tanam (MST) dilakukan pemindahan tanaman ke dalam polybag (35x35 cm2) dengan media yang sama pada pesemaian.
Perbanyakan Serangga Vektor Bebas Virus
Imago B. tabaci koleksi Laboratorium Virologi Departemen Proteksi Tanaman dibiakkan pada tanaman kapas yang berumur dua minggu dalam kurungan kedap serangga (1x1x1 m3). Serangga dibiarkan berkembang biak hingga jumlah yang mencukupi untuk inokulasi virus.
Pemurnian dan Perbanyakan Isolat Virus
Imago B. tabaci yang bebas virus diinfestasikan pada tanaman tomat yang terinfeksi geminivirus isolat Segunung dan dibiarkan selama 24 jam melewati periode makan akuisisi. Kemudian, sebanyak sepuluh ekor imago dipindahkan ke tanaman cabai sehat yang telah berumur lima minggu dan di inkubasi hingga tanaman menunjukkan gejala penyakit kuning.
Inokulasi Virus pada Tanaman Uji
12
gangguan fisik (tanaman digoyang) untuk memastikan serangga vektor hinggap dan makan (inokulasi) pada setiap tanaman uji. Metode ini dilakukan khususnya untuk tanaman berumur 3 MST dan 5 MST, sedangkan untuk tanaman berumur 6 MST, inokulasi dilakukan secara langsung dengan menaruh sumber inokulum dan sejumlah vektor diantara tanaman yang akan diuji selama tiga hari. Hal ini dikarenakan ukuran kurungan inokulasi tidak dapat menampung seluruh tanaman berumur 6 MST yang telah dipindahkan ke polybag yang lebih besar.
Pengamatan Morfologi Tanaman
Variabel pengamatan terkait morfologi tanaman yaitu: 1) tinggi tanaman, diukur setiap minggu sejak tanaman berumur 5 hingga 15 MST; 2) jumlah cabang utama dan sekunder, dihitung pada saat tanaman mencapai masa pembungaan (15 MST); 3) jumlah daun dan bunga, dihitung pada 15 MST.
Pengukuran keparahan penyakit
Pengamatan mulai dilakukan 3 minggu setelah periode makan inokulasi (3 MSI) dengan mengukur skor penyakit pada masing-masing tanaman uji. Kategori skor yang digunakan (gambar lampiran 1):
0 : tidak bergejala
1 : penguningan tulang daun
2 : seluruh tulang daun menguning, daun keriting 3 : lamina daun menguning, daun keriting
4 : tanaman kerdil, seluruh daun menguning, reduksi ukuran daun, daun keriting sepenuhnya
13
Variabel lain dari penyakit kuning yaitu masa inkubasi yang diamati sejak periode makan inokulasi berakhir hingga munculnya gejala awal dari penyakit tersebut.
Rancangan Percobaan
Percobaan disusun dalam rancangan acak faktorial dengan dua faktor utama yaitu jenis bakteri dan waktu inokulasi virus. Unit perlakuan terdiri dari enam taraf yang terdiri dari strain bakteri (BG25, SB3, PG01 dan ES32), kombinasi keempat strain bakteri (Campuran) dan pembanding (Kontrol) yang disusun dalam empat blok dengan tiga unit contoh pada masing-masing blok.
Analisis Data
HASIL DAN PEMBAHASAN
Pengaruh PGPR dan Waktu Inokulasi Terhadap Morfologi Tanaman
Pengaruh PGPR terhadap Perkembangan Tinggi Tanaman, Jumlah Cabang, Daun dan Bunga
PGPR mampu meningkatkan pertumbuhan tanaman secara langsung melalui hormon-hormon pertumbuhan yang dihasilkan seperti Giberilin (GAs) dan indole-3-acetic acid (IAA) (Barazani & Friedman 1999; Glickman et al. 1997). Disamping itu, PGPR juga mampu mensintesis sitokinin dan beberapa fitohormon lain (Nelson 2004). Oleh karena itu, umumnya tanaman yang diberi perlakuan PGPR mampu tumbuh lebih baik.
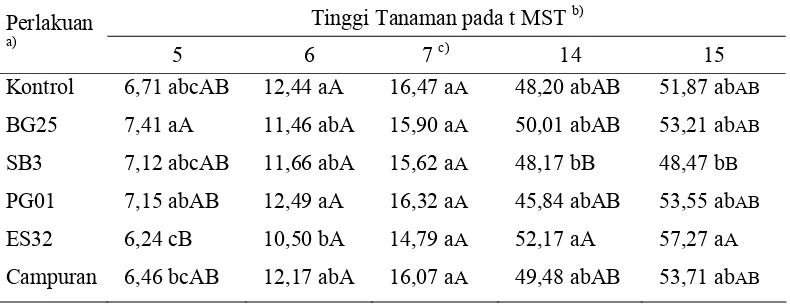
Tabel 1 Pengaruh perlakuan bakteri terhadap tinggi tanaman cabai pada beberapa waktu pengamatan
P. fluorescens PG01; ES32 = P. fluorescens ES32.
b)
angka yang diikuti dengan huruf kecil dan huruf kapital yang sama dalam satu kolom tidak berbedanyata pada α = 0,05 dan α = 0,01;
c)
tinggi tanaman pada minggu ke-8 hingga minggu ke-13 menunjukkan hasil yangtidak berbeda nyata pada semua perlakuan sehingga data tidak ditampilkan.
15
Hal ini mengindikasikan bahwa perlakuan bakteri tidak mampu meningkatkan pertumbuhan tinggi tanaman cabai secara nyata.
Perbedaan pertumbuhan tinggi tanaman justru diperlihatkan diantara perlakuan bakteri yang diberikan. Pada 5 dan 6 MST, tanaman yang diberi perlakuan P. fluorescens PG01 dan perlakuan bakteri lainnya mampu memacu pertumbuhan tanaman lebih baik (nyata pada PG01) dibandingkan tanaman yang diberi perlakuan P. fluorescens ES32. Akan tetapi, sejak 7 MST, tanaman dengan perlakuan P. fluorescens ES32 mulai menunjukkan kemampuan dalam meningkatkan pertumbuhan tinggi tanaman. Kemampuan P. fluorescens ES32 ini terus ditunjukkan hingga akhir pengamatan, bahkan dalam perkembangannya, tanaman dengan bakteri ini cenderung menunjukkan pertumbuhan yang lebih baik dibandingkan perlakuan lainnya. Pseudomonas fluorescens ES32 juga menunjukkan kemampuan memacu pertumbuhan yang lebih baik dibandingkan B. subtilis SB3. Hal ini bertentangan dengan penelitian serupa yang dilakukan oleh Wirianti (2005), dimana tanaman dengan perlakuan B. subtilis SB3 selalu menunjukkan tinggi tanaman yang lebih baik dibandingkan perlakuan bakteri lainnya dan berbeda nyata dengan tanaman kontrol.
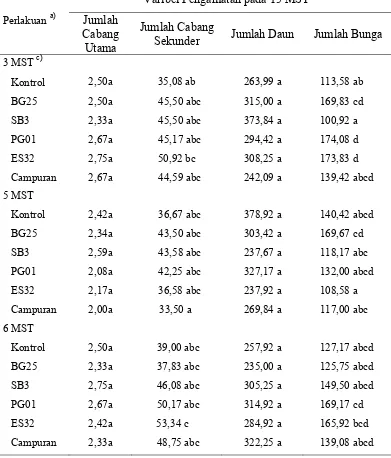
Tabel 2 Pengaruh perlakuan bakteri terhadap jumlah cabang utama, cabang sekunder, daun dan bunga
Variabel Pengamatan pada 15 MST b) Perlakuan a) Jumlah
P. fluorescens PG01; ES32 = P. fluorescens ES32.
b)
16
Melalui Tabel 2 terlihat adanya keragaman pada kemampuan bakteri dalam memacu perkembangan vegetatif dan generatif tanaman. Secara umum, tidak ada perbedaan jumlah cabang utama pada tanaman cabai. Akan tetapi pada perkembangan cabang sekunder, perlakuan B. subtilis SB3, P. fluorescens PG01 dan P. fluorescens ES32 menunjukkan hasil yang berbeda nyata dengan kontrol dimana jumlah cabang sekunder yang dihasilkan lebih banyak. Pada penghitungan jumlah daun perlakuan bakteri tidak menunjukkan pengaruh yang nyata, walau demikian tanaman dengan perlakuan P. fluorescens PG01 menunjukkan jumlah daun yang lebih banyak dari perlakuan lainnya. Pada penelitian ini, B. subtilis SB3 terlihat hanya mampu memacu pertumbuhan vegetatif tanaman melalui peningkatan jumlah cabang sekunder tanaman.
Perlakuan bakteri juga mampu memacu pertumbuhan generatif tanaman. Perlakuan P. fluorescens PG01 menunjukkan hasil yang berbeda nyata dibanding kontrol, sementara perlakuan bakteri lainnya tidak. Selain itu, perlakuan bakteri ini juga berbeda nyata dengan perlakuan B. subtilis SB3. Dengan mengasumsikan bahwa persentase keberhasilan bunga menjadi buah adalah sama pada tiap perlakuan, maka dapat diketahui bahwa P. fluorescens PG01 dapat membantu tanaman menghasilkan produksi yang lebih banyak dibanding perlakuan lainnya.
Hasil ini menunjukkan adanya perbedaan bakteri dalam memacu pertumbuhan tanaman. Secara umum adanya bakteri mampu meningkatkan kemampuan pertumbuhan tanaman, akan tetapi kemampuan tersebut tidak selalu terjadi pada kedua fase pertumbuhan tanaman (vegetatif dan generatif) dan tidak selalu memacu semua variabel ukur pertumbuhan pada masing-masing fase pertumbuhan.
Pengaruh Waktu Inokulasi Virus terhadap Perkembangan Tinggi Tanaman, Jumlah Cabang, Daun dan Bunga
17
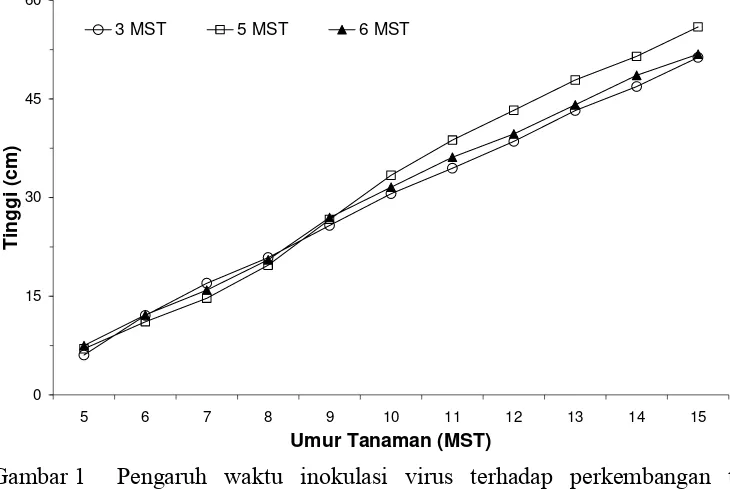
virus pada umur tanaman yang lebih tua. Dari Gambar 1 terlihat bahwa tanaman yang diinokulasi virus pada 5 MST secara nyata memiliki pertumbuhan tinggi yang lebih baik (55,93 cm) dibandingkan dengan tanaman yang diinokulasi virus pada 3 MST (51,31 cm) dan 6 MST (51,81 cm). Hal ini kemungkinan disebabkan karena nilai tengah yang digunakan merupakan rata-rata nilai tinggi tanaman dari semua unit perlakuan pada taraf waktu inokulasi yang sama.
0
Gambar 1 Pengaruh waktu inokulasi virus terhadap perkembangan tinggi tanaman; nilai rata-rata yang digunakan berasal dari seluruh perlakuan pada waktu inokulasi yang sama; 3, 4 dan 5 MST menunjukkan umur tanaman saat inokulasi virus.
18
tanaman (waktu inokulasi 6 MST) sudah memasuki fase generatif, sehingga pertumbuhan vegetatifnya mulai terhambat.
Tabel 3 Pengaruh waktu inokulasi terhadap jumlah cabang utama, cabang sekunder, daun dan bunga
Variabel Pengamatan pada 15 MST a) Waktu
Inokulasi
(MST) Jumlah Cabang Utama Jumlah Cabang Sekunder Jumlah Daun Jumlah Bunga 3 2,57 aA 44,46 aAB 299,60 aA 145,28 abA 5 2,26 bB 39,35 bB 292,49 aA 130,97 bA 6 2,50 aAB 45,86 aA 286,71 aA 146,10 aA
a)
angka yang diikuti dengan huruf kecil dan huruf kapital yang sama dalam satu kolom berturut-turut tidak berbedanyata pada α = 0,05 dan α = 0,01.
Perbedaan waktu inokulasi virus yang berimplikasi pada perbedaan waktu infeksi dan perkembangan penyakit juga menunjukkan perbedaan pada variabel pengamatan lain yang ditunjukkan pada Tabel 3. Jumlah cabang pada tanaman yang diinokulasi pada umur 5 MST berbeda nyata dengan dua waktu inokulasi lainnya. Pada waktu inokulasi tersebut, jumlah cabang utama dan cabang sekundernya lebih banyak dibanding dua taraf lainnya. Jumlah daun pada ketiga waktu inokulasi tidak memiliki pengaruh yang berbeda nyata. Jumlah daun yang lebih tinggi pada waktu inokulasi 3 dan 5 MST kemungkinan terjadi sebagai respon terhadap adanya gangguan penyakit sehingga tanaman menghasilkan daun yang lebih banyak.
Bertentangan dengan hal diatas, jumlah bunga pada tanaman yang diinokulasi virus pada 5 MST menunjukkan hasil yang lebih rendah (berbeda nyata) dibanding dua taraf waktu lainnya. Akan tetapi, tidak dapat diketahui secara pasti faktor penyebab jumlah bunga yang lebih rendah ini.
Pengaruh Interaksi antara Perlakuan PGPR dan Waktu Inokulasi Virus terhadap Perkembangan Tinggi Tanaman, Jumlah Cabang, Daun dan Bunga
19
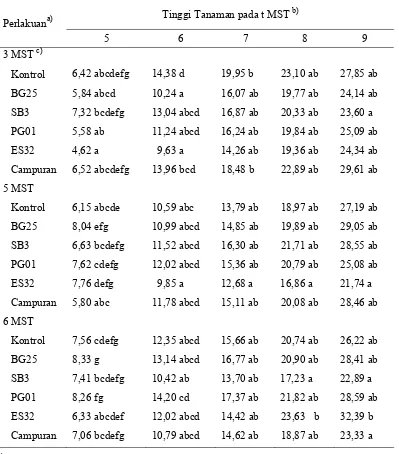
Tabel 4 Pengaruh interaksi antara perlakuan bakteri dan waktu inokulasi virus terhadap perkembangan tinggi tanaman cabai
Tinggi Tanaman pada t MST b) Perlakuana)
P. fluorescens PG01; ES32 = P. fluorescens ES32.
b)
angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbedanyata pada
α = 0,05.
c)
20
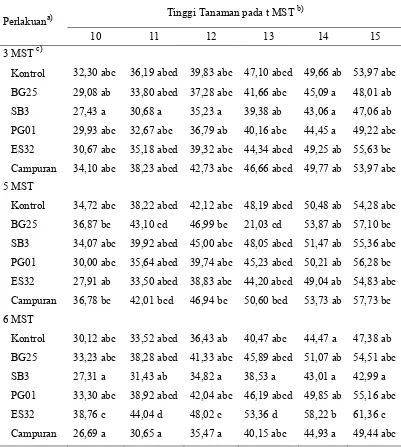
Tabel 4 Pengaruh interaksi antara waktu inokulasi virus dan perlakuan bakteri terhadap perkembangan tinggi tanaman cabai (lanjutan)
Tinggi Tanaman pada t MST b)
P. fluorescens PG01; ES32 = P. fluorescens ES32.
b)
angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbedanyata pada
α = 0,05.
c)
21
dan bunga). Secara umum tidak terdapat perbedaan tinggi tanaman yang nyata antara perlakuan pada taraf waktu inokulasi yang berbeda. Pertumbuhan tinggi tanaman antara waktu inokulasi virus yang berbeda cenderung menunjukkan hasil yang sama.
Hasil percobaan ini menunjukkan bahwa perlakuan bakteri tidak dapat meningkatkan pertumbuhan tinggi tanaman walaupun terdapat variasi waktu terinfeksinya tanaman oleh geminivirus. Walaupun terdapat keragaman dalam peningkatan tinggi tanaman, tetapi tiap perlakuan didalamnya tidak menunjukkan perbedaan yang nyata dengan perlakuan yang sama pada tiap taraf waktu inokulasi virus yang berbeda. Hal ini juga mengindikasikan bahwa perlakuan bakteri tidak dapat membantu tanaman dalam mengurangi gangguan fisiologis yang berdampak pada penghambatan pertumbuhan tanaman yang disebabkan infeksi geminivirus.
Melalui Tabel 4, dapat diketahui bahwa perlakuan P. fluorescens ES32 dengan waktu inokulasi geminivirus pada 6 MST memiliki perbedaan nyata dibanding tanaman kontrol. Hal ini menunjukkan bahwa P. fluorescens ES32 mampu meningkatkan pertumbuhan tanaman yang lebih baik jika geminivirus baru menginfeksi tanaman pada umur 6 MST. Sedangkan, jika tanaman terinfeksi sebelum berumur 6 MST maka P. fluorescens ES32 tidak dapat meningkatkan pertumbuhan tanaman secara nyata jika dibandingkan dengan tanaman kontrol. Disamping itu, tanaman dengan perlakuan P. fluorescens ES32 menunjukkan pertumbuhan yang lambat pada awal penanaman tetapi ketika tanaman mulai memasuki umur 9 MST terjadi peningkatan tinggi tanaman yang lebih baik. Ada kemungkinan bahwa P. fluorescens SB3 memacu pertumbuhan tanaman dengan lebih baik ketika tanaman mulai memasuki fase vegetatif cepat.
22
Tabel 5 Pengaruh interaksi perlakuan bakteri dan waktu inokulasi virusterhadap jumlah cabang utama, cabang sekunder, daun dan bunga
Varibel Pengamatan pada 15 MST b)
Perlakuana) Jumlah Cabang Utama
Jumlah Cabang
Sekunder Jumlah Daun Jumlah Bunga
3 MST c)
P. fluorescens PG01; ES32 = P. fluorescens ES32.
b)
angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbedanyata pada
α = 0,05.
c)
waktu inokulasi geminivirus pada tiap unit contoh perlakuan
23
bakteri yang digunakan tidak memiliki sifat yang sinergistik sehingga tidak dapat meningkatkan pertumbuhan tanaman dengan lebih baik.
Tabel 5 menunjukkan interaksi antara waktu inokulasi geminivirus dan strain bakteri terhadap jumlah cabang, daun dan bunga. Secara umum, tidak terdapat perbedaan yang nyata antara perlakuan bakteri dan kontrol pada tiap waktu inokulasi. Hal ini juga menunjukkan bahwa adanya perbedaan waktu inokulasi geminivirus pada tiap perlakuan tidak terlalu berpengaruh terhadap perkembangan cabang, daun dan bunga pada tiap perlakuan bakteri yang diberikan. Artinya, walaupun geminivirus menginfeksi hanya pada saat tanaman berumur 6 MST, maka pengaruhnya terhadap pembentukan cabang, daun dan bunga tidak akan berbeda dengan tanaman yang terinfeksi sebelum tanaman berumur 6 MST.
Melalui uraian di atas, dapat kita ketahui bahwa interaksi antara waktu inokulasi geminivirus dan perlakuan bakteri tidak terlalu mempengaruhi pertumbuhan tanaman. Tidak semua kombinasi dua faktor perlakuan yang ada menunjukkan hasil positif terhadap morfologi tanaman.
Pengaruh PGPR dan Waktu Inokulasi Virus terhadap Perkembangan Penyakit Kuning
Pengaruh PGPR terhadap Masa Inkubasi dan Perkembangan Keparahan Penyakit
24
Gambar 2 Pengaruh perlakuan bakteri terhadap masa inkubasi penyakit
Kontrol = tanpa bakteri; BG25= B. polymixa BG25; SB3 = B. subtilis SB3; PG01 = P. fluorescens PG01; ES32 = P. fluorescens ES32; Huruf kecil dan huruf kapital yang sama tidak berbeda nyata pada α = 0,05 dan α = 0,01
Walaupun perlakuan bakteri menunjukkan kemampuan dalam menghambat masa inkubasi penyakit, akan tetapi perlakuan bakteri belum dapat dikatakan efektif secara ekonomi. Hal ini dikarenakan selisih waktu penghambatan masa inkubasi penyakit hanya berbeda sekitar satu hari. Selisih waktu tersebut tidak akan menunjukkan penghambatan yang berarti dalam perkembangan penyakit yang disebabkan oleh virus.
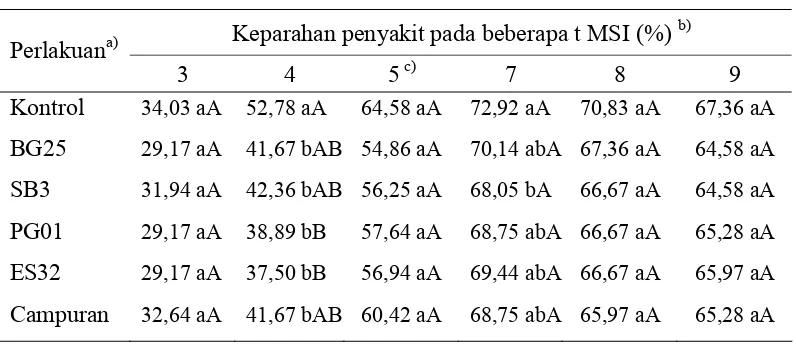
Daya penghambatan terhadap pemunculan gejala ini berimplikasi pada perkembangan keparahan penyakit geminivirus. Tanaman pada perlakuan bakteri menunjukkan perbedaan tingkat keparahan penyakit yang lebih rendah pada minggu awal pengamatan. Tanaman yang diberi perlakuan P. fluorescens PG01 dan ES32 menunjukkan perbedaan yang nyata pada 4 MSI (Tabel 6). Memasuki 5 MSI, seluruh perlakuan bakteri menunjukkan tingkat keparahan penyakit yang tidak berbeda nyata dengan perlakuan kontrol. Hal ini menunjukkan bahwa geminivirus mulai dapat berkembang dengan baik di semua tanaman pada perlakuan bakteri sehingga menunjukkan tingkat keparahan yang sama dengan
25
tanaman kontrol. Perlakuan bakteri yang diberikan secara umum tidak dapat melindungi tanaman terhadap serangan geminivirus.
Tabel 6 Pengaruh perlakuan bakteri terhadap keparahan penyakit kuning Keparahan penyakit pada beberapa t MSI (%) b) Perlakuana)
P. fluorescens PG01; ES32 = P. fluorescens ES32.
b)
angka yang diikuti dengan huruf kecil dan huruf kapital yang sama dalam satu kolom berturut-turut tidak berbedanyata pada α = 0,05 dan α = 0,01.
c)
seluruh perlakuan pada pengamatan minggu ke-6 setelah inokulasi tidak berbeda nyata
Pada minggu ke-8 setelah inokulasi terjadi penurunan tingkat keparahan penyakit, baik pada seluruh tanaman yang diberi perlakuan bakteri maupun tanaman kontrol. Hal ini menunjukkan adanya penghambatan kembali terhadap perkembangan geminivirus dalam tanaman inang. Terhambatnya perkembangan geminivirus ini belum mengindikasikan adanya induksi ketahanan tanaman, akan tetapi lebih mengindikasikan adanya proses penyembuhan (recovery) yang berasal dari tanaman itu sendiri.
26
Pengaruh Waktu Inokulasi terhadap Masa Inkubasi dan Perkembangan Keparahan Penyakit
Perbedaan waktu inokulasi virus tidak menyebabkan perbedaan waktu munculnya gejala tanaman (masa inkubasi) secara nyata (Gambar 3). Hal ini mengindikasikan bahwa dalam mengekspresikan gejala awal, geminivirus tidak bergantung pada waktu inokulasi oleh serangga vektor. Perlakuan waku inokulasi mungkin akan memberikan pengaruh jika waktu inokulasi dilakukan pada tanaman yang berumur lebih dari 6 MST.
Gambar 3 Pengaruh waktu inokulasi virus terhadap masa inkubasi penyakit Huruf kecil dan huruf kapital yang sama tidak berbeda nyata pada α = 0,05 dan α = 0,01
27
Kejadian ini dapat disebabkan karena pada umur yang lebih tua saat inokulasi, tanaman telah membentuk senyawa-senyawa yang dapat menghambat infeksi patogen. Dapat pula disebabkan ketersediaan zat yang dibutuhkan oleh virus untuk berkembang dan mengekspresikan gejala lebih sedikit dibandingkan pada tanaman muda. Hasil ini menunjukkan terjadinya peningkatan ketahanan terhadap infeksi virus pada tanaman yang diinokulasi pada umur yang lebih tua. Tabel 7 Pengaruh waktu inokulasi virus terhadap keparahan penyakit
Keparahan Penyakit pada t MSI (%) a)
angka yang diikuti dengan huruf kecil dan huruf kapital yang sama dalam satu kolom berturut-turut tidak berbedanyata pada α = 0,05 dan α = 0,01.
b)
keparahan penyakit pada 3 MSI memiliki komposisi beda nyata yang sama dengan pengamatan pada 4 MSI
28
Pengaruh Interaksi Perlakuan Bakteri dan Waktu Inokulasi terhadap Masa Inkubasi dan Tingkat Keparahan Penyakit
Melalui analisis statistik, dapat terlihat bahwa interaksi antara waktu inokulasi virus dan strain bakteri yang digunakan tidak memberikan pengaruh yang nyata dalam menghambat waktu pemunculan gejala awal penyakit kuning. Perlakuan bakteri yang diberikan tidak mempengaruhi lama terlihatnya ekspresi awal gejala baik dalam waktu inokulasi yang sama maupun antara waktu inokulasi yang berbeda (Gambar 4). Walaupun demikian, perlakuan bakteri pada tiap waktu inokulasi virus menunjukkan ekspresi gejala yang secara rata-rata lebih lambat dibanding pada tanaman kontrol.
0
Kontrol BG25 SB3 PG01 ES32 Mix
Gambar 4 Pengaruh interaksi antara perlakuan bakteri dan waktu inokulasi virus terhadap masa inkubasi penyakit
Kontrol = tanpa bakteri; BG25= B. polymixa BG25; SB3 = B. subtilis SB3; PG01 = P. fluorescens PG01; ES32 = P. fluorescens ES32
29
Tabel 8 Pengaruh interaksi perlakuan bakteri dan waktu inokulasi virus terhadap perkembangan keparahan penyakit
Keparahan Penyakit pada t msi b)
Perlakuana)
P. fluorescens PG01; ES32 = P. fluorescens ES32.
b)
angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbedanyata pada
α = 0,05.
c)
waktu inokulasi geminivirus pada tiap unit contoh perlakuan
d)
seluruh perlakuan pada pengamatan minggu ke-3 setelah inokulasi tidak berbeda nyata
30
semakin tinggi) maka tingkat keparahan penyakit pada tiap perlakuan akan semakin rendah. Pengamatan yang dilakukan sejak tanaman berusia 3 MSI hingga 9 MSI menunjukkan kecenderungan yang sama dimana terjadi penurunan tingkat serangan geminivirus, walaupun pada 7 MSI beberapa perlakuan bakteri menunjukkan keparahan yang lebih tinggi pada taraf waktu inokulasi 5 MST tetapi keparahannya kembali lebih rendah ketika memasuki 8 MSI. Hal ini mengindikasikan adanya peningkatan toleransi tanaman terhadap penyakit kuning jika infeksi terjadi pada usia tanaman yang lebih tua.
Peningkatan toleransi tanaman pada waktu inokulasi virus yang lebih lama dapat terlihat pada 7 hingga 9 MSI dimana secara umum terdapat perbedaan yang nyata antara perlakuan pada taraf waktu inokulasi yang lebih rendah (3 MST) dan lebih tinggi (8 MST). Perbedaan nyata di 7 dan 8 MSI pada beberapa strain bakteri (tidak terjadi pada kontrol) dengan taraf waktu inokulasi 3 dan 6 MST menunjukkan bahwa perlakuan bakteri akan efektif menekan tingkat keparahan penyakit geminivirus dengan syarat tanaman tidak terinfeksi pada perkembangan awal tanaman (3 MSI) melainkan baru terinfeksi saat tanaman sudah dapat berkembang dengan baik (6 MST).
Hasil penelitian ini tidak dapat menunjukkan adanya induksi ketahanan sistemik oleh bakteri terhadap adanya infeksi geminivirus. Tanaman kontrol pada taraf waktu inokulasi 5 dan 6 MST menunjukkan kemampuan penekanan tingkat keparahan penyakit. Hal ini menunjukkan bahwa secara alami tanaman mampu meningkatkan toleransinya terhadap infeksi geminivirus seiring dengan bertambahnya umur tanaman tersebut. Mekanisme yang terjadi tidak dapat dijelaskan secara pasti tetapi kemungkinannya adalah terjadinya perubahan fisiologis pada tanaman yang dalam perkembangannya dapat menghasilkan senyawa-senyawa yang mampu memberikan tekanan pada virus secara langsung ataupun tidak langsung sehingga mengurangi virulensi virus tersebut.
KESIMPULAN
Kesimpulan
Aplikasi empat strain PGPR dengan waktu inokulasi yang berbeda tidak dapat meningkatkan pertumbuhan tanaman secara nyata. Perlakuan bakteri kelompok Pseudomonas mampu memperlambat masa inkubasi penyakit tetapi pada taraf waktu inokulasi yang berbeda tidak menunjukkan hambatan terhadap pemunculan gejala awal.
Keparahan penyakit berkorelasi negatif dengan taraf waktu inokulasi yang digunakan, semakin tinggi taraf waktu inokulasi semakin rendah tingkat keparahan penyakitnya. Taraf waktu inokulasi memiliki peran yang lebih tinggi dalam menekan keparahan penyakit. Pada taraf waktu yang lebih tinggi, perlakuan bakteri memperlihatkan kemampuan menekan keparahan penyakit lebih baik.
Saran
DAFTAR PUSTAKA
Agrios GN. 1997. Plant Pathology. Edisi ke-4. San Diego: Academic Press. Ahmed NE, Kanan HO, Sugimoto Y, Ma YQ, Inanaga S. 2001. Effect of
imidacloprid on incidence of tomato yellow leaf curl virus. Plant Dis. 85(1):84-87.
[Anonim]. 2004. Penyakit kuning rusak tanaman cabai. Lampung post 5 Juli 2004:8(kolom 1-4).
Bloemberg GV, Lugtenberg BJJ. 2001. Molecular basis of plant growth-promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 4:343-350.
Brown JK. 2003. General information begomoviruses. Arizona Biological Science. http://gemini.biosci.arizona.edu/mini-blast.html [23 Mei 2005]. Brown JK, Bird J. 1992. Whitefly-transmitted geminiviruses and associated
disorders in the Americas and the Caribbean Basin. Plant Dis. 76:220-225. Compant S, Duffy B, Nowak J, Clement C, Barka EA. 2005. Use of plant
growth-promoting rhizobacteria for biocontrol of plant diseases: principles, mechanisms of action and future prospect. Applied and Environmental Microbiology 71(9): 4951-4959.
De Meyer G, Hofte M. 1997. Salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 induced resistance to leaf infection by Botrytis cinerea on Bean. Phytopathology 87:588-593.
Gray EJ, Smith DL. 2005. Intracellular and extracellular PGPR: commonalities and distinctions in the plant-bacterium signalling processes. Soil. Biol. Biochem. 37:395-412.
Hadidi A, Khetarpal RK, Koganezawa H. 1998. Plant Virus Disease Control. USA: APS Press.
Hull R. 2002. Matthew’s Plant Virology. Edisi ke-4. New York: Academic Press.
Jamaliyah W. 2005. Ketahanan hidup dan kemampuan antagonisme Bacillus subtilis ERB21, dan Pseudomonas flourescens ES32 dalam berbagai formulasi [skripsi] Bogor: IPB. Fakultas Pertanian.
Jetiyanon K, Fowler WD, Kloepper JW. 2003. Broad-spectrum protection against several pathogens by PGPR mixtures under field conditions in Thailand. Plant Dis. 87:1390-1394.
33
Kloepper JW, Ryu CM, Zhang S. 2004. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 94:1259-1266. Liu L, Kloepper JW, Tuzun S. 1995. Induction of systemic resistance in cucumber
against bacterial angular leaf spot by plant growth-promoting rhizobacteria. Phytopathology 85:843-847.
Maurhofer M, Hase C, Meuwly P, Metraux JP, Defago G. 1994. Induction of systemic resistance of tobacco to tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens strain CHA0: influence of the gacA gene and pyoverdine production. Phytopathology 84:139-146.
Murphy JF, Zehnder GW, Schuster DJ, Sikora EJ, Polston JE, Kloepper JW. 2000. Plant growth-promoting rhizobacterial mediated protection in tomato against tomatomottlevirus. PlantDis. 84:779-784.
Nirajan Raj S, Chaluvaraju G, Amruthesh KN, Shetty HS, Reddy MS, Kloepper JW. 2003. Induction of growth promotion and resistance against downey mildew on pearl millet (Pennisetum glaucum) by Rhizobacteria. Plant Dis. 87:380-384.
Nelson LM. 2004. Plant growth promoting rhizobacteria (PGPR): Prospects for New Inoculants. Crop Management.
Oostendorp M, Sikora RA. 1990. In vitro interrelationship between rhizosphere bacteria and Heterodera schachtii. Rev. Nematol. 13(3):269-274.
Pieterse CMJ, van Wees SCM, Hoffland E, van Pelt JA, van Loon LC. 1996. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell. 8:1225-1237.
Ramamoorthy V, Viswanathan R, Raguchander T, Prakasam V, Samiyappan R. 2001. Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Prot. 20:1-11. Raupach GS, Kloepper JW. 1998. Mixture of plant growth-promoting
rhizobacteria enhance biological control of multiple cucumber pathogens. Phytopathology 88:1158-1164.
Raupach GS, Liu L, Murphy JF, Tuzun S, Kloepper JW. 1996. Induced systemic resistance in cucumber and tomato against cucumber mosaic cucumovirus using plant growth-promoting rhizobacteria (PGPR). Plant Dis. 80:891-94. Rusli ES, Hidayat SH, Suseno R, Tjahjono B. 1999. Geminivirus pada cabai:
variasi gejala dan studi cara penularan. Buletin HPT 2:26-31.
34
Smith IM. 2003. Data sheets on quarantine pests: Bemisia tabaci. EPPO Quarantine Pests. http://gemini.biosci.arizona.edu/links.html [30 Mei 2005].
Sukamto. 2005. Mengenali virus tanaman cabai. http://www.beritaiptek.com/. [13 Juli 2006].
Van Loon LC, Bakker PAHM, Pietersen CMJ. 1998. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 36:453-83.
Widodo 1993. Penggunaan Pseudomonas spp. kelompok fluorescens untuk pengendalian penyakit akar gada (Pseudomonas brassicae Wor) pada caisin (Brassica campestris L) var Chinensis (Rupr) Olson [tesis]. Bogor: IPB. Program Pasca Sarjana.
Wirianti DA. 2005. Pengaruh penggunaan bakteri pemacu pertumbuhan tanaman dalam menekan populasi kutu kebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) pada tanaman cabai [skripsi]. Bogor: IPB. Fakultas Pertanian.
Yan Z, Reddy MS, Ryu CM, McInroy JA, Wilson M, Kloepper JW. 2002. Induced systemic protection against tomato late blight elicited by plant growth-promoting rhizobacteria. Phytopathology 92:1329-1333.
36
Lampiran 1 Gambar gejala tanaman yang terinfeksi geminivirus dengan berbagai tingkat keparahan; (a) gejala dengan nilai skor 1; (b) gejala dengan nilai skor 2; (c) dan (d) gejala dengan nilai skor 3.
(a) (b)
37
Lampiran 2 Tabel pengaruh perlakuan bakteri terhadap perkembangan tinggi tanaman cabai Tinggi Tanaman pada t MST b) Perlakuana)
5 6 7 8 9 10 11 12 13 14 15
Kontrol 6,71abc 12,44a 16,47a 20,94a 27,09a 32,38a 35,97a 39,46a 45,26a 48,20ab 51,87ab
BG25 7,41a 11,46ab 15,90a 20,18a 27,20a 33,06a 38,39a 41,87a 46,19a 50,01ab 53,21ab
SB3 7,12abc 11,66ab 15,62a 19,75a 25,01a 29,60a 34,01a 38,35a 41,98a 48,17b 48,47b
PG01 7,15ab 12,49a 16,32a 20,82a 26,25a 31,07a 35,74a 39,52a 43,86a 45,84ab 53,55ab
ES32 6,24c 10,50b 14,79a 19,95a 26,16a 32,45a 37,57a 42,05a 47,30a 52,17a 57,27a
Campuran 6,46bc 12,17ab 16,07a 20,61a 27,13a 32,53a 36,96a 41,71a 45,80a 49,48ab 53,71ab
Lampiran 3 Tabel pengaruh waktu inokulasi terhadap perkembangan tinggi tanaman cabai
Tinggi Tanaman pada t MST b) Waktu
Inokulasi
(MST) 5 6 7 8 9 10 11 12 13 14 15
3 6,05b 12,08ab 16,98a 20,88a 25,77a 30,58b 34,46b 38,53b 43,22b 46,88b 51,31b
5 7,00a 11,12b 14,68b 19,72a 26,68a 33,39a 38,73a 43,27a 47,88a 51,49a 55,93a
6 7,49a 12,15a 15,92ab 20,53a 26,97a 31,57ab 36,14b 39,68b 44,09b 48,59ab 51,81b
a)
Kontrol = tanpa bakteri; BG25= B. polymixa BG25; SB3 = B. subtilis SB3; PG01 = P. fluorescens PG01; ES32 = P. fluorescens ES32 (lampiran 2)
b)
angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbedanyata pada α = 0,05.
38
Lampiran 4 Tabel pengaruh perlakuan bakteri terhadap masa inkubasi penyakit
Perlakuan Masa Inkubasi (hari) a)
Kontrol 8,14 b B
BG25 9,28 ab AB
SB3 9,30 a AB
PG01 9,69 a A
ES32 9,61 a A
Campuran 9,14 ab AB
a) angka yang diikuti dengan huruf kecil dan huruf kapital yang sama dalam satu kolom
berturut-turut tidak berbedanyata pada α = 0,05 dan α = 0,01..
Lampiran 5 Tabel waktu inokulasi virus terhadap masa inkubasi penyakit
Waktu Inokulasi (MST) Masa Inkubasi (hari) a)
3 8,93a A
5 9,29a A
6 9,36a A
a)
39
Lampiran 6 Tabel pengaruh perlakuan bakteri dan waktu inokulasi terhadap masa inkubasi
Masa Inkubasi (hari) pada waktu inokulasi virus t MST b) Perlakuan a)
3 5 6
Kontrol 8,00 a 8,75 a 7,67 a
BG25 9,00 a 9,59 a 9,25 a
SB3 9,17 a 9,17 a 9,58 a
PG01 9,08 a 9,92 a 10,08 a
ES32 9,83 a 9,50 a 9,50 a
Campuran 8,49 a 8,84 a 10,08 a
a)
Kontrol = tanpa bakteri; BG25= B. polymixa BG25; SB3 = B. subtilis SB3; PG01 =
P. fluorescens PG01; ES32 = P. fluorescens ES32
b)
angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbedanyata pada