VEABILITAS Trichoderitta Izavzianunz Rifai PADA BEBERAPA
JENIS SERASAH DAN TANAH
DARUSSALIM
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan dengan sebenamya bahwa tesis yang berjudul, Viabilitas Triclzoderitta harziartunz Rifai pada Beberapa Jenis
Serasah dan Tanah adalah hasil karya saya dengan arahan Komisi Pembimbing
dan belum pernah digunakan untuk memperoleh gelar sejenis pada perguman tinggi manapun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan secara jelas dalam teks dan dicanturnkan dalam Dafiar Pustaka di bagian akhir eesis ini.
Bogor, Agustus 2009
ABSTRACT
DARUSSALIM. Viability of Trichoderma harzianum Rifai on Several Kinds of Leaf Litters and Soils. Under direction of LISDAR I. SUDIRMAN and OKKY SETYAWATI DHARMAPUTRA.
Trichoderma harzianum have been extensively studied as antagonistic fungal agent againts several plant pathogenic fungi caused by soil-borne fungi. In the soil, the leaf litter residues affected of fungal viability. This research was aimed to study the effect of four groups of media on viability T. harzianum. The first group was leaf litters of acacia, onion, and peanut, respectively; the second group was the soils containing decomposed leaf litters of each plant of the first group; the third group was the mixtures of each media of both group above (1:1), and the fourth group was natural top soil as a control. Each plastic bag filled with 0.5 kg of each medium was inoculated with ten ml of T harzianum conidia (10~1ml of concentration) and incubated in open area for 8 weeks. Statistically, the results showed that all treatments and control was significantly affected the fungal viability in field, but they did not significantly reduce the abilities of growth, conidial production and germination of T. harzianum on Potato Sucrose Agar medium. The lowest number of colony was found on peanut's leaf litters. The decreasing number of colony was found on all treatments and control during eight weeks of incubation. The result analysis showed that carbon concentration, nitrogen concentration, carbon to nitrogen ratio, lignin and cellulose concentration on all treatments and control were negative correlation with T. harzianunz population. It is suggested that T. harzianurn can be used as potential biofungicide in the soils containing either those leaf litters or composts except peanut's leaf litters.
DARUSSALIM. Viabilitas Trichoderma harzianum Rifai pada Beberapa Jenis Serasah dan Tanah. Dibimbing oleh LISDAR I. SUDIRMAN dan OKKY SETYAWATI DHARMAPUTRA.
Dalam upaya pengendalian berbagai penyakit pada tanaman hortikultura maupun hutan tanaman industri terutama yang disebabkan cendawan patogen tanaman, petani menggunakan pestisida kimia (termasuk fungisida) yang berlebihan dan kurang bijaksana. Hal ini dapat menimbulkan pengaruh negatif terhadap keseimbangan ekosistem, kesehatan manusia, serta dapat meningkatkan biaya produksi. Oleh karena itu perlu dicari altematif teknologi pengendalian cendawan patogen tanaman untuk menekan penggunaan fungisida kimia. Altematif pengendalian yang dapat dikembangkan adalah penggunaan cendawan antagonis sebagai agens pengendali hayati. Salah satu cendawan antagonis yang dilaporkan efektif menekan berbagai cendawan patogen terbawa tanah adalah T. harziantrm. Di lapangan, persistensi T. harzianum masih rendah dan hasilnya tidak konsisten setelah diaplikasikan, sehingga menjadi faktor pembatas dalam penggunaannya sebagai agens pengendali hayati. Hal ini terjadi karena perkembangan populasinya dipengamhi berbagai faktor lingkungan seperti kelembaban, suhu, pH, aplikasi pestisida, jenis sisa tanaman, kandungan bahan organik, senyawa inetabolit tanaman, dan keberadaan mikroorganisme lain di dalam tanah. Umumnya penelitian tentang pengaruh faktor lingkungan lebih banyak dilakukan pada skala laboratorium, sedangkan di lapangan seperti pengaruh serasah tanaman terhadap viabilitas T. harzianum beluln banyak dilaporkan.
Penelitian dilakukan sejak bulan Januari 2009 sampai dengan April 2009 di Laboratorium Mikrobiologi dan Biokimia Pusat Penelitian Studi Havati dan Bioteknologi (PPSHB) ~nstitut Pertanian Bogor (IPB) Dramaga Bogor. Pkrlakuan menggunakan rancangan acak lengkap (RAL) terdiri dari sepuluh perlakuan - - deng& masing-masing tiga ulangan. Media perlakuan dikelokpokkai menjadi empat kelompok. Kelompok pertama ( K1) terdiri dari jenis serasah: akasia (SA), bawang merah (SB), dan kacang tanah (SK). Kelompok kedua (K2) yaitu tanah yang mengandung masing-masing jenis serasah pada K1 yang telah terdekomposisi yaitu akasia (TAD), bawang merah (TBD), dan kacang tanah (TKD). Kelompok ketiga (K3) yaitu campuran masing-masing media K1 dan K2 dengan perbandingan 1:l. Kelompok keempat (TK) adalah tanah alami sebagai kontrol. Media perlakuan dimasukkan ke dalam kantong plastik polybag (0.5 kg) dan diinokulasi sebanyak 10 in1 konidium T. harzianurn umur 6 hari (konsentrasi 1 06/ml) dan diinkubasikan selama 8 minggu di lapangan terbuka.
PKb dilakukan pada suspensi konidium yang telah diinkubasikan selama 24, 48, 72, dan 96 jam dengan bantuan hemasitometer di bawah mikroskop. Penghitungan dilakukan pada 10 bidang pandang berdasarkan metode Sudirman et al. (2008). Parameter yang digunakan untuk mengetahui dekomposisi yang terjadi pada masing-masing media perlakuan adalah penentuan kandungan C-organik, N-total, rasio C:N, lignin, dan selulosa pada minggu ke-1, ke-5, dan ke-8 setelah introduksi T. harzianum. Kandungan C-organik ditentukan dengan metode Walkey dan Black, N-total dengan metode Kjeldahl, kandungan lignin dan selulosa dengan metode Van Soest.
Data dianalisis menggunakan program SPSS V.13 for Windows ~nelalui One-way Analysis of Varians (ANOVA). Apabila terdapat perbedaan yang nyata dari perlakuan maka dilanjutkan dengan uji jarak berganda Duncan Multiple Range Test (DMRT) pada taraf kepercayaan 95%.
Hasil penelitian menunjukkan bahwa semua media perlakuan pada K1, K2, K3, dan TK berpengamh terhadap populasi T. harzianum. Rata-rata populasi T. harzianum pada K1, K2, dan K3 lebih rendah dibandingkan pada TK. Pada minggu ke-1, populasi T. harziantlm pada K1 berkisar 5.33-29.83 x lo3 kolonilg (rata-rata 18.28 x lo3 kolonilg), K2 24.17-32.50 x lo3 kolonilg (rata-rata 29.06
x
1o3
kolonilg), K3 13.1 7-26.50 x 10' kolonilg (rata-rata 21.44 x 10' kolonilg), dan TK dengan rata-rata 43 x lo3 kolonilg. Setelah delapan minggu inkubasi, rata-rata populasi T. harzianum inengalami penurunan pada K1, K2, dan K3 masing- masing sebesar 13.16 x lo3, 14 x lo3, dan 12.11 x lo3 kolonilg. Sebaliknya, rata- rata populasi T. harzianum pada TK relatif tinggi yaitu 25.33 x lo3 kolonilg setelah delapan minggu inkubasi. Hasil analisis ragam menunjukkan bahwa semua media perlakuan berpengaruh sangat nyata (P < 0.01) terhadap fluktuasi populasiT. harzianum pada minggu ke-1 sampai minggu ke-7, tetapi tidak berpengamh nyata (P > 0.05) terhadap populasi T. harziantrm pada minggu ke-8.
Diameter koloni isolat M1 dari semua media perlakuan pada K1, K2, dan K3 yang ditumbuhkan pada media PSA pada hari ke-1 pengamatan (HI) antara 13.17-16.17 mm, hari ke-2 (H2) antara 34.83-47.67 mm, hari ke-3 (H3) antara 64.67-84.17 mm, sedangkan TK pada HI, H2, dan H3 masing-masing 17 m n , 51 mm, dan 85.5 mm. Diameter koloni isolat M5 pada H1 antara 30.67-33.5 nun, H2 antara 65.5-70.5 mm, H3 antara 85.83-87 mm, sedangkan TK pada HI, H2, dan H3 masing-masing 31.5 mm, 66.5 mm, dan 87 mm. Diameter koloni isolat MS pada HI antara 26.33-36 mm, H2 antara 64.67-72.67 mm, H3 sebesar 87 mm, sedangkan TK pada HI, H2, dan H3 masing-masing 26.83 mm, 64.67 mm, dan 87 mm. Berdasarkan data tersebut menunjukkan bahwa rata-rata diameter koloni isolat M1 pada H1 dan H2 lebih rendah dibandingkan diameter koloni isolat M5 dan M8 pada H1 dan H2. Sementara itu, rata-rata diameter koloni isolat MI pada H3 relatif tidak berbeda dengan diameter koloni isolat M5 dan M8 pada H3. Semakin lama isolat (M5 dan M8) berada di media perlakuan, maka pertumbuhannya semakin baik ketika ditumbuhkan pada media PSA. Efek media perlakuan hanya berpengaruh pada isolat MI. Jadi secara umum pengamh media perlakuan TK tidak berbeda dengan pengamh media perlakuan lainnya terhadap diameter koloni isolat MI, M5, dan M8 pada media PSA.
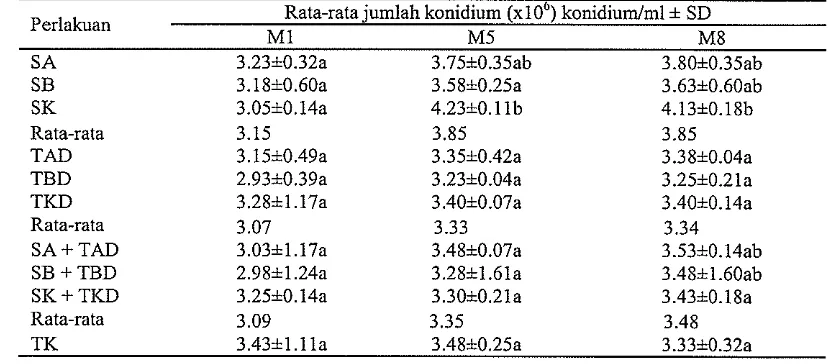
sebesar 3.48 x lo6 konidiudg. Jk isolat M8 antara 3.25-4.13 x
lo6
konidiudg, sedangkan pada TK sebesar 3.33 x lo6 konidiudg. Jadi pengaruh media perlakuan TK tidak berbeda dengan pengaruh media perlakuan lainnya terhadap Jk isolat MI, M5, dan M8 pada media PSA.PKb isolat M1 semua media perlakuan pada K1,
K2,
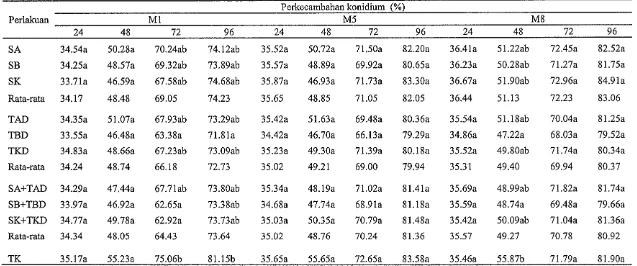
dan K3 yang diinkubasikan dalam akuades setelah 24 jam inkubasi antara 33.71-34.83%, 48 jam antara 46.48-51.07%, 72 jam antara 62.65-70.24%, dan 96 jam antara 71.81-74.68%, sedangkan pada TK setelah 24, 48, 72, dan 96 jam inkubasi masing-masing sebesar 35.17%, 55.23%, 75.06%, dan 81.15%. PKb isolat M5 setelah 24 jam inkubasi antara 34.42-35.87%, 48 jam antara 46.70-51.63%, 72 jam antara 66.13-71.73%, dan 96 jam antara 79.29-83.30%, sedangkan pada TK setelah 24, 48, 72, dan 96 jam inkubasi masing-masing sebesar 35.65%, 55.65%, 72.65%, dan 83.58%. PKb isolat M8 setelah 24 jam inkubasi antara 34.86- 36.67%, 48 jam antara 47.22-51.90%, 72 jam antara 68.03-72.96%, dan 96 jam antara 79.52-84.91%, sedangkan pada TK setelah 24, 48,72, dan 96 jam inkubasi masing-masing 35.46%, 55.87%, 71.79%, dan 81.90%. Jadi pengaruh media perlakuan TK tidak berbeda dengan pengaruh media perlakuan lainnya terhadap PKb isolat M1, M5, dan M8 pada media PSA.Berdasarkan hasil uji korelasi Pearson menunjukkan bahwa kandungan C-organik, N-total, rasio C:N, kandungan lignin dan selulosa semua media perlakuan berkorelasi negatif terhadap populasi T. harzianunt pada minggu ke-1, ke-5, dan ke-8.
OHak Cipta milik IPB tahun 2009 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh katya tulis ini tanpa nzencantzrnzkan atau nzenyebutkan sumbernya. Pengutipan hanya untulc kepentingair pendidikan, penelitian, penulisan katya ilmiah, penyusunan laporan, penulzsan iiritik atazr tinjazran suatu masalah; dan pengutipan tersebut tidalc merugilcan kcepentingan yang wajar IPB.
VIABILITAS
Trichodevnza harzianum
Rifai PADA BEBERAPA
JENIS SERASAH DAN TANAH
DARUSSALIM
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Mikrobiologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
LEMBAR PENGESAHAN
Judul Tesis : Viabilitas Trichoderma harzianum Rifai pada Beberapa Jenis Serasah dan Tanah
Nama : Darussalim NRP : G351070191
Disetujui Komisi Pembimbing
/ .
Dr. Ir. Llsdal I. Sudirman Ketua
/
Dr. Okkv Setvawati Dharma~utra Anggota
Diketahui :
. .-
Ketua Mayor Mikrobiologi
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Judul yang dipilih dalam penelitian ini ialah Viabilitas Trichoderma harzianum Rifai pada Beberapa Jenis Serasah dan Tanah. Penelitian dilaksanakan dari bulan Januari 2009 sampai dengan bulan April 2009.
Penulis menyampaikan penghargaan dan ucapan terima kasih yang sebesar- besarnya temtama kepada Pembimbing, yaitu Dr. Ir. Lisdar I. Sudirman dan Dr. Okky Setyawati Dharmaputra yang telah banyak memberikan arahan, bimbingan dan saran selama penulis menempuh studi S2. Terima kasih penulis sampaikan kepada Prof. Dr. Ir. Meity S. Sinaga selaku Penguji Luar Komisi yang telah banyak memberikan koreksi dan arahan untuk perbaikan tesis.
Terima kasih yang sebesar-besarnya penulis sampaikan kepada Departemen Agama Republik Indonesia yang telah mengadakan kerjasama program beasiswa pascasarjana dengan Institut Pertanian Bogor. Selanjutnya ungkapan telima kasih juga disampaikan kepada Ayahanda (almarhum) H. Rajali, Ibunda Hj. Siti Rahrnah, Ayahanda dan Ibunda mertua Sofyan Dt. Sinarosati dan Ridasmar Dahlan, Isteriku Zulhima S.Pd dan anak-anak tercinta Muhammad Raihan Pratama Salim dan Salsabila Salim atas do'a, pengorbanan dan kasih sayangnya. Serta teman-teman dan semua pihak yang tidak dapat saya sebutkan satu persatu yang telah membantu dalam penyelesaian penulisan tesis penelitian ini. Semoga budi baiknya mendapat imbalan dari Allah SWT.
Penulis menyadari bahwa tesis ini masih jauh dari sempuma, oleh karena itu kritik dan saran sangat diharapkan. Demikian karya iliniah ini kami susun semoga dapat bermanfaat bagi semua pihak yang berkepentingan.
Bogor, Agustus 2009
RIWAYAT HIDUP
Penulis dilahirkan di Desa Durian pada tanggal 8 Mei 1972 dari ayah (Almarhum) H. Rajali dan ibu Hj. Siti Rahmah. Penulis nlerupakan putra ketujuh dari sembilan bersaudara.
Pendidikan dasar penulis selesaikan di Sekolah Dasar Inpres 105341 Desa Durian tahun 1985, Sekolah Lanjutan Tingkat Pertama di SMP Negeri Pantai Labu tahun 1988, kemudian penulis melanjutkan ke Sekolah Menengah Atas di SMA Negeri Lubuk Pakam dan lulus pada tahun 1991. Pada tahun yang sama, penulis melanjutkan pendidikan di Jurusan Tadris Biologi, Fakultas Tarbiyah Institut Agama Islam Negeri (IAIN) Sumatera Utara dan lulus pada tahun 1997. Setelah menyelesaikan pendidikan sarjana, penulis menjadi guru di Madrasah Aliyah Negeri (MAN) 2 Medan Sumatera Utara sampai sekarang. Selanjutnya pada tahun 2001, penulis mendapatkan kesempatan beasiswa untuk melanjutkan pendidikan di Jurusan Pendidikan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam (MIPA) Universitas Negeri Medan (UNIMED) Medan dan lulus pada tahun 2004. Pada bulan Juli tahun 2007 penulis mendapat beasiswa melanjutkan pendidikan pada Program Pascasarjana Institut Pertanian Bogor dengan mengambil Mayor Mikrobiologi dan dinyatakan lulus pada tanggal 14 Agustus 2009.
Bogor, Agustus 2009
DAFTAR
IS1
DAFTAR TABEL
...
...
DAFTAR GAMBAR
...
DAFTAR LAMPIRAN
PENDAHULUAN
...
Latar Belakang...
Tujuan...
Manfaat
...
...
TINJAUAN PUSTAKA
.
...
Biologi T harzianum
...
Potensi T. harzianum sebagai Agens Pengendali Hayati
Persistensi T. harzianum
...
...
Dekomposisi Serasah
...
BAHAN DAN METODE
...
Waktu dan Tempat
Penyiapan Media Perlakuan
...
Perbanyakan T.
harzianum...
...
Introduksi Konidium T harzianum ke dalam Media Perlakuan
...
Populasi T harzianum dari Masing-masing Media Perlakuan
Pertumbuhan. Jumlah Konidium. dan Persentase Perkecambahan Konidium T. harzianum
...
Penentuan Kandungan C.organik. N.total. Rasio C.N. Lignin. dan Selulosa...
...
Analisis Data
HASIL DAN PEMBAHASAN
...
...
Populasi T
.
harzianum pada Media Perlakuan...
Pertumbuhan T harzianum pada Media PSA
...
Produksi Konidium T
.
harzianum pada Media PSAPerkecambahan Konidium T Izarzianum
...
...
Kandungan C.organik, N.total. dan Rasio C:N
...
Kandungan Lignin-dan Selulosa
SIMPULAN DAN SARAN
...
DAFTAR PUSTAKA...
xi xii
...
DAFTAR
TABEL
Halaman
Populasi T. harzianum (x103 kolonitg) pada keeinpat kelompok media
yang dihitung setiap minggu selama inkubasi
...
16 Hasil analisis ragam pengaruh semua media perlakuan terhadap...
populasi T. harzianum selama 8 minggu inkubasi 17 Pertumbuhan isolat minggu ke-1 (MI), ke-5 (MS), dan ke-8 (M8) dari
semua media perlakuan pada media PSA yang diamati pada hari ke-1
(HI), ke-2 (H2), dan ke-3 (H3)
...
22 Produksi konidium isolat MI, M5, dan M8 dari semua mediaperlakuan pada media PSA
...
23 Hasil analisis ragam pengaruh semua media perlakuan terhadapproduksi konidium isolat MI, M5, dan M8 pada media PSA
...
24 Perkecambahan konidiuin isolat MI, M5, dan M8 dari semua mediaperlakuan setelah 24,48, 72, dan 96 jam inkubasi dalam akuades
...
25 Hasil analisis ragam pengaruh semua media perlakuan terhadap PKbisolat MI, M5, dan M8 setelah 24,48, 72, dan 96 jam inkubasi dalam
akuades
...
26 Kandungan C-organik, N-total, rasio C:N, kandungan lignin danxii
DAFTAR GAMBAR
Halaman
Koloni T. harzianum setelah 6 hari inkubasi pada media PSA pada
...
suhu kamar dan foto mikrograf T. harzianum (400 x) 5 Populasi T. harzianum pada berbagai kelompok perlakuan setelah
...
diinkubasi beberapa minggu ke dalam media perlakuan 17 Populasi T. harzianum pada berbagai serasah setelah diinkubasi
...
beberapa minggu ke dalam media perlakuan 18 Populasi T. harziantlm pada berbagai tanah yang mengandung
serasah yang telah terdekomposisi setelah diinkubasi beberapa
minggu ke dalam media perlakuan
...
19 Populasi T. harzianum pada berbagai campuran dari masing-masing...
media kelompok pertama dan kedua 19
Morfologi konidium T. harzianum, konidium yang mulai
berkecambah setelah 24,48,72, dan 96 jam, dan tabung kecambah 27 Kandungan lignin semua inedia perlakuan pada minggu ke-1, ke-5,
...
dan ke-8 setelah diintroduksi T. harzianum 31 Kandungan selulosa semua media perlakuan pada minggu ke-1,
...
DAFTAR
LAMPIRAN
Halaman
1 Komposisi media Potato Sucrose Agar (PSA)
...
40 2 Temperatur, kelembaban relatif, dan curah hujan harian (mm)...
pada bulan Januari, Februari, dan Maret tahun 2009 41 3 Kandungan C-organik dan N-total semua media perlakuan pada
...
minggu ke-1, ke-5, dan ke-8 setelah introduksi T. harzianum 44 4 Nilai rasio C:N (%) semua media perlakuan pada minggu ke-1,
...
ke-5, dan ke-8 setelah introduksi T. harzianum 45 5 Hasil analisis kandungan lignin (%) dan selulosa (%) semua
media perlakuan pada minggu ke-1, ke-5, dan ke-8 setelah introduksi T. harzianum
...
46 6 pH semua media perlakuan sejak tninggu ke-1 sampai dengan...
minggu ke-8 setelah introduksi T. harziantim 47 7 Populasi T. harzianum (x10' koloniig) pada keempat kelompok
...
media yang dihitung setiap minggu selaina inkubasi 48 8 Pertumbuhan isolat MI, M5, dan M8 dari semua media perlakuan
pada media PSA yang diamati pada hari ke-1 (HI), ke-2 (H2),
...
dan ke-3 (H3) 49
9 Jumlah konidium isolat MI, M5, dan M8 dari seinua media perlakuan pada media PSA
...
52 10 Perkecambahan konidium (%) isolat MI, M5, dan M8 dari semuamedia perlakuan pada media PSA setelah 24, 48, 72, dan 96 jam
inkubasi dalam akuades
...
55 11 Analisis ragam pengaruh semua media perlakuan terhadap...
populasi T. harzianum pada minggu ke-1 inkubasi 59 12 Analisis ragam pengaruh semua media perlakuan terhadap
...
populasi T. harzianum pada minggu ke-2 inkubasi 59 13 Analisis ragam pengaruh semua media perlakuan terhadap
...
14 Analisis ragam pengaruh semua media perlakuan terhadap populasi T. harzianum pada minggu ke-4 inkubasi
...
15 Analisis ragam pengaruh semua media perlakuan terhadappopulasi T. harzianum pada minggu ke-5 inkubasi
...
16 Analisis ragam pengaruh semua media perlakuan terhadap...
populasi 7: harzianum pada minggu ke-6 inkubasi
17 Analisis ragam pengaruh semua media perlakuan terhadap populasi T. harzianum pada minggu ke-7 inkubasi
...
18 Analisis ragam pengaruh semua media perlakuan terhadappopulasi T. harzianum pada minggu keg inkubasi
...
19 Analisis ragam pengaruh semua media perlakuan terhadappertumbuhan isolat MI pada media PSA pada hari ke-1 (HI)
...
20 Analisis ragam pengamh semua media perlakuan terhadappertumbuhan isolat MI pada media PSA pada hari ke-2 (H2)
...
21 Analisis ragam pengaruh semua media perlakuan terhadappertumbuhan isolat M1 pada media PSA pada hari ke-3 (H3)
...
22 Analisis ragam pengaruh semua media perlakuan terhadap pertumbuhan isolat M5 pada media PSA pada hari ke-1 (HI)...
23 Analisis ragam pengaruh semua inedia perlakuan terhadappertumbuhan isolat M5 pada media PSA pada hari ke-2 (H2)
...
24 Analisis ragam pengaruh semua inedia perlakuan terhadappertumbuhan isolat M5 pada media PSA pada hari ke-3 (H3)
...
25 Analisis ragam pengaruh semua media perlakuan terhadappertumbuhan isolat M8 pada media PSA pada hari ke-1 (HI)
...
26 Analisis ragam pengamh semua media perlakuan terhadap...
pertumbuhan isolat M8 pada media PSA pada hari ke-2 (H2) 27 Analisis ragam pengaruh semua media perlakuan terhadap
pertumbuhan isolat M8 pada media PSA pada hari ke-3 (H3)
...
28 Analisis ragam pengaruh semua media perlakuan terhadap jumlah29 Analisis ragam pengaruh semua media perlakuan terhadap
...
jumlah konidium isolat M5 pada media PSA
30 Analisis ragam pengaruh semua media perlakuan terhadap
...
jumlah konidium isolat M8 pada media PSA
31 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M1 setelah 24 jam inkubasi 32 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M1 setelah 48 jam inkubasi 33 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat MI setelah 72 jam inkubasi 34 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M1 setelah 96 jam inkubasi 35 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M5 setelah 24 jam inkubasi 36 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M5 setelah 48 jam inkubasi 37 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M5 setelah 72 jam inkubasi 38 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M5 setelah 96 jam inkubasi 39 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M8 setelah 24 jam inkubasi 40 Analisis ragam penga~uh semua media perlakuan terhadap
...
perkecambahan konidium isolat M8 setelah 48 jam inkubasi 41 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M8 setelah 72 jam inkubasi 42 Analisis ragam pengaruh semua media perlakuan terhadap
...
perkecambahan konidium isolat M8 setelah 96 jam inkubasi 43 Uji korelasi Pearson C-organik, N-total, rasio C:N, kandungan
lignin dan selulosa semua media perlakuan terhadap populasi T.
xvi
44 Uji korelasi Pearson C-organik, N-total, rasio C:N, kandungan lignin dan selulosa sernua media perlakuan terhadap populasi T.
harzianum pada minggu ke-5
...
65 45 Uji korelasi Pearson C-organik, N-total, rasio C:N, kandunganlignin dan selulosa semua media perlakuan terhadap populasi T
PENDAHULUAN
Latar Belakang
Dalam upaya pengendalian berbagai penyakit pada tanaman hortikultura maupun hutan tanaman industri terutama yang disebabkan cendawan patogen tanaman, petani menggunakan pestisida kimia yang berlebihan untuk mengendalikannya. Adiyoga et al. (1997) melaporkan bahwa 62-93% petani responden melakukan penyemprotan pestisida (termasuk fungisida) secara rutin 3-7 hari sekali dan inelakukan pencampuran 2-4 jenis pestisida. Penggunaan fungisida kimia yang berlebihan dan kurang bijaksana akan menimbulkan pengaruh negatif terhadap keseimbangan ekosistem dan kesehatan manusia seperti berkembangnya patogen yang resisten terhadap fungisida, kemunglunan terjadinya penurunan kualitas genetik dalam populasi, dan berbahaya bagi organisme yang bukan sasaran (Benitez et al. 2004), pencemaran lingkungan, serta dapat meningkatkan biaya produksi mencapai 30-50% dari total biaya produksi (Koster 1990). Oleh karena itu perlu dicari altematif teknologi pengendalian cendawan patogen tanaman untuk menekan penggunaan fungisida kimia.
Altematif pengendalian yang dapat dikembangkan adalah penggunaan mikroorganisme seperti cendawan antagonis sebagai agens pengendali hayati (biological control agents). Sitepu (1993) melaporkan bahwa dalam konsep pengendalian hama terpadu (PHT), pemanfaatan agens pengendali hayati merupakan teknik pengendalian yang perlu diutamakan, yang dalam aplikasinya hams kompatibel dengan pengendalian lainnya. Selain itu penggunaan mikroorganisme untuk pengendalian hayati relatif lebih aman, tidak terakumulasi dalam rantai makanan, pemakaiannya tidak berulang-ulang, dapat dikombinasikan dengan teknik pengendalian lain yang bukan bahan kimia dan tidak terjadi resistensi pada organisme sasaran (Suwanto & Tjahjoleksono 1993).
luas, terdapat dimana-mana, mudah diisolasi dari tanah, kayu membusuk dan organ lain dari bahan organik tanaman, mudah dikulturkan, pertumbuhannya cepat, menyerang patogen tanaman pada kisaran yang luas, dan jarang patogenik pada tanaman (Howell 2003).
Salah satu spesies Trichoderma yang dilaporkan efektif sebagai agens pengendali hayati patogen terbawa tanah pada berbagai tanaman adalah Trichoderma harzianum (Elad et al. 1980). Aplikasinya telah banyak dilaporkan pada skala laboratorium, sedangkan di lapangan belum banyak dilaporkan. Trichoderma harzianum dapat inengendalikan cendawan patogen tanaman seperti Fusarium oxysporum dan Alternaria porri pada bawang merah (Imtiaj & Lee 2008), Sclerotium rolfsii dan Aspergillt~sj7avus pada kacang tanah (Dharmaputra et al. 2003; Kuswinanti 2006; Gachomo & Kotchoni 2008), d m Rhizoctonia solani Kuhn. pada akasia (Umarella 2006).
Mengingat persistensi T. harzianum yang sangat rendah dan tidak konsisten, serta data pengaruh serasah tanaman terhadap viabilitas T. harzianum belum banyak dilaporkan, maka perlu dilakukan penelitian tentang viabilitas
T.
harzianum pada beberapa jenis serasah tanaman seperti akasia, bawang merah,kacang tanah, dan tanah yang mengandung serasah yang telah terdekomposisi dari masing-masing tanaman tersebut.
Tujuan
Penelitian ini bertujuan untuk mempelajari viabilitas T. harzianum TBPH pada beberapa jenis serasah tanaman seperti akasia, bawang merah, kacang tanah, dan tanah yang mengandung serasah yang telah terdekomposisi dari masing- masing tanaman tersebut.
Manfaat
Hasil penelitian ini diharapkan dapat inemberikan infomasi tentang viabilitas
T.
harzianum TBPH serta waktu yang tepat dan efektif ketikaTINJAUAN PUSTAKA
Biologi
T.
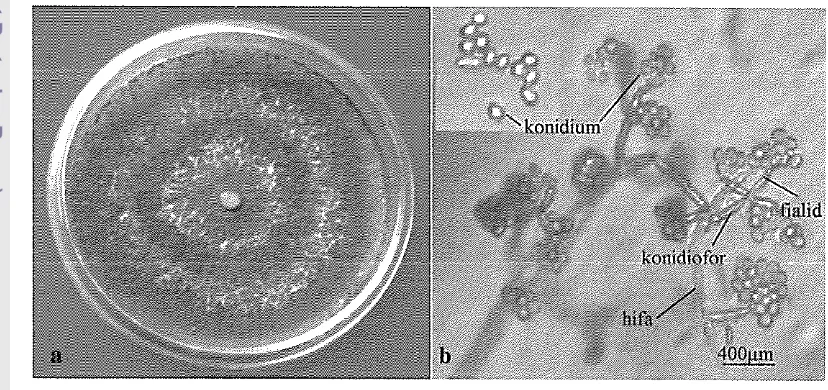
knrziaizzc~~zTrichoderma harzianum termasuk ke dalam kelas Deuteromycetes, famili Moniliaceae, ordo Moniliales (Alexopolous & Mims 1996). Trichoderma harzianum termasuk imperfect fungi (cendawan tidak sempurna), tingkat anamorfnya Hypocrea dan tidak mengenal tingkat teleomorf (Barnet & Hunter 1999; Monte 2001). Koloni T. harzianum yang ditumbuhkan pada media Potato Sucrose Agar (PSA) memiliki pertumbuhan yang cepat, setelah 3-4 hari inkubasi pada suhu kamar (27 OC) mampu memenuhi cawan Petri (7-9 cm), pertumbuhan hifa datar dan membentuk daerah melingkar yang benvarna hijau terang sampai gelap, kemudian koloni menjadi seperti beludru sampai menjadi tepung dan membentuk cincin konsentris (Gambar la). Pada media Czapek Yeast Extract Agar (CYA) dan Malt Extract Agar (MEA) pada umumnya koloni tumbuh memenuhi cawan Petri, bentuk garis melingkar jelas dengan miselium benvarna putih sampai kuning, sedangkan koloni pada media 25% Glycerol Nitrate Agar (G25N) tumbuh lambat yaitu berdiameter kurang dari 5 mm (Pitt & Hocking 1997).
Trichoderma harzianum memiliki hifa bersekat, hercabang, dindingnya halus, hialin, berukuran 1.5-12 pm, konidiofor tegak, bercabang, bentuknya verticillate, menyangga fialid, fialid ampulliform (berbentuk botol), berukuran 3.5-7.5
x
2.5-3.8 pm. Konidium kecil, berbentuk bulat atau lonjong, berukuran 2.7-3.5 x 2.1-2.6 pm, dindingnya halus, benvarna subhialin sampai hijau muda dan berkumpul pada bagian ujung fialid (Gambar lb) (Rifai 1969; Pitt & Hocking1997; Fisher & Cook 1998; Gams & Bissett 1998).
Pada kondisi laboratorium, T. harzianum tumbuh pada pH 2-7 dan optimum pH 4 dengan suhu sekitar optimum 30 OC dan maksimum kurang dari 36 OC
(Domsch et al. 1980; Kredics et al. 2003). Sebaliknya, pada suhu 37 OC tidak ada
keberadaan mikroorganisme lain di dalam tanah (Gardner & Storey 1977; Papavizas 1985; Kredics et al. 2003).
Pertumbuhan
T.
harzianum dipengaruhi oleh berbagai faktor lingkungan di dalam tanah, seperti pH tanah yang tinggi dapat mendegradasi beberapa antibiotik yang dihasilkan olehT.
harzianum (Delgado et al. 2000), suhu yang tingg menghambat pertumbuhan hlfa (Pitt & Hocking 1997), dan logam berat seperti besi, seng, nikel, dan kobal bersifat toksik pada konsentrasi yang tinggi (Benitez et al. 2004). Triclzoderma harzianum toleran terhadap pestisida hmia sehingga dapat dikombinasikan dalam pengendalian patogen tanaman (Benitez et al. 2004), dan toleran terhadap beberapa senyawa metabolit sekunder tanaman (Klein & [image:23.595.97.511.356.551.2]Eveleigh 1998).
Gambar 1 Koloni
T.
harzianum setelah 6 hari inkubasi pada media PSA pada suhu kamar (a) dan foto mikrograf % lzarzianum (400x) (b) (Foto: Darussalim 2009)Potensi
T.
Izarzianum Sebagai Agens Pengendali Hayatiagens pengendali hayati (Pal & Gardener 2006). Menurut Monte (2001) agens pengendali hayati bekerja dengan beberapa cara yaitu (1) agens pengendali hayati tumbuh sangat cepat atau menggunakan sumber nutrisi secara efisien dibandingkan patogen, (2) melepaskan suatu produk yang dapat menekan atau membunuh patogen, (3) menginduksi ketahanan tanaman terhadap patogen, dan (4) membunuh patogen secara langsung (parasitisme).
Aplikasi pengendalian hayati di lapangan dapat dilakukan melalui (1) praktik budidaya dan pengelolaan habitat dengan menciptakan suatu lingkungan yang sesuai bagi organisme antagonis, (2) pemuliaan tanaman untuk meningkatkan resistensi tanaman terhadap patogen, dan (3) introduksi massal satu atau lebih organisme antagonis (Papavizas 1985; Cook & Baker 1988).
Trichoderma harzianzrm merupakan agens pengendali hayati yang efektif terhadap berbagai cendawan patogen terbawa tanah antara lain S. rolfsii dan R. solani (Elad et al. 1980), Verticilliunz dahliae (Chet & Inbar 1994), Drechslera sorokiniana, Fusarium moniliforme, Fusarium cttlmortrnz dan Gaeumannomyces graminis var. tritici (Kucuk & Kivanc 2002), A. flavus (Dharmaputra et al. 2003), Alternaria alternata (Sempere & Santamarina 2007), Fusarium solani f. sp. phaseoli (Abeysinghe 2007), Sclerotinia sclerotiorum (Akrami et al. 2008), A. porri (Nur 2005; Imtiaj & Lee 2008), dan F. oqsportrm f. sp. ciceri (Jayalakshmi et al. 2009).
Mekanisme antagonis T. harzianum sangat kompleks dan bervariasi dalam menekan cendawan patogen dan meningkatkan pertumbuhan tanaman. Howell (2003) mengemukakan bahwa reaksi antara T. harzianum dengan berbagai inang cendawan mendasari perbedaan mekanisme antagonis. Mekanisme antagonis
T.
harzianum meliputi kompetisi (Delgado et al. 2004; Ozbay & Newman 2004), rnikoparasitisme (Limon et al. 1999), antibiosis (Howell 2003; Benitez et al. 2004), dan induksi ketahanan (Howell 2003).Kondisi kelaparan umumnya menyebabkan kematian mikroorganisme, terutama pada kondisi nutrisi yang sangat terbatas antara agens pengendali hayati dengan cendawan patogen. Trichoderma harziantlm mampu berkompetisi dengan cendawan patogen dalam pengambilan oksigen, air, nutrisi, dan ruang untuk kelangsungan hidup (Cook & Baker 1988). Tjamos et al. (1992) melaporkan
T. harzianum T35 mampu berkompetisi dengan F. oxysporum dalam pengambilan nutrisi dan berkolonisisasi di daerah i-izosfir.
Mikoparasitisme merupakan salah satu mekanisme antagonis T. harziantmz terhadap cendawan patogen. Prosesnya sangat kompleks ineliputi pengenalan terhadap hifa inang, penempelan, penetrasi pada dinding sel inang dengan menghasilkan enzim pendegradasi dinding sel seperti kitinase, glukanase, protease, dan terakhir membunuh sel inang patogen dengan cara mengambil isi hifa sebagai sumber nutrisi (Benitez et al. 2004). Trichoderma Izarziantcin ~nenghasilkan enzim Chit33 kitinase yang menghambat pertumbuhan R. solani (Limon et al. 1999). Enzim kitinase dan a-1,3-glukanase yang dihasilkan
T. harziantlm galur T24 menghainbat cendawan patogen tanaman S. rolfsii (El-Katatny et al. 2001). Mekanisme antagonis lainnya dari T. harziantlm terhadap cendawan patogen adalah antibiosis. Antibiosis merupakan proses sekresi senyawa antimikroba oleh cendawan antagonis nntuk menekan dan atau membunuh cendawan patogen (Schirmbock et al. 1994). Benitez et al. (2004) mengemukakan bahwa banyak isolat T. harzianum yang memproduksi antibiotik pyrone yang efektif menekan Gaeumannomyces graminis var. tritici.
dan chalcone synthase (CHS), yang meliputi biosintesis phytoalexins (HR
response), kitinase, dan glukanase (Benitez et al. 2004). Harrnan et al. (2004a) mengemukakan bahwa mekanisme induksi ketahanan berhubungan dengan mikoparasitisme dan antibiosis. Harrnan et al. (2004b) melaporkan bahwa aplikasi
T. harzianum T22 pada tanaman jagung dapat meningkatkan pertumbuhan tanaman secara signifikan, cendawan ini mampu menekan dan mengendalikan patogen pada akar dan daun tanaman melalui induksi ketahanan, mengubah komposisi mikroflora di daerah perakaran, meningkatkan penyerapan nutrisi, meningkatkan solubilitas nutrisi tanah, meningkatkan perkembangan akar, meningkatkan jumlah rambut akar, serta kedalaman perakaran. Selain itu, T. harzianum yang diaplikasikan pada tanaman lettuce mampu meningkatkan berat segar bibit dari 1.61 g/m2 inenjadi 15 g/m2 (Bal & Altintas 2008). Induksi ketahanan penting bagi tanaman agar tanaman resisten terhadap toksin yang dihasilkan oleh mikroflora tanah atau induksi akibat aktivitas manusia seperti fungisida, logam berat, dan sekresi oleh berbagai faktor seperti antibiotik dan enzim pendegradasi dinding sel.
Persistensi
T.
harzianu~~tKemampuan agens pengendali hayati untuk dapat bidup dan bertahan di lingkungan merupakan salah satu faktor penting dalam keberhasilan pengendalian hayati. Propagul cendawan antagonis yang meiniliki persistensi yang baik akan mempunyai peluang yang lebih besar untuk bisa kontak dan menimbulkan penyakit pada cendawan patogen terbawa tanah.
2003). Selain itu persistensi agens pengendali hayati juga bergantung pada metode dan strategi dalam inengintroduksikan dan mengaktifkan agens tersebut dalam berasosiasi dengan tanaman (Cook & Baker 1988).
Tanaman seperti akasia, bawang merah, dan kacang tanah adalah jenis-jenis tanaman yang mengandung bahan kimia yang berbeda, ha1 ini sangat besar pengaruhnya terhadap persistensi agens pengendali hayati. Bawang merah menghasilkan senyawa metabolit tanaman berupa senyawa isotiosianat yang terakumulasi dalam konsentrasi tinggi di lahan pertanaman (Inyang et al. 1999; Klingen et al. 2001). Sudirman et al. (2008) melaporkan bahwa serasah daun dan tanah yang mengandung serasah bawang merah yang telah terdekomposisi dapat menurunkan populasi dan inenghambat pertumbuhan Beauvevia bassiana (Bals.) Vuill.
Kacang tanah merupakan jenis tanaman yang banyak mengandung unsur nitrogen dan mampu bersimbiosis dengan bakteri Rhizobium dalam ha1 memfiksasi nitrogen. Unsur nitrogen merupakan unsur yang sangat dibutuhkan oleh semua mikroorganisme termasuk cendawan berfungsi dalam proses sintesis protein dalam sel. Sedangkan akasia merupakan tanaman hutan industri yang mengandung selulosa dan lignin yang sangat tinggi. Menurut Rohiani (1996) kandungan lignin pada serasah daun akasia umur 2 dan 3 tahun masing-masing sebesar 45.82% dan 45.85%, sedangkan kandungan selulosa masing-masing sebesar 51.36% dan 51.45%.
Dekomposisi Serasah
Serasah merupakan bahan organik yang berasal dari tumbuhan yang terdapat di atas permukaan tanah dan tersusun dari bahan-bahan yang telah mati (Spurr & Burton 1980 dalam Hilwan 1993). Pada dasarnya proses dekomposisi serasah melibatkan berbagai mikroorganisme tanah yang mampu mendaur ulang nutrisi ke dalam lingkungan (Kunito & Nagaoka 2009). Mikroorganisme tanah yang memilih peranan paling dominan sebagai dekomposer adalah cendawan (Moore & Landecker 1996).
Proses dekomposisi yang dilakukan cendawan mencakup proses pembusukan bahan organik yang dimulai dengan sekresi enzim ekstraseluler yang mampu menghidrolisis molekul kompleks berukuran besar menjadi molekul yang lebih kecil sehingga dapat dimanfaatkan oleh organisme lain (Irawan et al. 2008). Prosesnya dapat berlangsung beberapa bulan bahkan bertahun-tahun sanpai bahan organik tersebut menjadi humus, dan akhirnya inenjadi tanah (Dix
& webster 1995).
BAHAN DAN METODE
Waktu dan Ternpat
Penelitian dilakukan di Laboratorium Mikrobiologi dan Biokimia Pusat Penelitian Studi Hayati dan Bioteknologi (PPSHB) Institut Pertanian Bogor (IPB) Darmaga Bogor, dari bulan Januari 2009 sarnpai dengan April 2009. Untuk penentuan kandungan C-organik dan N-total dilaksanakan di Services Laborato~y SEAMEO BIOTROP Bogor, sedangkan penentuan kandungan lignin dan selulosa dilaksanakan di Laboratorium Ilmu dan Teknologi Pakan, Departemen Ilmu Nutrisi dan Teknologi Pakan Fakultas Petemakan IPB Darmaga Bogor.
Penyiapan Media Perlakuan
Perbanyakan
T.
harziaizuittIsolat TBPH adalah T. harzianum Rifai diperoleh dari Balai Penelitian dan Pengembangan PT Riau Andalan Pulp & Paper (RAPP) Riau, dan telah diidentifikasi sampai spesies oleh Dr. Okky Setyawati Dharmaputra berdasarkan pustaka acuan Rifai (1969). Biakan T. havzianum (diameter 3 mm) ditumbuhkan pada media Potato Sucrose Agar (PSA) dalam cawan Petri (diameter 9 cm) dan diinkubasi pada suhu ruang (27 OC) selama 6 hari, kemudian ditambah akuades
steril (10 ml/cawan), selanjutnya digunakan kuas halus untuk melepaskan konidium dari fialid. Setelah itu suspensi konidium dibuat pengenceran berseri dalam tabung reaksi yang berisi akuades steril dan dikocok dengan vorteks selama 30 detik. Pada pengenceran 10" diperoleh suspensi konidium dengan konsentrasi 1o61ml akuades yang dihitung dengan menggunakan hemasitometer Levy-Neubauer (Hadioetomo 1993).
lntroduksi Konidium
T.
itarziant~trt ke dalam Media PerlakuanSuspensi konidium T. hartianum berulnur 6 hari (konsentrasi 1o61ml) diintroduksikan pada masing-masing media perlakuan dengan cara disemprotkan pada permukaan media (10 mllpolybag). Masing-masing perlakuan dibiarkan di lahan terbuka di kebun percobaan PPSHB IPB Darmaga Bogor hingga 8 minggu.
Penghitungan Populasi T. harziartztnz dari Masiug-masing Media Perlakuan
Dari masing-masing media perlakuan diambil sampel masing-masing sebanyak 5 g pada bagian tengah media dengan kedalaman yang bervariasi yaitu lapisan permukaan, tengah (5 cm), dan bawah (10 cm). Setelah itu, masing- masing sampel dicampur hingga homogen dan diambil 5 g. Isolasi T. harzianum
cawan Petri (diameter 9 cm), selanjutnya dituang media PSA 10 ml (suhu 40 OC)
yang mengandung kloramfenikol (250 mgIL). Cawan Petri diputar secara rotasi dengan tangan dan diinkubasi pada suhu ruang (27 OC) selama 2 hari. Setiap perlakuan dibuat 2 ulangan. Populasi T harzianum ditentukan dengan cara men&tung rata-rata jumlah koloni dikalikan dengan faktor pengenceran.
Pertumbuhan, J u d a h Konidium, dan Persentase Perkecambahan Konidium
T.
IzarzianrmzTrichoderma harzianum yang digunakan adalah isolat cendawan hasil isolasi pada minggu ke-1 (MI), ke-5 (M5), dan ke-8 (M8) dari semua media perlakuan. Biakan T harziantr17z (diameter 3 mm) selanjutnya ditumbuhkan pada media PSA di cawan Petri (diameter 9 cm), dan diinkubasikan pada suhu ruang. Setiap perlakuan dibuat 3 ulangan. Pertumbuhan berdasarkan diameter koloni (mm) dilakukan setiap 24 jam sampai biakan benunur 3 x 24 jam. Selanjutnya koloni diinkubasikan sampai 30 hari, untuk penghitungan jumlah konidium (Jk) yang dilakukan dengan cara kultur dipotong-potong lalu ditimbang sebanyak 1 g dengan menggunakan neraca analitik merk Sartorius tipe BP 210s dan dimasukkan ke dalam labu Erlenmeyer dan diberi akuades steril sebanyak 100 ml, kemudian dikocok dengan vorteks selama 60 detik dan disaring dengan kain kasa steril untuk memisahkan konidium dari miselium dan media PSA. Selanjutnya Jk dihitung dengan inengguilakan heinasitolneter Levy-Neubauer (Hadioetomo 1993). Persentase konidium yang berkecambah (PKb) dihitung pada suspensi konidium tersebut di atas yang telah diinkubasikan selama 24,48, 72, dan 96 jam pada suhu ruang dengan bantuan hemasitometer di bawah mikroskop (400x). Penghitungan dilakukan pada 10 bidang pandang. PKb dihitung berdasarkan Sudirman et aI. (2008) rumus berikut :
PKb (%) = a h x 100 Keterangan :
PKb : Persentase konidium yang berkecambah
Penentuan Kandungan C-organik, N-total, Rasio C:N, Lignin, dan Selulosa
Parameter yang digunakan untuk mengetahui dekomposisi yang terjadi pada masing-masing serasah adalah penentuan kandungan C-organik, N-total, rasio C:N, lignin, dan selulosa pada masing-masing media perlakuan pada minggu ke-1, 5, dan 8 setelah introduksi T. harziantrm. Kandungan C-organik ditentukan dengan metode Walkey dan Black (Walkey 1947), N-total dengan metode Kjeldahl (Kjeldahl 1883), kandungan lignin dan selulosa menggunakan metode Van Soest (Amrullah & Suryahadi 1992). Selain itu faktor-faktor lain yang diduga berpengaruh nyata terhadap proses dekomposisi dan pertumbuhan T. harzianum seperti pH, curah hujan, dan suhu lingkungan juga diamati.
Analisis Data
HASIL DAN PEMBAHASAN
Populasi
T.
harzianunt pada Media PerlakuanHasil penelitian menunjukkan bahwa semua media perlakuan pada K1 (serasah daun), K2 (tanah yang mengandung masing-masing jenis serasah pada IC1 yang telah terdekomposisi), K3 (campuran masing-masing media K1 dan K2), d m TK berpengaruh terhadap populasi T. harzianum. Hasil tersebut berdasarkan pada rata-rata populasi T. harzianum pada masing-masing kelompok perlakuan. Rata-rata populasi T. harzianum pada pada K1, K2, dan K3 lebih rendah dibandingkan pada tanah kontrol (TK) (Tabel 1).
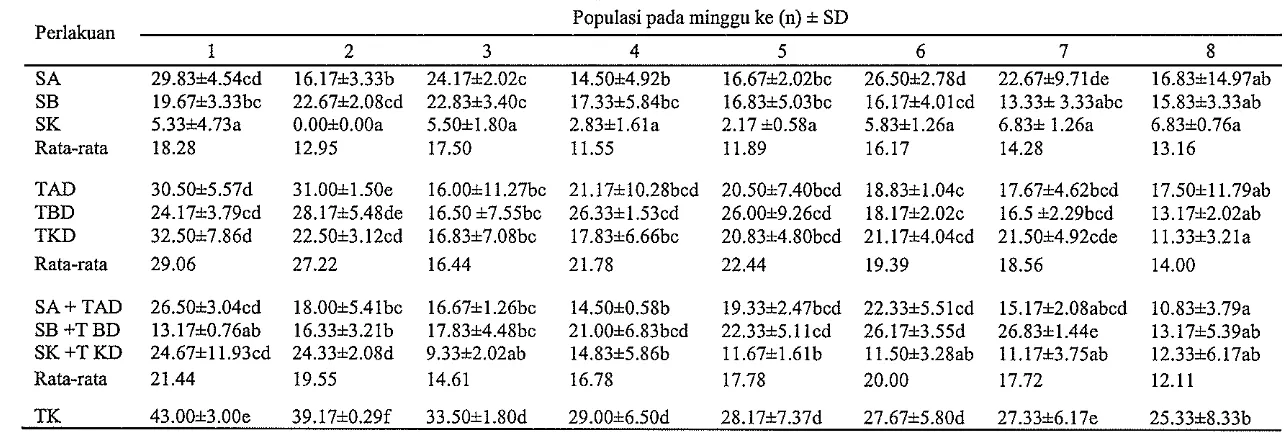
Pada minggu ke-1, populasi T. harzianum pada K1 berkisar 5.33-29.83 x lo3 kolonilg dengan rata-rata 18.28 x lo3 kolonilg, K2 24.17-32.50 x 10' kolonilg (rata-rata 29.06 x lo3 kolonilg), K3 13.17-26.50 x lo3 kolonilg (rata-rata 21.44 x lo3 kolonilg), dan TK dengan rata-rata 43 x lo3 kolonilg (Gambar 2). Setelah delapan minggu inkubasi, rata-rata populasi T. harzianum mengalami penurunan pada K1, K2, dan K3 masing-masing sebesar 13.16 x lo3 kolonilg, 14 x lo3 d m
12.1 1 x lo3 kolonilg. Sebaliknya, rata-rata populasi T. harzian~rm pada TK relatif tinggi yaitu 25.33 x lo3 kolonilg setelah delapan minggu inkubasi.
Hasil analisis ragam pengaruh semua media perlakuan terhadap populasi T. harzianum dari minggu ke-1 sampai minggu ke-8 disajikan pada Tabel 2. Berdasarkan analisis ragam dapat diketahui bahwa pada rninggu ke-1 sampai minggu ke-7 seinua media perlakuan berpengaruh sangat nyata (P < 0.01) terhadap populasi T. harzianum di dalam media perlakuan, sedangkan pada minggu ke-8 semua media perlakuan tidak berpengaruh nyata (P > 0.05) terhadap populasi T. harzianum di dalam media perlakuan.
Tabel 1 Populasi T. harzianum (x103 kolonilg) pada kee~npat kelompok media yang dihitung setiap minggu selarna inkubasi Populasi pada minggu ke (n) 1 SD
Perlakuan
1 2 3 4 5 6 7 8
S A 29.8314.54cd 16.1713.33b 24.1712.02~ 14.5014.92b 16.6712.02bc 26.5012.78d 22.6719.71de 16.83114.97ab SB 19.6713.33bc 22.67i2.08cd 22.8313.40~ 17.3315.84bc 16.8315.03bc 16.1714.01cd 13.331 3.33abc 15.8313.33ab SK 5.3314.73a 0.0010.00a 5.5011.80a 2.8311.61a 2.17 10.58a 5.8311.26a 6.831 1.26a 6.8310.76a
Rata-rata 18.28 12.95 17.50 11.55 11.89 16.17 14.28 13.16
TAD 30.5015.57d 31.0011.50e 16.00+11.27bc 21.17110.28bcd 20.5017.40bcd 18.8311.04~ 17.6714.62bcd 17.50-tl1.79ab TBD 24.1713.79cd 28.17+5.48de 16.50 17.55bc 26.3311.53cd 26.0019.26cd 18.1712.02~ 16.5 12.29bcd 13.1712.02ab TKD 32.5017.86d 22.5013.12cd 16.8317.08bc 17.8316.66bc 20.83i4.80bcd 21.1714.04cd 21.50+4.92cde 11.3313.21a
Rata-rata 29.06 27.22 16.44 21.78 22.44 19.39 18.56 14.00
S A + TAD 26.5053.04cd 18.0015.41bc 16.67i1.26bc 14.5010.58b 19.3312.47bcd 22.3315.51cd 15.1712.08abcd 10.8313.79a SB+TBD 13.17+0.76ab 16.3313.21b 17.83-L.4.48bc 21.0016.83bcd 22.3315.11cd 26.1713.55d 26.8311.44e 13.1715.39ab SK +T KD 24.67111.93cd 24.3312.08d 9.33i2.02ab 14.8315.86b 11.6711.61b 11.5013.28ab 11.1713.75ab 12.3316.17ab
Rata-rata 21.44 19.55 14.61 16.78 17.78 20.00 17.72 12.11
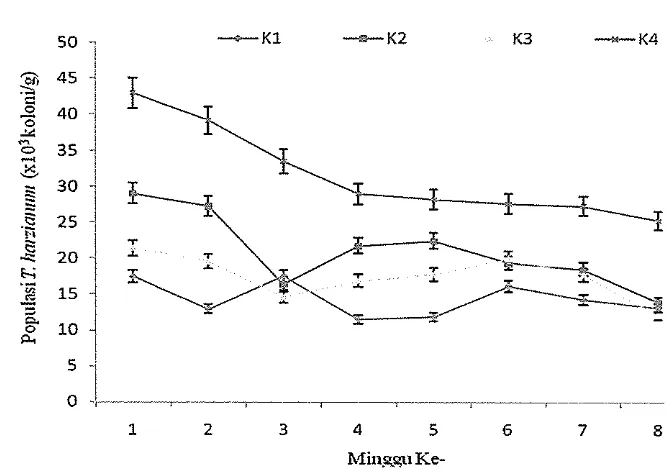
[image:34.842.89.731.141.360.2]Gambar 2 Populasi
T.
harzianum pada berbagai kelompok perlakuan setelahdiinkubasi beberapa minggu ke dalam media perlakuan. K1: kelompok pertama, K2: kelompok kedua, K3: kelompok ketiga, K4: kelompok keempat
Tabel 2 Hasil analisis ragam pengaruh semua media perlakuan terhadap populasi
T.
harzzanum selama 8 minggu inkubasiMinggu ke- Pengaruh
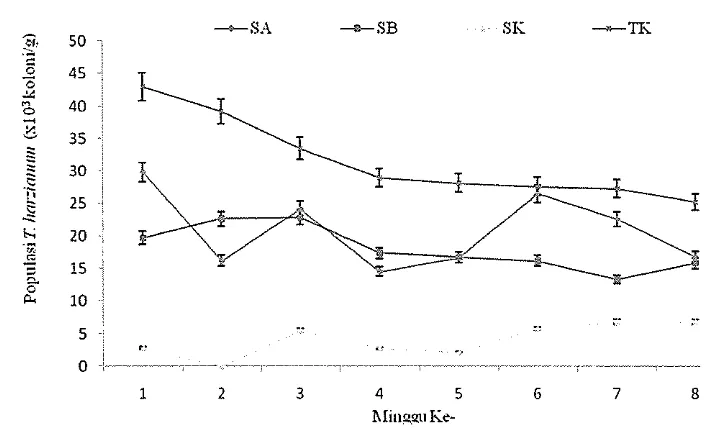
[image:35.595.115.450.94.332.2]Gambar 3 Populasi T. harzianurn pada berbagai serasah setelah dinkubasi beberapa minggu ke dalam media perlakuan. SA: serasah akasia, SB: serasah bawang merah, SK: serasah kacang tanah, d m TK: tanah alami sebagai kontrol
[image:36.595.119.473.113.328.2]Gambar 4 Populasi T. harzianum pada berbagai tanah yang inengandung serasah yang telah terdekomposisi setelah diinkubasi beberapa minggu ke dalam media perlakuan. TAD: tanah yang mengandung serasah akasia yang telah terdekomposisi, TBD: tanah yang mengandung serasah bawang merah yang telah terdekomposisi, TKD: tanah yang mengandung serasah kacang yang telah tanah terdekomposisi, dan TK: tanah alami sebagai kontrol
50
,-.
-8-- T;\D -TBD TIiD-
TIi [image:37.595.131.476.111.294.2]Berdasarkan hasil penelitian, rata-rata populasi pada media K2 ditemukan relatif lebih tinggi sampai minggu ke-5 dibandingkan pada K1 dan K3. Fenomena ini mengindikasikan bahwa viabilitas cendawan pada K2 lebih tinggi dibandingkan pada K1 dan K3. Sebaliknya hasil penelitian Sudirman et al. (2008) melaporkan bahwa populasi B. bassiana pada K2 relatif lebih rendah dibandingkan pada K1 dan K3, ha1 ini karena yang diuji berbeda yaitu serasah Brassicaceae (caisin, kubis, sawi) dan bawang merah.
Populasi T. harzianum pada SK jauh lebih rendah dibandingkan pada SA dan SB (Gambar 3). Rendahnya populasi T. harzianum diduga dipengambi oleh cendawan lain yang menutupi peimukaan media sewaktu inengisolasi T. harzianum dari SK. Populasi T. harzianum pada SK berkorelasi negatif dengan kandungan karbon organik (C-organik), nitrogen total (N-total), dan rasio C:N pada minggu ke-1, ke-5, dan ke-8 (Lampiran 43-45). Bae et al. (2002) melaporkan bahwa peningkatan biomassa mikroorganisme tanah sebagai faktor total biotik berpengamh terhadap pertumbuhan dan efikasi T. harzianum di dalam tanah. Kandungan karbon, nitrogen, dan rasio C:N berpengaruh terhadap pertumbuhan Trichodernza spp. (Venna et al. 2007). Menumt Gao dan Liu (2009) cendawan tumbuh optimal pada rasio C:N yang berbeda-beda sebagai bentuk respon cendawan terhadap konsentrasi C dan N dalam substrat.
Pertumbuhan
T.
harziaizuiii pada Media PSAHasil penelitian menunjukkan bahwa diameter koloni isolat M1 dari semua media perlakuan pada K1, K2, dan K3 yang ditumbuhkan pada media PSA pada hari ke-1 pengamatan (HI) antara 13.17-16.17 mm, hari ke-2 (H2) antara 34.83- 47.67 mm, hari ke-3 (H3) antara 64.67-84.17 mm, sedangkan TK pada H1, H2, dan H3 masing-masing 17 min, 51 min, dan 85.5 mm (Tabel 3). Diameter koloni isolat M5 pada H1 antara 30.67-33.5 mm, H2 antara 65.5-70.5 mm, H3 antara 85.83-87 mm, sedangkan TK pada H1, H2, dan H3 masing-masing 31.5 mm, 66.5 mm, dan 87 mm. Diameter koloni isolat M8 pada H1 antara 26.33-36 mm, H2 antara 64.67-72.67 mm, H3 87 mm, sedangkan TK pada HI, H2, dan H3 masing- masing 26.83 mm, 64.67 mm, dan 87 mm.
Berdasarkan data tersebut di atas menunjukkan bahwa rata-rata diameter koloni isolat M1 pada HI dan H2 lebih rendah dibandingkan diameter koloni isolat M5 dan M8 pada H1 dan H2. Sementara itu, rata-rata diameter koloni isolat M1 pada H3 relatif tidak berbeda dengan diameter koloni isolat M5 dan M8 pada H3. Semakin lama isolat (M5 dan M8) berada di media perlakuan, maka pertumbuhannya semakin baik ketika ditumbuhkan pada media PSA. Efek media perlakuan hanya berpengaruh pada isolat MI. Jadi secara umum pengaruh media perlakuan TK tidak berbeda dengan pengaruh media perlakuan lainnya terhadap diameter koloni isolat MI, M5, dan M8 pada media PSA.
Tabel 3 Pertumbuhan isolat minggu ke-1 (MI), ke-5 (M5), dan ke-8 (M8) dari semua media perlakuan pada media PSA yang diamati pada hari ke-1 (HI), ke-2 (H2), dan ke-3 (H3)
TAD 15.17-tO.29ab 41.3311.61b 73.5010.50b 32.6711.76ab 68.3311.76abc 87.0010.00b 34.1711.26b 72.6710.76bc 87.00*0.00a TBD c 34.8310.29a 64.6711.04a 33.50-tO.50b 69.3311.53bc 87.0010.00b 26.3314.86a 65.6716.93ab 87.0010.00a TKD 13.1711.15a 45.3312.89bc 81.5M4.27~ 31.17?C0.76ab 67.3310.58ab 85.8311.04ab 35.6710.58b 74.1710.76~ 87.0010.00a
15.83k1.53bc
Rata-rata 14.72 40.50 73.89 32.45 68.33 73.89 32.06 70.84 87.00
SA+TAD 14.8311.04ab 43.3311.76bc 74.17k2.47b 32.83k1.04ab 68.3312.57abc 87.0010.00b 34.0011.00b 71.6711.53abc 87.0010.00a SB + TBD 14.1710.58ab 36.67+1.04a 66.00*3.00a 32.0010.50ab 65.5010.87a 85.3311.76a 33.50*0.87b 70.8311.26abc 87.0010.00a SK+TKD 15.0011.18ab 47.67k0.26cd 84.1711.37~ 32.1711.81ab 66.1711.81a 86.0010.89ab 27.00*1.18a 66.5011.34ab 87.0010.00a Rata-rata 14.67 42.56 74.78 32.33 66.67 86.11 31.50 69.67 87.00
[image:40.842.84.733.148.376.2]Produksi Konidium T . hnrzinnuin pada Media PSA
[image:41.595.102.516.351.533.2]Berdasarkan hasil penelitian menunjukkan bahwa isolat MI, M5, dan M8 ternyata masih mampu memproduksi konidium ketika ditumbuhkan pada media PSA. Jumlah konidium (Jk) isolat M1 semua media perlakuan pada K1, K2, dan K3 antara 2.93-3.28 x lo6 konidiudg, sedangkan pada TK sebesar 3.43 x lo6 konidiudg (Tabel 4). Jk isolat M5 antara 3.23-4.23 x lo6 konidiudg, sedangkan pada TK sebesar 3.48 x lo6 konidiudg. Jk isolat M8 antara 3.25-4.13 x lo6 konidiudg, sedangkan pada TK sebesar 3.33 x lo6 konidiudg. Jadi pengaruh media perlakuan TK tidak berbeda dengan pengaruh media perlakuan lainnya terhadap Jk isolat MI, M5, dan M8 pada media PSA.
Tabel 4 Produksi konidium isolat MI, M5, dan M8 dari semua media perlakuan pada media PSA
Rata-rata jumlah konidium (xlOU) konidiudml
+
SD PerlakuanMI M5 M8
S A 3.2310.32a 3.7510.35ab 3.80*0.35ab
SB 3.1810.60a 3.5810.25a 3.6310.60ab
SK 3.0510.14a 4.2310.11b 4.13-tO.lSb
Rata-rata 3.15 3.85 3.85
TAD 3.15A0.49a 3.3510.42a 3.3810.04a
TBD 2.93h0.39a 3.2310.04a 3.2510.21a
TKD 3.2811.17a 3.4010.07a 3.4010.14a
Rata-rata 3.07 3.33 3.34
SA + TAD 3.0311.17a 3.4810.07a 3.53A0.14ab SB + TBD 2.9811.24a 3.2811.61a 3.48A1.60ab SK + TKD 3.2510.14a 3.3010.21a 3.4310.18a
Rata-rata 3.09 3.35 3.48
TK 3.4311.11a 3.4810.25a 3.3310.32a
Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata berdasarkan uji DMRT pada taraf kepercayaan 95%. SA: serasah akasia, SB: serasah bawang merah, SK: serasah kacang tanah, AD: tanah yang mengandung serasah akasia terdekomposisi, BD: tanah yang mengandung serasah bawang merah terdekomposisi, KD: tanah yang mengandung serasah kacang tanah terdekomposisi, dan TK: tanah alami sebagai kontrol
Tabel 5 Hasil analisis ragam pengaruh semua media perlakuan terhadap produksi konidium isolat M1, M5, dan M8 pada media PSA
Isolat Pengaiuh
M1
tn
M5
*
Keterangan
*
= nyata pada taraf kepercayaan 95% tn = tidak nyataKemampuan produksi konidium isolat MI, M5, dan M8 dari media perlakuan meningkat antara 3.53-26.15%, tetapi pada TK menurun sebesar 3% selama periode 8 minggu. Klein dan Eveleigh (1998) melaporkan bahwa viabilitas konidium Trichoderma mengalami penurunan sekitar 10% akibat pengaruh fungistatis di dalam tanah selaina periode dua tahun.
Perkecambahan Konidium T. harzinrturrz
Tabel 6 Perkecambahan konidium isolat MI, M5, dan MS dari semua media perlakuan setelah 24, 48, 72, dan 96 jam inkubasi dalam akuades
Perkecambahan konidium (%)
Perlakuan M1 M5 M8
24 48 72 96 24 48 72 96 24 48 72 96
TAD 34.35a 51.07a 67.93ab 73.29ab 35.42a 51.63a 69.48a 80.36a 35.54a 51.18ab 70.04a 81.25a
TBD 33.55a 46.48a 63.38a 71.81~1 34.42a 46.70a 66.13a 79.29a 34.86a 47.22a 68.03a 79.52a
TKD 34.83a 48.66a 67.23ab 73.09ab 35.23a 49.30a 71.39a 80.18a 35.52a 49.80ab 71.74~1 80.34a Rata-rata 34.24 48.74 66.18 72.73 35.02 49.21 69.00 79.94 35.31 49.40 69.94 80.37
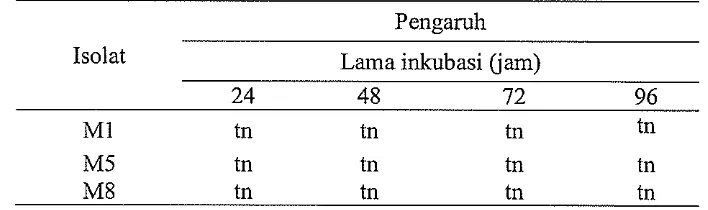
[image:43.842.99.736.156.422.2]Hasil analisis ragam pengaruh semua media perlakuan terhadap PKb isolat M1, M5, dan M8 setelah 24, 48, 72, dan 96 jam inkubasi disajikan pada Tabel 7. Berdasarkan hasil analisis ragam dapat diketahui bahwa semua media perlakuan tidak berpengaruh nyata (P > 0.05) terhadap PKb isolat MI, M5, dan M8 setelah diinkubasikan selama 24,48,72, dan 96 jam dalam akuades.
Tabel 7 Hasil analisis ragam pengaruh semua media perlakuan terhadap PKb isolat MI, M5, dan M8 setelah 24, 48, 72, dan 96 jam inkubasi dalam akuades
Pengaruh Isolat Lama inkubasi (jam)
24 48 72 96
M1 tn tn tn tn
M5 tn tn tn tn
M8 tn tn tn tn
Keterangan tn = tidak nyata pada taraf kepercayaan 95%
Berdasarkan data tersebut di atas menunjukkan bahwa semua media perlakuan tidak berpengaruh terhadap PKb isolat MI, M5, dan M8 yang diinkubasikan dalam akuades setelah 24,48, 72, dan 96 jam inkubasi. Sebaliknya Sudirman et al. (2008) melaporkan bahwa efek media perlakuan serasah daun tanaman Brassicaceae dan bawang merah dan tanah yang mengandung serasah yang telah terdekomposisi dari tanaman tersebut berpengaruh negatif terhadap perkecambahan konidium B. bassiana yang diinkubasikan dalam akuades setelah 10 jam inkubasi. Penelitian yang sama dilaporkan Inyang et al. (1999) bahwa isotiosianat dari tanaman Brassicaceae menghambat perkecambahan konidium M. anisopliae dan Paecilomyces fumosoroseus.
[image:44.595.154.508.265.370.2]ukuran konidium semakin cepat waktu yang dibutuhkan konidium untuk berkecambah. Dalam penelitian ini penghitungan PKb dimulai dari proses konidium berkecambah (Gambar 6b). Sedangkan menurut Junianto & Sukamto (1995) bahwa karakteristik konidium yang berkecambah dapat dikenali apabila panjang tabung kecambah telah melebihi diameter konidium. Dengan demikian, pembentukan tabung kecambah merupakan stadium awal yang sangat menentukan cepat atau lambatnya cendawan berkolonisasi dan berpenetrasi pada cendawan inang di lapangan.
[image:45.602.104.501.304.650.2]Kandungan C-organik, N-total, dan Rasio C:N
Hubungan populasi T. harzianum dengan kandungan C-organik, N-total, dan rasio C:N semua media perlakuan dapat dilihat pada Tabel 8. Hasil penelitian menunjukkan bahwa populasi T. harzianum pada minggu ke-1 tertinggi pada TK dan terendah pada SK, hasil yang sama juga terlihat pada minggu ke-5 dan ke-8. Kandungan C-organik, N-total, dan rasio C:N media perlakuan TK pada minggu ke-1 masing-masing 0.96%, 0.14%, dan 6.9: 1, minggu ke-5 masing-masing 0.84%, 0.19%, dan 4.4:1, dan minggu ke-8 masing-masing 0.14%, 0.12%, dan 1.2:1. Sementara itu kandungan C-organik, N-total, dan rasio C:N media perlakuan SK pada minggu ke-1 masing-masing 28.17%, 2.05%, dan 13.7:1, minggu ke-5 masing-masing 22.46%, 2.07%, dan 10.9:1, dan minggu ke-8 masing-masing 21.8%, 2.12%, dan 10.3:l.
Berdasarkan data tersebut di atas menunjukkan bahwa populasi T. harzianum pada media perlakuan TK tinggi pada rasio C:N yang rendah, tetapi rasio C:N yang semakin rendah pada minggu ke-5 dan ke-8 tidak meningkatkan populasi T. harzianum pada media perlakuan. Verma et al. (2007) melaporkan bahwa rasio C:N berpengaruh terhadap pertumbuhan Trichoderma spp..
Tabel 8 Kandungan C-organik, N-total, rasio C:N, kandungan lignin dan selulosa semua media perlakuan pada minggu ke-1, ke-5, dan ke-8
Minggu ke-l Minggu ke-5 Minggu ke-8
Perlakuan
C-org N-tot C:N Lig Sel Pop C-org N-tot C:N Lig Sel Pop C-org N-tot C:N Lig Sel POP
Rata-rata 17.44 11.86 13.16
TAD 2.85 0.31 9.2 12.19 8.70 30.50 2.82 0.31 9.1 18.00 7.91 20.50 2.70 0.31 8.7 16.82 8.12 17.5
TBD 1.09 0.11 9.9 13.50 2.23 24.17 0.78 0.12 6.5 8.72 2.93 26.00 0.77 0.13 5.9 11.07 4.27 13.17
TKD 1.39 0.19 7.3 11.81 5.76 32.50 1.43 0.19 7.5 8.00 7.04 20.83 1.50 0.20 7.5 21.39 10.38 11.33
Rata-rata 29.06 22.55 14.00
SA+TAD 17.25 0.68 25.4 22.56 17.08 26.50 16.80 0.82 20.5 21.00 21.39 19.33 14.98 0.96 5 23.82 18.19 10.83 SB+TBD 6.68 0.40 16.7 12.65 2.92 13.17 3.73 0.50 7.5 6.44 7.12 22.33 3.71 0.46 8.1 11.40 6.93 13.17
SK+TKD 7.78 0.51 15.3 10.20 6.55 24.67 7.19 0.64 11.2 13.00 15.27 11.67 4.18 0.47 8.9 15.68 6.67 12.33
Rata-rata 2 1.45 17.78 12.11
TK 0.96 0.14 6.9 7.58 11.00 43.00 0.84 0.19 4.4 10.00 13.15 28.17 0.14 0.12 1.2 9.28 12.31 25.33
[image:47.842.84.738.155.389.2]Pada inedia perlakuan SA terlihat adanya kecenderungan penunman kandungan C-organik dan peningkatan kandungan N-total. Kandungan C-organik pada minggu ke-1, ke-5, dan ke-8 masing-masing 49.06%, 48.49%, dan 45%, sedangkan kandungan N-total inasing-masing 1.76%, 1.95%, dan 1.98% (Tabel 8). Hasil ini sama dengan pola penelitian sebelurnnya pada serasah akasia yang dilakukan oleh Samingan (2009). Penurunan kandungan C-organik berkaitan dengan pemanfaatan energi oleh inikroorganisme yang terlibat di dalam proses dekomposisi. Menurut Atlas dan Bartha (1993) bahwa selama proses dekomposisi sebagian besar karbon akan dibebaskan dalam bentuk C02 oleh mikroorganisme pada kondisi aerobik, sedangkan pada kondisi anaerohik karbon dibebaskan sebagai metan, garam-garam organik, alkohol dan C02. Tetapi tidak semua karbon dilepaskan dalam bentuk C02, karena menurut Tuomela et al. (1999) bahwa sebagian sumber karbon digunakan oleh mikroorganisme sebagai sumber energi dan bahan untuk pembentukan sel-sel baru bersama-sama dengan unsur lainnya. Peningkatan kandungan nitrogen disebabkan adanya degradasi protein pada serasah oleh cendawan, selain i t - juga berasal dari mobile nitrogen pada hifa cendawan dan immobile nitrogen di dalam enzim yang diekskresikan cendawan ke dalam substrat serasah. Menurut Miyamoto dan Hiura (2007) pertuinbuhan miselium cendawan pada serasah selama proses dekomposisi dapat inenyebabkan peningkatan konsentrasi nitrogen.
Kandungan Lignin dan Selulosa
Gambar 7 Kandungan lignin semua media perlakuan pada minggu ke-1, ke-5, dan ke-8 setelah diintroduksi T. harzianum. SA: serasah akasia, SB: serasah bawang merah, SK: serasah kacang tanah, TAD: tanah yang mengandung serasah akasia terdekomposisi, TBD: tanah yang mengandung serasah bawang merah terdekomposisi, TKD: tanah yang mengandung serasah kacang tanah terdekomposisi, dan TK: tanah alami sebagai kontrol
[image:49.595.98.476.77.839.2] [image:49.595.122.458.98.302.2]Berdasarkan data di atas menunjukkan bahwa populasi T. harzianum pada media perlakuan TK tinggi pada kandungan lignin dan selulosa yang rendah, sebaliknya populasi T. harzianum pada SA cukup tinggi pada minggu ke-1, ke-5, dan ke-8 meskipun kandungan lignin dan selulosa tertinggi.
Hasil uji korelasi menunjukkan bahwa kandungan lignin dan selulosa semua media perlakuan berkorelasi negatif dengan populasi T. harzianum pada minggu ke-1, ke-5, dan ke-8. Selengkapnya hasil uji korelasi tersebut disajikan pada Lampiran 43-45.
Kandungan lignin pada minggu ke-1, ke-5, dan ke-8 ditemukan relatif lebih tinggi dibandingkan kandungan selulosa pada semua media perlakuan. Hal ini mengindikasikan bahwa mikroorganisme dekomposer lebih sulit mendegradasi lignin dibandingkan selulosa. Menurut Pometto dan Crawford (1986) bahwa lignin merupakan senyawa polimer aromatik yang resisten terhadap degradasi berbagai mikroorganisme dekomposer termasuk cendawan. Samingan (2009) melaporkan bahwa T. harzianum mampu mendekomposisi selulosa lebih tinggi dibandingkan lignin. Hal yang sama dilaporkan Schmidt (2006) bahwa Trichoderma merupakan cendawan selulolitik yang memiliki potensi yang baik mendekoinposisi selulosa dan hemiselulosa dibandingkan lignin. Kemampuan Trichoderma mendekomposisi selulosa karena cendawan ini menghasilkan enzim
p
1-4 selobiohidrolase, endoglukanase, eksoglukanase dan duaP
1-4 glukosidase (Dix & Webster 1995; Evans & Hedger 2001). Disamping itu, aktivitas cendawan dalam mendekomposisi lignin dan selulosa dipengaruhi oleb faktor lingkungan seperti kelembaban dan suhu dalam substrat (Pometto & Crawford 1986; Donnelly et al. 1990). Kondisi lingkungan yang tidak menguntungkan seperti kelembaban yang rendah menghambat cendawan mendekomposisi selulosa dan lignin, sebaliknya dekomposisi selulosa dan lignin mengalami peningkatan yang signifikan pada kondisi kelembaban (40-60%) dan suhu 24 OC (Donnelly et al.SIMPULAN DAN SARAN
Simpulan
Semua media perlakuan berpengaruh sangat nyata (P < 0.01) terhadap fluktuasi populasi T. harzianum di lapangan sejak minggu ke-1 sampai minggu ke-7, tetapi tidak berpengaruh nyata (P > 0.05) terhadap populasi T. harzianum di lapangan pada minggu ke-8 sehingga viabilitas T. harzianzrm rendah pada semua media perlakuan. Semua media perlakuan tidak berpengaruh nyata (P < 0.05) terhadap pertumbuhan, kemampuan produksi konidium, dan perkecambahan konidium pada media PSA. Populasi T. harzianum berkorelasi negatif dengan kandungan C-organik, N-total, rasio C:N, kandungan lignin dan selulosa semua media perlakuan pada minggu ke-1, ke-5, dan ke-8.
Saran
Dari penelitian yang telah dilakukan maka dapat disarankan beberapa ha1 sebagai berikut:
1. Perlu dilakukan penelitian lanjutan untuk mengetahui faktor lain yang mempengamhi tingginya populasi T. harzianum Rifai isolat TBPH pada tanah kontrol pada kondisi laboratorium dan lapangan.
2. Perlu dilakukan penelitian lanjutan untuk mengetahui faktor lain yang berpengaruh terhadap viabilitas T. harzianurn Rifai isolat TBPH pada serasah kacang tanah pada kondisi laboratorium dan lapangan.
DAFTAR
PUSTAKA
Abeysinghe S. 2007. Biological control of Fusarium solani f. sp. phaseoli the causal agent of root rot of bean using Bacillus subtilis CA32 and Trichoderma harzianum RUOI. Ruhuna J S c i 2: 82-88.
Adiyoga W, Basuki RS, Hilman Y, Udiarto BK. 1997. Studi base line identifikasi dan pengembangan teknologi PHT pada tanaman cabai di Jawa Barat. Makalah Hasil Seminar Hasil Penelitian Pendukung PHT. Jakarta: Departemen Pertanian.
Akrami M, lbrahimov As, Valizadeh E, Dizaji AA. 2008. The ability of Trichodertna harzianum isolates in controlling rapeseed Sclerotinia rot in comparison with the effects of Bacillus subtilis and Benomyl fungicide. Res JBiol Sci 3(8): 856-859.
Alexopolous CJ, Mims CW. 1996. Introductory Mycology. 41h Ed. Canada: John Wiley & Sons.
Amrullah IK, Suryahadi. 1992. Kumptrlan Bahan Penunttrn Praktikum Ilmu Malcanan Ternak. Bogor: Departemen Pendidikan dan Kebudayaan DIKTI-
PAU Ilmu Hayat IPB.
Atlas RM, Bartha R. 1993. Microbial Ecology, Fundameiztal and Application. Redwood City California: The BenjaminICummings Publishing Company. Bae YS, Knudsen GR, Dandurand LMC. 2002. Influence of soil microbial
biomass on growth and biocontrol efficacy of Trichoderma harzianunz. Plant Pathol J 18(1): 30-35.
Bal U, Altintas S. 2008. Effects of Trichodernza harziantrm on lettuce in protected cultivation. J Central Euro Agri 9(1): 63-70.
Barnett HL, BB Hunter. 1999. Illustrated Genera of Imperfect Fungi. 4th Ed. Minnesota: APS Press.
Benitez T, Rincon AM, Limon MC, Codon AC. 2004. Biocontrol mechanism of Trichodernza strains. Int Microbiol 7: 249-260.
Broder MW, Wagner GH. 1988. Microbial colonization and decomposition of corn, wheat and soybean resdue. Soil Sci Soc Am J 52: 112-1 17.
Chet I, Inbar J. 1994. Biological control of fungal pathogens. Appl Biochem Biotechno148: 37-43.
Delgado JJ, Pintor-Toro JA, Benitez T. 2000. Overproduction of p-1,6-glucanase (BGN16,l) in Trichoderma hnrzianum is controlled by extracellular acidic proteases and pH. Biochim Biophys Acta 1481: 289-296.
Delgado JJ, Moreno MMA, Benitez T. 2003. Glucose uptake in Trichoderma harzianum: role of gttl. Euk Cell 2: 708-717.
Dharmaputra OS, Putri ASR, Retnowati, Saraswati S. 2003. Penggunaan Trichoderma harzianum untuk mengendalikan Aspergillusflavus penghasil aflatoksin pada kacang tanah. JFitopat Indones 7(1): 28-37.
Dix NJ, Webster J. 1995. Fungal Ecology. London: Chapman & Hall.
Domsch KH, Gams W, Anderson TH. 1