PERBANDINGAN ANTARA PERTUMBUHAN MORFOMETRIK
KOMODO (
Varanus komodoensis
) PADA PULAU BESAR
DENGAN PULAU KECIL DI TAMAN NASIONAL KOMODO
RINI RISMAYANI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Perbandingan Pertumbuhan Morfometrik Komodo (Varanus komodoensis) antara Pulau Besar dengan Pulau Kecil di TN. Komodo adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
Rini Rismayani
RINGKASAN
RINI RISMAYANI. Perbandingan Pertumbuhan Morfometrik Komodo (Varanus komodoensis) antara Pulau Besar dengan Pulau Kecil di TN. Komodo. Dibimbing oleh YANTO SANTOSA dan NOVIANTO BAMBANG WAWANDONO.
Biawak komodo di Taman Nasional Komodo tersebar di dua pulau besar (Pulau Komodo dan Pulau Rinca) dan dua pulau kecil (Nusa Kode dan Gili Motang). Pertumbuhan biawak komodo telah diukur dalam studi capture mark recapture pada tahun 2002 sampai dengan 2012. Penelitian ini bertujuan membandingkan laju pertumbuhan morfometrik antara populasi komodo di pulau besar dengan populasi komodo di kecil, model pertumbuhan biawak komodo diantara pulau besar dan kecil serta membandingkan karakteristik habitatnya. Uji beda t dilakukan untuk membandingkan laju pertumbuhan jenis morfometrik, periode pertumbuhan serta kelas umur yang sama, antara komodo yang tinggal di pulau kecil dengan komodo yang tinggal di pulau besar.
Populasi komodo kelas umur anak pada pulau kecil signifikan memiliki laju pertumbuhan morfometrik yang lebih rendah (151.18%) dibanding laju pertumbuhan morfometrik populasi komodo di pulau besar. Pada kelas umur remaja, nilai tengah laju pertumbuhan morfometrik komodo di pulau besar lebih tinggi (75.61%) dibandingkan komodo di pulau kecil tetapi tidak menunjukkan perbedaan yang signifikan. Karakteristik habitat biawak komodo yang meliputi ukuran dan jarak antar pulau, tipe ekosistem, keberadaan pemukiman dan aktivitas wisata serta kelimpahan mangsa, di pulau kecil mengarah pada terjadinya tekanan pada populasi biawak komodo di pulau kecil yang laju pertumbuhannya rendah dan jumlahnya semakin menurun.
SUMMARY
RINI RISMAYANI. Comparison of Morphometric Growth on Komodo (Varanus komodoensis) in Islands and Islets at Komodo NP. Supervised by YANTO SANTOSA dan NOVIANTO BAMBANG WAWANDONO.
Komodo dragons on Komodo National Park is spread across two large islands (Komodo and Rinca) and two small islands (Nusa Kode and Gili Motang). Komodo dragons growth has been measured in the study of capture mark and recapture in 2002 up to 2012. This study aimed to compare the growth of morphometric population of dragons on islands to population of dragons in the islets, to compare dragons growth model on islands to islets, and to compare the characteristics of the islands habitat to islets habitat. The T different test was performed on the same growth rate of morphometric types, the growth period and age class, between the dragons on island and islets.
Juvenile dragons on islets significantly has a growth rate of morphometric lower (151.18%) than juvenile dragons growth rate of morphometric on island. While median growth rate of morphometric on subadult dragons in island was higher (75.61%) than subadult dragons on islets but did not show significant differences. Habitat characteristics of dragons that include the size and spacing between islands, types of ecosystems, residential and tourist activity and the abundance of prey on a islets, leads to the pressure on dragons population in islets so that growth rate was low and its amount decreasing.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Konservasi Biodiversitas Tropika
PERBANDINGAN ANTARA PERTUMBUHAN MORFOMETRIK
KOMODO (
Varanus komodoensis
) PADA PULAU BESAR
DENGAN PULAU KECIL DI TAMAN NASIONAL KOMODO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji syukur kepada Allah SWT atas berkat karuniaNya sehingga karya ini berhasil diselesaikan. Obyek yang diambil dalam penelitian ini adalah biawak komodo (Varanus komodoensis) pada lokasi penulis bekerja yaitu di Taman Nasional Komodo. Karya ilmiah ini berjudul Perbandingan Pertumbuhan Morfometrik Biawak Komodo (Varanus Komodoensis) pada Pulau Besar dan Pulau Kecil di Taman Nasional Komodo.
Terimakasih penulis ucapkan kepada Bapak Prof. Dr. Ir. Yanto Santosa, DEA serta Bapak Dr. Ir. Novianto Bambang Wawandono, M.Si selaku komisi pembimbing yang telah banyak mencurahkan waktu dan pikiran selama proses penulisan. Terimakasih penulis sampaikan kepada Kementerian Kehutanan yang telah memberi tugas belajar dan mendanai seluruh tahapan studi termasuk penelitian. Penghargaan penulis sampaikan untuk Kepala Balai Taman Nasional Komodo dan seluruh staf, serta Yayasan Komodo Survival Program, yang telah memberi ijin akses database komodo. Ucapan terimakasih juga penulis tujukan kepada Heru Rudiharto, Aganto Seno, Deni Purwandana, Achmad Ariefandy dan Tim Jessop, yang telah berbagi pengalaman lapangannya dan berdiskusi dengan penulis. Ungkapan terima kasih juga disampaikan kepada ibu, suami, anak-anak dan seluruh keluarga, serta rekan-rekan studi, atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2015
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 1
Tujuan Penelitian 2
Hipotesis Penelitian 2
Manfaat Penelitian 2
2 METODE 3
Lokasi dan Waktu 3
Bahan dan Alat 3
Prosedur Analisis Data 6
3 HASIL DAN PEMBAHASAN 7
Perbandingan laju pertumbuhan antara pulau besar dan pulau kecil 7 Perbandingan model laju pertumbuhan morfometrik pada pulau besar dan
dan pulau kecil 13
Perbandingan karakteristik habitat antara pulau besar dan pulau kecil 17
Implikasi Konservasi 23
4 SIMPULAN DAN SARAN 23
Simpulan 23
Saran 24
DAFTAR PUSTAKA 24
LAMPIRAN 27
DAFTAR TABEL
1 Jenis dan sumber data yang menjadi bahan penelitian 4 2 Rata-rata laju pertumbuhan morfometrik biawak komodo kelas umur anak
di pulau besar dan pulau kecil 11
3 Rata-rata laju pertumbuhan morfometrik biawak komodo kelas umur
remaja di pulau besar dan pulau kecil 12
4 Perbandingan korelasi antara pertambahan umur dengan laju pertumbuhan morfometrik panjang kepala, lebar kepala dan SVL 14 5 Perbandingan korelasi antara pertambahan umur dengan laju pertumbuhan
morfometrik ventral TBL, panjang ekor, lingkar ekor dan berat tubuh 14 6 Model regresi pendugaan pertambahan umur dari variabel persentase
pertumbuhan morfometrik panjang kepala, lebar kepala dan SVL 15 7 Model regresi pendugaan pertambahan umur dari variabel persentase
pertumbuhan morfometrik ventral TBL, panjang ekor, lingkar ekor dan
berat tubuh 15
8 Proporsi tipe ekosistem pada masing-masing pulau di Taman Nasional
Komodo 20
9 Indeks kelimpahan mangsa komodo di pulau besar dan pulau kecil 21
DAFTAR GAMBAR
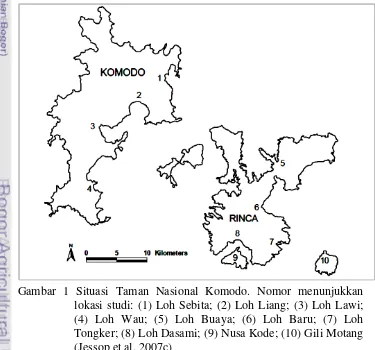
1 Situasi Taman Nasional Komodo. Nomor menunjukkan lokasi studi: (1) Loh Sebita; (2) Loh Liang; (3) Loh Lawi; (4) Loh Wau; (5) Loh Buaya; (6) Loh Baru; (7) Loh Tongker; (8) Loh Dasami; (9) Nusa Kode; (10) Gili
Motang 3
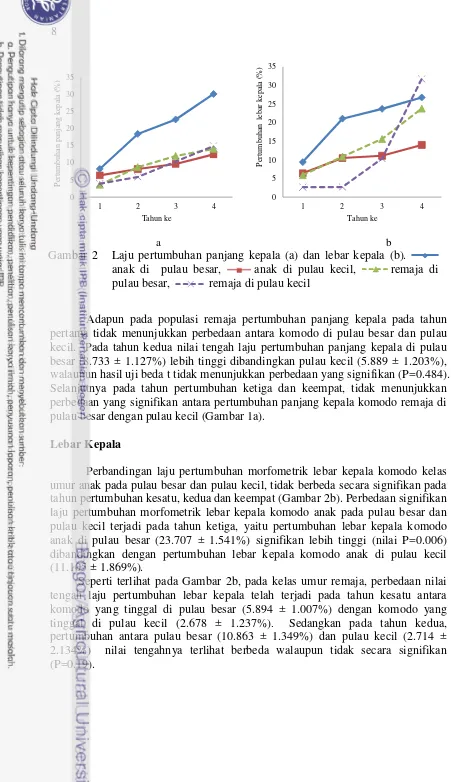
2 Laju pertumbuhan panjang kepala (a) dan lebar kepala (b). anak di pulau besar, anak di pulau kecil, remaja di pulau besar,
remaja di pulau kecil 8
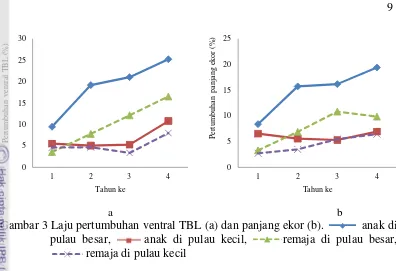
3 Laju pertumbuhan ventral TBL (a) dan panjang ekor (b). anak di pulau besar, anak di pulau kecil, remaja di pulau besar,
remaja di pulau kecil 9
4 Laju pertumbuhan lingkar ekor (a) dan berat tubuh (b). anak di pulau besar, anak di pulau kecil, remaja di pulau besar, remaja
di pulau keci 10
5 Laju pertumbuhan SVL anak di pulau besar, anak di pulau kecil, remaja di pulau besar, remaja di pulau keci 11 6 Peta situasi dan penutupan vegetasi di Taman Nasional Komodo 18
DAFTAR LAMPIRAN
1 Laju pertumbuhan jenis-jenis morfometrik kelas umur anak di lokasi-lokasi pulau besar dan pulau kecil pada setiap tingkat pertumbuhan 27 2 Laju pertumbuhan jenis-jenis morfometrik kelas umur remaja di
lokasi-lokasi pulau besar dan pulau kecil pada setiap tingkat pertumbuhan 37 3 Nilai tengah pertumbuhan morfometrik dan uji beda t pada kelas umur anak 41 4 Nilai tengah pertumbuhan morfometrik dan uji beda t pada kelas umur
1
PENDAHULUAN
Latar Belakang
Biawak komodo (Varanus komodoensis) adalah kadal terbesar di dunia (Auffenberg 1981). Oleh International Union for the Conservation of Nature and Natural Resources (IUCN) dikategorikan sebagai rentan atau vulnerable, karena penurunan demografis dan sebarannya yang terbatas. Selain itu berdasarkan peraturan di Indonesia, biawak komodo juga dilindungi berdasarkan beberapa peraturan yaitu Peraturan Binatang Liar tahun 1931, Surat Keputusan Menteri Kehutanan Nomor 301/Kpts-II/1991, dan Peraturan Pemerintah Nomor 7 tahun 1999 tentang Pengawetan Jenis Tumbuhan dan Satwa.
Penyebaran utama biawak komodo adalah di empat pulau dalam kawasan Taman Nasional Komodo (Jessop et al. 2003). Dua diantaranya merupakan pulau terbesar di Taman Nasional Komodo, yaitu Pulau Komodo (luas 31159 ha) dan Pulau Rinca (luas 20478 ha). Jumlah populasi biawak komodo di Pulau Komodo adalah 2921 ekor dan Pulau Rinca adalah 3238 ekor (BTNK 2013a). Sementara dua pulau lainnya memiliki luas yang lebih kecil yaitu Nusa Kode (luas 733 ha) dan Gili Motang (luas 948 ha), dengan jumlah populasi komodo pada masing-masing pulau adalah 99 ekor dan 65 ekor (BTNK 2013a).
Ukuran pulau dan isolasi akan mempengaruhi keragaman habitat, relung serta keberadaan sumberdaya habitat sehingga mempengaruhi spesies yang disangga oleh pulau tersebut (McArthur dan Wilson 1967, Blondel 2000). Akibatnya spesies di pulau kecil pada akhirnya akan menampakkan perubahan dibandingkan kerabatnya yang tinggal di pulau besar dan salah satu perubahan yang paling mendapat perhatian sehingga disebut sebagai island rule, adalah perubahan ukuran tubuh (Clegg dan Owen 2002).
Populasi komodo di pulau kecil memiliki ukuran tubuh dan berat tubuh yang lebih kecil dan lebih ringan dibandingkan kerabatnya di pulau besar (Jessop
et al. 2006). Akan tetapi penelitian tersebut hanya dilakukan dalam periode yang singkat yaitu tahun 2002 sampai 2005. Sementara pengukuran pertumbuhan morfometrik biawak komodo di Taman Nasional Komodo terus dilakukan sampai dengan tahun 2012. Oleh karena itu untuk memantau tren perubahan ukuran tubuh pada populasi komodo di pulau kecil, perlu dilakukan penelitian mengenai perbandingan antara pertumbuhan morfometrik biawak komodo di pulau besar dengan pulau kecil dalam periode pengamatan yang lebih panjang, yaitu mempergunakan data hasil pengukuran pertumbuhan tahun 2002 sampai 2012. Pemantauan tren perbedaan laju pertumbuhan biawak komodo di pulau besar dan pulau kecil juga perlu diikuti dengan pemantauan perbedaan karakteristik habitat pulau besar dan pulau kecil untuk mengungkapkan fakta-fakta mendasar yang diduga menjadi penyebab perbedaan laju pertumbuhan biawak komodo antara pulau besar dengan pulau kecil.
Perumusan Masalah
2
dilakukan pada 15% komodo terbesar yang tertangkap di pulau besar dan 15% komodo terbesar yang tertangkap di pulau kecil. Perbandingan ukuran satwa seharusnya dilakukan pada umur yang sama. Akan tetapi pendugaan umur komodo sampai saat ini belum diketahui. Oleh karena itu untuk membandingkan ukuran komodo di pulau besar dan pulau kecil maka penelitian ini melakukan pendekatan dengan mengukur laju pertumbuhan.
Selain itu adanya variasi ukuran tubuh yang berbeda antar pulau (Jessop et al. 2006), berimplikasi pada kategori umur berdasarkan ukuran antar pulau juga berbeda, terutama untuk pulau besar dan pulau kecil. Bisa jadi individu-individu komodo yang diukur di pulau kecil merupakan komodo dengan umur yang tua sementara komodo yang diukur di pulau besar memang komodo dengan umur lebih muda. Oleh karena itu penelitian ini menganalisis laju pertumbuhan biawak komodo berdasarkan kelas umur, yaitu anak dan remaja, dengan asumsi laju pertumbuhan setiap kelas umur adalah berbeda.
Tujuan Penelitian
Tujuan dari penelitian ini adalah mengungkapkan fakta perbedaan ukuran antara populasi biawak komodo yang tinggal di pulau besar dengan pulau kecil, dengan cara:
1. Membandingkan pertumbuhan morfometrik biawak komodo pada populasi komodo di pulau besar dan populasi komodo di kecil.
2. Membandingkan model pertumbuhan morfometrik biawak komodo di pulau besar dan populasi komodo di kecil.
3. Membandingkan karakteristik habitat biawak komodo di pulau besar dan di pulau kecil.
Hipotesis Penelitian
Hipotesis dari penelitian ini adalah:
1. Pertumbuhan morfometrik biawak komodo yang hidup di pulau kecil, lebih lambat dibandingkan pertumbuhan morfometrik biawak komodo di pulau besar. 2. Model pertumbuhan morfometrik biawak komodo yang hidup di pulau besar berbeda dengan model pertumbuhan morfometrik biawak yang hidup di pulau kecil.
3. Terdapat perbedaan karakteristik habitat komodo di pulau besar dan pulau kecil yang mendorong perbedaan ukuran biawak komodo.
Manfaat Penelitian
Penelitian ini bermanfaat untuk menunjukkan bahwa populasi di pulau kecil terisolasi bila terindikasi terdapat penurunan ukuran tubuh dan keterbatasan pakan, perlu mendapat perhatian khusus, melalui manipulasi habitat maupun populasi. Agar jumlah populasi efektif terpelihara untuk menghindari peningkatan
3
2
METODE
Lokasi dan Waktu
Pengumpulan data morfometrik serta ekologi populasi biawak komodo di Taman Nasional Komodo dilakukan di Kantor Balai Taman Nasional Komodo yang berada di Labuan Bajo, Flores. Penelitian dilaksanakan selama tiga bulan yaitu bulan Januari sampai Maret 2015.
Bahan Penelitian
Penelitian ini dilaksanakan dengan mengolah data yang tersedia dari hasil studi capture mark recapture biawak komodo di Taman Nasional Komodo tahun 2002 sampai 2012 (Tabel 1). Selain itu dipergunakan hasil studi lainnya yaitu data populasi, penutupan tipe ekosistem, mangsa dan genetika. Ijin tertulis untuk akses dan penggunaan data hasil studi capture mark recapture biawak komodo diperoleh dari Kepala Balai Taman Nasional Komodo dan Ketua Program Yayasan Komodo Survival Program.
Studi capture mark recapture biawak komodo di Taman Nasional Komodo pada tahun 2002 sampai 2012 dilaksanakan pada kawasan Taman Nasional Komodo di Nusa Tenggara Timur, pada dua pulau besar yaitu Pulau Rinca dan
4
Data yang dipergunakan mencakup data individu biawak komodo yang dua Tabel 1 Jenis dan sumber data yang menjadi bahan penelitian
Jenis data Sumber data
Lokasi studi Waktu studi Pelaksana studi
Morfometrik Studi
capture 10 lokasi. Dua lokasi mewakili pulau kecil yaitu Gili Motang dan Nusa kode. Delapan lokasi mewakili pulau besar yaitu empat lokasi di Pulau Komodo (Loh Liang, Loh Lawi, Loh Sebita dan Loh Wau) dan empat lokasi di Pulau Rinca (Loh Buaya,
Tahun 2006 Rudiharto (2007)
Penutupan
5 Pulau Komodo serta dua pulau kecil yaitu Gili Motang dan Nusa Kode. Studi pada Pulau Rinca dilaksanakan pada empat lokasi yaitu Loh Buaya, Loh Baru, Loh Tongker dan Loh Dasami. Sedangkan studi di Pulau Komodo dilaksanakan pada lokasi Loh Sebita, Loh Liang, Loh Lawi dan Loh Wau (Gambar 1). Pemilihan empat lokasi pada masing-masing pulau besar ditentukan dengan mempertimbangkan frekuensi perjumpaan komodo yang relatif tinggi dan keterjangkauan medan. Sementara pemilihan lokasi studi di pulau kecil ditujukan sebagai perwakilan populasi kecil di pulau kecil.
Studi capture mark recapture dimulai dengan memasang perangkap pada titik-titik yang tetap di lokasi-lokasi studi sepanjang masa studi. Pengumpanan dilakukan dengan daging kambing yang disimpan dalam perangkap serta digantung di atas perangkap untuk memancing komodo masuk dalam perangkap. Komodo yang tertangkap diukur morfometriknya dan diberi penanda berupa PIT (passive integrated transporder) Trovan Inc. yang berbentuk silinder kaca, sebagai identitas setiap individu yang tertangkap. Pemasangan penanda dilakukan dengan cara disuntikkan antara kulit dan daging pada tungkai belakang sebelah kanan. Setelah itu komodo tersebut dilepaskan kembali. Pada periode studi selanjutnya (tahun berikutnya), setiap komodo yang tertangkap akan dipindai sebelah tungkai kanan belakangnya dengan alat pembaca khusus. Apabila pemindai menampilkan kombinasi angka dan huruf tertentu, berarti komodo tersebut pernah tertangkap sebelumnya. Komodo yang pernah tertangkap tersebut, kembali dilakukan pengukuran morfometrik untuk mengetahui laju pertumbuhannya.
Setiap individu komodo memiliki peluang yang sama untuk tertangkap kembali setiap tahunnya, walau demikian terindikasi adanya proses belajar dari komodo dalam mengatasi perangkap. Selain itu komodo dalam keadaan kenyang tidak akan tertarik pada umpan dalam perangkap. Demikian pula bila cuaca hujan maka komodo cenderung akan berlindung, oleh karena itu studi ini dijadwalkan pada waktu yang diperkirakan musim kemarau.
Data yang dipergunakan pada penelitian ini mencakup data individu biawak komodo yang tertangkap, tertandai dan tertangkap kembali, meliputi informasi tanggal, lokasi, kelas umur dan morfometrik. Pada penelitian ini tidak diperhitungkan jenis kelamin karena pada satwa komodo tidak ada penanda morfologi luar yang pasti untuk membedakan jantan dan betina, sehingga laju pertumbuhan morfometrik diasumsikan sama untuk jenis kelamin jantan dan betina.
Kriteria penentuan kelas umur didasarkan pada ukuran panjang moncong hingga kloaka/snout to vent lenght (SVL). Kelas umur tetasan adalah komodo dengan SVL kurang dari 35 cm. Kelas umur anak berukuran SVL 26 cm sampai 80 cm. Kelas umur remaja mempunyai panjang SVL 81 cm sampai 110 cm. Sedangkan kelas umur dewasa adalah komodo dengan ukuran SVL lebih dari 110 cm. Pada penelitian ini laju pertumbuhan yang dianalisis hanya kelas umur anak dan remaja, karena keterbatasan jumlah data komodo kelas umur tetasan dan dewasa di pulau kecil.
6
Pengertian masing-masing morfometrik sebagaimana disebutkan dalam Jessop et al. (2007c) sebagai berikut:
1. Panjang kepala: panjang dari ujung ujung moncong sampai ke tulang leher pertama
2. Lebar kepala: tulang dahi kiri ke tulang dahi kanan
3. Panjang moncong: panjang dari rahang sampai ujung moncong
4. Panjang moncong hingga pangkal ekor: panjang dari ujung moncong sampai ke kloaka
5. Total panjang tubuh bagian ventral: panjang dari ujung moncong sampai ke ujung ekor
6. Panjang ekor: panjang dari kloaka ke ujung ekor
7. keliling pangkal ekor: keliling lingkaran tepat di belakang kloaka, pada bagian paling gemuk
8. Berat badan: penimbangan berat badan komodo
Prosedur Analisis Data
Dalam penelitian ini telah dianalisis morfometrik komodo kelas umur anak dan remaja masing-masing sebanyak 353 ekor dan 163 ekor yang tinggal di pulau besar, serta 46 ekor komodo anak dan 14 ekor komodo remaja yang tinggal di pulau kecil (Lampiran 1 dan 2). Laju pertumbuhan morfometrik dihitung dengan persamaan sebagai berikut (Krebs 1999):
�
=
�2−�1�1 X 100%Keterangan : H1 = ukuran morfometrik awal
H2 = ukuran morfometrik setelah 1,2,3 atau 4 tahun G = persentase pertumbuhan morfometrik
Pengukuran morfometrik pada satwaliar komodo tidak selalu berjalan sempurna sehingga kadang tidak semua pengukuran terlaksana dengan baik. Seperti terlihat pada Lampiran 1 dan 2, dalam satu data individu kerap tidak dapat dihitung pertumbuhan semua jenis morfometriknya. Periode waktu pertumbuhan yang dapat dianalisis adalah satu, dua, tiga dan empat tahun. Singkatnya periode pertumbuhan ini disebabkan rendahnya ukuran populasi komodo di pulau kecil sehingga peluang tertangkapnya komodo juga lebih rendah dibanding pulau besar dan data yang diperoleh pun terbatas.
Diasumsikan setiap individu dapat mewakili lebih dari satu periode pertumbuhan untuk setiap jenis morfometrik, sehingga jumlah unit contoh setiap periode berkisar antara 4 sampai 147. Oleh karena itu perbandingan persentase pertumbuhan dilakukan melalui uji beda t bagi perbandingan rata-rata dua kelompok yang berbeda (Walpole 1995). Uji beda dilakukan terhadap kelompok data jenis morfometrik serta periode pertumbuhan yang sama antara pulau besar dengan pulau kecil. Analisis uji beda t dijalankan dengan program SPSS 16.
Hipotesis yang akan diuji adalah:
H0: x1- x2 = 0
7
Keterangan: x1 = rata-rata laju pertumbuhan morfometrik biawak komodo
pada periode pertumbuhan tertentu di pulau besar
x2 = rata-rata laju pertumbuhan morfometrik biawak komodo
pada periode pertumbuhan tertentu di pulau kecil
Uji korelasi pearson dilakukan untuk melihat korelasi antara pertumbuhan jenis morfometrik dengan pertambahan umur. Kemudian dibangun model pendugaan pertambahan umur komodo dengan variabel bebas persentase pertumbuhan jenis morfometrik yang berkorelasi dengan pertambahan umur. Model regresi selanjutnya diuji secara parsial pada masing-masing koefisien regresi mempergunakan uji t, serta secara bersama mempergunakan uji analysis of varians dan menilai kesesuaian model mempergunakan koefisien determinasi. Pada penelitian ini pembangunan model hanya dapat dilakukan pada populasi komodo kelas umur anak, karena pada kelas umur lainnya terdapat keterbatasan data pada populasi komodo di pulai kecil.
Karakteristik habitat dianalisis secara deskriptif melalui perbandingan habitat biawak komodo pada pulau besar maupun pulau kecil. Adapun unsur habitat yang dianalisis meliputi ukuran dan jarak antar pulau, tipe ekosistem, keberadaan pemukiman dan aktivitas wisata, serta ketersediaan mangsa. Perbedaan karakteristik habitat antara pulau besar dan pulau kecil dipergunakan untuk menelaah faktor-faktor penyebab perbedaan pertumbuhan morfometrik biawak komodo.
3
HASIL DAN PEMBAHASAN
Perbandingan Laju Pertumbuhan antara Pulau Besar dan Pulau Kecil
Panjang Kepala
8
Adapun pada populasi remaja pertumbuhan panjang kepala pada tahun pertama tidak menunjukkan perbedaan antara komodo di pulau besar dan pulau kecil. Pada tahun kedua nilai tengah laju pertumbuhan panjang kepala di pulau besar (8.733 ± 1.127%) lebih tinggi dibandingkan pulau kecil (5.889 ± 1.203%), walaupun hasil uji beda t tidak menunjukkan perbedaan yang signifikan (P=0.484). Selanjutnya pada tahun pertumbuhan ketiga dan keempat, tidak menunjukkan perbedaan yang signifikan antara pertumbuhan panjang kepala komodo remaja di pulau besar dengan pulau kecil (Gambar 1a).
Lebar Kepala
Perbandingan laju pertumbuhan morfometrik lebar kepala komodo kelas umur anak pada pulau besar dan pulau kecil, tidak berbeda secara signifikan pada tahun pertumbuhan kesatu, kedua dan keempat (Gambar 2b). Perbedaan signifikan laju pertumbuhan morfometrik lebar kepala komodo anak pada pulau besar dan pulau kecil terjadi pada tahun ketiga, yaitu pertumbuhan lebar kepala komodo anak di pulau besar (23.707 ± 1.541%) signifikan lebih tinggi (nilai P=0.006) dibandingkan dengan pertumbuhan lebar kepala komodo anak di pulau kecil (11.105 ± 1.869%).
Seperti terlihat pada Gambar 2b, pada kelas umur remaja, perbedaan nilai tengah laju pertumbuhan lebar kepala telah terjadi pada tahun kesatu antara komodo yang tinggal di pulau besar (5.894 ± 1.007%) dengan komodo yang tinggal di pulau kecil (2.678 ± 1.237%). Sedangkan pada tahun kedua,
9
Total Tubuh Bagian Ventral (Ventral TBL)
Perbandingan laju pertumbuhan ukuran panjang total tubuh bagian ventral (ventral TBL) pada komodo kelas umur anak di pulau besar dan kecil tidak menunjukkan perbedaan signifikan pada tahun kesatu dan kedua (Gambar 3a). Perbedaan signifikan mulai terjadi pada pertumbuhan ventral TBL antara komodoanak di pulau besar dan pulau kecil adalah pada tahun ketiga dan keempat. Pada tahun keempat pertumbuhan ventral TBL komodo anak di pulau besar mencapai 25.218 ± 2.61%, signifikan lebih tinggi (P=0.007) dibandingkan pertumbuhan ventral TBL komodo anak di pulau kecil (10.797 ± 1.857%).
Sebagaimana terlihat pada Gambar 3a, pada komodo remaja pertumbuhan tahun ketiga pada pulau besar (12.140 ± 1.406%) lebih tinggi dibanding pulau kecil (3.347 ± 3.002%), tetapi tidak berbeda signifikan (P=0.48).
Panjang Ekor
Pertumbuhan panjang ekor komodo kelas umur anak di pulau besar (8.394 ± 1.067%) tidak signifikan berbeda dengan pertumbuhan panjang ekor komodo anak pada pulau kecil (6.541 ± 1.285%) pada tahun pertama. Tetapi pada tahun kedua, ketiga dan keempat pertumbuhan panjang ekor komodo anak di pulau besar dan pulau kecil berbeda secara signifikan. Pertumbuhan panjang ekor komodo anak di pulau besar lebih tinggi dibandingkan pertumbuhan biawak komodo di pulau kecil (Gambar 3b).
Sementara pada komodo kelas umur remaja, laju pertumbuhannya di pulau besar (12.140 ± 1.406%) pada tahun ketiga lebih tinggi dibanding pulau kecil (3.347 ± 3.002%), tetapi tidak berbeda signifikan dengan nilai P=0.426 (Gambar 3b). Demikian pula dengan pertumbuhan tahun kesatu, kedua dan ketiga juga tidak menunjukkan perbedaan signifikan.
a b
Gambar 3 Laju pertumbuhan ventral TBL (a) dan panjang ekor (b). anak di pulau besar, anak di pulau kecil, remaja di pulau besar,
10
Lingkar Ekor
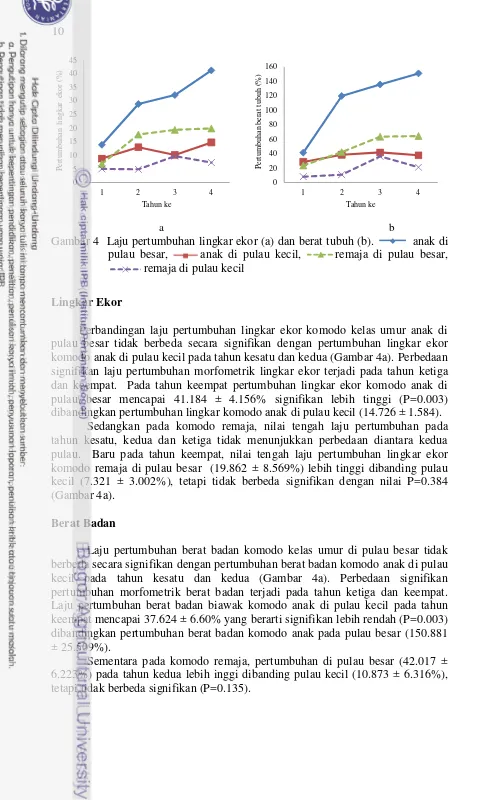
Perbandingan laju pertumbuhan lingkar ekor komodo kelas umur anak di pulau besar tidak berbeda secara signifikan dengan pertumbuhan lingkar ekor komodo anak di pulau kecil pada tahun kesatu dan kedua (Gambar 4a). Perbedaan signifikan laju pertumbuhan morfometrik lingkar ekor terjadi pada tahun ketiga dan keempat. Pada tahun keempat pertumbuhan lingkar ekor komodo anak di pulau besar mencapai 41.184 ± 4.156% signifikan lebih tinggi (P=0.003) dibandingkan pertumbuhan lingkar komodo anak di pulau kecil (14.726 ± 1.584). Sedangkan pada komodo remaja, nilai tengah laju pertumbuhan pada tahun kesatu, kedua dan ketiga tidak menunjukkan perbedaan diantara kedua pulau. Baru pada tahun keempat, nilai tengah laju pertumbuhan lingkar ekor komodo remaja di pulau besar (19.862 ± 8.569%) lebih tinggi dibanding pulau kecil (7.321 ± 3.002%), tetapi tidak berbeda signifikan dengan nilai P=0.384 (Gambar 4a).
Berat Badan
Laju pertumbuhan berat badan komodo kelas umur di pulau besar tidak berbeda secara signifikan dengan pertumbuhan berat badan komodo anak di pulau kecil pada tahun kesatu dan kedua (Gambar 4a). Perbedaan signifikan pertumbuhan morfometrik berat badan terjadi pada tahun ketiga dan keempat. Laju pertumbuhan berat badan biawak komodo anak di pulau kecil pada tahun keempat mencapai 37.624 ± 6.60% yang berarti signifikan lebih rendah (P=0.003) dibandingkan pertumbuhan berat badan komodo anak pada pulau besar (150.881 ± 25.509%).
Sementara pada komodo remaja, pertumbuhan di pulau besar (42.017 ± 6.223%) pada tahun kedua lebih inggi dibanding pulau kecil (10.873 ± 6.316%), tetapi tidak berbeda signifikan (P=0.135).
a b
Gambar 4 Laju pertumbuhan lingkar ekor (a) dan berat tubuh (b). anak di pulau besar, anak di pulau kecil, remaja di pulau besar,
11
Panjang Moncong Hingga Pangkal Ekor/Snout to Vent Lenght (SVL)
Penelitian ini memperlihatkan bahwa laju pertumbuhan tahun kesatu morfometrik SVL komodo kelas umur anak di pulau besar lebih tinggi dibandingkan laju pertumbuhan SVL komodo anak di pulau kecil, tetapi tidak menunjukkan perbedaan yang signifikan (Gambar 5). Laju pertumbuhan SVL pada pulau besar di tahun kesatu adalah 10.216 ± 0.657% sedangkan laju pertumbuhan SVL pada pulau kecil adalah 7.569 ± 1.213%. Perbandingan laju pertumbuhan SVL komodo anak pada pulau besar dan pulau kecil mengalami perbedaan yang signifikan pada tahun kedua, ketiga dan keempat. Perbedaan paling signifikan terjadi pada pertumbuhan tahun ketiga. Laju pertumbuhan SVL komodo anak di pulau besar (24.237 ± 1.159%) signifikan lebih tinggi daripada pertumbuhan SVL komodo anak di pulau kecil (8.630 ± 2.066%) dengan nilai P=0.
Pada komodo remaja, laju pertumbuhan SVL di pulau kecil (6.301 ± 3.462%) lebih rendah dibandingkan pulau besar (9.304 ± 1.079%) pada tahun kedua, tetapi tidak berbeda signifikan (P=0.496). Sedangkan pada tingkat waktu pertumbuhan yang lain, nilai tengah laju pertumbuhan SVL komodo remaja tidak menunjukkan perbedaan antara pulau besar dan pulau kecil.
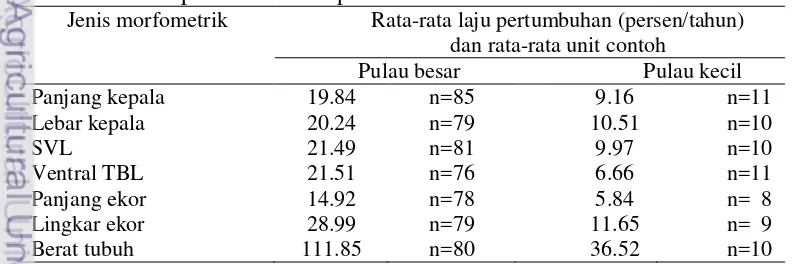
Tabel 2 Rata-rata laju pertumbuhan morfometrik biawak komodo kelas umur anak di pulau besar dan pulau kecil
Jenis morfometrik Rata-rata laju pertumbuhan (persen/tahun) dan rata-rata unit contoh
Pulau besar Pulau kecil Panjang kepala 19.84 n=85 9.16 n=11
remaja di pulau besar, remaja di pulau kecil
12
Secara keseluruhan pada semua jenis morfometrik biawak komodo kelas umur anak, rata-rata laju pertumbuhan pertahun di pulau besar lebih tinggi dibandingkan pulau kecil (Tabel 2). Rata-rata laju pertumbuhan SVL biawak komodo di pulau besar adalah 21.49 persen/tahun yang berarti 215.55% dibanding pulau kecil (9.97 persen/tahun). Demikian juga dengan rata-rata laju pertumbuhan lebar kepala, panjang kepala, ventral TBL, panjang ekor, lingkar ekor dan berat badan, berturut-turut nilai perbandingannya 192.58%, 216.59%, 322.97%, 255.48%, 248.84%, 306.27% pada pulau besar dibandingkan rata-rata laju pertumbuhan pertahun di pulau kecil. Pada kelas umur anak, laju pertumbuhan morfometrik yang paling pesat adalah berat tubuh, yang terjadi pada komodo anak di pulau besar maupun pulau kecil. Sedangkan yang paling rendah laju pertumbuhannya adalah pada panjang ekor.
Sama halnya dengan kelas umur anak, pada kelas umur remaja laju pertumbuhan morfometrik yang paling pesat pada pulau besar dan kecil juga adalah berat tubuh. Sementara secara keseluruhan, rata-rata laju pertumbuhan pada semua jenis morfometrik pertahun di pulau besar lebih tinggi dibandingkan pulau kecil (Tabel 2). Rata-rata laju pertumbuhan SVL biawak komodo di pulau besar adalah 11.20 persen/tahun yang berarti 146.21% lebih besar dibanding pulau kecil (7.66 persen/tahun). Demikian juga dengan rata-rata laju pertumbuhan lebar kepala, panjang kepala, ventral TBL, panjang ekor, lingkar ekor dan berat badan, berturut-turut nilai perbandingannya 109.45%, 118.28%, 193.99%, 170.42%, 237.11%, 253.78% pada pulau besar dibandingkan rata-rata laju pertumbuhan pertahun di pulau kecil. Dengan demikian terlihat bahwa perbedaan laju pertumbuhan antara pulau besar dibanding pulau kecil, pada kelas umur anak lebih besar dibandingkan kelas umur remaja.
Persentase pertumbuhan morfometrik yang lambat pada populasi komodo di pulau kecil sejalan dengan studi Jessop et al. (2005). Pada populasi yang sama dengan data pengamatan tahun 1994, 2002 sampai 2004, Jessop et al. (2005) menyatakan pertumbuhan SVL biawak komodo di Pulau Gili Motang (salah satu pulau kecil) sebesar 0.514±0.086 cm/tahun, yang berarti 49.54% lebih kecil dibandingkan pertumbuhan SVL biawak komodo di Pulau Rinca (salah satu pulau besar) yang tercatat sebesar 1.019±0.192 cm/tahun. Sementara hasil penelitian ini menunjukkan perbedaan yang lebih besar antara laju pertumbuhan SVL antara pulau besar dengan pulau kecil, yaitu laju pertumbuhan SVL biawak komodo di pulau kecil (Nusa Kode dan Gili Motang) 115.54% lebih kecil dibandingkan laju
Tabel 3 Rata-rata laju pertumbuhan morfometrik biawak komodo kelas umur remaja di pulau besar dan pulau kecil
Jenis morfometrik Rata-rata laju pertumbuhan (persen/tahun) dan rata-rata unit contoh
Pulau besar Pulau kecil
Panjang kepala 9.61 n=38 8.78 n=3
Lebar kepala 14.04 n=31 11.87 n=2
SVL 11.20 n=34 7.66 n=2
Ventral TBL 10.01 n=31 5.16 n=2
Panjang ekor 7.72 n=31 4.53 n=2
Lingkar ekor 15.91 n=30 6.71 n=2
13 pertumbuhan SVL biawak komodo di pulau besar. Hal ini dikarenakan studi Jessop et al. (2005) dilakukan pada populasi komodo pada kelas umur dewasa, sedangkan penelitian ini dilakukan pada kelas umur anak. Laver et al. (2012) menyebutkan laju pertumbuhan biawak komodo pada kelas umur anak adalah paling cepat dibandingkan laju pertumbuhan biawak komodo kelas umur lainnya.
Penelitian ini menunjukkan pada kelas umur anak, laju pertumbuhan biawak komodo di pulau kecil signifikan lebih rendah dibandingkan laju pertumbuhan biawak komodo di pulau besar. Hal tersebut diduga akan mempengaruhi ukuran tubuh maksimal pada saat dewasa. Pada populasi biawak komodo yang sama dengan data pengukuran morfometrik tahun 2002 sampai 2004, Jessop et al. (2006) menyatakan rata-rata SVL pada 15% biawak komodo terbesar di Pulau komodo adalah 145.61 ± 0.83 cm, lebih panjang dibandingkan rata-rata SVL pada 15% biawak komodo terbesar di Gili Motang adalah 96.91 ± 1.19 cm. Biawak komodo terpanjang terdapat di Pulau Komodo dengan SVL 154.05 cm. Sedangkan rata-rata dari 15% biawak komodo terbesar di Pulau Gili Motang berukuran sedang (SVL antara 60 sampai120 cm). Sedangkan untuk ukuran berat tubuh, diketahui biawak komodo pulau besar signifikan lebih besar dan berat dibandingkan dengan biawak komodo di pulau kecil. Rata-rata berat badan biawak komodo di Pulau Komodo adalah 66.39 ± 3.06 kg, sedangkan di Pulau Gili Motang adalah 13.50 ± 1.06 kg. Dari studi tersebut dapat disimpulkan bahwa biawak komodo di pulau-pulau kecil Gili Motang dan Nusa Kode mempunyai ukuran tubuh maksimal yang lebih kecil dibandingkan dengan ukuran maksimal biawak komodo di pulau- pulau besar Komodo dan Rinca.
Pertumbuhan morfometrik pada populasi komodo di pulau kecil yang yang lebih lambat dibandingkan kerabatnya di pulau besar, sejalan dengan Raia et al. (2010) pada studi terhadap kadal biru spandrel yang tinggal di pulau kecil di Selatan Italia terdeteksi memiliki laju pertumbuhan yang rendah dibanding kerabatnya di pulau besar, karena populasi di pulau kecil menghadapi keadaan lingkungan ekstrim yang tak terduga. Kadal-kadalan merupakan kelompok satwa yang terdokumentasi mengalami endemisitas sebagai akibat dari terisolasi dalam pulau (Whittaker 1998). Schall (1983) menyebutkan bahwa perubahan ukuran tubuh pada kadal-kadalan yang tinggal di pulau merupakan salah satu perubahan sifat populasi pada pulau yang terisolasi.
Perbandingan Model Laju Pertumbuhan Morfometrik pada Pulau Besar dan Pulau Kecil
14
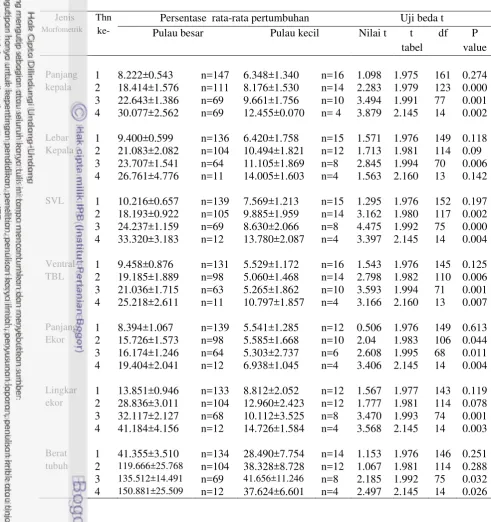
dengan pertambahan umur di pulau besar menunjukkan nilai di atas 0.5, kecuali pada ventral TBL (r=0.236), dengan tingkat kepercayaan 0.01. Sementara nilai korelasi pearson pada korelasi persentase pertumbuhan ukuran morfometrik dengan pertambahan umur di pulau kecil menunjukkan nilai di bawah 0.5 kecuali lebar kepala (r=0.544), dengan tingkat kepercayaan 0.01 dan 0.05 (Tabel 4 dan 5).
Pada populasi komodo di pulau besar, korelasi paling tinggi dalam hubungan antara persentasi pertumbuhan jenis morfometrik dengan pertambahan umur adalah pada morfometrik SVL (r=0.742). Setelah SVL, berturut-turut dari yang korelasinya tertinggi hingga terendah adalah ukuran morfometrik adalah panjang kepala (r=0.704), lingkar ekor (r=0.680), lebar kepala (r=0.663), berat badan (r=0.580), panjang ekor (r=0.577) dan ventral TBL (r=0.239). Pada populasi komodo di pulau besar sejalan dengan studi Meiri (2010) mengenai allometri kadal-kadalan termasuk genus Varanus, menyimpulkan bahwa SVL merupakan ukuran yang baik untuk mengukur pertumbuhan tubuh.
Sedangkan pada populasi komodo di pulau kecil, jenis ukuran morfometrik yang paling kuat korelasinya dengan pertambahan umur adalah lebar kepala (r=0.544). Selanjutnya jenis ukuran morfometrik berikut ini berturut-turut menunjukkan korelasi yang semakin lemah dengan pertambahan umur, yaitu panjang kepala (r=0.492), SVL (r=0.433), ventral TBL (r=0.354), panjang ekor (r=0.354), berat badan (r=0.361) dan lingkar ekor (r=0.341).
Tabel 5 Perbandingan korelasi antara pertambahan umur dengan laju pertumbuhan morfometrik vebtral TBL, panjang ekor, lingkar ekor dan berat tubuh
*korelasi signifikan pada tingkat kepercayaan 0.01 ** korelasi signifikan pada tingkat kepercayaan 0.05
Tabel 4 Perbandingan korelasi antara pertambahan umur dengan laju pertumbuhan morfometrik panjang kepala, lebar kepala dan SVL
Panjang kepala
Lebar
kepala SVL
Pulau besar Pertumbuhan tahun ke
Pearson
Correlation 0.704** 0.663** 0.742** Sig. (2-tailed) 0 0 0
N 467 443 452
Pulau kecil Pertumbuhan tahun ke
Pearson
Correlation 0.337* 0.385* 0.239 Sig. (2-tailed) 0.025 0.016 0.132
N 44 39 41
15
Persentase pertumbuhan lingkar pangkal ekor dan berat badan pada populasi komodo di pulau kecil menunjukkan korelasi yang rendah terhadap pertambahan umur. Pangkal ekor merupakan tempat penimbunan lemak yang berfungsi sebagai cadangan makanan. Komodo dapat makan hingga 80% berat tubuhnya (Aufenberg 1981), oleh karena itu lingkar pangkal ekor dan berat badan dapat berfluktuasi tergantung kecukupan pakan.
Model regresi untuk menduga pertambahan umur pada biawak komodo dapat mempergunakan variabel bebas semua jenis persentase pertumbuhan morfometrik, karena semua jenis persentase pertumbuhan morfometrik tersebut berkorelasi positif dengan pertambahan umur. Pada populasi komodo di pulau besar, semua jenis persentase morfometrik dapat dipergunakan untuk memprediksi pertambahan umur. Hal ini ditunjukkan dengan nilai P value pada model regresi yang dibangun untuk semua variabel bebas persentase ukuran morfometrik adalah signifikan (Pvalue<0.000). Sementara pada populasi komodo di pulau kecil, model regresi dengan variabel bebas persentase pertumbuhan lingkar ekor dan berat badan menunjukkan nilai P value >0.05 (Tabel 6 dan 7). Tabel 7 Model regresi pendugaan pertambahan umur dari variabel persentase
pertumbuhan morfometrik ventral TBL, panjang ekor, lingkar ekor dan berat tubuh
Ventral TBL Panjang ekor Lingkar ekor Berat tubuh
Pulau
besar R square 5.7 33.3 46.3 33.6
Model Y=1.377+6.877X Y=2.905+0.9447X Y=1.576+ 3.9027X Y=2.294+0.4287X
t hitung 21.398 24.492 13.987 21.719
Pvalue <0.001 <0.001 <0.001 <0.001
Pulau
kecil R square 12.5 3.6 11.6 13.1
Model Y=1.419 +.829X Y=1.723 +6.1749X Y=1.691 +4.211X Y=1.68 +1.165X
t hitung 3.707 7.923 5.842 6.632
Pvalue <0.001 0.017 0.039 0.024
Tabel 6 Model regresi pendugaan pertambahan umur dari variabel persentase pertumbuhan morfometrik panjang kepala, lebar kepala dan SVL
Panjang kepala Lebar kepala SVL
Pulau
besar R square 49.6 43.9 55.1
Model Y=1.377 +6.877X Y=1.577 +5.621X Y=1.331 +6.182X t hitung 21.398 13.8 12.614
Pvalue <0.001 <0.001 <0.001 Pulau
kecil R square 24.2 29.6 18.8
Model Y=1.419 +7.829X Y=1.375 + .028X Y=1.48 + .222X t hitung 3.707 5.863 5.85
16
Nilai koefisien determinasi (nilai R square) model penduga umur pada populasi komodo di pulau besar memiliki nilai di atas 30%, kecuali pada persentase pertumbuhan ventral TBL (R2=5.7%). Sedangkan koefisien determinasi model penduga umur pada populasi komodo di pulau kecil, semuanya memiliki nilai di bawah 30%.
Pada populasi komodo di pulau besar, nilai koefisien determinasi (nilai R square) yang paling tinggi dalam model penduga pertambahan umur, adalah pada variabel bebas persentase pertumbuhan morfometrik SVL, yaitu sebesar 0.551. Hal ini menunjukkan bahwa pendugaan pertambahan umur biawak komodo sebesar 55.1 % dapat diduga dari persentasi pertumbuhan morfometrik SVL dan 44.9 % dipengaruhi oleh faktor lain, dengan model regresi linier Y=1.331 + 6.182X (t=21.398, P<0.000). Sedangkan pada populasi komodo di pulau kecil nilai koefisien determinasi paling tinggi (R2=29.,6 %) adalah pada model pendugaan pertambahan umur dengan variabel persentase pertumbuhan lebar kepala, dengan model regresi linear Y=1.419 + 7.829X (t=3.707, P<0.000).
Model pertumbuhan komodo yang terbentuk adalah model linier. Dalam penelitian ini data yang diolah adalah data komodo kelas umur juvenil saja. Hal ini sejalan dengan studi Laver et al. (2012), bahwa awal hidupnya, pertumbuhan tubuh komodo adalah linear dengan pertambahan umurnya dan laju pertumbuhan tertinggi adalah pada kelas umur juvenile. Pada kelas umur ini mahluk hidup banyak mengalokasikan energinya untuk pertumbuhan ukuran tubuh, lalu pada umur tertentu akan mengalihkan energinya untuk reproduksi.
Pada populasi biawak komodo yang sama dengan data tahun 2002 s.d. 2010, berdasarkan model pertumbuhan yang dibangun oleh Laver et al. (2012), disimpulkan bahwa laju pertumbuhan terendah adalah di pulau kecil yaitu Nusa Kode (0.132 ± 2.341 cm/tahun) dan Gili Motang (2.883 ± 0.495 cm/tahun). Sementara laju pertumbuhan tertinggi adalah lokasi di pulau besar yaitu Loh Baru (5.974 ± 0.358 cm/tahun). Laju pertumbuhan ini berhubungan dengan kepadatan populasi dengan bentuk model kurva terbalik dan tidak menunjukkan hubungan dengan koefisien inbreeding dan kelimpahan mangsa. Pernyataan ini sejalan untuk kasus populasi biawak komodo di pulau besar, bahwa pada populasi yang pertumbuhan tubuhnya tergantung pada kepadatan maka akan berimplikasi pada kondisi kompetisi intraspesifik, interaksi sosial agonistik yang selanjutnya menurunkan kemampuan jelajah individu (Kolbe 2008, Bjorndal 2000).
Penelitian ini menunjukkan pertumbuhan biawak komodo di Pulau Rinca lebih tinggi dibandingkan pertumbuhan di Pulau Komodo, sementara Purwandana
et al. (2014) menyebutkan kepadatan populasi di Pulau Rinca (14.20 ± 1.12 ekor/km2) lebih tinggi dibandingkan kepadatan populasi di Pulau Komodo (12.53 ± 1.13 ekor/km2). Dengan demikian dapat diperkirakan bahwa persaingan intraspesifik di Pulau Rinca lebih tinggi dibandingkan dengan Pulau Komodo. Demikian pula dengan pergerakan individu, Sastrawan dan Ciofi (2002) menyebutkan terdapat korelasi positif antara ukuran home range dengan SVL maupun berat tubuh. Jessop et al. (2007a) juga menyatakan bahwa pertumbuhan ukuran tubuh berkorelasi signifikan dengan pertambahan ukuran wilayah jelajah.
17 populasi komodo di pulau kecil lebih rendah dibandingkan dengan pulau besar (Gili Motang 4.05 ± 0.81 ekor/km2 dan Nusa Kode 14.02 ± 4.85 ekor/km2). Laver et al. (2012) menyebutkan fenomena ini sebagai Alle effect yang timbul karena inbreeding dan hilangnya gen heterozigot, perubahan demografi seperti fluktuasi sex ratio dan kurangnya interaksi antar individu sehingga menurunkan peluang reproduksi (Chourchamp et al. 1999). Dengan kondisi jumlah populasi yang sedikit dan rendahnya variasi genetik maka Arida (2011) menggambarkan populasi komodo di pulau kecil sebagai populasi yang menuju kepunahan lokal.
Dalam model sindrom pulau (Alder dan Levin 1994), populasi pulau akan mengarah pada peningkatan kepadatan, mendorong pada seleksi calon keturunan yang berukuran besar dan tumbuh menjadi individu dewasa berukuran besar. Kasus populasi komodo pada pulau kecil dalam penelitian ini menunjukkan kondisi sebaliknya dari model sindrom pulau. Sejalan dengan yang dinyatakan oleh sisi Raia et al. (2010) yang menyatakan bahwa populasi pulau yang menghadapi lingkungan ektrim akan menampilkan arah yang berlawanan dengan sindrom pulau. Tetapi pernyataan Raia et al. (2010) selanjutnya yang menyatakan populasi akan mempertahankan kepadatan populasi tetap rendah dan mendorong produksi telur yang banyak dengan ukuran tetasan yang kecil, menjadi tidak sesuai dengan kondisi populasi komodo di pulau kecil, yang jumlah populasinya semakin menurun. Kondisi ukuran tubuh yang lebih kecil, laju pertumbuhan yang rendah dan ukuran populasi yang mengecil disebutkan Jessop et al. (2005) sebagai bukti adanya tekanan dan keterbatasan energetik pada populasi komodo di pulau kecil.
Perbandingan Karakteristik Habitat antara Pulau Besar dan Pulau Kecil
Pulau besar dan pulau kecil di Taman Nasional Komodo memiliki perbedaan dan karakteristik habitat berupa ukuran dan jarak antar pulau, tipe ekosistem, keberadaan pemukiman dan aktivitas wisata, serta ketersediaan mangsa. Keempat faktor tersebut patut diduga memberi pengaruh pada laju pertumbuhan morfometrik biawak komodo.
Ukuran dan Jarak antar Pulau
18
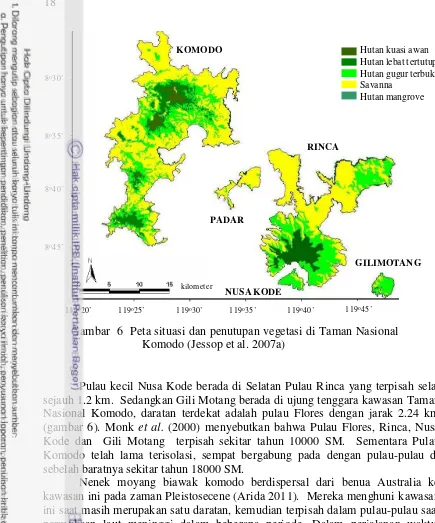
Gambar 6 Peta situasi dan penutupan vegetasi di Taman Nasional Komodo (Jessop et al. 2007a)
Pulau kecil Nusa Kode berada di Selatan Pulau Rinca yang terpisah selat sejauh 1.2 km. Sedangkan Gili Motang berada di ujung tenggara kawasan Taman Nasional Komodo, daratan terdekat adalah pulau Flores dengan jarak 2.24 km (gambar 6). Monk et al. (2000) menyebutkan bahwa Pulau Flores, Rinca, Nusa Kode dan Gili Motang terpisah sekitar tahun 10000 SM. Sementara Pulau Komodo telah lama terisolasi, sempat bergabung pada dengan pulau-pulau di sebelah baratnya sekitar tahun 18000 SM.
Nenek moyang biawak komodo berdispersal dari benua Australia ke kawasan ini pada zaman Pleistosecene (Arida 2011). Mereka menghuni kawasan ini saat masih merupakan satu daratan, kemudian terpisah dalam pulau-pulau saat permukaan laut meninggi dalam beberapa periode. Dalam perjalanan waktu, setiap populasi pada pulau-pulau tersebut mengalami isolasi reproduksi dan perubahan genetik (mutasi) dari nenek moyangnya, sehingga terjadi divergensi (perbedaan) genetik antar populasi dalam kurun waktu yang beragam. Tingkat kekerabatan antara populasi yang telah terpecah tersebut dapat diestimasi melalui waktu divergensi (Zhivotovsky 2001). Studi oleh Ciofi dan Bruford (1999b) menunjukkan bahwa waktu divergensi antara populasi biawak komodo di Pulau Gili Motang dengan Pulau Komodo adalah 3616 tahun, sedangkan dengan Pulau Rinca adalah 1000 tahun. Waktu divergensi terpendek adalah antara populasi biawak komodo di Pulau Gili Motang dengan Pulau Flores yaitu 98 tahun. Sementara waktu divergensi populasi komodo antara dua pulau besar adalah 4453 tahun. Dengan demikian jarak kekerabatan genetik yang paling jauh adalah antara
19 populasi biawak komodo di Pulau Komodo dan Pulau Rinca. Sementara populasi biawak komodo di Pulau Gili Motang dan Pulau Flores menunjukkan jarak kekerabatan yang paling dekat. Sedangkan tingkat kekerabatan genetik populasi biawak komodo pada Pulau Rinca dan Nusa Kode lebih dekat dibandingkan dengan Gili Motang.
Walaupun populasi biawak komodo terpisah antar pulau tetapi terdapat fakta bahwa biawak komodo memiliki kemampuan berenang sehingga terdapat peluang migrasi antar pulau dan terjadi aliran genetik. Tentu saja kemampuan migrasi antar pulau dibatasi oleh jarak dan kondisi perairan antar pulau, serta keterbatasan daya renang biawak komodo. Apabila ditinjau dari posisi dan jarak antara pulau diperkirakan biawak komodo di pulau komodo tidak dapat bermigrasi ke pulau lainnya. Hal tersebut sejalan dengan Ciofi et al. (1999) yang memperkirakan migrasi dari dan ke Pulau Komodo sudah tidak terjadi pada akhir periode glasial terakhir. Sementara pada akhir masa glasial antara Flores, Rinca, Nusa Kode dan Rinca masih mempertahankan aliran genetik. Hingga kemudian Gili Motang yang pertama kali terpisah. Lebih lanjut Ciofi et al. (1999) menyatakan laju migrasi biawak dari atau menuju Gili Motang adalah seekor setiap 60 tahun, merupakan nilai yang sangat rendah dan diperkirakan terjadi hanyutan genetik terstruktur pada populasi biawak komodo di Gili Motang. Dengan demikian pulau kecil (Gili Motang dan Nusa Kode) tidak mengalami isolasi dibandingkan dengan Pulau Komodo. Akan tetapi setelah Pulau Komodo maka yang kedua terisolasi adalah pulau kecil Gili Motang, kemudian Pulau Rinca dan Nusa Kode.
Populasi biawak komodo di Pulau Komodo yang memiliki tingkat kekerabatan paling jauh dengan ketiga pulau lainnya ternyata memiliki tingkat keanekaragaman genetik yang paling tinggi (Ciofi dan Bruford 1999). Sedangkan populasi biawak komodo di Pulau Gili Motang memiliki tingkat keanekaragan genetik yang paling rendah, disertai jumlah populasi yang kurang dari 50 ekor, Arida (2011) mengindikasikan adanya tingkat inbreeding yang tinggi. Yang berarti bahwa di Gili Motang kerap terjadi perkawinan antar individu yang memiliki pertalian keluarga dekat, yang terlihat dari tingginya homozigositas. Ditegaskan oleh Purwandana (2015) bahwa koefisien inbreeding populasi biawak komodo di Gili Motang (0.45 ± 0.02) adalah paling tinggi dibandingkan lokasi-lokasi lain. Sementara koefisien inbreeding pada lokasi-lokasi di pulau besar berkisar antara 0.12 sampai 0.27, dan koefisien inbreeding terkecil adalah di Nusa Kode (0.11 ± 0.02).
Tipe Ekosistem
Secara umum formasi tumbuhan pada pulau-pulau di Taman Nasional Komodo adalah tumbuhan yang bertahan pada iklim yang kering. Auffenberg (1980) menyebutkan terdapat tiga formasi tumbuhan yang dapat dijumpai di Pulau Komodo dan pulau-pulau di sekitarnya yaitu hutan gugur (musim) tropis, savana dan stepa. Selain itu terdapat juga hutan kuasi awan di daerah pegunungan dan hutan mangrove di sebagian pantai.
20
hutan lebat tertutup berfungsi baik sebagai tempat berteduh saat cuaca panas dan tumbuhan banyak meranggas. BTNK (2013a) menyebutkan bahwa ekosistem hutan gugur terbuka memiliki unsur habitat yang paling diperlukan oleh komodo, yaitu tempat berjemur, tempat berteduh dan tempat bersarang. Demikian pula dengan ketersediaan mangsa besar dan mangsa kecil paling banyak ditemui di hutan gugur terbuka. Sehingga ditemui nilai kepadatan biawak komodo yang tinggi dibandingkan ekosistem lainnya.
Apabila dibandingkan maka pulau besar memiliki tipe ekosistem yang lebih beragam dibandingkan pulau kecil (Tabel 8). Pulau Rinca dan Komodo ditutupi oleh tipe ekosistem mangrove, savana, hutan gugur terbuka, hutan gugur tertutup. Khusus di Pulau Komodo juga terdapat hutan kuasi berawan. Tipe ekosistem savana menutupi 59.4% Pulau Komodo dan 55.0% Pulau Rinca. Sementara di Pulau Gili Motang tidak terdapat ekosistem magrove, sedangkan di Nusa Kode tidak terdapat hutan lebat tertutup. Seluas 80% Pulau Gili Motang dan 84.3% Nusa Kode ditutupi oleh hutan gugur terbuka. Hasil pengukuran indeks struktur vegetasi yang dihitung dari persentase luas pulau dibandingkan luas hutan gugur terbuka dan hutan gugur tertutup, maka terlihat pulau kecil memiliki indeks di bawah 50%, sedangkan lokasi-lokasi di pulau besar seluruhnya memiliki indeks di atas 50% (Purwandana 2015).
Hasil analisis vegetasi oleh Rudiharto (2007) menunjukkan bahwa jumlah jenis tumbuhan perplot yang ditemukan di Gili Motang (salah satu pulau kecil) signifikan lebih sedikit dibandingkan lokasi lainnya di pulau besar. Tetapi jumlah jenis perplot yang ditemukan di pulau kecil lainnya yaitu Nusakode tidak signifikan berbeda dengan lokasi lain di pulau besar, karena kedekatannya dengan pulau Rinca sehingga kelembabannya relatif lebih terpelihara dibanding Gili Motang. Demikian juga komposisi INP di Gili Motang terlihat didominasi oleh satu jenis yaitu walikukun (Schoutenia ovata). Sebagaimana disebutkan oleh Whittaker (1998) bahwa dalam kondisi fisik yang ekstrim, sedikitnya jenis akan dikompensasi dengan banyaknya jumlah individu perjenis.
Secara umum kondisi vegetasi di pulau Gili Motang terlihat ekstrim ditinjau dari dominasi habitat berupa hutan kering (Jessop et al. 2007a), ketiadaan sumber air (Rudiharto 2007) dan berulang kali mengalami kebakaran (BTNK 2011). Kondisi ekstrim di pulau kecil patut diduga sebagai penyebab rendahnya laju pertumbuhan biawak komodo di pulau kecil.
Tabel 8 Proporsi tipe ekosistem pada masing-masing pulau di Taman Nasional Komodo*
Tipe ekosistem
21 Keberadaan Pemukiman dan Aktivitas Wisata
Pulau-pulau kecil tidak dihuni oleh manusia dan tidak terdapat aktivitas wisata. Sementara di pulau besar terdapat pemukiman manusia, yaitu di Pulau Komodo terdapat Kampung Komodo dengan jumlah penduduk 1406 jiwa dan di Pulau Rinca terdapat Kampung Rinca dengan jumlah penduduk 887 jiwa dan Kampung Kerora dengan jumlah penduduk 250 jiwa. Di kedua pulau besar juga terdapat areal wisata alam, yaitu Loh Liang yang dikunjungi 19935 wisatawan dan Loh Buaya yang dikunjungi 30047 wisatawan pada tahun 2012 (BTNK 2013b). Keberadaan aktivitas manusia di pulau besar menimbulkan bertambahnya preferensi pakan komodo. Di areal wisata, pada kondisi kegiatan wisata tertentu dilakukan pengumpanan dengan kambing untuk menarik kedatangan biawak komodo. Sementara di pemukiman komodo kerap makan sampah dapur, ikan dan hewan ternak. Ardiantiono (2014) menyebutkan, hasil wawancara dengan penduduk kampung Komodo dan Rinca, terhitung telah terjadi 405 kali serangan komodo terhadap hewan ternak kambing dan ayam dalam waktu tiga tahun, tetapi serangan tersebut tidak selalu berakhir dengan pemangsaan karena kerap ternak tersebut dapat diselamatkan.
Ketersediaan Mangsa
Setiap kelas umur biwak komodo memiliki ketertarikan jenis mangsa yang berbeda. Mangsa utama komodo dewasa dan remaja adalah ungulata besar seperti rusa dan babi hutan (Aufenberg 1981), sedangkan komodo anak memangsa tikus, tokek dan serangga. Apabila ditinjau dari ketersediaan mangsa, terlihat bahwa kelimpahan dan keragaman mangsa bagi komodo di pulau kecil lebih sedikit dibandingkan dengan pulau besar (Tabel 9).
Kepadatan rusa di pulau besar Komodo (88.6 ekor/km2) dan Rinca (41.6 ekor/km2),lebih tinggi dibandingkan kepadatan rusadi pulau kecil Nusa Kode (2.5 ekor/km2) dan Gili Motang (6.5 ekor/km2) (Ariefandy et al. 2013). Di Pulau Rinca dan Pulau Komodo juga terdapat mangsa biawak komodo yang tidak terdapat di pulau kecil, yaitu kerbau dan babi hutan.
Tabel 9 Indeks kelimpahan mangsa komodo di pulau besar dan pulau kecil Jenis Mangsa
Rata-rata indeks kelimpahan Pulau besar Pulau kecil
Komodo Rinca Gili Motang Nusa Kode Mangsa besar
Rusa 27.364 21.211 7.186 7.178
Babi hutan 0.675 0.525 0.000 0.000
Kerbau 0.115 0.430 0.000 0.000
Mangsa kecil
Tokek - 0.790 2.500 1.000
Tikus - 0.030 0.002 0.161
22
Sementara itu untuk ketersediaan mangsa kecil, Rudiharto (2007) menyebutkan bahwa kepadatan populasi mangsa kecil yaitu tikus dan tokek signifikan lebih tinggi di pulau kecil dibandingkan pulau besar. Dari hasil pengamatan didapatkan bahwa di Gili Motang (salah satu pulau kecil) terdapat 2.05 ekor tokek pernight-transect, signifikan lebih tinggi dibanding tiga tempat lain di pulau besar (Loh Buaya 1.07 ekor tokek pernight-transect, Loh Dasami 1.05 ekor tokek pernight-transect, Loh Baru 0.27 ekor tokek pernight-transect
dan Nusa kode 1.00). Sedangkan rata-rata jumlah tangkapan tikus di Nusakode (salah satu pulau kecil) adalah 0.161 ekor pernight-trap signifikan lebih tinggi dibanding tempat lain di pulau besar (Loh Buaya 0.039 ekor pernight-trap, Loh Baru 0.045 ekor pernight-trap, Loh Dasami 0.018 ekor pernight-trap dan Gili Motang 0.002 ekor pernight-trap).
Dengan langkanya mangsa besar di pulau kecil, maka biawak komodo dewasa diduga mencari alternatif makanan lain misalnya mangsa kecil. Sejalan dengan Rudiharto (2007) yang menyatakan mangsa kecil berkontribusi dalam mensubtitusi pakan komodo di pulau kecil. Walaupun demikian keterbatasan mangsa besar juga dapat menyebabkan terjadinya kematian pada biawak komodo dewasa, karena kemampuan untuk mendapatkan mangsa kecil juga tidak dapat diandalkan berhubung perubahan ontogeni ukuran tubuhnya (Jessop et al. 2007a). Sehingga mendorong tingkat kematian selektif yang mengarah pada penurunan populasi di pulau kecil.
Lebih sedikitnya mangsa di pulau kecil diduga merupakan penyebab lebih rendahnya laju pertumbuhan biawak komodo di pulau kecil dibandingkan pulau besar. Sejalan dengan pernyataan Jessop et al. (2005) dan Whittaker (1998) yang menyebutkan ekosistem pulau kecil kadang tidak menguntungkan beberapa spesies, misalnya dengan langkanya pakan sehingga yang terjadi adalah spesies akan berkurang pertumbuhannya serta menyusut ukurannya. Hal ini dipertegas dengan oleh studi Jessop et al. (2006) yang menyebutkan bahwa indeks kepadatan rusa berkorelasi kuat dengan variasi ukuran dan berat tubuh biawak komodo antar pulau.
23 pulau besar yaitu Pulau Komodo adalah 0.07 dan Pulau Rinca adalah 0.17. Sementara untuk pulau kecil Gili Motang adalah 0.71. Sebagaimana disebutkan oleh Ciofi dan Bruford (1999), ukuran populasi efektif secara tidak langsung untuk populasi di Pulau Komodo adalah 130.1 ekor, Pulau Rinca 189.9 ekor dan Gili Motang adalah 85.8 ekor. Dengan demikian dapat disimpulkan bahwa ditinjau dari ukuran populasi efektif secara tidak langsung, populasi biawak komodo di pulau kecil memiliki kemampuan bereproduksi lebih rendah dibandingkan populasi komodo di pulau besar.
Implikasi Konservasi
Laju pertumbuhan morfometrik populasi komodo di pulau kecil pada kelas umur anak maupun remaja yang lebih rendah dibandingkan populasi komodo anak dan remaja di pulau besar, menunjukkan adanya mekanisme ekologis yang berbeda antara pulau kecil dan pulau besar, yang dapat mengarah pada keterancaman populasi pada pulau kecil. Pulau kecil yang prioritas memperoleh perhatian adalah Gili Motang karena ketersediaan mangsa paling rendah, tutupan vegetasi kurang ideal untuk komodo, rawan terjadi kebakaran, paling terisolasi dan keanekaragaman genetiknya paling rendah. Sementara itu pulau besar yang patut menjadi pusat konservasi adalah Pulau Komodo, dengan tingkat keanekaragaman genetik yang paling tinggi serta kondisi ekologi yang kondusif, Pulau Komodo berperan sebagai mainland bagi pulau-pulau habitat populasi komodo dimanapun.
Kondisi fisik dan biologi yang beragam pada pulau-pulau habitat biawak komodo dan keadaan populasinya, mempengaruhi keberlangsungan hidup biawak komodo. Purwandana (2015) menganalisis ragam tingkat survival populasi komodo di Taman Nasional Komodo, dari kovariate indeks struktur vegetasi, indeks kualitas lingkungan, biomassa mangsa, koefisien inbreeding dan kepadatan populasi. Hasilnya menunjukkan bahwa elemen yang paling berpengaruh pada tingkat survival populasi komodo adalah ketersediaan mangsa atau yang disebutkan oleh Alikodra (1990) sebagai faktor pembatas. Faktor tersebut berpengaruh pada kelimpahan, pertumbuhan dan penyebaran. Dengan demikian implikasi konservasi bagi manajemen adalah memperhatikan ketersediaan mangsa pada setiap pulau populasi biawak komodo di Taman Nasional Komodo.
4
SIMPULAN DAN SARAN
Simpulan
24
2. SVL merupakan peubah morfometrik yang paling tinggi korelasinya dengan pertambahan umur. Adapun model laju pertumbuhan dari variable morfometrik SVL pada populasi komodo di pulau kecil adalah Y = 1.48 + 0.222X, sedangkan untuk pulau besar yaitu Y = 1.331 + 6.182X.
3. Perbedaan karakteristik habitat komodo di pulau besar dibandingkan pulau kecil adalah ketersediaan mangsa, keragaman tipe ekosistem, serta proporsi penutupan vegetasi antara savana, hutan gugur terbuka dan hutan gugur tertutup. Dengan demikian pulau besar dapat memberikan dukungan yang lebih baik bagi kehidupan komodo.
Saran
1. Penelitian menunjukkan bahwa spesies langka dengan ukuran populasi kecil pada pulau yang terisolasi perlu mendapatkan perhatian khusus terutama ketersediaan pakan.
2. Penelitian dapat ditindaklanjuti dengan perbandingan bentuk tubuh antara populasi komodo di pulau besar dengan pulau kecil, untuk mempelajari perubahan ontogeni tubuh komodo sebagai respon perubahan ekologi.
DAFTAR PUSTAKA
Alder GH, Levin R. 1994. The island syndrome in rodent populations. Quarter Review Biology (69): 473-489.
Alikodra HS. 1990. Pengelolaan Satwaliar Jilid 1. Bogor (ID): Departemen Pendidikan dan Kebudayaan Ditjen Pendidikan Tinggi Pusat Antar Universitas Institut Pertanian Bogor.
Ardiantiono. 2014. Analisis Spasial Konflik Manusia-Biawak Komodo di Desa Komodo, Taman Nasional Komodo [skripsi]. Depok (ID): Universitas Indonesia.
Arida AA. 2011. Phylogeography of the Komodo monitor Varanus komodoensis (Reptilia: Varanidae) inferred from mitochondrial DNA Control Region I and the implications for in situ management plans [Dissertation]. Bonn (DE): Rheinischen Friedrich-Wilhelms-Universität Bonn.
Ariefiandy A, Purwandana D, Coulson G, Forsyth DM, Jessop TS. 2013. Monitoring the primary prey of the Komodo dragon: distance sampling or faecal counts? Wildl. Biol. 19: 1-12.
Auffenberg W. 1981. The Behavioral Ecology of the Komodo Monitor. Gainesville (US): University Presses of Florida.
[BTNK] Balai Taman Nasional Komodo. 2011. Ekologi Pulau Gili Motang.
Labuan Bajo (ID): Balai Taman Nasional Komodo.
[BTNK] Balai Taman Nasional Komodo. 2013a. Laporan Inventarisasi Komodo.
Labuan Bajo (ID): Balai Taman Nasional Komodo.
25 Bjorndal KA, Bolten AB, Chaloupka MY. 2000. Green turtle somatic growth
model: evidence for density dependence. Ecol Appl 10: 269–282.
Blondel J. 2000. Evolution and ecology of bird on island: trend and prospect. Vie etMilieu (50): 250-220.
Chourchamp F, Clutton-Brock T, Grenfell B. 1999. Inverse density dependence and the Allee effect. Trends Ecol Evol 14: 405-410.
Ciofi C, Beaumont MA, Swingland IR, Bruford MW. 1999. Genetic divergence and units for conservation in komodo dragon Varanus komodoensis. Pro R Lond. B(266):2269-2274.
Ciofi C, Bruford MW. 1999. Genetic structure and gene flow among komodo dragon populations inferred by microsatellite loci analysis. Molecular Ecology 8: S17-S30.
Cleeg SM, Owens PF. 2002. The island rule in bird: medium body size and its ecological explanation. Proc. R. Soc. Land B(269): 1359-1365.
Jessop TS, Imansyah MJ, Purwandana D, Ariefiandi A, Rudiharto H, Seno A, Opat DA, Noviandi T, Payung I, Ciofi C. 2007a. Ekologi Populasi, Reproduksi dan Spasial Biawak Komodo (Varanus komodoensis) di Taman Nasional Komodo, Indonesia. Labuan Bajo (ID): Balai Taman Nasional Komodo, Center for Conservation and Research of Endangered Species, Zoological Society of San Diego, The Nature Conservancy.
Jessop TS, Madsen T, Ciofi C, Imansyah MJ, Purwandana, Rudiharto H, Ariefiandy A, Phillips JA. 2007b. Island differences in population size structure and catch per unit effort and their conservation implications for Komodo dragons. Biological Conservation. 135: 247 –255.
Jessop TS, Imansyah MJ, Purwandana D, Ariefiandi A, Rudiharto H. 2007c.
Panduan Teknis Pemantauan Ekologi dan Hidupan Liar di Taman Nasional Komodo Indonesia. Labuan Bajo (ID): Center for Conservation and Research of Endangered Species, Zoological Society of San Diego, Balai Taman Nasional Komodo, The Nature Conservancy.
Jessop TS, Madsen T, Sumner J, Rudiharto H, Phillips JA, and Ciofi C, 2006. Maximum body size among insular komodo dragon populations covaries with large prey density. Oikos. 112(2): 422 – 429.
Jessop TS, MadsenT, Purwandana D, Imansyah MJ, Rudiharto H, Ciofi C. 2005.
Evidence for Energetic Constraints Affecting A Small Island Komodo Dragon Population. Labuan Bajo (ID): The Zoological Society of San Diego, Komodo National Park, and The Nature Conservancy.
Jessop TS, Sumner J, Rudiharto H, Purwandana D, Imansyah MJ, Phillips JA. 2003. Distribution, use and selection of nest type by komodo dragons.
Biological Conservation (117): 463 – 470.
Kolbe JJ, Colbert PR, Smith BE. 2008. Niche Niche relationship and interspesific interactions in antiguan lizard communities. Copeia (2): 261 – 272.
Krebs CJ. 1999. Ecological Methodology, Second Edition. California (US): Addison-Wesley Educational Publisher.
Laver RJ, Purwandana D, Ariefandy A, Imansyah J, Forsyth D, Ciofi C, Jessop TS. 2012. Life-History and spatial determinants of somatic growth dynamics in komodo dragon populations. Plos One 7( 9)(e45398):1-10. Monk KA, de Fretes Y, Reksodiharjo-Lilley G, Kartikasari S.N.(editor). 2000.
26
Purwandana D, Ariefiandy A, Imansyah M J, Ciofi C, Forsyth DM, Gormley AM, Rudiharto H, Seno A, Fordham DA, Gillespie G, Jessop TS. 2015. Evaluating environmental, demographic and genetic effects on population-level survival in an island endemic. Ecography (38): 001–011.
Purwandana D, Ariefiandy A, Imansyah M J, Rudiharto H, Seno A, Ciofi C, Fordham DA, Jessop TS. 2014. Demographic status of Komodo dragons populations in Komodo National Park. Biological Conservation (171): 29– 35.
MacArthur RH, Wilson EO. 1967. The Theory of Island Biogeography. Princeton (US): Princeton University Press.
Raia P, Guarino FM, Turano M, Polese G, Rippa D, Carotenuto F, Monti DM, Cardi M, Fulgione. 2010. The blue lizard spandrel and island syndrom.
BMC Evolutionary 10:289.
Rudiharto H. 2007. Hubungan antara karakteristik habitat dengan densitas Komodo [tesis]. Yogyakarta (ID):Universitas Gadjah Mada.
Sastrawan P, Ciofi C. 2002. Population, Distribution and Home Range. In Murphy J B, Ciofi C, de La Panose C, Walsh T (editors). Komodo Dragons: Biology and Conservation. Washington (US) and London (UK): Smithsonian Institution Press.
Schall JJ. 1983. Small clutch size in a tropical whiptail lizard (Cnemidophorus arubensis). Journal of Herpetology 17 (4):406-408.
Walpole RE. 1995. Pengantar statistika. Jakarta (ID): Gramedia Pustaka Utama. Whittaker RJ. 1998. Island Biogeography: Ecology, Evolution and Conservation.
New York (US): Oxford University Press.
27 Lampiran 1 Laju pertumbuhan jenis morfometrik komodo kelas umur anak di lokasi- lokasi
pulau besar dan pulau kecil pada setiap tingkat pertumbuhan
36
Data Ind.
Thn
ke- Pulau Lokasi
Laju Pertumbuhan (%) Panjang
Kepala
Lebar
kepala SVL
Ventral TBL
Panjang ekor
Lingkar ekor
Berat badan
375. 3 Kecil Gili Motang 7.500 8.772 6.601 5.015 1.026 16.590 48.413
376. 3 Kecil Gili Motang 9.052 9.259 7.500 1.502 - - 22.069
377. 3 Kecil Gili Motang 11.881 6.863 2.824 1.577 - 2.000 9.149
378. 3 Kecil Nusa Kode 17.954 19.527 17.596 16.133 13.959 21.858 86.992
379. 3 Kecil Gili Motang 1.917 - - 2.099 0.923 - -
380. 3 Kecil Gili Motang 7.500 8.772 6.601 5.015 1.026 16.590 48.413
381. 3 Kecil Gili Motang 9.052 9.259 7.500 1.502 - - 22.069
382. 3 Kecil Gili Motang 11.881 6.863 2.824 1.577 - 2.000 9.149
383. 3 Kecil Nusa Kode 17.954 19.527 17.596 16.133 13.959 21.858 86.992
384. 4 Besar Loh Lawi 19.435 - 21.364 15.323 10.600 41.304 149.242
385. 4 Besar Loh Lawi 24.449 30.414 35.043 24.381 17.844 37.255 144.131 386. 4 Besar Loh Lawi 20.143 20.638 21.463 17.068 12.614 28.333 84.118 387. 4 Besar Loh Lawi 33.379 30.028 39.310 33.364 28.308 42.742 95.946 388. 4 Besar Loh Lawi 37.551 43.573 39.167 32.220 28.622 56.250 317.492 389. 4 Besar Loh Lawi 41.871 57.647 39.082 33.831 29.689 67.568 287.156 390. 4 Besar Loh Sebita 17.308 7.488 15.290 10.366 8.511 25.980 48.387
391. 4 Besar Loh Buaya 24.865 - 28.258 - 17.564 13.333 24.194
37 Lampiran 2 Laju pertumbuhan jenis morfometrik komodo kelas umur remaja di lokasi-lokasi
pulau besar dan pulau kecil pada setiap tingkat pertumbuhan