ALLOMETRY ANALYSIS OF FIDDLER CRABS Uca forcipata AND U. dussumieri
(Crustacea: Decapoda) IN THE WEST OF TANJUNG JABUNG JAMBI
TIA WULANDARI
GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY BOGOR
STATEMENT ABOUT THESIS
I hereby declare that thesis entitled Allometry Analisys of Fiddler Crabs Uca forcipata and U. dussumieri (Crustacea: Decapoda) in the West of Tanjung Jabung Jambi is original result of my own research supervised under advisory committee and has never been submitted in any form at any institution before. All information from other authors cited here are mentioned in the text and listed in the reference at the end part of the thesis.
Bogor, May 2015
RINGKASAN
TIA WULANDARI. Analisis Allometri Kepiting Biola Uca forcipata dan U. dussumieri (Crustacea: Decapoda) di Tanjung Jabung Barat Jambi. Dibimbing oleh BAMBANG SURYOBROTO dan BERRY JULIANDI.
Kepiting biola jantan dewasa memiliki seksual dimorfisme yaitu salah satu capitnya tumbuh lebih cepat dari capit lainnya. Kondisi ini menunjukkan bentuk tubuh asimetri. Capit besar dapat berada pada bagian kanan (dekstral) atau bagian kiri (sinistral) tubuh. Capit besar dan capit kecil pada Uca memiliki fungsi yang berbeda. Capit besar berfungsi sebagai armamen, yaitu untuk bertarung dengan individu jantan lainnya dalam memperebutkan teritori dan individu betina; capit besar juga berfungsi sebagai ornamen, yaitu untuk menarik perhatian individu betina. Capit kecil baik pada individu jantan maupun individu betina berfungsi sebagai alat makan.
Studi mengenai asimetri dilakukan di Tanjung Jabung Barat Jambi, Indonesia dari Agustus 2014 sampai Februari 2015. Jumlah sampel (individu) yang digunakan dalam penelitian ini adalah 153. Terdiri dari dua spesies, yaitu 89 individu Uca forcipata dan 64 individu U. dussumieri.
SUMMARY
TIA WULANDARI. Allometry Analysis of Fiddler Crabs Uca forcipata and U. dussumieri (Crustacea: Decapoda) in the West of Tanjung Jabung Jambi. Supervised by BAMBANG SURYOBROTO and BERRY JULIANDI.
Adult male of fiddler crabs have sexual dimorfism that one of cheliped grow faster than the other. This condition leads to asymmetrical body pattern. Enlarged cheliped can be on right (dextral) or left side (sinistral) of the body. Major and minor cheliped in Uca have different function. Major cheliped use as armament, for fighting with another male for territory and female; and as ornament for female attractor. Minor cheliped in male and female have same function, that as a feeding tools.
A study of asymmetry was conducted in the West of Tanjung Jabung Jambi, Indonesia from early August 2014 to the end of February 2015. The number of samples (individuals) used in this research was 153. That consisted of two species, in the following quantities 89 individu of Uca forcipata and 64 individu of U. dussumieri.
The frequency of handedness (position of enlarged cheliped) for either species at each sampling station was equal for right-handed and left-handed (p-value 0.05; chi-square test). Allometry analysis was done to study the covariation of chelipeds, relative to different body sizes using bivariate analysis. Carapace width (CRW) was used as a proxy for body size because it varies in parallel with the hypothesized positive allometry of the major cheliped. Carapace length (CRL) showed a negative allometrical growth pattern relative to CRW. The major cheliped showed positive allometry, and the minor cheliped showed isometry, in both species. This might have caused the growth imbalance between chelipeds that leads to the asymmetry of body shape.
© Copy Right by IPB, 2015
All rights reserved
It is prohibited to cite all or a part of this thesis without referring to and mentioning the source. Citation is permitted for the purpose of education, research, scientific paper, report, or critism writing only; and it does not defame the name and honour of Bogor Agricultural University.
A Graduate Thesis
in partial fulfilment of Master Science degree in Animal Biosciences Faculty of Mathematics and Natural Science
ALLOMETRY ANALYSIS OF FIDDLER CRABS Uca forcipata AND U. dussumieri
(Crustacea: Decapoda) IN THE WEST OF TANJUNG JABUNG JAMBI
GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY BOGOR
2015
ENDORSEMENT PAGE
Title : Allometry Analysis of Fiddler Crabs Uca forcipata and U. dussumieri (Crustacea: Decapoda) in the West of Tanjung
Jabung Jambi Name : Tia Wulandari Student ID : G352130221
Approved by, Supervisory Committee
Dr Bambang Suryobroto Chair
Dr Berry Juliandi Member
Agreed by,
Chair of Major of Animal Biosciences
Dr Ir R. R. Dyah Perwitasari
Dean of Graduate School,
Dr Ir Dahrul Syah, MSc.Agr
PREFACE
The title of this thesis is Allometry Analysis of Uca forcipata and U. dussumieri (Genus:Uca) in the West of Tanjung Jabung Jambi. The samples of these research took from the west of Tanjung Jabung Jambi. I want to send my gratitude to my supervisor Dr Bambang Suryobroto and Dr Berry Juliandi for all guidance and endcouragement as well as invaluable academic advices for the whole period of my study and research, and to my examiner Dr. Yusli Wardiatno, M.Sc. for the generous support and great discussion. Thank to Mr. Taharudin, Mr. Mulyo and Rizki for their help throughout in the field; Mrs. Tini, Mrs. Dewi Citra Murniati for their help in the laboratory. Thank to all people in Zoo Corner, BSH 2013, and special thank to my parents and my family, for their supports for my study. Therefore, above all, I thank God Allah SWT for His almighty.
Bogor, May 2015
CONTENTS
LIST OF TABLES vi
LIST OF FIGURES vi
INTRODUCTION 1
MATERIALS AND METHOD 1
Sample 1
Data Analysis 2
RESULT AND DISCUSSION 4
Results 4
Discussion 7
CONCLUSION 9
REFFERENCES 9
APPENDIXES 11
LIST OF TABLES
1 Frequency of handedness 4
2 Growth pattern comparison of CRL and CRW 5
3 Elevation regression line value of U. forcipata and U. dussumieri 6 4 Allometry analysis of U. forcipata and U. dussumieri 6 5 Allometry analysis of handedness U. forcipata and U. dussumieri 6
LIST OF FIGURES
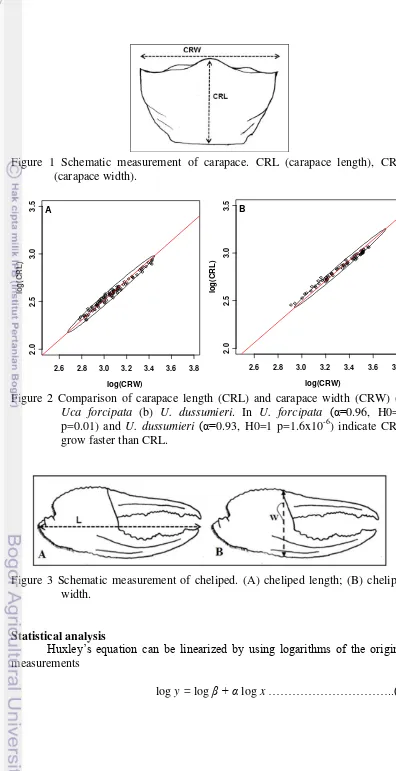
1 Schematic measurement of carapace 3
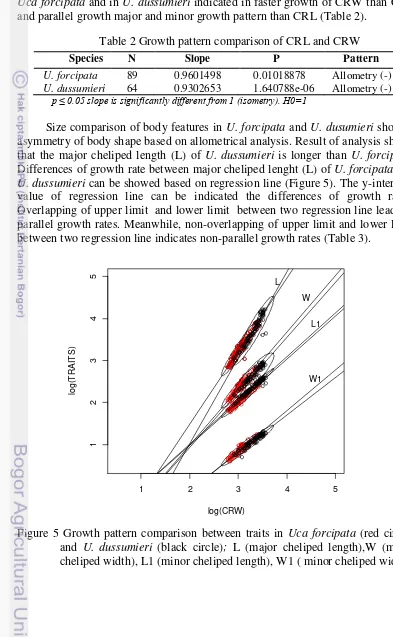
2 Comparison of carapace length (CRL) and carapace width (CRW)
(a) Uca forcipata (b) U. dussumieri 3
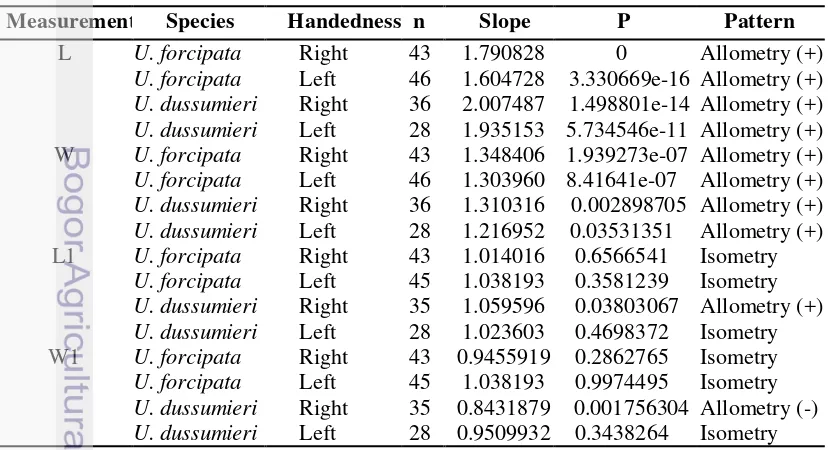
3 Schematic measurement of cheliped 3
4 Antisymmetry in U. forcipata and U. dussumieri 4 5 Growth pattern comparison between traits in Genus Uca 5 6 Growth pattern comparison between traits in (A) right-handed and
(B) left-handed of U. forcipata 7
7 Growth pattern comparison between traits in (A) right-handed and
(B) left-handed U. dussumieri 7
LIST OF APPENDIX
1 Location of research sampling 12
INTRODUCTION
Fiddler crabs (Uca spp.) are intertidal crabs that show sexual dimorphism during the adult stage. The sexual dimorphism manifests as asymmetry in male chelipeds, while the female remains symmetric (Rosenberg 2001). Adult males
have one enlarged cheliped, with the other remaining ‘normal’. The major cheliped can be twice the size of the carapace. This cheliped is thought to be used for armament and ornament, while the minor cheliped is used for feeding in the male as are both chelipeds in the female (Rosenberg 2002, Murniati 2009).
In fiddler crabs, the enlarged cheliped can be on the right (dextral) or left (sinistral) side of the body. This random asymmetry within a population is known as antisymmetry (Valen 1962). In antisymmetry, development of asymmetrical traits is almost never inherited and is triggered by the external environment (Vanberg and Costlow 1966; Yamaguchi 1977; Palmer 2004; Palmer 2012). Selection pressure in a population could affect the dominance of right or left enlarged chelipeds (Palmer 1996; Croll and McClintock 2002; Rosenberg 2002). Many traits that are associated with combat or aids to courtship, evolved by sexual selection; for example, in fiddler crabs the enlarged cheliped is used as weapon in male combat, and for courtship display to attract females (Boundriansky and Day 2003).
Asymmetry of bilateral features develops through an allometrical growth mechanism. Huxley (1924) coined the elementary law of relative growth, maintaining the same shape over a range of sizes, it is said to be in an isometrical growth pattern; when the shape changes following changes in size, it is allometric (Claude 2008).
Although asymmetry has been extensively observed in genus Uca, quantification and the cause of antisymmetry in this genus - especially Uca forcipata (Adams and White, 1848)and U. dussumieri (Milne-Edwards, 1852) – has remained unknown. This present study reports that the frequency of left-handed or right-left-handed in cheliped of U. forcipata and U. dussumieri is equal and the handedness occurs in an antisymmetrical way. An asymmetrical body shape is caused by the faster growth of one cheliped relative the other.
MATERIALS AND METHOD
Sample
2
were taken from an area in the West of Tanjung Jabung, Jambi, Indonesia. Crabs of this size have already developed asymmetrical chelipeds. They were collected from two stations with different types of habitat 0o48’44.2”S-103o29’2.8”E: sandy clay substrate and 0o49’3.1”S-103o28’48.7”E: clay substrate; (appendix 1). Specimens were collected from 08.00-15.00 WIB (low tide and sunshine). The sample was identified at the Museum Zoologicum Bogoriense (MZB) in Cibinong. Sample identification was based on criteria established by Crane (1975). Identification was taken to species level. The number of samples used in this research was 153 individuals: comprising two species, in the following quantities: 89 individu of Uca forcipata and 64 individu of U. dussumieri.
Data Analysis Frequency of either handedness
The frequency of ‘right-handed’ and ‘left-handed’ individuals at each station was compared using the chi-square test (Agresti 2007).
Allometry analysis
Following Huxley’s morphological law (Huxley 1924), allometry analysis was carried out, in order to study the covariation of size of major and minor chelipeds (y) relative to different body sizes (x). When α=1, y grows in an isometric way relative to x; that is, increases of one unit measurement of x will be followed by increases of the same amount of y. When α≠1, the rate of growth of y is longer (allometry positive) or smaller (allometry negative) than that of x.
In this study, the body size was estimated using carapace size. Carapace width (CRW, the distance between two antero-lateral angles in the carapace) was the most representative of overall size of the crabs (Martins and Masunari 2013). In a preliminary allometrical comparison (see statistical analysis below) of the respective growths of CRW and carapace length (CRL, the distance between front and intestinal of carapace) (Figure 1), it was shown that CRW grew faster than CRL (Figure 2). Therefore CRW was used as an estimate of body size, as it varies in parallel with the hypothesized positive allometry of major cheliped.
4
This equation can be used to describe allometrical growth patterns in respect of chelipeds and carapace width. The constancy of α is an important parameter in the assessment of the static allometry of adult crabs. The regression model II was chosen to find the regression line (Sokal and Rohlf 1969), because this model takes into consideration the inherent variability of both the carapace and the chelipeds. This is in contrast to model I, where one of the variables should be measured without error. The estimate and confidence intervals for the slope of the model II regression line were measured using the standardised major axis (SMA) method (Sokal and Rohlf 1969). The Standardised Major Axis Tests and Routines (SMATR) software provides tools for the estimation, and inferences about regression lines relative to hypothetical isometric lines (Warton et al. 2012).
RESULTS AND DISCUSSION
Results
Frequency of either handedness
In adult male genus Uca,one of the cheliped grew faster than the other. The enlarged cheliped can be found on right or left side of the body. This phenomena is called antisymmetry (Figure 4). This study shows that the frequency of
No Species Station Right-handed Left-handed p-value
1 U. forcipata I 36 38 0.8162
2 U. forcipata II 6 9 0.4386
3 U. dussumieri II 36 28 0.3173
Total 78 75
5 Allometry analysis
In this study, the asymmetrical growth pattern of major and minor chelipeds was compared by reference to carapace size. Comparison of CRL and CRW in Uca forcipata and in U. dussumieri indicated in faster growth of CRW than CRL and parallel growth major and minor growth pattern than CRL (Table 2).
Table 2 Growth pattern comparison of CRL and CRW
Species N Slope P Pattern
U. forcipata 89 0.9601498 0.01018878 Allometry (-)
U. dussumieri 64 0.9302653 1.640788e-06 Allometry (-)
p ≤ 0.05 slope is significantly different from 1 (isometry). H0=1
Size comparison of body features in U. forcipata and U. dusumieri showed asymmetry of body shape based on allometrical analysis. Result of analysis shows that the major cheliped length (L) of U. dussumieri is longer than U. forcipata. Differences of growth rate between major cheliped lenght (L) of U. forcipata and U. dussumieri can be showed based on regression line (Figure 5).The y-intercept value of regression line can be indicated the differences of growth rates. Overlapping of upper limit and lower limit between two regression line leads to parallel growth rates. Meanwhile, non-overlapping of upper limit and lower limit between two regression line indicates non-parallel growth rates (Table 3).
Figure 5 Growth pattern comparison between traits in Uca forcipata (red circle) and U. dussumieri (black circle); L (major cheliped length),W (major cheliped width), L1 (minor cheliped length), W1 ( minor cheliped width).
6
Table 3 Elevation regression line value of U. forcipata and U. dussumieri
Traits U. forcipata U. dussumieri
Elevation Lower limit Upper limit Elevation Lower limit Upper limit L -1.897593 -2.203430 -1.591755 -2.838938 -3.412637 -2.265238 species, irrespective of handedness (Table 4); and second, data separated for right-handed and left-right-handed (Table 5). When the slope is significantly different from 1,
an allometric growth pattern (p ≤ 0.05) was indicated. The major cheliped shows
positive allometry, and the minor cheliped shows an isometric growth pattern, with respect to CRW, for both of groups of data (Figures 6 and 7).
Table 4 Allometry analysis of U. forcipata and U. dussumieri Measurement Species N Slope Intercept P Pattern
L U. forcipata 89 1.705365 -1.897593 0 Allometry (+) cheliped width); p ≤ 0.05 slope is significantly different from 1 (isometry); H0=1.
Table 5 Allometry analysis of handedness U. forcipata and U. dussumieri Measurement Species Handedness n Slope P Pattern
L U. forcipata Right 43 1.790828 0 Allometry (+)
U. forcipata Left 46 1.604728 3.330669e-16 Allometry (+)
U. dussumieri Right 36 2.007487 1.498801e-14 Allometry (+)
U. dussumieri Left 28 1.935153 5.734546e-11 Allometry (+)
W U. forcipata Right 43 1.348406 1.939273e-07 Allometry (+)
U. forcipata Left 46 1.303960 8.41641e-07 Allometry (+)
U. dussumieri Right 36 1.310316 0.002898705 Allometry (+)
U. dussumieri Left 28 1.216952 0.03531351 Allometry (+)
L1 U. forcipata Right 43 1.014016 0.6566541 Isometry
U. forcipata Left 45 1.038193 0.3581239 Isometry
U. dussumieri Right 35 1.059596 0.03803067 Allometry (+)
U. dussumieri Left 28 1.023603 0.4698372 Isometry
W1 U. forcipata Right 43 0.9455919 0.2862765 Isometry
U. forcipata Left 45 1.038193 0.9974495 Isometry
U. dussumieri Right 35 0.8431879 0.001756304 Allometry (-)
U. dussumieri Left 28 0.9509932 0.3438264 Isometry
7
Figure 6 Growth pattern comparison between traits in (A) right-handed and (B) left-handed of U. forcipata. L (major cheliped length), W (major
8
equal frequency of either handedness (p≤0.05). Jones and George (1982) also found that U. dussumieri had equal proportions of right-handed and left-handed males. In contrast, another study in southeast Sulawesi found 14 left-handed and two right-handed males of U. dussumieri (Weis and Weis 2004). In Thallasuca, another subgenera of Uca showed a predominance of right-handed males (Jones and George 1982).
Development of antisymmetrical traits is almost never inherited. Antisymmetrical development is triggered by the external environment (Vanberg and Costlow 1966; Yamaguchi 1977; Palmer 2004; Palmer 2012). The major cheliped in genus Uca develops in parallel with sexual maturation. It develops for the first time in the seventh juvenile stage of juvenile, although there is no obvious sign of asymmetry at that stage (Pralon and Negreiros-Fransozo 2007; Vieira 2010). Sexual maturation is not the only possible factor however; loss of one cheliped before the immature stage, also determined handedness. It has not been established whether handedness in Uca is caused by genetic factors, or an imbalance differentiation during growth because of the loss of one claw (Vanberg and costlow 1966; Yamaguchi 1977).
The major cheliped is used for combat and the attraction of mates. When this cheliped is lost, it will grow back on the same side. The respective handedness in the male Uca could affect fight activity. In one population, the choice of fighting a rival with the same or different handedness can affect fighting tactics. Right-handed males are significantly more likely to fight than left-handed males. When left-handed males do fight, they are less likely to win (Backwell et al. 2007).
Allometry analysis
Standard growth of body dimensions between individuals in the same species or different species has biological consequences (Pralon and Negreiros-Fransozo 2007). In this study, asymmetrical growth patterns of chelipeds were compared with carapace size. CRW was considered the best predictor, relative to other body dimensions, and considered an appropriate proxy for body size since it is most representative of the overall size of crabs (Araujo et al. 2012; Martins and Masunari 2013).
Major cheliped length in U. dussumieri is longer than in U. forcipata. The propudus and the dactilus in U. dusssumieri are both slimmer than in U. forcipata. These factors caused a larger total length of cheliped in U. dussumieri than the thicker one found in U. forcipata. The relative dimensions of cheliped size can be caused by genetic factors, but in some cases can be influenced by environmental factors (Murniati 2010). Based on the major cheliped length (L) of U. dussumieri longer than U. forcipata, further analysis separated these two species (interspecies analysis).
9 asymmetrical pattern in U. forcipata and U. dussumieri. Isometry shows a linear regression of 45o and indicates similar proportions of size growth between predictor and response. Meanwhile, allometry leads to a linear regression with an
angel ≠ 45o
; more than 45o for positive allometry, and less than 45o for negative allometry.
CONCLUSION
The frequencies of handedness in U. forcipata and U. dussumieri were equal for right-handed and left-handed. The occurrence of major chelipeds on either side in U. forcipata and U. dussumieri shows an antisymmetrical growth pattern. Major chelipeds showed positive allometry and minor chelipeds showed isometry for both species. This might have caused the growth imbalance between chelipeds that leads to the asymmetry of body shape.
REFERENCES
Agresti A. 2007. An Introduction to Categorical Data Analysis. 2nd ed. New York, NY: University of Florida.
Araujo MSL, Coelho PA, Castiglioni DA. 2012. Relative growth and determination of morphological sexual maturity of the fiddler crab Uca thayeri Rathbun (Crustacea, Ocypodidae) in two mangrove areas from the Brazilian tropical coast. PanamJAS.7:156-170.
Backwell PRY, Matsumasa M, Double M, Roberts A, Murai V, Keogh JS, Jennions MD. 2007. What are the consequences of being left-clawed in a predominantly right-clawed fiddler crab?. Proc R Soc B: Biol Sci.274:2723–2729.
Boundriansky R, Day T. 2003. The Evolution of Static Allometry in Sexually Selected Traits. Evol.57: 2450-2458.
Claude J. 2008. Morphometrics in R. France, FR: University de Montpellier II. Crane J. 1975. Fiddler Crabs of The World, Ocypodidae: genus Uca. Princeton,
US: Princeton University Pr.
Croll GA, McClintock JB. 2002. An analysis of cheliped asymmetry in three species of fiddler crabs. Gulf Mex Sci.20: 106-109.
Gebo DL. 1993. Postcranial Adaptation in Nonhuman Primates. DeKalb, US: Northern Illinois University Press.
Hirose GL, Junior EAB, Fransozo MLN. 2010. Post-lrval morphology, and development of Uca cumulata, 1943 (Crustacea, Decapoda, Ocypodidae) under laboratory conditions. Invertebr Reprod Dev. 54: 95-109.
Huxley JS. 1924. Constant differential growth-ratios and their significance. Nature.114:895-896.
10
Klingenberg PC. Multivariate Allometry. New York, US: Plenum Press; 1996. Martins SB, Masunari S. 2013. Relative growth in the fiddler crab Uca
uruguayensis Nobili, 1901 (Brachyura, Ocypodidae) from Garças River mangrove, Guaratuba Bay, southern Brazil. Nauplius. 21: 35-41.
Murniati DC. 2009. Perbandingan luas tutupan spoon tiped setae maksiliped kedua pada Uca spp. (Brachyura: Ocypodidae). Fauna Ind. 18:1-8.
Murniati DC. 2010. Pola dominansi capit pada Uca spp. (Decapoda: Ocypodidae). J Biol Sci Res.16:15-20.
Palmer AR. 1996. From symmetry to asymmetry: Prhylogenetic patterns of asymmetry variation in animals and their evolutionary significance. Proc Natl Acad Sci. 93:14279–14286.
Palmer AR. 2004. Symmetry Breaking and the Evolution of Development. Science.36:828-833.
Palmer AR. 2012. Developmental origins of normal and anomalous random right-left asymmetry; lateral inhibition versus developmental error in a threshold trait. Contrib Zool. 81: 111-124.
Pralon BGN, Nigreiros-Fransozo MC. 2007. Relative growth and morphological sexual maturity of Uca cumulate (Crustacea: Decapoda: Ocypodidae) from tropical Brazilian mangrove population. J Mar Biol Assoc U.K. 83: 569-574.
Rosenberg MS. 1997. Evolution of shape differences between the major and minor chelipeds of Uca pugnax (decapoda: Ocypodidae). J Crust Biol.17: 52-59.
Rosenberg MS. 2001. The systematics and taxonomy of fiddler crabs: a phylogeny of the genus Uca. J Crust Biol. 21: 839–869.
Rosenberg MS. 2002. Fiddler crab claw shape variation: a geometric morphometric analysis across the genus Uca (Crustacea: Brachyura: Ocypodidae). Biol J Linnean Soc.75:147-162.
Sokal RR, Rohlf FJ. 1969. Biometry, The Principles and Practice of Statistics in Biological Research. 3th ed. New York, US: State University of New York at Stony Brook.
Valen LV. 1962. A Study of Fluctuating Asymmetry. Evolution. 16: 125-142. Vanberg FJ, Costlow JD. 1966. Handedness in fiddler crabs (Genus Uca).
Crustaceana.11: 61-64.
Vieira RRR, Pinho GLL, Rieger PJ. 2010. Juvenile development of Uca (Minuca) Burgersi Holthuis, 1967 (Crustacea, Brachyura, Ocypodidae) in the laboratory. Atlantica (Rio Grande). 32: 59-70.
Warton DI, Remko AD, Daniel SF, Sara T. 2012. SMATR 3 – an R package for estimation and inference about allometric lines. Methods Ecol Evol. 3: 257–259.
Weis JS, Peddrick W. 2004. Behavior of four species of fiddler crabs, genus Uca, in southeast Sulawesi, Indonesia. Hydrobiologia. 34: 47-58.
11
12
APPENDIXES
11
Appendix 2 Linear regression equation between body features (CRW is predictor) in Uca forcipata and U. dussumieri
Relation between Species y = ax+b Slopes Intercepts
L vs CRW U. forcipata L = 1.705365 CRW - 1.897593 p ≤ 0.05 (Allometry)
Non-parallel U. dussumieri L = 1.973034 CRW - 2.838938 p ≤ 0.05 (Allometry)
W vs CRW U. forcipata W = 1.327799 CRW - 1.659258 p ≤ 0.05 (Allometry)
Parallel U. dussumieri W = 1.267016 CRW - 1.552041 p ≤ 0.05 (Allometry)
L1 vs CRW U. forcipata L1 = 1.025352 CRW - 1.073495 p ≥0.05 (Isometry)
Parallel U. dussumieri L1 = 1.042195 CRW - 1.080059 p ≥0.05 (Isometry)
W1 vs CRW U. forcipata W1 = 0.971302 CRW - 1.080059 p ≥0.05 (Isometry)
Parallel U. dussumieri W1 = 0.898517 CRW - 1.080059 p ≥0.05 (Isometry)
14