ANALISIS POPULASI UNDUR-UNDUR LAUT Hippa adactyla Fabricius, 1787 (CRUSTACEAN: HIPPIDAE) BERDASARKAN PENDEKATAN MORFOMETRIK
DAN GENETIK DI PANTAI BERPASIR CILACAP DAN KEBUMEN
WAHYU MUZAMMIL
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Analisis Populasi Undur-Undur Laut Hippa adactyla Fabricius, 1787 (Crustacean: Hippidae) Berdasarkan Pendekatan Morfometrik dan Genetik di Pantai Berpasir Cilacap dan Kebumen adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari Penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
WAHYU MUZAMMIL. Analisis Populasi Undur-Undur Laut Hippa adactyla Fabricius, 1787 (Crustacean: Hippidae) Berdasarkan Pendekatan Morfometrik dan Genetik di Pantai Berpasir Cilacap dan Kebumen. Dibimbing oleh YUSLI WARDIATNO dan NURLISA ALIAS BUTET.
Undur-undur laut (mole crab) hidup di ekosistem pantai berpasir zona intertidal. Organisme yang hidup di lingkungan ini salah satunya beradaptasi dengan meliangkan dirinya di pasir, guna membantu mereka bertahan hidup. Wilayah pesisir Indonesia merupakan salah satu daerah sebaran undur-undur laut terutama famili Hippidae, salah satunya di pesisir selatan Jawa Tengah terutama di Kabupaten Cilacap dan Kebumen. Masyarakat dikedua daerah ini memanfaatkan undur-undur laut sebagai umpan memancing dan bahan makanan khas berupa rempeyek. Pengambilan sampel undur-undur laut dilakukan di bagian pasang surut (zona intertidal) pantai berpasir di Kecamatan Adipala, Kabupaten Cilacap dan pantai berpasir di Kecamatan Buluspesantren, Kabupaten Kebumen. Pengambilan sampel dilakukan pada bulan Agustus 2014 dengan menggunakan alat tangkap sorok dengan luas sapuan sepanjang 3 km (1.5 km dengan satu kali ulangan) menyusuri pantai.
Total Hippa adactyla yang ditangkap dan dianalisis sebanyak 118 individu asal Cilacap yang terdiri atas 27 jantan (23%), 91 betina (77%); sebanyak 102 individu asal Kebumen yang terdiri atas 46 jantan (45%) dan 56 betina (55%). Kedua populasi ini memiliki nilai rasio panjang dan lebar karapas di atas 1, hal ini menunjukkan pergerakan H. adactyla bergerak dominan dengan arah vertikal (backward locomotion). Terdapat delapan rasio karakter morfometrik yang dapat dijadikan penciri utama yang membedakan undur-undur laut asal Cilacap dan Kebumen berdasarkan analisis diskriminan. Pusat sebaran karakter morfometrik Cilacap (1.042) terpisah dengan pusat sebaran karakter morfometrik Kebumen (-1.205).
Amplifikasi gen 16S rRNA dari kedelapan DNA total H. adactyla dilakukan dengan mesin PCR pada suhu penempelan primer (annealing) 45oC, dimana kedelapan sampel ini dikategorikan sebagai kelompok kaya A-T (A-T rich) berdasarkan komposisi basa nukleotida. Terdapat 54 situs mutasi pada gen 16S rRNA antara H. adactyla asal Cilacap dan Kebumen dibandingkan dengan H. adactyla (KF051307.1) yang berasal dari luar Indonesia, terdiri dari 3 situs mutasi insersi, 2 situs mutasi delesi, 29 situs transisi, dan 20 situs transversi. Terdapat 26 situs spesifik pada H. adactyla asal Cilacap dan Kebumen jika dibandingkan dengan H. adactyla yang berasal dari luar Indonesia serta Emerita talpoida (KF182557.1) yang diperoleh dari GenBank.
Tidak adanya jarak genetik antara spesies H. adactyla asal Cilacap dan Kebumen mengindikasikan antara H. adactyla asal Cilacap dan Kebumen diperkirakan merupakan satu populasi. Pola arus di perairan Cilacap dan Kebumen memiliki peran yang sangat penting dalam hal konektivitas H. adactyla asal Cilacap dan Kebumen.
SUMMARY
WAHYU MUZAMMIL. Population Analysis of Mole Crab Hippa adactyla Fabricius, 1787 (Crustacean: Hippidae) Based On Morphometric and Genetic Study in Cilacap and Kebumen Sandy Beach. Supervised by YUSLI WARDIATNO and NURLISA ALIAS BUTET.
Mole crab (Hippa adactyla) lives in the intertidal zone of sandy beach ecosystems. Mole crab usually burries into substrate to cope with environmental stress. Indonesian coastal waters is one of the mole crab distribution area, especially families Hippidae, such as in the south coast of Central Java, especially in Cilacap and Kebumen sandy beach. Cilacap and Kebumen communities utilizing mole crab as bait fishing and food ingredients such as ‘rempeyek’.
The samples was carried out on August 2014 at intertidal zone of Adipala sandy beach in Cilacap district and intertidal zone of Buluspesantren sandy beach in Kebumen district. Samples were caught using traditional fishing gear called ‘sorok’ along 3 km (1.5 km with one repetition).
Total Hippa adactyla were 118 individuals which caught in Cilacap that consisted of 27 males (23%) and 91 females (77%); while 102 individuals that were caught in Kebumen consisted of 46 males (45%) and 56 females (55%). Carapace length to width ratio is above 1 in both population, it shows that the locomotion of H. adactyla is dominant in the vertical direction (backward locomotion).
There were eight morphometric characters ratio that can be used as major factors to identify both populations based on discriminant analysis such as total length:carapace length (TL:CL), carapace width:carapace length (CW:CL), left and right coxa length:carapace length (LCo/RCo:CL), left merus length:carapace length (LM:CL), left and right propodus length:carapace length (LP/RP:CL), and left dactylus length:carapace length (LD:CL). Group centroid of morphometric characters in Cilacap (1.042) were separate to group centroid of morphometric characters in Kebumen (-1.205).
16S rRNA gene amplification of eight H. adactyla DNA by PCR machine at 450C annealing temperature. Composition of the nucleotide mtDNA 16S rRNA gene of H. adactyla shows rich of adenine (A) and thymine (T) (A-T rich). There are 54 mutations in the 16S rRNA gene between H. adactyla from Cilacap and Kebumen compared with H. adactyla (KF051307.1) from outside of Indonesia, consists of three sites of insertion, two sites of deletion, twenty nine sites of transition, and twenty sites of transversion. There are 26 specific sites on H. adactyla from Cilacap and Kebumen compared with H. adactyla from outside of Indonesia and Emerita talpoida (KF182557.1) data from GenBank.
There is no genetic distance between species H. adactyla from Cilacap dan Kebumen that indicates there are estimated one population. Current pattern in Cilacap and Kebumen are important role in connectivity between H. adactyla from Cilacap and Kebumen.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pengelolaan Sumberdaya Perairan
ANALISIS POPULASI UNDUR-UNDUR LAUT Hippa adactyla FABRICIUS, 1787 (CRUSTACEAN: HIPPIDAE) BERDASARKAN PENDEKATAN MORFOMETRIK DAN GENETIK DI PANTAI BERPASIR CILACAP DAN
KEBUMEN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
Judul Penelitian : Analisis Populasi Undur-Undur Laut Hippa adactyla Fabricius, 1787 (Crustacean: Hippidae) Berdasarkan Pendekatan Morfometrik dan Genetik di Pantai Berpasir Cilacap dan Kebumen
Nama : Wahyu Muzammil
NIM : C251120181
Disetujui oleh Komisi Pembimbing
Dr Ir Yusli Wardiatno, MSc Ketua
Dr Ir Nurlisa A. Butet, MSc Anggota
Diketahui oleh
Ketua Program Studi Pengelolaan Sumberdaya Perairan
Dr Ir Sigid Hariyadi, MSc
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MscAgr
PRAKATA
Puji syukur kepada Allah swt, karena atas izin-Nya Penulis dapat menyajikan tulisan ilmiah berdasarkan kegiatan penelitian yang dilakukan sejak Agustus 2014. Karya ilmiah ini merupakan pengembangan bidang ilmu biologi dan molekuler perikanan yang berjudul Analisis Populasi Undur-Undur Laut Hippa adactyla Fabricius, 1787 (Crustacean: Hippidae) Berdasarkan Pendekatan Morfometrik dan Genetik di Pantai Berpasir Cilacap dan Kebumen.
Pelaksanaan penelitian dan penulisan karya ilmiah ini tidak lepas dari bantuan dan dukungan dari banyak pihak. Oleh karena itu, Penulis mengucapkan terima kasih kepada:
1. Institut Pertanian Bogor (IPB) yang telah menyediakan berbagai fasilitas sehingga penelitian ini dapat diselesaikan.
2. Dr Ir Enan M Adiwilaga selaku Ketua Program Studi SDP untuk tahun studi 2010-2013 yang telah membantu tahapan penyelesaian studi dan penelitian. 3. Dr Ir Sigid Hariyadi, MSc selaku Ketua Program Studi SDP untuk tahun studi
2014-2017 yang telah membantu tahapan penyelesaian studi dan penelitian. 4. Dr Ir Yusli Wardiatno, MSc dan Dr Ir Nurlisa A. Butet, MSc selaku dosen
pembimbing yang telah memberikan arahan dan masukan kepada Penulis dari tahap awal pelaksanaan penelitian sampai pada tahap akhir penulisan karya ilmiah ini.
5. Dr Ir Isdradjad Setyobudiandi, MSc selaku dosen penguji dari program studi yang telah memberikan masukan dan saran untuk penyempurnaan tulisan ini. 6. Dr Majariana Krisanti, SPi MSi selaku dosen perwakilan dari program studi
yang telah memberikan masukan dan saran untuk penyempurnaan tulisan ini. 7. Seluruh keluarga, terutama kepada Mama, Papa, Teh Lian, Teh Fani, Ridha,
Bang Ulas, My Nephew and Niece (Amer, Ayesha, Maher, Akmal), saudara (Mba Ita, Mas Yadin, Uak Nur, A Opik) atas doa dan dukungan yang tidak pernah putus sehingga tulisan ini berhasil diselesaikan.
8. Seluruh kolega laboratorium biologi molekular akuatik MSP IPB (Panji, Findra, Agus, Yuyun, Syamsul, Dewi, Siska, Lella, Fajrin, Lita, Yustin, Lusita, Febi, Ningsih, Dani Ajeng), staf laboratorium terpadu FPIK IPB (Ani dan Faqih), unconditionally buddies (Fuquh, Panji, Bambang, Dede, Alim, Dita, Nta, Mba Pepen, Bang Apri, Julmi), Keluarga Pak Sarmo (Kebumen) serta Keluarga Pak Sugeng (Cilacap) atas kerjasamanya dalam pengambilan sampel penelitian.
9. Seluruh rekan SDP 2011, SDP 2012, SDP 2013, rekan pondok d’qaka serta teman-teman lainnya yang tidak dapat disebutkan satu per satu atas dukungan yang telah diberikan.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan dan Manfaat Penelitian 3
2 METODE 4
Lokasi dan Waktu Penelitian 4
Bahan dan Alat 4
Pengambilan dan Identifikasi Sampel 4
Rasio Morfometrik 5
Analisis Hubungan Panjang-Bobot 6
Marka Gen 16S rRNA 7
Isolasi dan Ekstraksi DNA 7
Amplifikasi mtDNA dengan PCR 7
Produk PCR 8
Sekuensing dan Analisis Data Molekuler 8
Konservasi 8
3 HASIL DAN PEMBAHASAN 9
Analisis Diskriminan 9
Hubungan Panjang-Bobot 12
Amplifikasi DNA Gen 16S rRNA 13
Sekuensing DNA dan Pensejajaran Urutan Basa Nukleotida 14
Analisis Jarak Genetik dan Filogeni 15
Situs Mutasi Gen 16S rRNA H. adactyla 17
Nukleotida Spesifik Gen 16S rRNA H. adactyla 17
Konektivitas 17
Konservasi 18
4 KESIMPULAN DAN SARAN 20
Kesimpulan 20
Saran 20
DAFTAR PUSTAKA 21
LAMPIRAN 24
DAFTAR TABEL
1 Nilai minimum, rataan (±SD), dan maksimum panjang karapas (PK) serta bobot basah (BB) H. adactyla jantan dan betina di Cilacap dan
Kebumen 9
2 Fungsi diskriminan, nilai maksimum, minimum dan rataan untuk karakter morfometrik H. adactyla asal Cilacap dan Kebumen 10 3 Persamaan regresi linear antara tiga belas variabel morfometrik (PK
sebagai variabel acuan) pada H. adactyla asal Cilacap dan Kebumen,
dengan hasil ANCOVA. 12
4 Kisaran panjang karapas dan pola pertumbuhan H. adactyla di perairan
pantai berpasir Cilacap dan Kebumen 13
5 Jarak genetik fragmen gen 16S rRNA pada H. adactyla yang berasal dari Cilacap (C1, C2, C3, dan C4) dan Kebumen (K1, K2, K3, dan K4), serta H. adactyla dan Emerita talpoida yang diperoleh dari
GenBank (*) 16
DAFTAR GAMBAR
1 Lokasi Pengambilan Sampel 4
2 Karakter morfometrik yang diukur 5
3 Grafik fungsi diskriminan karakter morfometrik H. adactyla. 11 4 Visualisasi produk PCR dengan satu kali ulangan (kiri: amplifikasi
pertama; kanan: amplifikasi kedua) pada gel agarosa 1.2%, kolom kiri sampai kanan: marker, sampel Cilacap (1, 2, 3, 4), sampel kebumen (5,
6, 7, 8) 14
5 Konstruksi filogeni berdasarkan gen 16S rRNA pada genus Hippa (H. adactyla asal Cilacap, Kebumen, dan yang berasal dari luar Indonesia) dengan genus Emerita (E. talpoida).
16
6 Pola arus Pulau Jawa berdasarkan olahan data real time Ocean Surface Current Analyses NOAA pada tahun 2014 (a) dan pada Bulan Agustus 2014 (b) berasal dari Cilacap dan Kebumen dibandingkan dengan H. adactyla yang berasal dari luar Indonesia yang didapatkan dari GenBank (KF051307.1)
24
3 Situs Nukleotida Spesifik gen 16S rRNA Hippa adactyla yang berasal dari Cilacap dan Kebumen.
25
1
1
PENDAHULUAN
Latar Belakang
Undur-undur laut (mole crab) hidup di ekosistem pantai berpasir, lingkungan ini bukanlah tempat yang mudah bagi organisme untuk hidup, karena tekanan lingkungan seperti ombak, perubahan pasang surut, dan predator. Hewan-hewan yang hidup di lingkungan ini salah satunya beradaptasi dengan meliangkan dirinya di pasir guna membantu mereka bertahan hidup. Undur-undur laut banyak ditemukan di Amerika Selatan (Defeo et al. 2001; Boere et al. 2011; Veas et al. 2014), Amerika Utara (Dugan and Hubbard 1996; Amend and Shanks 1999), Taiwan (Chan et al. 2010), dan Thailand (Boonruang and Phasuk 1975). Wilayah pesisir Indonesia merupakan salah satu daerah sebaran undur-undur laut terutama famili Hippidae, diantaranya yang sudah dipublikasikan oleh Wardiatno et al. (2015) adalah di pesisir Sumatera (Aceh Barat, Padang, dan Bengkulu), pesisir Jawa (Pelabuhanratu dan Cilacap), pesisir Bali (Pantai Lebih dan Singaraja), pesisir Nusa Tenggara Barat (Gili Meno), pesisir Sulawesi (Tangkoko, Pantai Talise, Lero, Banggai, dan Buton), dan Maluku Tenggara (Kepulauan Kei), serta pesisir selatan Yogyakarta (Mursyidin 2007). Selain itu, undur-undur laut ini juga sudah bernilai ekonomis terutama di pesisir selatan Jawa Tengah, yaitu di Kabupaten Cilacap dan Kebumen.
Undur-undur laut atau biasa disebut yutuk (Cilacap dan Kebumen), tempenyol (Lombok), dan penyon (Bali) biasa digunakan masyarakat Cilacap dan Kebumen sebagai umpan memancing dan dijadikan bahan konsumsi untuk dijadikan rempeyek. Selain dimanfaatkan sebagai sumber protein, undur-undur laut juga dapat dimanfaatkan sebagai bio-indikator pencemaran (Powell et al. 2002). Penelitian undur-undur laut di Indonesia belum banyak dilakukan, sehingga data dan informasi mengenai undur-undur laut yang ada di Indonesia sangat minim. Padahal undur-undur laut merupakan sumberdaya penting dalam siklus rantai makanan, kepiting ini merupakan konsumen tingkat awal dalam tingkat trofik level di daerah pantai berpasir (Lercari and Defeo 1999).
2
dibandingkan sekuen COI (An et al. 2005) dan menurut Quan et al. (2004), fragmen gen 16S rRNA cocok untuk memeriksa hubungan filogenetik pada level spesies atau genus pada krustasea.
Informasi dasar mengenai kekerabatan dan penciri (spesies dan populasi) undur-undur laut di Indonesia melalui pendekatan karakter morfometrik dan karakter molekuler sampai saat ini belum ada yang melakukan, padahal kajian seperti ini diperlukan sebagai informasi dasar rasio morfometrik, taxonomy certainty (kepastian taksonomi), dan population certainty (kepastian populasi) yang dapat menjadi dasar pengelolaan sumberdaya undur-undur laut di pantai berpasir Cilacap dan Kebumen. Oleh sebab itu perlu adanya kajian tentang analisis populasi Hippa adactyla berdasarkan pendekatan morfometrik dan genetik sebagai acuan kepastian populasi, konektivitas dan filogeografi dalam pengelolaan sumberdaya undur-undur laut di pantai berpasir Cilacap dan Kebumen.
Perumusan Masalah
Saat ini penggunaan karakter morfometrik tidaklah cukup untuk mengidentifikasi suatu spesies. Penggunaan karakter morfometrik pada biota akuatik seringkali memperlihatkan fenomena cryptic species (hampir mirip) dan kompleks spesies sehingga dapat mengakibatkan masalah sinonim yaitu terdapat nama ganda pada satu spesies yang sama atau sebaliknya (Bickford et al. 2006). Kepastian taksonomi dan populasi atau taxonomy and population certainty sangat diperlukan dalam acuan pengelolaan berbasis wilayah pengelolaan perikanan (WPP), mengingat sangat memungkinkan adanya spesies dengan populasi yang berbeda ataupun populasi yang sama dalam satu wilayah pengelolaan. Pendekatan molekuler menjadi salah satu metode yang populer dan berkembang pesat saat ini dalam mengatasi masalah kepastian taksonomi dan populasi yang cepat dan akurat dengan menggunakan fragmen urutan nukleotida yang pendek.
Wilayah pantai berpasir di Indonesia merupakan salah satu habitat undur-undur laut, diantaranya ditemukan di pantai berpasir Cilacap dan Kebumen (Jawa Tengah). Kedekatan secara geografis dari perairan pesisir Cilacap dan Kebumen memungkinkan adanya konektivitas Hippa adactyla melalui salah satu stadia perkembangan hidupnya. Larva H. adactyla yang bersifat meroplankton dapat menempati lokasi yang berbeda dari lokasi asal penyebaran, dengan demikian konektivitas genetik bukan suatu hal yang mustahil. Kajian molekuler sangat membantu dalam memecahkan permasalahan kepastian taksonomi dan populasi yang sering kali menjadi kendala permasalahan bagi unit pengelolaan dan konservasi.
3
Tujuan dan Manfaat Penelitian
4
2
METODE
Lokasi dan Waktu Penelitian
Sampel undur-undur laut (Hippa adactyla) ditangkap dari daerah pantai berpasir selatan Jawa Tengah, meliputi pantai berpasir di Kecamatan Adipala, Kabupaten Cilacap dan pantai berpasir di Kecamatan Buluspesantren, Kabupaten Kebumen (Gambar 1). Pengambilan sampel undur-undur laut dilakukan pada bulan Agustus 2014 dengan menggunakan alat tangkap sorok dengan lebar sapuan sekitar 60-80 cm dan luas sapuan sepanjang 3 km (1.5 km dengan satu kali ulangan) menyusuri pantai. Data primer morfometrik yang diukur mencakup data panjang baku, bobot basah, jenis kelamin, panjang karapas, lebar karapas, panjang coxa (kanan dan kiri), panjang basis (kanan dan kiri), panjang merus (kanan dan kiri), panjang carpus (kanan dan kiri), panjang propodus (kanan dan kiri) dan panjang dactylus (kanan dan kiri). Analisis molekuler dilakukan di Laboratorium Biologi Molekuler Departemen Manajemen Sumber daya Perairan IPB dan Laboratorium Terpadu Fakultas Perikanan dan Ilmu Kelautan IPB.
Gambar 1. Lokasi Pengambilan Sampel
Bahan dan Alat
Bahan dan alat yang digunakan selama penelitian adalah alkohol 95%, mistar, jangka sorong, timbangan digital, botol sampel, cool box, gunting bedah, pinset, tube 1.5 ml, pengocok (shaker), mikro tip, mikro pipet, pistil, pemanas inkubator, sentrifuse, spin column, mesin visual ultraviolet, agarose 1.2%, akuades, kit Gene Aid, EtOH absolut, reagen PCR, sampel undur-undur laut (Hippa adactyla), dan buku identifikasi superfamili Hippoidea.
Pengambilan dan Identifikasi Sampel
5 dengan mengamati karakter morfometrik undur-undur laut dan disesuaikan dengan karakteristik yang ada pada buku identifikasi superfamili Hippoidea (Chan et al. 2010) dan juga pengiriman sampel yang diidentifikasi langsung oleh Boyko (mole crab scientist expert) pada tahun 2013 untuk memastikan identifikasi secara morfometrik sudah tepat. Sampel H. adactyla selanjutnya dipisahkan antara jantan (tanpa pleopod) dan betina (memiliki sepasang pleopod).
Rasio Morfometrik
Karakter morfometrik (Gambar 2) yang diukur menggunakan mistar (0.5 mm) dan jangka sorong (0.05 mm) mengacu pada Boyko and Harvey (1998) serta ilustrasi karakter morfologi superfamili Hippoidea (Chan et al. 2010) yang meliputi panjang karapas (PK), lebar karapas (LK), panjang coxa kanan (PCoKa) dan kiri (PCoKi), panjang basis kanan (PBKa) dan kiri (PBKi), panjang merus kanan (PMKa) dan kiri (PMKi), panjang carpus kanan (PCaKa) dan kiri (PCaKi), panjang propodus kanan (PPKa) dan kiri (PPKi), panjang dactylus kanan (PDKa) dan kiri (PDKi), serta bobot basah (BB). Untuk menghindari diferensiasi yang berkaitan dengan struktur umur pada setiap sampel, maka seluruh karakter morfometrik (kecuali panjang karapas) dirasiokan terhadap panjang karapas (PK). Rasio karakter morfometrik ini kemudian dianalisa untuk mengetahui rasio mana yang dapat digunakan sebagai penciri H. adactyla antar kedua populasi.
Gambar 2. Karakter morfometrik yang diukur
Pada spesies kepiting, karapas merupakan lokomotif untuk menentukan arah pergerakan kepiting (Vidal-Gadea et al. 2008). Pendekatan rasio panjang dan lebar karapas berkaitan dengan preferensi arah jalan kepiting (sideways atau forward/backward). Menurut Vidal-Gadea et al. (2008), rasio ini memberi informasi preferensi pergerakan kepiting (rasio > 1 = forward/backward locomotion; rasio < 1 = sideways locomotion).
Analisis diskriminan digunakan untuk mendeskripsikan pola pusat sebaran undur-undur laut (H. adactyla) berdasarkan empat belas karakter morfometrik yang dirasiokan terhadap panjang karapas. Analisis diskriminan menggunakan empat belas variabel morfometrik H. adactyla yang berperan sebagai variabel independen secara bersama-sama (simultan) yang mampu dengan
PK
6
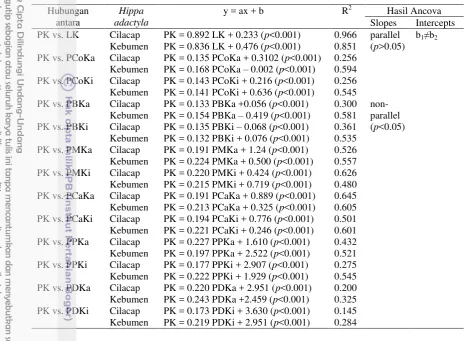
baik membedakan dan memprediksi pola keanekaragaman morfometrik dan populasi undur-undur laut di Cilacap dan Kebumen. Keanekaragaman morfometrik H. adactyla dianalisis menggunakan metode analisis diskriminan yang terdapat dalam perangkat lunak SPSS 16.0 untuk melihat pengelompokkan karakter. Selanjutnya untuk memperkuat hasil analisis diskriminan digunakan analysis of covariance (ANCOVA) untuk membandingkan hubungan linear panjang karapas (PK) dengan ketiga belas karakter morfometrik lainnya antara H. adactyla asal Cilacap dan Kebumen apakah memiliki slope dan intersep yang sama atau tidak.
Analisis Hubungan Panjang-Bobot
Analisis hubungan panjang dan bobot basah untuk mengetahui pola pertumbuhan H. adactyla.Model hubungan panjang bobot mengikuti pola hukum kubik dari dua parameter yang dianalisis. Asumsi hukum kubik ini adalah bahwa idealnya setiap pertambahan panjang akan menyebabkan pertambahan berat, sehingga untuk menganalisis hubungan panjang bobot H. adactyla menggunakan pendugaan sebagai berikut (Effendie, 2002):
Keterangan: W = Bobot basah H. adactyla (g) L = Panjang baku H. adactyla (mm) a, b = konstanta
Korelasi parameter dari hubungan panjang-berat dapat dilihat dari nilai konstanta b, yaitu:
Nilai b = 3, menunjukkan pola pertumbuhan isometrik Nilai b ≠ 3, menunjukkan pola pertumbuhan allometrik:
Jika b>3, maka pola pertumbuhan allometrik positif (pertumbuhan berat dominan)
Jika b<3, maka pola pertumbuhan allometrik negatif (pertumbuhan panjang dominan).
Kemudian dilakukan uji t untuk lebih menguatkan pengujian hipotesis berdasarkan Walpole (1992) dengan rumus sebagai berikut:
Keterangan:
Sb1 = Simpangan baku b1
b0 = Intercept b1 = Slope Dengan hipotesis:
H0 : b = 3 (isometrik)
7 Nilai thitung akan dibandingkan dengan nilai ttabel sehingga keputusan yang
akan diambil adalah sebagai berikut: thitung > ttabel, maka Tolak H0
thitung < ttabel, maka Gagal Tolak H0
Keeratan hubungan panjan-berat H. adactyla ditunjukkan oleh nilai koefisien korelasi (r). Nilai r yang mendekati 1 (r>0,7) menggambarkan hubungan yang erat antar keduanya, sedangkan nilai r yang mendekati 0 atau menjauhi 1 (r<0,7) menggambarkan hubungan yang tidak erat antar keduanya (Walpole 1992).
Marka Gen 16s rRNA
Saat ini penggunaan 16S rRNA telah banyak dilakukan dan telah menjadi prosedur baku untuk menentukan hubungan filogenetik dan menganalisis suatu ekosistem. Data urutan basa gen penyandi 16S rRNA memungkinkan digunakan untuk mengkonstruksi pohon filogenetik yang dapat menunjukkan nenek moyang dan hubungan kekerabatan organisme, tetapi organisme yang sekerabat atau identik berdasarkan parameter ini belum tentu memiliki kesamaan secara fisiologi. Hal ini disebabkan gen penyandi 16S rRNA bukan merupakan suatu gen yang fungsional untuk kelangsungan hidup dan adaptasi pada lingkungan tertentu. Molekul 16S rRNA memiliki beberapa daerah yang memiliki urutan basa yang relatif konservatif dan beberapa daerah urutan basanya variatif. Beberapa penelitian yang menggunakan gen 16S rRNA diantaranya Haye et al. (2002) mengenai analisis filogeni undur-undur laut (Hippidae: Emerita); Devine et al. (2012) mengenai analisis metagenomik keragaman bakteri pada Elysia chlorotica dan Vaucheria litorea; Goffredi et al. (2014) mengenai variasi ontogenetic: struktur komunitas epibion pada Kiwa puravida.
Isolasi dan Ekstraksi DNA
Isolasi dan ekstraksi DNA menggunakan kit komersial Gene Aid. Sampel jaringan yang digunakan sebagai sumber DNA adalah jaringan otot dari bagian telson Hippa adactyla. Proses pencucian alkohol pengawet dilakukan dengan cara merendam sekitar 30 mg potongan jaringan otot dalam aquades steril yang divortex dan dilakukan proses pengulangan sebanyak sepuluh kali. Selanjutnya prosedur ekstraksi ini mengikuti manual kit Gene Aid hingga didapatkan DNA yang telah berhasil diekstraksi.
Amplifikasi mtDNA dengan PCR
8
data). Amplifikasi ini pada suhu predenaturasi 94oC selama 5 menit, suhu denaturasi 94oC selama 1 menit, suhu penempelan 45oC selama 1 menit, suhu pemanjangan 72oC selama 1 menit, suhu pasca pemanjangan 72oC selama 5 menit, dan suhu penyimpanan 15oC selama 10 menit.
Produk PCR
Hasil PCR dapat langsung dianalisis dengan elektroforesis atau disimpan di dalam lemari pendingin. Bahan-bahan yang digunakan pada proses elektroforesis adalah gel agarosa 1.2%, TAE 1X, produk PCR, loading buffer dan penanda DNA. Sampel produk PCR sebanyak 2,5 µl dimasukkan ke dalam mesin elektroforesisuntuk dijalankan selama 60 menit, kemudian divisualisasi di bawah monitor UV.
Sekuensing dan Analisis Data Molekuler
Produk PCR dengan integritas yang baik dikirimkan ke perusahaan jasa sekuensing. Semua urutan basa nukleotida yang diperoleh akan dicocokkan di GenBank. Analisis data menggunakan perangkat lunak Mega versi 5 (Tamura et al. 2011) untuk menganalisis jarak genetik dan pohon kekerabatan.
Konservasi
9
3 HASIL DAN PEMBAHASAN
Analisis Diskriminan
Total Hippa adactyla yang ditangkap dan dianalisis sebanyak 118 individu asal Cilacap dan 102 individu asal Kebumen, yang terdiri atas 27 jantan (23%) dan 91 betina (77%) untuk H. adactyla asal Cilacap dan 46 jantan (45%) serta 56 betina (55%) untuk H. adactyla asal Kebumen. Rataan panjang karapas untuk jantan asal Cilacap adalah 24.91 ± 1.57 mm dan jantan asal Kebumen 22.17 ± 2.17 mm. Rataan panjang karapas untuk betina asal Cilacap adalah 25.51 ± 3.65 mm dan betina asal Kebumen 23.88 ± 2.20 mm. Rataan bobot basah (BB) untuk jantan asal Cilacap adalah 5.09 ± 0.65 g dan jantan asal Kebumen 3.65 ± 1.02 g. Rataan BB untuk betina asal Cilacap adalah 4.68 ± 2.18 g dan betina asal Kebumen 4.55 ± 1.35 g (Tabel 1). Penelitian Megawati (2012) menunjukkan undur-undur laut genus Hippa di Kebumen memiliki nisbah kelamin jantan sebesar 46% dan nisbah kelamin betina sebesar 54%. Jenis Emerita emeritus di Kebumen memiliki nisbah kelamin betina sebesar 91% dan nisbah kelamin jantan sebesar 9%; Undur-undur laut genus Hippa di Kebumen memiliki nisbah kelamin betina sebesar 54% dan nisbah kelamin jantan sebesar 46%.
10
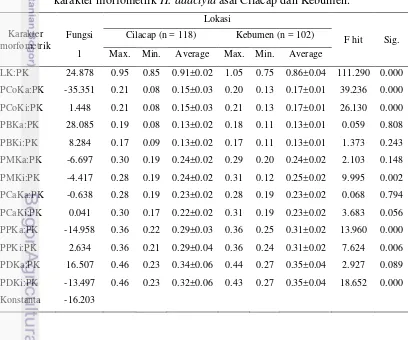
Analisis diskriminan berdasarkan perhitungan rasio tiga belas karakter morfometrik menggunakan koefisien fungsi diskriminan disajikan pada Tabel 2. Fungsi linear dari koefisien diskriminan dapat digunakan untuk memastikan karakter morfometrik yang diukur dari spesies H. adactyla masuk kedalam populasi Cilacap atau Kebumen berdasarkan formula diskriminan yang dihasilkan. Hasil analisis diskriminan menunjukkan rasio LK:PK, PCoKa:PK, PCoKi:PK, PMKi:PK, PPKa:PK, PPKi:PK, dan PDKi:PK dapat kita jadikan penciri utama yang membedakan undur-undur laut asal Cilacap dan Kebumen. Ketujuh karakter tersebut layak digunakan menjadi penciri utama disebabkan oleh karakter-karakter tersebut berbeda nyata antara Cilacap dan Kebumen (dilihat dari nilai signifikannya).
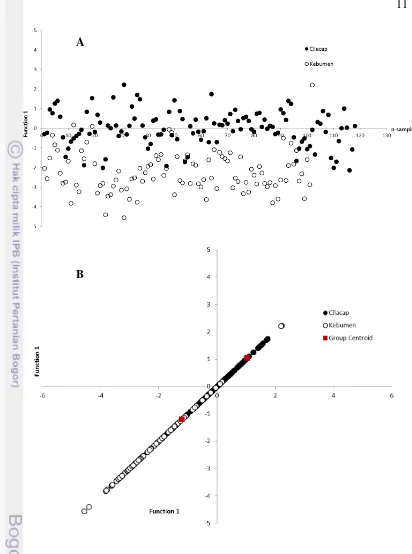
Berdasarkan analisis diskriminan rasio ketiga belas karakter morfometrik didapatkan perbedaan pusat sebaran dari H. adactyla asal Cilacap dan Kebumen. Grafik analisis diskriminan (Gambar 3) memperlihatkan pusat sebaran karakter morfometrik Cilacap (1.042) terpisah dengan pusat sebaran karakter morfometrik Kebumen (-1.205). Beberapa individu undur-undur laut Cilacap mendekati pusat sebaran karakter morfometrik undur-undur laut Kebumen (n = 13). Hal yang sama terjadi pula di Kebumen, beberapa individu undur-undur laut Kebumen mendekati pusat sebaran karakter morfometrik undur-undur laut Cilacap (n = 13).
Tabel 2. Fungsi diskriminan, nilai maksimum, minimum dan rataan untuk karakter morfometrik H. adactyla asal Cilacap dan Kebumen.
Karakter morfometrik
Lokasi
F hit Sig.
Fungsi Cilacap (n = 118) Kebumen (n = 102)
1 Max. Min. Average Max. Min. Average
LK:PK 24.878 0.95 0.85 0.91±0.02 1.05 0.75 0.86±0.04 111.290 0.000
PCoKa:PK -35.351 0.21 0.08 0.15±0.03 0.20 0.13 0.17±0.01 39.236 0.000
PCoKi:PK 1.448 0.21 0.08 0.15±0.03 0.21 0.13 0.17±0.01 26.130 0.000
PBKa:PK 28.085 0.19 0.08 0.13±0.02 0.18 0.11 0.13±0.01 0.059 0.808
PBKi:PK 8.284 0.17 0.09 0.13±0.02 0.17 0.11 0.13±0.01 1.373 0.243
PMKa:PK -6.697 0.30 0.19 0.24±0.02 0.29 0.20 0.24±0.02 2.103 0.148
PMKi:PK -4.417 0.28 0.19 0.24±0.02 0.31 0.12 0.25±0.02 9.995 0.002
PCaKa:PK -0.638 0.28 0.19 0.23±0.02 0.28 0.19 0.23±0.02 0.068 0.794
PCaKi:PK 0.041 0.30 0.17 0.22±0.02 0.31 0.19 0.23±0.02 3.683 0.056
PPKa:PK -14.958 0.36 0.22 0.29±0.03 0.36 0.25 0.31±0.02 13.960 0.000
PPKi:PK 2.634 0.36 0.21 0.29±0.04 0.36 0.24 0.31±0.02 7.624 0.006
PDKa:PK 16.507 0.46 0.23 0.34±0.06 0.44 0.27 0.35±0.04 2.927 0.089
PDKi:PK -13.497 0.46 0.23 0.32±0.06 0.43 0.27 0.35±0.04 18.652 0.000
11
B A
Gambar 3. Grafik fungsi diskriminan karakter morfometrik H. adactyla tanpa group centroid (A) dan dengan group centroid (B).
12
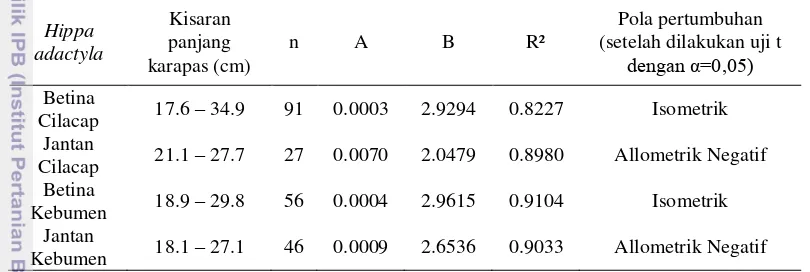
Selama pengamatan diketahui bahwa panjang karapas minimum dan maksimum H. adactyla betina Cilacap adalah 17.6 mm dan 34.9 mm, panjang karapas minimum dan maksimum H. adactyla jantan Cilacap adalah 21.1 mm dan 27.7 mm, sedangkan panjang karapas minimum dan maksimum H. adactyla betina Kebumen adalah 18.9 cm dan 29,8 mm, serta panjang karapas minimum dan maksimum H. adactyla jantan Kebumen adalah 18.1 mm dan 27.1 mm.
13 Menurut hasil analis menunjukan bahwa hubungan panjang bobot pada undur-undur laut baik jenis H. adactyla Cilacap maupun Kebumen memiliki hubungan korelasi yang sangat erat, hal ini terlihat dari nilai korelasi yang mendekati satu. Pertambahan panjang karapas undur-undur laut akan berpengaruh terhadap bobot tubuhnya. H. adactyla jantan dan betina di kedua populasi memiliki perbedaan dalam hal pola pertumbuhan. Undur-undur laut betina di kedua populasi memiliki pola pertumbuhan isometrik dimana pertumbuhan panjang karapas dan bobot seimbang. Undur-undur laut jantan Cilacap dan Kebumen menunjukkan pola perumbuhan yang berbeda yaitu allometrik negatif, yang artinya pertumbuhan panjang karapas lebih cepat dibandingkan dengan pertumbuhan bobot undur-undur laut.
Tabel 4. Kisaran panjang karapas dan pola pertumbuhan H. adactyla di perairan pantai berpasir Cilacap dan Kebumen
Hippa dibandingkan dengan undur-undur laut jantan. Hal ini mungkin terjadi karena perbedaan upaya reproduksi antar jenis kelamin. Pada undur-undur laut betina, lebih dari 50% ditemukan dalam kondisi menyimpan telur yang terlindungi oleh telson. Hal ini memungkinkan undur-undur laut betina memiliki bobot yang lebih tinggi dibandingkan dengan undur-undur laut jantan. Undur-undur laut betina dan jantan memiliki perilaku bergerak aktif meliangkan tubuhnya di zona pasang surut sehingga energinya banyak terpakai dalam aktivitas bergerak aktif. Menurut Pinheiro and Fransozo (1999), pada kepiting jantan jenis Arenaeus cribrarius biasanya memiliki perilaku menjaga pasangannya selama dan setelah kopulasi serta menjaga pasangannya saat fase setelah molting. Hal ini mungkin saja terjadi pada undur-undur laut jantan H. adactyla yang menyebabkan energinya banyak terpakai untuk menjaga pasangannya.
Amplifikasi DNA gen 16S rRNA
14
Berdasarkan isolasi dan ekstraksi DNA yang dilakukan diperoleh DNA total dengan kualitas yang baik pada keempat contoh H. adactyla asal Cilacap dan Kebumen. DNA total dengan kualitas yang baik ditunjukkan oleh keberadaan pita DNA yang terang dan tebal.
DNA total H. adactyla yang telah berhasil diisolasi dan diekstraksi dengan kualitas yang baik ini selanjutnya dapat dijadikan cetakan DNA dalam tahap amplifikasi gen 16S rRNA yang dilakukan menggunakan metode PCR (polymerase chain reaction).
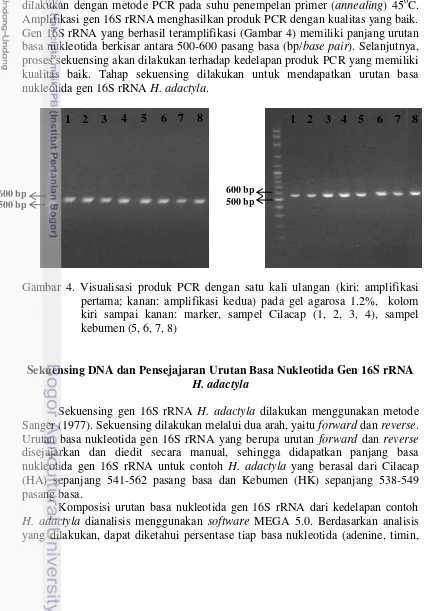
Amplifikasi gen 16S rRNA dari kedelapan DNA total H. adactyla dilakukan dengan metode PCR pada suhu penempelan primer (annealing) 45oC. Amplifikasi gen 16S rRNA menghasilkan produk PCR dengan kualitas yang baik. Gen 16S rRNA yang berhasil teramplifikasi (Gambar 4) memiliki panjang urutan basa nukleotida berkisar antara 500-600 pasang basa (bp/base pair). Selanjutnya, proses sekuensing akan dilakukan terhadap kedelapan produk PCR yang memiliki kualitas baik. Tahap sekuensing dilakukan untuk mendapatkan urutan basa nukleotida gen 16S rRNA H. adactyla.
Gambar 4. Visualisasi produk PCR dengan satu kali ulangan (kiri: amplifikasi pertama; kanan: amplifikasi kedua) pada gel agarosa 1.2%, kolom kiri sampai kanan: marker, sampel Cilacap (1, 2, 3, 4), sampel kebumen (5, 6, 7, 8)
Sekuensing DNA dan Pensejajaran Urutan Basa Nukleotida Gen 16S rRNA
H. adactyla
Sekuensing gen 16S rRNA H. adactyla dilakukan menggunakan metode Sanger (1977). Sekuensing dilakukan melalui dua arah, yaitu forward dan reverse. Urutan basa nukleotida gen 16S rRNA yang berupa urutan forward dan reverse disejajarkan dan diedit secara manual, sehingga didapatkan panjang basa nukleotida gen 16S rRNA untuk contoh H. adactyla yang berasal dari Cilacap (HA) sepanjang 541-562 pasang basa dan Kebumen (HK) sepanjang 538-549 pasang basa.
Komposisi urutan basa nukleotida gen 16S rRNA dari kedelapan contoh H. adactyla dianalisis menggunakan software MEGA 5.0. Berdasarkan analisis yang dilakukan, dapat diketahui persentase tiap basa nukleotida (adenine, timin,
600 bp 500 bp
600 bp 500 bp
15 sitosin, dan guanin) yang menyusun gen 16S rRNA dari kedelapan contoh. Komposisi basa nukleotida gen 16S rRNA dari H. adactyla yang berasal dari Cilacap sampel-1 (C1) terdiri dari 34.9% basa timin (T), 12.4% basa sitosin (C), 35.8% basa adenin (A), dan 16.8% basa guanin (G) dengan panjang sekuen 558 bp. Untuk sampel-2 (C2) terdiri dari 35.2% basa T, 12.6% basa C, 35.2% basa A, dan 16.9% basa G dengan panjang sekuen 562 bp. Untuk sampel-3 (C3) terdiri dari 35.5% basa T, 12.2% basa C, 35.7% basa A, 16.6% basa G dengan panjang sekuen 541 bp. Untuk sampel-4 (C4) terdiri dari 35.4% basa T, 13% basa C, 35.2 basa A, 16.4% basa G dengan panjang sekuen 560 bp. Komposisi nukleotida H. adactyla yang berasal dari Kebumen sampel-1 (K1) terdiri dari 34.8% basa timin (T), 12.2% basa sitosin (C), 36.1% basa adenin (A), dan 16.9% basa guanin (G) dengan panjang sekuen 549 bp. Untuk sampel-2 (K2) terdiri dari 35.5% basa T, 12.2% basa C, 35.7% basa A, dan 16.6 basa G dengan panjang sekuen 541 bp. Untuk sampel-3 (K3) terdiri dari 35.5% basa T, 12.1% basa C, 35.9% basa A, dan 16.7% basa G dengan panjang sekuen 538 bp. Untuk sampel-4 (K4) terdiri dari 35.5% basa T, 12.2% basa C, 35.7% basa A, dan 16.6% basa G dengan panjang sekuen 541 bp (Lampiran 1). Berdasarkan komposisi basa nukleotida, diketahui bahwa basa nukleotida adenine (A) dan timin (T) mendominasi urutan basa nukleotida gen 16S rRNA dari kedelapan sampel, sehingga gen 16S rRNA dari kedelapan contoh dikategorikan sebagai kelompok kaya A-T (A-T rich).
Urutan basa nukleotida gen 16S rRNA dari kedelapan contoh H. adactyla (genus Hippa) disejajarkan dengan H. adactyla yang berasal dari luar Indonesia yang diperoleh dari GenBank (KF051307.1) dengan panjang sekuen 481 bp, serta outgroup (genus Emerita) yaitu Emerita talpoida (KF182557.1) yang diperoleh dari GenBank dengan panjang sekuen 541 bp. Berdasarkan hasil pensejajaran, diperoleh H. adactyla Cilacap dan Kebumen dibandingkan dengan H. adactyla yang diperoleh dari GenBank didapatkan nilai conserved sebesar 89.7% (479/534), variable sebesar 10.1% (54/534) dan singleton sebesar 10.1% (54/534). Berdasarkan pensejajaran kedelapan sampel H. adactyla (Cilacap dan Kebumen) dengan H. adactyla yang berasal dari luar Indonesia serta genus Emerita (E. talpoida) yang diperoleh dari GenBank, didapatkan nilai conserved sebesar 83.5% (446/534), variable sebesar 16.7% (89/534) dan singleton sebesar 4.5% (24/534).
Analisis Jarak Genetik dan Filogeni Gen 16S rRNA H. adactyla
16
dibandingkan antara H. adactyla (Cilacap dan Kebumen) dengan spesies outgroup dari spesies Emeritatalpoida (Tabel 5).
Tabel 5. Jarak genetik fragmen gen 16S rRNA Hippa adactyla yang berasal dari Cilacap (C1, C2, C3, dan C4) dan Kebumen (K1, K2, K3, dan K4), serta H. adactyla dan Emeritatalpoida yang diperoleh dari GenBank (*).
Spesies 1 2 3 4 5 6 7 8 9
1. H. adactyla (C1)
2. H. adactyla (C2) 0.000
3. H. adactyla (C3) 0.000 0.000
4. H. adactyla (C4) 0.000 0.000 0.000
5. H. adactyla (K1) 0.000 0.000 0.000 0.000
6. H. adactyla (K2) 0.000 0.000 0.000 0.000 0.000
7. H. adactyla (K3) 0.000 0.000 0.000 0.000 0.000 0.000
8. H. adactyla (K4) 0.000 0.000 0.000 0.000 0.000 0.000 0.000
9. H. adactyla * 0.093 0.093 0.093 0.093 0.093 0.093 0.093 0.093
10. E. talpoida * 0.110 0.110 0.110 0.110 0.110 0.110 0.110 0.110 0.127
Data jarak genetik dijadikan sebagai dasar dalam analisis filogeni. Analisis filogeni dilakukan untuk mengetahui hubungan kekerabatan antar spesies. Analisis filogeni (Gambar 5) menunjukkan pemisahan yang nyata antara genus Hippa (H. adactyla asal Cilacap, Kebumen, dan yang berasal dari luar Indonesia) dengan genus Emerita (E. talpoida). Pohon filogeni yang dikonstruksi berdasarkan metode pairwise-distance atau p-distance menghasilkan dua nodus. Nodus pertama terdiri dari genus Hippa dan nodus kedua terdiri dari genus Emerita.
17
Situs Mutasi Gen 16S rRNA H. adactyla
Berdasarkan hasil pensejajaran urutan basa nukleotida gen 16S rRNA H. adactyla yang berasal dari Cilacap dan Kebumen dibandingkan dengan H. adactyla yang berasal dari luar Indonesia diperoleh situs mutasi pada gen 16S rRNA H. adactyla sebanyak 54 situs. Situs mutasi yang ditemukan terdiri dari 3 situs mutasi insersi, 2 situs mutasi delesi, 29 situs transisi, dan 20 situs transversi (Lampiran 2).
Nukleotida Spesifik Gen 16S rRNA Hippa adactyla
Berdasarkan hasil pensejajaran gen 16S rRNA H. adactyla dengan gen 16S rRNA H. adactyla yang berasal dari luar Indonesia serta Emerita talpoida, diperoleh 26 situs spesifik pada H. adactyla (Lampiran 3). Situs spesifik ini merupakan penciri yang membedakan spesies H. adactyla Cilacap dan Kebumen dengan spesies H. adactyla yang berasal dari luar Indonesia dan juga genus Emerita. Keberadaan situs spesifik ini menunjukkan adanya evolusi dari spesies H. adactyla.
Konektivitas
18
tersebut (Gambar 6), sehingga konektivitas antara populasi H. adactyla Cilacap dan Kebumen salah satunya dipengaruhi oleh percampuran arus di kedua daerah tersebut.
(a)
(b)
Gambar 6. Pola arus Pulau Jawa berdasarkan olahan data real time Ocean Surface Current Analyses NOAA pada tahun 2014 (a) dan pada Bulan Agustus 2014 (b).
Konservasi
Berdasarkan sekuen H. adactyla asal Cilacap dan Kebumen memiliki nukleotida yang sama. Hal ini menunjukkan tidak adanya keragaman nukleotida antara H. adactyla Cilacap dan Kebumen. Isu utama dalam konservasi genetik salah satunya adalah hilangnya keragaman genetik dari suatu biota, dimana hal ini terjadi pada H. adactyla di Cilacap dan Kebumen. Konsekuensinya adalah kerentanan yang tinggi dalam menghadapi perubahan tekanan lingkungannya. Kegiatan manusia secara langsung dan tidak langsung merupakan salah satu tekanan lingkungan yang mengancam keberlangsungan hidup suatu biota perairan. Adanya seleksi alam sangat mungkin terjadi, hal ini memungkinkan bisa saja sekuen nukleotida H. adactyla Cilacap dan Kebumen merupakan sekuen nukleotida yang telah terseleksi alam dengan baik, sehingga sekuen ini yang mampu bertahan dan mampu merespon perubahan lingkungan yang terjadi dengan baik. Meskipun demikian, pada masa yang akan datang hal ini dapat menimbulkan kerentanan kepunahan jika sekuen nukleotida tersebut tidak dapat beradaptasi dengan perubahan yang terjadi di masa yang akan datang.
19 perlindungan (conserved) dengan membuat aturan yang memperhatikan aspek sustainability, seperti pengaturan maksimal penangkapan dan ukuran yang boleh ditangkap. Jika ketiga pilar ini dijalankan dengan baik, maka H. adactyla di Cilacap dan Kebumen dapat lestari.
20
4 KESIMPULAN DAN SARAN
Kesimpulan
Pendekatan analisis diskriminan menggunakan data rasio morfometrik menunjukkan pusat sebaran yang berbeda antara H. adactyla Cilacap dan Kebumen. Terdapat delapan karakter rasio morfometrik yang dapat membedakan H. adactyla Cilacap dan Kebumen. Penggunaan marka gen 16S rRNA pada spesies yang diteliti menunjukkan memang benar spesies yang diteliti adalah H. adactyla. Terdapat 54 situs mutasi pada gen 16S rRNA antara H. adactyla asal Cilacap dan Kebumen dibandingkan dengan H. adactyla (KF051307.1) yang berasal dari luar Indonesia, terdiri dari 3 situs mutasi insersi, 2 situs mutasi delesi, 29 situs transisi, dan 20 situs transversi. Terdapat 26 situs spesifik pada H. adactyla asal Cilacap dan Kebumen jika dibandingkan dengan H. adactyla yang berasal dari luar Indonesia serta Emerita talpoida (KF182557.1) yang diperoleh dari GenBank. Tidak adanya jarak genetik antara spesies H. adactyla asal Cilacap dan Kebumen mengindikasikan antara H. adactyla asal Cilacap dan Kebumen diperkirakan merupakan satu populasi. Pola arus di perairan Cilacap dan Kebumen memiliki peran yang sangat penting dalam hal konektivitas H. adactyla asal Cilacap dan Kebumen.
Saran
21
DAFTAR PUSTAKA
Amend M, Shanks A. 1999. Timing of Larval Release in the Mole Crab Emerita talpoida. Marine Ecology Progress Series 183: 295-300.
An H-S, Jee Y-J, Min K-S, Kim B-L, Han S-J. 2005. Phylogenetic Analysis of Six Species of Pacific Abalone (Haliotidae) Based on DNA Sequences of 16S rRNA and Cytochrome c Oxidase Subunit I Mitochondrial Genes. Marine Biotechnology 7: 373-380.
Bickford D. Lohman DJ. Sodhi NS. Ng PKL. Meier R. Winker K. Ingram KK. Das I. Cryptic Species as a Window on Diversity and Conservation. Ecology and Evolution 22: 148-155.
Boere V, Cansi ER, Alvarenga ABB, Silva IO. 2011. The Burying Behavior of the Mole Crab Before and After an Accident with Urban Sewage Effluents in Bombinhas Beach, Santa Catarina, Brazil. Ambi-Agua, Taubaté 6: 70-76. Boonruang P, Phasuk B. 1975. Species Compotition and Abundance Distribution
of Anomuran Sand Crab and Population Bionomic of Emerita emeritus (L) Along the Indian Ocean Coast of Thailand (Decapoda: Hippidae). Phuket Marine Biological Center. Thailand. Research Bulletin 8: 1-17.
Bors EK, Rowden AA, Maas EW, Clark MR, Shank TM. 2012. Pattern of Deep-Sea Genetic Connectivity in the New Zealand Region: Implications for Management of Bentic Ecosystems. Plos One 7(11): 1-16.
Boyko CB. Harvey AW. 1999. Crustacea Decapoda: Albuneidae and Hippidae of the Tropical IndoWest Pacific Region. Résultats des campagnes MUSORSTOM 20: 379-406
Brown WM, George M Jr, Wilson AC. 1979. Rapid Evolution of Animal Mitochondrial DNA. Proceedings of the National Academy of Sciences of the United States of America 76: 1967-1971.
Chan T-Y, Osawa M, Boyko CB, Ahyong ST, Macpherson E. 2010. Crustacean Fauna of Taiwan: Crab-Like Anomurans (Hippoidea, Lithodoidea and Porcellanidae). Keelung (Taiwan): National Taiwan Ocean University. 106 p.
Da Silva JM, Dos Santos A, Cunha MR, Costa FO, Creer S, Carvalho GR. 2011. Multigene Molecular Systematics Confirm Species Status of Morphologically Convergent Pagurus Hermit Crabs. Plos One 6(12): 1-10.
Defeo O, Gomez J, Lercari D. 2001. Testing the Swash Exclusion Hypothesis in Sandy Beach Populations: the Mole Crab Emerita brasiliensis in Uruguay. Marine Ecology Progress Series 212: 159-170.
Devine SP, Pelletreau KN, Rumpho ME. 2012. 16S rDNA-Based Metagenomic Analysis of Bacterial Diversity Associated With Two Populations of the Kleptoplastic Sea Slug Elysia chlorotica and Its Algal Prey Vaucheria litorea. Biol Bull 223: 138-154.
Dugan JE, Hubbard DM. 1996. Local Variation in Populations of the Sand Crab Emerita analoga on Sandy Beaches in Southern California. Revista Chilena de Historia Natural 69: 579-588.
22
Frankham R. 2003. Genetics and Conservation Biology. Comptes Rendus Biologies. Elsevier 326: S22-S29.
Goffredi SK, Gregory A, Jones WJ, Morella NM, Sakamoto RI. 2014. Ontogenetic Variation in Epibiont Community Structure in the Deep-Sea Yeti Crab, Kiwa puravida: Convergence Among Crustaceans. Molecular Ecology 23: 1457-1472.
Haye PA, Tam YK, Kornfield I. 2002. Molecular Phylogenetics of Mole Crabs (Hippidae: Emerita). Journal of Crustacean Biology 22: 903-915.
Lercari D, Defeo O. 1999. Effect of Freshwater Discharge in Sandy Beach Populations: the Mole Crab Emerita brasiliensis in Uruguay. Estuarine, Coastal and Shelf Science 49: 457-468.
Mashar A, Wardiatno Y. 2013. Aspek Pertumbuhan Undur-Undur Laut, Hippa adactyla dari Pantai Berpasir Kabupaten Kebumen. Jurnal Biologi Tropis 2(13): 119-127.
Megawati E. 2012. Studi Beberapa Aspek Biologi Undur-undur laut di Kecamatan Buluspesantren Kabupaten Kebumen [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Miya M, Nishida M. 2015. The Mitogenomic Contributions to Molecular Phylogenetics and Evolution of Fishes: a 15-Year Retrospect. Ichthyol Res 62: 29-71.
Mursyidin DH. 2007. Kandungan Asam Lemak Omega 6 Pada Ketam Pasir (Emerita spp.) di Pantai Selatan Yogyakarta. Bioscientiae 4: 79-84.
Nei M, Kumar S. 2000. Molecular Evolution and Phylogenetics. New York (US): Oxford University Pr.
Palumbi SR, Cipriano F. 1998. Species Identification Using Genetic Tools: The Value of Nuclear and Mitochondria Gene Sequences in Whale Conservation. J Hered 89: 459-464.
Pfeiler E, Hurtado LA, Knowles LL, Torre-Cosio J, Bourillon-Moreno L, Marquez-Farias JF, Montemayor-Lopez G. 2005. Population Genetics of the Swimming Crab Callinectes bellicosus (Brachyura: Portunidae) from the Eastern Pacific Ocean. Marine Biology 146: 559-569.
Pinheiro MAA, Fransozo A. 1999. Reproduction of the Speckled Swimming Arenaeus cribrarius (Brachyura: Portunidae) on the Brazilian Coast Near 23°30’S. Journal of Crustacean Biology 22: 416-428.
Powell CL, Ferdin ME, Busman M, Kvitek RG, Doucette GJ. 2002. Development of a Protocol for Determination of Domoic Acid in the Sand Crab (Emerita analoga): A Possible New Indicator Species. Toxicon 40: 485-492.
Quan J, Zhuang Z, Deng J, Dai J, Zhang Y-P. 2004. Phylogenetic Relationships of 12 Penaeoidea Shrimp Species Deduced from Mitochondrial DNA Sequences. Biochemical Genetics 42: 331-345.
Schreiner JN. 2004. Adaptations by the Locomotor Systems of Terrestial and Amphibious Crabs Walking Freely on Land and Underwater [tesis]. United States of America (USA): Louisiana State University.
23 Shih H-T, Ng PKL, Liu M-Y. 2013. Systematics of the Indo-West Pacific
Broad-Fronted Fiddler Crabs (Crustacea: Ocypodidae: Genus Uca). The Raffles Bulletin of Zoology 61(2): 641-649.
Shih H-T, Saher NU, Kamrani E, Ng PKL, Lai Y-C, Liu M-Y. 2015. Population Genetics of the Fiddler Crab Uca sindensis (Alcock, 1900) (Crustacea: Brachyura: Ocypodidae) from the Arabian Sea. Zoological Studies 54: 1-10.
Sleinis S, Silvey GE. 1980. Locomotion in a Forward Walking Crab. Journal of Comparative Physiology 136: 301-312.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. Mega 5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol Biol Evol. 28(10): 2731-2739.
Veas R, Hernandez-Miranda E, Quinones RA. 2014. Body Shape and Burial Behavior of the Sand Crab Emerita analoga (Stimpson 1857) in a Reflective to Intermediate Morphodynamic Range of Sandy Beaches. Marine Biology 161: 2345-2357.
Vidal-Gadea AG, Rinehart MD, Belanger JH. 2008. Skeletal Adaptations for Forwards and Sideways Walking in Three Species of Decapod Crustaceans. Arthropod Structure & Development 37: 95-108.
Walpole RE. 1992. Pengantar Statistika. Jakarta: Gramedia Pustaka Utama. 515p. Wardiatno Y, Ardika PU, Farajallah A, Butet NA, Mashar A, Kamal MM, Renjaan EA, Sarong MA. 2015. Biodiversity of Indonesian Sand Crabs (Crustacea, Anomura, Hippidae) and Assessment of Their Phylogenetic Relationship. AACL Bioflux 2(8): 224-235.
24
LAMPIRAN
25
Lampiran 3. Situs Nukleotida Spesifik gen 16S rRNA Hippa adactyla yang berasal dari Cilacap dan Kebumen.
26