PENGARUH PENAMBAHAN UREA PADA MEDIA BAGAS TERHADAP PRODUKSI DAN KARAKTERISASI ENZIM SELULASE ISOLAT
Aspergillusspp. 1
Oleh
RATNA JAYA INDAH
Proses pengolahan tebu menjadi gula menghasilkan limbah padat yang disebut bagas. Bagas memiliki kandungan selulosa sebesar 50-55% (w/w) sehingga bagas dapat digunakan sebagai substrat untuk produksi enzim selulase. Unsur-unsur pembentuk enzim yaitu unsur karbon, hidrogen dan nitrogen. Senyawa nitrogen merupakan unsur kunci dalam asam amino dan ini menjadikan nitrogen penting untuk proses sintesis protein enzim. Asupan nitrogen akan meningkatkan sintesis protein, sehingga produksi enzim yang dihasilkan juga besar. Namun kandungan protein dalam bagas sangat kecil yaitu 1,6% (w/w) sehingga produksi enzim selulase sedikit, untuk itu perlu adanya penambahan nitrogen. Nitrogen yang ditambahkan dapat berupa urea (CO(NH2)2). Penelitian ini bertujuan untuk mengetahui pengaruh penambahan urea pada media bagas terhadap produksi dan karakterisasi enzim selulase dari isolatAspergillusspp.1.
Penelitian ini dilaksanakan di Laboratorium Mikrobiologi Jurusan Biologi Universitas Lampung dari bulan Juni 2011 sampai dengan Januari 2012. Rancangan percobaan yang digunakan pada penelitian yaitu Rancangan Acak Lengkap (RAL) dengan 3 ulangan. Parameter yang diamati yaitu aktivitas enzim selulase dari masing-masing konsentrasi urea serta karakterisasi enzim selulase meliputi pH, suhu, dan termostabilitas. Kadar konsentrasi urea yang digunakan yaitu 0% (w/v); 0,03% (w/v); 0,06% (w/v); 0,09% (w/v); dan 0,12% (w/v). Enzim selulase yang dihasilkan oleh isolatAspergillusspp.1 mempunyai aktivitas
tertinggi pada konsentrasi urea 0,06% yaitu sebesar 0,07 U/ml. Hasil karakterisasi enzim yang dihasilkan oleh isolatAspergillusspp.1 memiliki aktivitas optimum pada pH 4 yaitu sebesar 0,44 U/ml, suhu optimum pada 400C dengan aktivitas enzim selulase sebesar 0,25 U/ml, serta relatif stabil pada suhu 400C dan 500C.
A. Tinjauan Umum Fungi
Fungi merupakan organisme eukoriotik, berbentuk hifa atau sel tunggal, tidak
berklorofil dan memiliki siklus reproduksi seksual dan aseksual (Gandjar dkk,
1999). Sebagai organisme eukariotik fungi memilki nukleus yang jelas dan
sitoplasma yang dikelilingi oleh membran. Fungi mempunyai dinding sel
yang sedikit selulosa tetapi mengandung banyak kitin dan polisakarida
lainnya (Paul dan Clark, 1996).
Fungi memperoleh zat organik dari lingkungan melalui hifa dan miselium
untuk mendapatkan makanan kemudian menyimpan dalam bentuk glikogen.
Keberlangsungan hidup fungi bergantung pada substrat yang banyak
mengandung karbohidrat, protein, vitamin, dan senyawa kimia lainnya yang
diperoleh dari lingkungan. Zat organik dari sisa makhluk hidup yang telah
mati, misalnya kayu tumbang atau buah jatuh dimanfaatkan oleh fungi
pelapuk yang merupakan parasit saprofit. Fungi saprofit mampu
mengeluarkan enzim hidrolase untuk mendekomposisi molekul kompleks
hifa juga dapat menyerap secara langsung bahan-bahan organik dalam
bentuk sederhana yang dikeluarkan oleh inangnya (Deacon, 1997).
Untuk sumber energi dan sintesis sel, fungi membutuhkan nutrien organik.
Nutrien yang digunakan untuk pertumbuhan fungi adalah senyawa organik
seperti glukosa, asam-asam organik, disakarida, polisakarida, pektin, selulosa,
dan lignin (Alexander, 1997). Fungi hanya mampu mengabsorpsi nutrien
terlarut yang berukuran kecil seperti monosakarida dan asam amino. Jika
nutrien yang tersedia dalam bentuk disakarida maupun polisakarida, maka
substrat didegradasi terlebih dahulu oleh fungi menjadi monosakarida dengan
mengeluarkan enzim ekstraseluler untuk melakukan proses depolimerisasi
yaitu pemecahan senyawa polimer kompleks menjadi senyawa sederhana
(Campbell, Reece, dan Mitchel. 2000).
B. Aspergillus
Aspergillusmempunyai hifa bersepta dan dilengkapi dengan spora aseksual. Ciri lain yang dimiliki olehAspergillusyaitu terdiri darifoot cell, konidiofor, vesikel, sterigma, serta konidia. Karakteristik yang membedakan antara
spesies yang satu dengan spesies yang lain adalah jumlah lapisan sterigma dan
kedudukannya pada vesikel (Dwidjoseputro, 1978). Menurut Frazier &
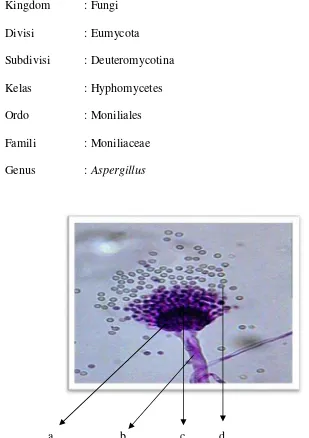
Kingdom : Fungi
Divisi : Eumycota
Subdivisi : Deuteromycotina
Kelas : Hyphomycetes
Ordo : Moniliales
Famili : Moniliaceae
Genus :Aspergillus
a b c d
Gambar 1.Morfologi talusAspergillussp. dengan perbesaran sedang (10x45). Keterangan: (a)Vesikula (b) Konidiofor (c) Sterigmata (d) Konidia
(Pertiwi, 2012)
Morfologi isolatAspergillusspp.1 yaitu koloni berwarna putih terdiri dari kumpulan hifa setelah diinkubasi selama satu hari pada media PDA,
berbentuk bundar dengan tepian menyebar, tepian seperti wol, dengan
dengan bentuk konsentris, tepian bercabang dengan elevasi datar sedangkan
ciri-ciri mikroskopisAspergillusspp.1 yaitu hifa bersepta, berwarna cokelat muda, dan berbentuk spiral. Spora aseksual berbentuk konidia yang
menjuntai. Panjang juntaian 10 atau lebih konidia. Konidia terbentuk di
atas vesikula. Konidia berbentuk bulat, dan berwarna cokelat. Ukuran 5
µm. konidia di produksi berantai dan bercabang, anamorf. Konidiofor
pembentukannya tunggal dan sederhana (Arivo,2010).
C. Sintesis Enzim
Proses sintesis enzim terbagi menjadi tiga tahap, yaitu sintesis asam amino,
sintesis protein, dan sintesis enzim.
1. Sintesis asam amino
Sintesis asam amino merupakan reaksi aminasi (pengikatan gugus amin)
karboksilat. Gugus amin biasanya berasal dari amonia (Purwoko, 2007).
Amonia dapat diperoleh dari reaksi pemecahan urea dan air dengan reaksi
sebagai berikut:
Urea + Air Enzim 2NH3+ CO2
Enzim yang bekerja yaitu enzim urease (urea amidihidrolase). Kemudian
NH3masuk dalam proses aminasi untuk menghasilkan asam amino. Berikut
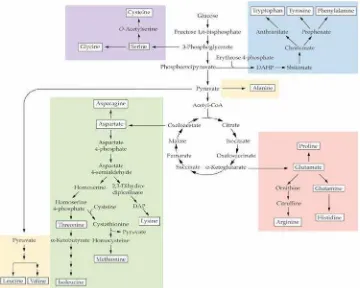
Gambar 2. Reaksi sintesis asam amino (Syafitri, 2012)
Berdasarkan gambar 2, terdapat lima asam amino yang merupakan prekursor
dalam biosintesis asam amino, yaitu glutamat, fenilalanin, aspartat, serin, dan
treonin. Pengelompokkan biosintesis asam amino berdasarkan prekusor
metaboliknya dibagi menjadi 6, yaitu prekusor dariαketoglutarat 3-fosfogliserat,
oksaloasetat, piruvat, fosfoenolpiruvat dan erythrose 4 fosfat, dan ribosa 5 fosfat.
Glutamat merupakan salah satu asam amino yang berperan penting dalam reaksi
pembentukan asam-asam amino lainnya. Glutamat dibentuk dari ammonia dan
α-ketoglutarat, suatu senyawa antara siklus asam sitrat, melalui kerja L-glutamat
dehidrogenase (GDH). α-ketoglutarat dan ammonia membentuk glutamat dengan
NH4++-ketoglutarat + NADPH ===== Glutamat + NADP + H2O
Reaksi ini merupakan dasar dalam biosintesis asam amino karena glutamat
merupakan donor gugus amino dalam biosintesis asam amino yang lain melalui
reaksi transaminasi. Sedangkan glutamin dibentuk dari kerja enzim glutamin
sintesis. Glutamat sintase merupakan enzim yang bereaksi pada reaksi yang
irreversible (tidak balik), namun glutamat dehydrogenase berperan dalam reaksi
yang dapat balik (reversible). Glutamin dibentuk langsung dari glutamat dan
ammonia, energi untuk sintesis ini didapatkan dari adenosine tri phosphate (ATP)
(Webster, 1952). Aktivitas glutamat sintetase berlokasi di sitoplasma (Forde dan
Lea, 2007).
Prolin disintesis dari glutamat atau ornitin. Prolin disintesis dari glutamat melalui
reaksi bertahap. Sebelumnyaglutamat direduksi menjadi α-semialdehida dengan
bantuan glutamatkinase dehidrogenase. Kemudian metabolit ini mengalami penutupan menjadi pirolin 5-karboksilat dan reduksi lebih lanjut menjadi prolin
dengan bantuan enzimpirolin karboksilat reduktase. Prolin adalah penghambat alosterik pada reaksi awal biosintesisnya. Langkah utama dari biosintesis prolin
yaitu dari katalisis glutamat menggunakan dua enzim, yaituΔ
1-pyrroline-5-karboksilat sintetase (P5CS) yang menghasilkanγ-glutamil kinase (γ –GK) dan
asam glutamat semialdehid (GSA) dehidrogenase (γ-glutamil fosfat reduktase).
GSA yang dihasilkan akan dikonversi menjadi prolin-5-karboksilat (P5C) yang
nantinya akan direduksi dengan P5C reduktase (P5CR) menjadi prolin (Zhang,
1995 dalam Raggio dan Raggio, 2007). Selain dari glutamat, prolin juga dibentuk
Alanin berasal dari piruvat dan oksaloasetat melalui transaminasi dari glutamat
(Lehninger, 1982). Seperti halnya glutamat, glutamin, dan prolin, alanin juga
berasal dari metabolit sentral yang didapatkan melalui kerja enzim alanin
transaminase.
Biosintesis aspartat seperti halnya glutamat, aspartat ini disintesis dengan satu
langkah sederhana melalui reaksi transaminasi dibantu dengan kerja enzim
pengkatalisis, yaitu aspartat aminotransferase. Reaksi ini menggunakan analog
asamα-keto aspartat, oksaloasetat, dan glutamat sebagai donor amino. Aspartat
juga diturunkan dari asparagin dengan bantuan asparaginase. Sedangkan
pembentukan asam amino asparagin berasal langsung dari prekursornya yaitu
aspartat dengan dikatalisis oleh asparagin sintetase.
Sintesis asam amino dari kelompok serin-glisin lebih sederhana. 3 fosfogliserat
mengalami dehidrogenasi (oksidasi) menjadi fosfohidroksi fosfat. Reaksi tersebut
dikatalisis oleh fosfogliserat dehidrogenase. Aminasi fosfogliserat dehidrogenase
menjadi fosfoserin. Fosfoserin mengalami hidrasi menjadi serin. Serin dapat
langsung didemetilasi menjadi glisin. Reaksi tersebut dikatalisis oleh serin
hidroksimetiltransferase. Serin juga dapat diasetilasi menjadi asetil serin
kemudian asetil serin mengalami sulfurasi menghasilkan sistein.
Asam amino yang lain seperti fenilalanin, tirosin, dan triptofan disintesis pertama
kali dari kondensasi fosfoeneol piruvat dan eritrosa 4 fosfat. Hasilnya berupa
mengalami reduksi menjadi shikimat, lalu shikimat diubah menjadi krosimat.
Krosimat mengalami mutasi menjadi prefenat, kemudian prefenat mengalami
dekarboksilasi menjadi hidroksi fenilpiruvat selanjutnya diubah menjadi tirosin.
Fenil piruvat diubah menjadi fenilalanin oleh tirosin aminotransferase. Krosimat
juga dapat mengalami transaminasi menjadi antranilat. Antranilat menerima
transfer gugus fosforibosa sehingga menjadi fosforibosil antranilat. Gugus
fosforibosil mengalami desiklisasi sehingga menjadi CDRP (karboksifenilamino
deoksiribosa 5 fosfat). Siklisasi pada gugus aminoribosil sehingga menjadi
indogliserol fosfat. Transaminasi indogliserol fosfat menjadi triptofan oleh
triptofan sintase (Syafitri, 2012 dan Purwoko, 2007).
Asam amino-asam amino yang dihasilkan dalam proses aminasi masuk ke dalam
sitoplasma, yang selanjutnya akan dibawa oleh tRNA ke ribosom untuk proses
sintesis protein.
Sumber nitrogen lain yang dapat ditambahkan dapat berupa Ammonia (NH4+), Nitrat (NO3-), Cyanamide (CaCN2), Ammonium Chlorida ( NH4Cl), Natrium Nitrat (NaNO3), dan Urea (CO(NH2)2). Urea digunakan sebagai sumber nitrogen karena memiliki kadar nitrogen yang cukup tinggi, mudah diperoleh, dan harga
yang relatif murah.
Urea adalah suatu senyawa organik yang terdiri dari unsur karbon, hidrogen,
oksigen dan nitrogen dengan rumus CON2H4atau CO(NH2)2. Urea
merupakan pupuk buatan yang mengandung unsur hara utama nitrogen,
berbentuk butiran (prill) atau gelintiran (glanular) (Nasih, 1996). Unsur
hara N yang terkandung dalam urea sebesar 46% dengan pengertian setiap
100 kg urea mengandung 46 kg Nitrogen (Palimbani, 2007).
2. Sintesis Protein
Translasi adalah proses penerjemahan kode genetik oleh tRNA ke dalam
urutan asam amino. Translasi menjadi tiga tahap yaitu inisiasi, elongasi,
dan terminasi. Semua tahapan ini memerlukan faktor-faktor protein yang
membantu mRNA, tRNA, dan ribosom selama proses translasi. Inisiasi dan
elongasi rantai polipeptida juga membutuhkan sejumlah energi. Energi ini
disediakan oleh GTP (guanosin triphosphat), suatu molekul yang mirip
dengan ATP.
Inisiasi
Tahap inisiasi terjadi karena adanya tiga komponen yaitu mRNA, sebuah
tRNA yang memuat asam amino pertama dari polipeptida, dan dua sub unit
ribosom. mRNA yang keluar dari nukleus menuju sitoplasma didatangi oleh
ribosom, kemudian mRNA masuk ke dalam “celah” ribosom. Ketika
mRNA masuk ke ribosom, ribosom “membaca” kodon yang masuk.
Pembacaan dilakukan untuk setiap 3 urutan basa hingga selesai seluruhnya.
hanya satu, melainkan beberapa ribosom yang dikenal sebagai polisom
membentuk rangkaian miriptusuk satu, di mana tusuknya adalah “mRNA”
dan daging adalah “ribosomnya”. Dengan demikian, proses pembacaan
kodon dapat berlangsung secara berurutan. Ketika kodon I terbaca ribosom
(misal kodonnya AUG), tRNA yang membawa antikodon UAC dan asam
amino metionin dating, tRNA masuk ke celah ribosom.
Elongasi
Proses pemanjangan polipeptida secara umum mempunyai mekanisme 3
tahapan: pengikatan aminoasil–tRNA pada sisi A yang ada di ribosom,
pemindahan rantai polipeptida yang tumbuh dari tRNA yang ada pada sisi P
ke arah sisi A dengan membentuk ikatan peptide, dan translokasi ribosom
sepanjang mRNA ke posisi kodon selanjutnya yang ada di sisi A.
Di dalam kompleks ribosom, molekul fMet-tRNAfMetmenempati sisi P (peptidil), sisi yang lain pada ribosom, yaitu sisi A (aminoasil), masih
kosong pada saat awal sintesis protein. Berpasangannya triplet kodon
inisiasi (AUG/GUG) pada mRNA dengan antikodon pada
metionil-tRNAfMetdi tapak P menentukan urutan triplet kodon dan aminoasil-tRNAfMetberikutnya yang akan masuk ke tapak A. Pengikatan aminoasil-tRNAfMetberikutnya, misalnya alanil- tRNAala, ke tapak A memerlukan protein-protein elongasi EF-Ts dan EF-Tu. Pembentukan ikatan peptida
suatu enzim yang terikat pada subunit ribosom 50S. Reaksi ini
menghasilkan dipeptida yang terdiri atas f-metionin dan alanin yang terikat
pada tRNAala di tapak A. Langkah berikutnya adalah translokasi, yang
melibatkan (1) perpindahan f-met-ala- tRNAala dari tapak A ke tapak P dan
(2) pergeseran posisi mRNA pada ribosom sepanjang tiga basa sehingga
triplet kodon yang semula berada di tapak A masuk ke tapak P. Dalam
contoh ini triplet kodon yang bergeser dari tapak A ke P tersebut adalah
triplet kodon untuk alanin. Triplet kodon berikutnya, misalnya penyandi
serin, akan masuk ke tapak A dan proses seperti di atas hingga translokasi
akan terulang kembali. Translokasi memerlukan aktivitas faktor elongasi
berupa enzim yang biasa dilambangkan dengan EF-G. Pemanjangan atau
elongasi rantai polipeptida akan terus berlangsung hingga suatu tripet kodon
yang menyandi terminasi memasuki tapak A.
Terminasi
Translasi akan berakhir pada waktu salah satu dari ketiga kodon terminasi
(UAA,UGA,UAG) yang ada pada mRNA mencapai posisi A pada ribosom.
Dimana RF1yang mengenali kodon UAA atau UAG sehingga rantai kodon
tersebut akan terlepas, kemudian RF2akan mengenali kodon UAA atau UGA sehingga rantai kodon tersebut terlepas. Proses terminasi ditandai oleh
terlepasnya mRNA, tRNA di tapak P, dan rantai polipeptida dari ribosom.
Selain itu kedua subunit ribosompun memisah, pada terminasi diperlukan
aktivitas dua protein yang berperan sebagai faktor pelepas atau releasing
Selama proses dan sesudah sintesisnya, suatu rantai polipeptida mulai
menggulung dan melipat secara spontan, membentuk protein fungsional
dengan konformasi yang spesifik: suatu molekul tiga dimensi dengan
struktur sekunder dan struktur tersier. Suatu gen menetukan struktur primer
dan struktur primer ini kemudian akan menentukan konformsi protein
(Campbell, Reece, dan Mitchel. 2000).
3. Sintesis Enzim
Polipeptida yang telah terbentuk akan dibawa menuju retikulum endoplasma
(RE). Rantai polipeptida ini dilengkapi dengan peptida sinyal. Peptida sinyal
merupakan suatu urutan kira-kira 20 asam amino di dekat atau pada ujung
leading(amino) dari polipeptida yang akan dikenali oleh partikel pengenal sinyal (SRP). Partikel ini akan mengikatkan diri pada peptida sinyal.
Selanjutnya SRP mengikatkan diri pada protein reseptor di dalam membran
RE. Reseptor ini merupakan bagian dari kompleks protein yang disebut
kompleks translokasi serta mencakup pori-pori membran dan enzim
pembelahan sinyal. Setelah itu SRP dilepaskan dan polipeptida yang sedang
tumbuh ditranslokasi melintasi membran. Peptida sinyal tetap melekat pada
membran kemudian enzim pembelah sinyal memotong peptida. Sisa dari
polipeptida yang sudah terbentuk sempurna kemudian meninggalkan ribosom
dan membentuk konformasi protein.
Protein-protein tersebut keluar dari RE dibungkus dalam membran vesikula
yang menggelembung dari daerah terspesialisasi yang disebut RE transisi.
vesikula transpor. Setelah meninggalkan RE, vesikula transpor berpindah
ke aparatus golgi. Produk dari aparatus golgi (protein enzim) yang akan
disekresi keluar dari mukatransakan berfusi dengan membran plasma. Selanjutnya produk tersebut menjadi enzim (protein) ekstraseluler
(Campbell, Reece, dan Mitchel. 2000).
D. Enzim Selulase
Enzim merupakan molekul polimer yang beragam yang dihasilkan oleh sel
hidup. Keragamannya dapat dilihat baik pada bentuk, ukuran, maupun
peranannya (Suhartono, 1989). Enzim merupakan protein yang memiliki
spesifikasi serta memiliki aktivitas katalitik. Terhadap substratnya,
spesifisitas enzim sangat tinggi. Enzim mempercepat reaksi kimiawi
spesifik tanpa pembentukan produk samping dan molekul ini berfungsi di
dalam larutan encer pada keadaan suhu dan pH normal (Lehninger, 1982).
Klasifikasi enzim secara internasional berdasarkan reaksi yang dikatalisis
antara lain:
1. Oksidoreduktase, enzim golongan ini dibagi dalam dua bagian yaitu
dehidrogenase dan oksidase. Dehidrogenase bekerja pada reaksi
dehidrogenasi yaitu reaksi pengambilan atom hidrogen dari suatu
senyawa. Sedangkan oksidase bekerja sebagai katalis pada reaksi
pengambilan hidrogen dari suatu substrat.
2. Transferase, enzim golongan ini bekerja sebagai katalis pada reaksi
Beberapa contoh enzim golongan ini yaitu metiltransferase,
hidroksimetiltransferase, karboksiltransferase, asiltransferase dan
aminotransferase.
3. Hidrolase, enzim golongan ini bekerja sebagai katalis pada reaksi
hidrolisis. Beberapa contoh ialah lipase, fosfatase, amilase, pepsin, tripsin
dan kimotripsin.
4. Liase, enzim golongan ini mempunyai peranan penting dalam reaksi
pemisahan suatu gugus dari suatu substrat atau sebaliknya. Contoh enzim
golongan ini yaitu dekarboksilase, aldose dan hidratase.
5. Isomerase, enzim golongan ini bekerja pada reaksi perubahan
intramolekuler, misalnya reaksi perubahan glukosa menjadi fruktosa.
6. Ligase, enzim golongan ini bekerja pada reaksi penggabungan dua
molekul. Contoh enzim golongan ini antara lain glutamin sintetase dan
piruvat karboksilase (Poedjiadi, 1994).
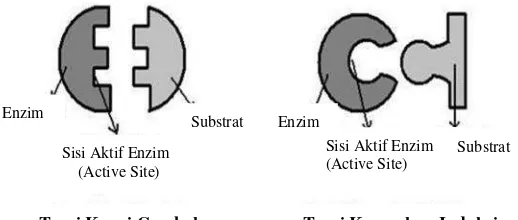
Enzim bekerja dengan dua cara, yaitu menurut Teori Kunci-Gembok
(Lock and Key Theory) dan Teori Kecocokan Induksi (Induced Fit Theory). Mekanisme kerja enzim dapat dilihat pada Gambar 4.
Teori Kunci-Gembok (Lock and Key Theory) dikemukakan oleh Emil Fisher yang menyatakan bahwa kerja enzim seperti kunci dan anak kunci, melalui
hidrolisis senyawa gula dengan enzim invertase. Terjadinya reaksi antara
substrat dengan enzim adalah karena adanya kesesuaian bentuk ruang antara
substrat dengan sisi aktif (active site) dari enzim, sehingga sisi aktif enzim cenderung kaku. Substrat berperan sebagai kunci (key) dan sisi aktif (lock) berperan sebagai gembok. Substrat masuk ke dalam sisi aktif sehingga terjadi
kompleks enzim-substrat. Hubungan antara enzim dan substrat membentuk
ikatan yang lemah. Pada saat ikatan kompleks enzim-substrat terputus,
produk hasil reaksi akan dilepas dan enzim akan kembali pada konfigurasi
semula.
Teori Kecocokan Induksi (Induced Fit Theory) dikemukakan oleh Daniel Koshland yang menyatakan bahwa sisi aktif tidak bersifat kaku tetapi lebih
fleksibel. Sisi aktif secara terus menerus berubah bentuknya sesuai dengan
interaksi antara enzim dan substrat. Ketika substrat memasuki sisi aktif
enzim, bentuk sisi aktif akan termodifikasi menyesuaikan bentuk substrat
sehingga terbentuk kompleks enzim substrat. Sisi aktif akan terus berubah
bentuknya sampai substrat terikat secara sepenuhnya, yang mana bentuk akhir
dan muatan enzim ditentukan. Ketika substrat terikat pada enzim, sisi aktif
enzim mengalami beberapa perubahan sehingga ikatan yang terbentuk antara
enzim dan substrat menjadi menjadi lebih kuat. Interaksi antara enzim dan
Menurut Da silva, Largo, Merheb, Machiome, Park, dan Gomes (2005),
berdasarkan hasil pemeriksaan pada fungi, sistem selulase
sekurang-kurangnya terdiri dari tiga enzim:
1. Enzim-enzim endo-β-1,4-glukanase
2. Enzim ekso-β-1,4-glukanase
3. Enzim-enzim β-glukosidase.
Menurut Salma dan Gunarto (1999), selulase merupakan enzim yang dapat
memutuskan ikatan glukosida β-1,4 didalam selulosa. Dalam menghidrolisis
senyawa selulosa, kemampuan selulase sangat digantungkan pada substrat
yang di gunakan.
E. Karakteristik Enzim
Enzim memiliki karakteristik tertentu yang dapat mempengaruhi laju reaksi
suatu enzim, antara lain:
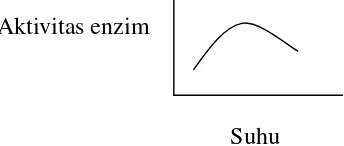
Suhu
Laju reaksi akan meningkat seiring dengan kenaikan suhu sampai batas
tertentu kemudian aktivitas enzim akan mengalami penurunan karena
enzim terdenaturasi oleh suhu yang terlalu tinggi.
Aktivitas enzim
Suhu
pH
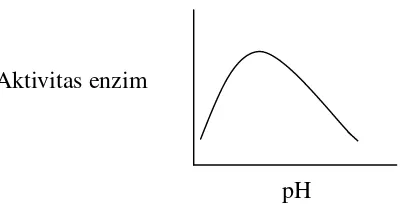
Laju reaksi meningkat pada pH optimum dan aktivitas enzim akan
mengalami penurunan pada kedua sisi pH optimum oleh pH yang terlalu
tinggi atau rendah. Hal ini disebabkan oleh beberapa hal:
Protein enzim dapat mengalami denaturasi akibat pH yang tinggi ataupun yang rendah
Protein enzim memerlukan gugus-gugus asam amino yang terionisasi pada rantai samping yang mungkin aktif hanya pada
satu keadaan ionisasi
Substrat dapat memperoleh atau kehilangan proton dan reaktif dalam satu bentuk muatan
Aktivitas enzim
pH
Gambar 6. Pengaruh pH terhadap aktivitas enzim
Konsentrasi enzim
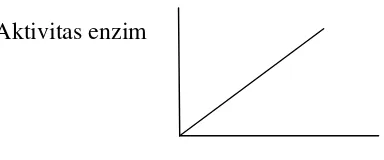
Pengaruh konsentrasi enzim terhadap aktivitas enzim yang dikatalisis oleh
enzim dapat diliihat pada gambar 7, aktivitas enzim meningkat secara
linier dengan bertambahnya konsentrasi enzim selama konsentrasi enzim
Aktivitas enzim
Konsentrasi enzim
Gambar 7. Pengaruh konsentrasi enzim terhadap aktivitas enzim
Konsentrasi substrat
Aktivitas enzim mula-mula meningkat seiring bertambahnya konsentrasi
substrat namun setelah konsentrasi substrat dinaikkan lebih lanjut akan
tercapai suatu laju limit atau laju maksimum. Penambahan konsentrasi
substrat tidak berpengaruh terhadap aktivitas enzim.
Aktivitas enzim
Konsentrasi substrat
Gambar 8. Pengaruh konsentrasi substrat terhadap aktivitas enzim (Page, 1989)
F. Selulosa
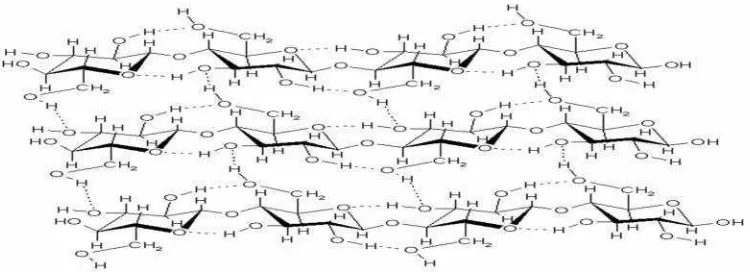
Selulosa (C6H1005)nmerupakan komponen struktural utama dari tumbuhan yang tidak dapat dicerna oleh manusia. Selulosa banyak terdapat pada
tumbuhan berkayu dan berserat, jumlahnya sangat melimpah di alam.
Gambar 9. Struktur Selulosa (Theo, 2007).
menempati hampir 60% komponen penyusun struktur tanaman (Salma dan
Gunarto, 1999). Selulosa merupakan komponen penting yang digunakan
sebagai bahan baku pembuatan kertas dan merupakan polimer linear dengan
berat molekul tinggi yang tersusun seluruhnya atas ß-D-glukosa dan dapat
memenuhi fungsinya sebagai komponen struktur utama dinding sel tumbuhan
karena sifat-sifat kimia dan fisiknya maupun struktur molekulnya ( Fengel
dan Wegener, 1995). Menurut Sjostrom (1981), selulosa merupakan
homopolisakarida yang tersusun atas unit ß-D-glukopironosa yang terikat satu
sama lain dengan ikatan glikosida.
Fungi merupakan mikroorganisme utama yang memiliki kemampuan untuk
menghidrolisis selulosa alami melalui aktivitas enzim selulase yang
dimilikinya (Salma dan Gunarto, 1999). Menurut Irawan (2003) fungi yang
mempunyai kemampuan mencerna selulosa terdapat pada kelompok fungi
yang tergolong ke dalam Ascomycotina dan Basidiomycotina. Fungi
aspergillus nudulans, Botrytis cinerea, Rhizoctonia solani, Trichoderma viridae(Schlegel, 1994).
G. Bagas
Gambar 10. Bagas
Bagas merupakan residu atau hasil sampingan dari proses ekstraksi
(pemerahan) cairan tebu menjadi gula, yang sejauh ini masih belum banyak
dimanfaatkan menjadi produk yang memiliki nilai tambah (Samsuri dkk.,
2006). Pada musim giling tahun 2006, data yang diperoleh dari Ikatan Ahli
Gula Indonesia (IKAGI) menunjukkan bahwa jumlah tebu yang digiling
oleh 57 pabrik gula di Indonesia mencapai sekitar 30 juta ton, sehingga
bagas yang di hasilkan diperkiran mencapai 9.640.000 ton. Namun,
sebanyak 60% dari bagas tersebut dimanfaatkan oleh pabrik gula sebagai
bahan bakar, bahan baku untuk kertas, bahan baku industri kanvas rem,
industri jamur dan lain-lain. Oleh karena itu diperkirakan sebanyak 40 %
dari bagas tersebut belum dimanfaatkan (Husin, 2007). Bagas sebagian
besar mengandunglignocellulose. Panjang seratnya antara 1,7 sampai 2 mm dengan diameter sekitar 20 mikro. Bagas mengandung air 48 - 52%,
dalam air dan sebagian besar terdiri dari selulosa, pentosan dan lignin
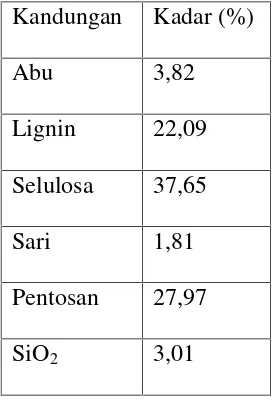
(Husin, 2007). Menurut Husin (2007) hasil analisis serat bagas adalah
seperti dalam Tabel 1 berikut:
Kandungan Kadar (%)
Abu 3,82
Lignin 22,09
Selulosa 37,65
Sari 1,81
Pentosan 27,97
SiO2 3,01
A. Tempat dan Waktu Penelitian
Penelitian ini di laksanakan di Laboratorium Mikrobiologi Jurusan Biologi
FMIPA Universitas Lampung dari bulan Juni 2011 sampai dengan Januari
2012
B. Alat dan Bahan
1. Alat
Alat-alat yang digunakan dalam penelitian ini yaitulaminar air flow cabinet,autoclave, neraca analitik, inkubator, jarum ose, erlenmeyer, kompor listrik,vortex mixer, mikropipet, pipet tips, spektrofotometer, botol selai, kapas, tabung reaksi, aluminium foil, batang pengaduk, botol aquades,
lampu spiritus, orbital shaker, centrifuge, mikrotube, water bath shaker,
2. Bahan
Bahan yang digunakan adalah bagas dari PT. GMP Gunung Sugih Lampung
Tengah, PDA (Potato Dextrose Agar), CMC (Carboxymethyl Cellulose), aquades, garam fisiologis, KH2PO4,FeSO4,buffer sitrat, pospat buffer, tris
buffer, DNS (3,5-dinitrosaliclic acid), urea (CO(NH2)2), alkohol 70%, dan spiritus.
C. Metode Penelitian
Uji pengaruh penambahan urea terhadap produksi dan karakterisasi enzim
selulase pada media bagas menggunakan Rancangan Acak Lengkap (RAL) 3
ulangan. Kadar konsentrasi urea yang digunakan yaitu 0% (w/v); 0,03%
(w/v); 0,06% (w/v); 0,09% (w/v); dan 0,12% (w/v). Parameter yang diamati
yaitu aktivitas enzim selulase dari masing-masing konsentrasi yang ditentukan
bedasarkan kadar glukosa yang terbentuk dari reaksi enzimatis antara ekstrak
enzim selulase dengan CMC (Carboxymetylcelullose) sebagai subsrat.
D. Analisis Data
Data yang diperoleh dilakukan analisis ragam, dan apabila di antara perlakuan
terdapat perbedaan nyata pada taraf kepercayaan 5%, analisis dilanjutkan
E. Produksi Enzim Selulase 1. Penyiapan Suspensi spora
Isolat fungi yang telah diremajakan pada PDA miring diinkubasi selama 3
hari. Pada masing-masing isolat tersebut dimasukkan larutan NaCl steril
dan dilakukan pemisahan antara spora dengan media menggunakanvortex mixer. Kemudian dilakukan pemisahan antara suspensi dengan medianya (PDA) sehingga diperoleh suspensi spora.
2. Penyiapan Media Fermentasi
Media fermentasi didasarkan pada media yang digunakan oleh Ahamed dan
Vermette (2008). Kedalam 1 liter aquades dilarutkan 2,0 g KH2PO4; 0,005
g FeSO4·7H2O. Kemudian dari larutan tersebut diambil sebanyak 200 ml lalu ditambahkan urea sesuai konsentrasi yang di tentukan. Sebanyak 1,35 g
bagas dimasukkan ke dalam botol selai kemudian ditambahkan 45 mL
larutan urea. Campuran tersebut disterilisasi pada temperatur 1210C selama 15 menit.
3. Produksi Enzim Selulase
Suspensi spora diambil sebanyak 1,8 ml lalu diinokulasikan pada media
fermentasi dan diinkubasi pada orbital shaker selama 4 hari (Purwadaria,
2003). Produksi enzim selulase ditentukan berdasarkan aktivitas enzim.
Aktivitas enzim tersebut dapat diketahui dari besarnya kadar glukosa
4. Penentuan Aktivitas Enzim Selulase
Uji aktivitas enzim selulase dilakukan berdasarkan kadar glukosa. Kadar
glukosa diukur dengan menggunakan metode DNS ( Miller, 1959). Isolat
fungi yang telah diinkubasi kemudian dipanen. Sebanyak 1 ml ekstrak
enzim diambil dan dicentrifuge selama 2 menit. Kemudian diambil
sebanyak 0,5 mL dan dicampurkan dengan CMC 0,5% sebanyak 0,5 mL
dan diinkubasi selama 30 menit padawater bathpada suhu 400C. Reaksi terakhir dilakukan dengan menambahkan 1 mL 3,5-dinitrosaliclic acid. Dihomogenkan dan dipanaskan kedalam air mendidih selama 15 menit.
Kemudian didinginkan ke dalam air dingin selama 20 menit selanjutnya
diukur absorbansinya dengan menggunakan spektrofotometer pada panjang
gelombang 540 nm. Absorbansi yang diperoleh digunakan untuk
menentukan kadar gula dengan rumus persamaan regresi linear Y= a + bx.
a dan b diperoleh dari perhitungan gula standar, Y adalah nilai absorbansi
pada panjang gelombang 540 nm, x adalah kadar glukosa yang dihasilkan.
Satu unit dari aktivitas selulase didefinisikan sebagai jumlah dari enzim
yang melepakan µmol glukosa dalam satu menit pada kondisi pengujian.
Penentuan aktivitas enzim selulase per unit dapat di tentukan dengan rumus:
Aktivitas enzim = kadar glukosa x faktor pengenceran
Keterangan :
Faktor pengenceran : x kali
Berat Molekul glukosa : 180
Waktu Inkubasi : 30 menit
F. Karakterisasi Enzim
Setelah diperoleh konsentrasi optimum, maka dilakukan karakterisasi yang
meliputi:
1. Karakterisasi Suhu
Pengujian suhu optimum selulase dilakukan dengan mereaksikan enzim
selulase dengan substrat CMC 0,5% selama 30 menit pada suhu 300C -700C dengan selang suhu 100C. Pengujian dilakukan dengan cara
menginkubasi reaksi enzim-substrat pada suhu 300C, 400C, 500C, 600C dan 700C di waterbathshaker selama 30 menit.
2. Karakterisasi pH
Pengujian pH optimum selulase dilakukan dengan mereaksikan enzim
selulase dengan substrat CMC 0,5% selama 30 menit pada suhu 400C pada berbagai kondisi pH larutan buffer dengan selang pH 1 unit, yaitu
menggunakan buffer sitrat untuk pH 4 dan 5, buffer phospat untuk pH 6
dan 7, buffer tris untuk pH 8 dan 9.
3. Karakterisasi Termostabilitas Enzim
Supernatan enzim diinkubasi pada suhu 400C, 500C, 600C, dan 700C dengan waktu sampling setiap 30 menit selama 3 jam kemudian
G. Pembuatan Kurva Standar Glukosa
Larutan gula reduksi glukosa merupakan larutan gula standar yang
digunakan pada interval 0-300 µg yaitu 0 µg, 50 µg, 100 µg, 150 µg, 200
µg, 250 µg, 250 µg, dan 300 µg, masing-masing larutan diambil sebanyak
0,5 ml, ditambahkan 0,5 ml larutan CMC sebagai substrat dan pereaksi
DNS. Divortex hingga homogen, kemudian dipanaskan dalam air mendidih
selama 15 menit. Selanjutnya didinginkan dengan merendamnya kedalam
air dingin selama 20 menit. Pengukuran absorbansi dilakukan dengan
menggunakan spektrofotometer pada panjang gelombang 540 nm (Miller,
1959).
H. Prosedur Kerja
Uji aktivitas enzim selulase
Uji aktivitas enzim Uji gula pereduksi
Kadar gula Persamaan regresi linear
Data absorbansi
Aktivitas optimum
Karakterisasi enzim
Suhu pH Termostabilitas enzim
A. Simpulan
Berdasarkan hasil penelitian yang telah dilakukan maka dapat disimpulkan
bahwa:
1. Urea berpengaruh terhadap produksi enzim selulase isolat
Aspergillusspp. 1 dengan aktivitas tertinggi pada konsentrasi urea 0,06% (w/v) sebesar 0,07 U/ml, sedangkan aktivitas terendah terdapat pada
konsentrasi urea 0% (w/v) yaitu sebesar 0,01 U/ml.
2. Enzim selulase isolatAspergillusspp. 1 memberikan kondisi optimum aktivitas enzim yaitu pada pH 4, suhu 400C serta aktivitas enzim selulase relatif stabil pada suhu 400C dan 500C.
B. Saran
Berdasarkan hasil penelitian ini dapat disarankan adanya penambahan urea
pada proses fermentasi bagas untuk memperbesar produksi enzim serta
penyesuaian kondisi lingkungan meliputi pH dan suhu. pH 4 dan suhu 400C merupakan kondisi optimum untuk produksi enzim selulase. Selanjutnya
dapat dilakukan penelitian lanjutan untuk mengetahui produksi enzim
(Skripsi)
Oleh :
RATNA JAYA INDAH
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS LAMPUNG
PRODUKSI DAN KARAKTERISASI ENZIM SELULASE ISOLAT Aspergillusspp. 1
Oleh
RATNA JAYA INDAH
Proses pengolahan tebu menjadi gula menghasilkan limbah padat yang disebut bagas. Bagas memiliki kandungan selulosa sebesar 50-55% (w/w) sehingga bagas dapat digunakan sebagai substrat untuk produksi enzim selulase. Unsur-unsur pembentuk enzim yaitu unsur karbon, hidrogen dan nitrogen. Senyawa nitrogen merupakan unsur kunci dalam asam amino dan ini menjadikan nitrogen penting untuk proses sintesis protein enzim. Asupan nitrogen akan meningkatkan sintesis protein, sehingga produksi enzim yang dihasilkan juga besar. Namun kandungan protein dalam bagas sangat kecil yaitu 1,6% (w/w) sehingga produksi enzim selulase sedikit, untuk itu perlu adanya penambahan nitrogen. Nitrogen yang ditambahkan dapat berupa urea (CO(NH2)2). Penelitian ini bertujuan untuk mengetahui pengaruh penambahan urea pada media bagas terhadap produksi dan karakterisasi enzim selulase dari isolatAspergillusspp.1.
Penelitian ini dilaksanakan di Laboratorium Mikrobiologi Jurusan Biologi Universitas Lampung dari bulan Juni 2011 sampai dengan Januari 2012. Rancangan percobaan yang digunakan pada penelitian yaitu Rancangan Acak Lengkap (RAL) dengan 3 ulangan. Parameter yang diamati yaitu aktivitas enzim selulase dari masing-masing konsentrasi urea serta karakterisasi enzim selulase meliputi pH, suhu, dan termostabilitas. Kadar konsentrasi urea yang digunakan yaitu 0% (w/v); 0,03% (w/v); 0,06% (w/v); 0,09% (w/v); dan 0,12% (w/v). Enzim selulase yang dihasilkan oleh isolatAspergillusspp.1 mempunyai aktivitas
tertinggi pada konsentrasi urea 0,06% yaitu sebesar 0,07 U/ml. Hasil karakterisasi enzim yang dihasilkan oleh isolatAspergillusspp.1 memiliki aktivitas optimum pada pH 4 yaitu sebesar 0,44 U/ml, suhu optimum pada 400C dengan aktivitas enzim selulase sebesar 0,25 U/ml, serta relatif stabil pada suhu 400C dan 500C.
Oleh
RATNA JAYA INDAH
Skripsi
Sebagai Salah Satu Syarat untuk Mencapai Gelar SARJANA SAINS
Pada Jurusan Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS LAMPUNG
ISOLATAspergillusspp. 1
Nama Mahasiswa : Ratna Jaya Indah Nomor Pokok Mahasiswa : 0717021062
Jurusan : Biologi
Fakultas : Matematika dan Ilmu Pengetahuan Alam
MENYETUJUI
1. Komisi Pembimbing
Dra. Christina.N Ekowati, M.Si Dr. Sumardi
NIP. 195808181985032001 NIP 196503251991031003
2. Ketua Jurusan Biologi
1. Tim Penguji
Ketua : Dra. Christina N. Ekowati, M. Si ...
Sekretaris : Dr. Sumardi ...
Penguji
Bukan Pembimbing : Kusuma Handayani, M. Si ...
2. Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Prof. Suharso, Ph.D. NIP. 196905301995121001
Penulis dilahirkan di Sukadana, Lampung Timur pada
tanggal 01 Desember 1989, dari pasangan Bapak H.
Sutarman dan Ibu Hj. Tumini, sebagai anak bungsu dari
empat bersaudara. Penulis menyelesaikan pendidikan Taman
Kanak-kanak di TK PGRI Bandar Negeri Lampung Timur
pada tahun 1995. Pendidikan dasar diselesaikan pada tahun 2001 di SDN Kedung
Rejo Jombang, Jawa Timur kemudian melanjutkan Sekolah Lanjutan Tingkat
Pertama di SLTP Negeri Pasir Sakti Lampung Timur sampai tahun 2004, dan
Sekolah Menengah Atas di SMA Negeri 3 Bandar Lampung hingga tahun 2007.
Pada tahun yang sama, penulis terdaftar sebagai mahasiswa di Jurusan Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Lampung melalui
jalur SPMB (Seleksi Penerimaan Mahasiswa Baru).
Selama menjadi mahasiswa, penulis pernah menjadi asisten pada mata kuliah
Struktur Perkembangan Hewan I, Struktur Perkembangan Hewan II, Mikrobiologi
Pangan dan Industri, Mikrobiologi Umum, Mikrobiologi Tanah, dan Fisiologi
Mikroba Jurusan Biologi FMIPA, Mikrobiologi Jurusan Biologi FKIP,
Penulis juga aktif di UKM Rohani Islam tahun 2010 sebagai Ketua Biro
Keputrian, di HIMBIO sebagai sekretaris Biro Danus pada tahun 2009-2010.
Pada tahun 2007 hingga sekarang penulis tergabung di Tim Kerja Dakwah
Sekolah (TKS) SMAN 3 Bandar Lampung dan SMKN 2 Bandar Lampung. Saat
ini, penulis mendapat amanah disebuah yayasan pembinaan pelajar dan generasi
muda, yaitu Forum Kerjasama Alumni Rohis (FKAR) Bandar Lampung sebagai
Staff Kesekretariatan.
Tahun 2010 penulis melaksanakan kerja praktek di PT Neka Boga Perisa, sebuah
perusahaan yang mengekspor rempah-rempah. Penulis berkesempatan untuk
Dengan penuh kerendahan hati, kupersembahkan karya
sederhana ini untuk:
Alloh swt, sebagai salah satu wujud ibadahku
untuk mengharapkan keridhoanNya
Rosululloh Muhammad saw atas rasa cinta
dan rinduku yang tak terkira kepada beliau
Ibu dan Bapak yang mulia.. Bakti ananda
sepanjang hidup pun tak akan mampu
membalas apa yang telah Ibu&Bapak berikan
Saudara kandungku tercinta: Mba Tatik,
Mba Jar, Mas Agus beserta keluarga kecilnya
yang selalu mengukir pelangi kebahagiaan di
hatiku..
Alhamdulillah, rasa syukur yang tiada terhingga kepada Sang Maha Pencipta
karena ananda terlahir dari wanita mulia, keluarga sederhana Ibu&Bapak.
Segudang rasa terima kasih tak akan pernah mampu membalas kebaikan
Ibu&Bapak. Atas semua cinta dan kasih sayang yang sangat kental ananda
rasakan bahkan sampai ananda dewasa, atas nasihat serta petuah yang
diberikan untuk menuntun ananda menjadi orang yang bermanfaat bagi
sesama, atas setiap tetes keringat yang mengalir di tubuh Bapak demi
tercapainya mimpi ananda, atas rasa cemas dan khawatir yang sering hinggap
di hati Ibu karena ananda tinggal di perantauan, atas segalanya yang tak
mampu ananda sebutkan, ananda ucapakanjazaakumulloh khoiron katsir,
semoga Alloh membalas kebaikan Ibu&Bapak dengan pahala yang berlipat,
mendapatkan kemulian di dunia dan akhirat serta memasuki syurga Alloh
tanpa hisab. Izinkan ananda berbakti kepada Ibu&Bapak sepanjang usia
ananda, ananda ingin berbakti hingga tubuh ini lelah, hingga Alloh pun ridho
dengan apa yang telah ananda lakukan...karena syurga Alloh itu sangat
dekat dengan Anda...Maafkan ananda jika bakti ini dirasakan belum lengkap
dan tak sesuai dengan keinginan Ibu&Bapak, ananda hanya ingin
mempersembahkan yang terbaik seperti yang dulu Ibu&Bapak berikan untuk
ananda...
Salam bakti dan cinta sedalam samudra...
Putrimu,
Maka orang-orang yang beriman dan mengerjakan kebajikan , mereka memperoleh ampunan dan rezeki yang mulia
(Terjemahan QS Al Hajj ayat 50)
Alloh tidak membebani seseorang melainkan sesuai dengan kesanggupannya
Alhamdulillah, segala puji hanya milik Alloh swt, Robb semesta alam yang
Maha Menggenggam jiwa-jiwa hambaNya, yang memiliki kerajaan langit
dan bumi hingga tak ada satu pun yang dapat luput dari pengawasanNya.
Sholawat serta salam selalu tercurahkan untuk sosok manusia paling mulia,
Nabi Muhammad saw beserta keluarga, sahabat, serta umat yang senantiasa
mengikuti sunnahnya.
Skripsi ini juga merupakan syarat untuk memperoleh gelar Sarjana Sains di
Universitas Lampung. Penyelesaian skripsi ini melibatkan banyak pihak,
untuk itu penulis mengucapkan terimakasih yang tiada terkira kepada:
1. Ibu Tumini dan Bapak Sutarman tersayang atas limpahan cinta, doa dan
pengorbanan untuk ananda. Semoga Alloh memuliakan Anda dunia dan
akhirat.
2. Ibu Dra. C.N. Ekowati, M.Si, selaku pembimbing I. Terima kasih yang
sedalam-dalamnya atas semua ilmu, nasehat, saran dan kritik produktif
yang telah Ibu berikan, serta atas kesabaran dalam membimbing penulis
hingga skripsi ini terselesaikan.
3. Bapak Dr. Sumardi, selaku pembimbing II yang selalu meluangkan waktu
untuk membimbing penulis, terimakasih atas semua motivasi, saran, serta
pembelajaran yang tak ternilai, semoga Alloh swt membalas kebaikan
Bapak dengan pahala yang berlipat.
4. Ibu Kusuma Handayani, M.Si. selaku pembahas atas kritik, saran serta
kesabarannya dalam membimbing penulis, luapan rasa terimakasih
berbungkus doa, semoga Ibu senantiasa dalam lindunganNya.
5. Ibu Dra. Nuning Nurcahyani, M.Sc. selaku Ketua Jurusan Biologi Fakultas
Matematika dan Ilmu Pengetahuan Alam.
6. Ibu Tundjung Tripeni Handayani, M.S., yang bersedia menjadi “guru”
8. Bapak Drs. M. Kanedi, M.Sc. selaku Pembimbing Akademik.
9. Seluruh karyawan dan laboran Jurusan Biologi (special to: Mba Ida&Nunung, makasih atas keceriaan, canda dan tawa selama ini...)
10. Mamasku tercinta: Agus Harwanto, yang mengajariku banyak hal,
makasih atas cinta yang selalu mengalir tanpa henti...
11. Mba Jarmiati dan Mba Hartatik,my beloved sisters, atas semua kasih sayang dan doa-doa kebaikan yang selalu mengalir untukku.
12. Keponakan tercinta yang menjadi sumber semangat dan penghilang penat:
Eka, Uwi’,Reni, Ambar, Nonik, Yogi, Firda, serta si kecil Aji.
13. Keluarga besarku yang selalu mendoakan dan memberi semangat
tersendiri, semoga kelak Alloh mengumpulkan kita di syurgaNya.
14. Zahwa,gumawo....atas kebersamaan selama ini, suka duka saat penelitian, dari judul hingga cetak, akhirnya, toga terpasang juga..(man shabara zhafira...) #see u again at Seol Tower! Hehehe
15. Sahabatku tersayang yang menjadi sumber inspirasi: Eka, Siska, Tya,
Dian, Ovi, Diah, Dwi, Neng,jazkillahatas ukhuwah yang manis ini, episode hidupku menjadi lengkap karenamu...
16. Kaum BORJU (Rombongan Biologi 2007) makasih atas kekompakan dan
kebersamaan selama ini, semoga kesuksesan selalu menyertai kita!!
17.MicroHolic(Mb Win, Mb Ebby, Mb Tukul, “Koloni B4”, Kak Yan, Kak
Asep, Kak Zein) meski kita belajar hal2 yg kecil tapi kita menghasilkan
19.Untuk “lingkaran terdekatku”, mari istiqomah bersama meneruskan risalah
yang mulia ini..
20. Keluarga kecilku di Wisma Ananda atas kebersamaan yang membawa
kebahagiaan, canda tawa yang menghiasi episode hidupku selama jauh
dari keluarga besar.
21. Civitas akademik Jurusan Biologi angkatan 2005, 2006, 2008, 2009, 2010
dan 2011.
22. Serta semua pihak yang tidak dapat penulis sebutkan satu per satu yang
telah membantu baik secara langsung maupun tidak langsung.
Jazakumulloh khoiron katsirpenulis haturkan untuk seluruh pihak yang telah berkontribusi dalam penyelesaian skripsi ini. Penulis berharap karya kecil ini
dapat bermanfaat bagi para pembaca dan mohon maaf atas segala kekurangan
karena sejatinya kesempurnaan hanya milki Alloh semata. Semoga Alloh swt
senantiasa mengumpulkan kita dalam langkah-langkah kebaikan dan menjadikan
kita hamba yang selalu bersyukur. Aamiin.
Bandar Lampung, Juni 2012
Penulis