KERAGAMAN KOMUNITAS BAKTERI PEMFIKSASI
NITROGEN PADA TANAH SAWAH DI SUKABUMI
MUNJIATI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Keragaman Komunitas Bakteri Pemfiksasi Nitrogen pada Tanah Sawah di Sukabumi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
MUNJIATI. Keragaman Komunitas Bakteri Pemfiksasi Nitrogen pada Tanah Sawah di Sukabumi. Dibimbing oleh IMAN RUSMANA dan ALINA AKHDIYA.
Dinitrogenase merupakan enzim penting dalam proses fiksasi nitrogen. Enzim tersebut memiliki 2 subunit protein, yaitu protein Fe dan protein MoFe yang masing-masing secara berurutan disandikan oleh gen nifH dan nifD. DGGE merupakan salah satu teknik metagenom yang dapat merepresentasikan keragaman dan kelimpahan spesies mikrob yang mendiami suatu lingkungan berdasarkan perbedaan melting point dari tiap-tiap sekuen DNA pada gradien senyawa denaturan. Penelitian ini bertujuan mengetahui keragaman bakteri pemfiksasi nitrogen pada tanah sawah. Sampel tanah sawah yang telah diberi perlakuan pupuk NPK 100% takaran (250 kg/ha) (K), pupuk NPK 25% takaran (62.5 kg/ha) dengan penambahan pupuk hayati (C), dan pupuk NPK 25% takaran (62.5 kg/ha) tanpa pupuk hayati (TC) diambil untuk diekstraksi DNA-nya, kemudian diamplifikasi dan dianalisis menggunakan gel DGGE. Diversitas gen nifH lebih banyak dibandingkan dengan nifD. Hasil pendugaan ialah terdapat minimal 7 jenis bakteri pemfiksasi nitrogen pada sampel C, 5 jenis pada sampel TC, dan 1 jenis pada sampel K. Gen nifH lebih dapat membedakan keragaman sekuen daripada gen nifD.
Kata kunci: nifH, nifD, DGGE, fiksasi nitrogen.
ABSTRACT
MUNJIATI. Diversity of Nitrogen Fixing-Bacteria Community on Rice Field in Sukabumi. Supervised by IMAN RUSMANA and ALINA AKHDIYA.
Dinitrogenase is an important enzyme in nitrogen fixation. The enzyme has 2 sub unit proteins, Fe Protein and MoFe protein, each encoded by nifH and nifD. DGGE is one of metagenomic analysis technique that can represent the abundance of specieses in environment, based on melting behavior of amplicones in linearly increasing denaturant in polyacrylamide gel. The aim of this research was to assay diversity of nitrogen fixing-bacteria on rice field. The soil samples of rice field treated with inorganic fertilizer NPK 15:15:15 250 kg/ha (100% dosage) (K), biofertilizer and NPK 62.5 kg/ha (25% dosage) (C), and NPK only 62.5 kg/ha (25% dosage) without biofertilizer application (TC), were extracted for DNA. Those DNA were amplified using primer-GC, hence, analyzed using DGGE. The nifH gene had high variability more than nifD gene. It was expected that there were at least 7 kinds of nifH gene in sample C, 5 in sample TC, and 1 in sample K. High genetic variability of nifH genes was caused evolutionary line. In the other hand, nifD genes tended to be conserved.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
KERAGAMAN KOMUNITAS BAKTERI PEMFIKSASI
NITROGEN PADA TANAH SAWAH DI SUKABUMI
MUNJIATI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : Keragaman Komunitas Bakteri Pemfiksasi Nitrogen pada Tanah Sawah di Sukabumi
Nama : Munjiati NIM : G34090011
Disetujui oleh
Dr Ir Iman Rusmana, MSi Pembimbing I
Ir Alina Akhdiya, MSi Pembimbing II
Diketahui oleh
Dr Ir Iman Rusmana, MSi Ketua Departemen
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah Tuhan Yang Maha Esa, karena dengan rahmat dan petunjuk-Nya karya ilmiah ini berhasil diselesaikan. Topik penelitian yang dilaksanakan sejak bulan Januari 2013 ini ialah keragaman, dengan judul Keragaman Komunitas Bakteri Pemfiksasi Nitrogen pada Tanah Sawah di Sukabumi.
Terima kasih penulis ucapkan kepada Bapak Dr Iman Rusmana, MSi dan Ibu Alina Akhdiya, M.Si selaku pembimbing, serta Ibu Dr Sri Listiyowati, M.Si selaku penguji, atas bimbingan dan perbaikan skripsi ini. Ucapan terima kasih tak lupa penulis sampaikan kepada Ibunda tercinta dan keluarga besar atas segala dukungan baik materi maupun moril. Di samping itu, penghargaan penulis sampaikan pada Kak Lena Novita, S.Si atas saran dan kerja samanya, rekan-rekan Biologi 46, rekan se-almamater Smast, serta teman-teman di laboratorium Mikrobiologi yang telah banyak membantu pelaksanaan penelitian ini.
Penulis berharap agar karya ilmiah ini bermanfaat bagi penulis khususnya dan para pembaca umumnya.
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
Tujuan 1
METODE 1
Waktu dan Tempat 1
Alat dan Bahan 2
Prosedur Penelitian 2
HASIL 4
Hasil Ekstraksi dan Kuantifikasi DNA 4
Amplifikasi Gen nifH dan nifD 4
Running DGGE Gen nif 5
Nested PCR, Sekuensing dan Analisis Blast 5
PEMBAHASAN 6
SIMPULAN DAN SARAN 8
Simpulan 8
Saran 8
DAFTAR PUSTAKA 9
LAMPIRAN 11
DAFTAR TABEL
1 Primer yang digunakan untuk amplifikasi gen nifH dan nifD 3 2 Kondisi reaksi PCR untuk amplifikasi gen nifH dan nifD 3 3 Konsentrasi DNA total (ng/µ L) hasil ekstraksi 4 4 Perbandingan ukuran hasil PCR awal dan nested PCR (pb) 6
DAFTAR GAMBAR
1 Visualisasi elektroforesis amplikon menggunakan primer nifH dan
nifD. 4
2 Visualisasi hasil PCR-DGGE gen nifD 5
3 Hasil elektroforesis DGGE gen nifH 5
4 Visualisasi hasil nested PCR 6
DAFTAR LAMPIRAN
PENDAHULUAN
Siklus biogeokimia di alam berkontribusi pada penggunaan dan penyediaan kembali unsur hara, contohnya nitrogen. Siklus nitrogen meliputi fiksasi nitrogen, denitrifikasi dan nitrifikasi yang seluruhnya dilakukan secara aerob maupun anaerob, artifisial maupun biologis. Fiksasi nitrogen merupakan proses kunci dalam kaitannya dengan siklus nitrogen. Fiksasi nitrogen secara biologis, menyumbang sekitar 70% dari total penambatan nitrogen di alam, sedangkan 30% sisanya terjadi secara alami dengan petir sebagai sumber energinya maupun dilakukan secara artifisial. Fiksasi nitrogen secara biologis hanya dapat dilakukan oleh prokariot yang memiliki kompleks enzim dinitrogenase (Auman et al. 2001).
Enzim dinitrogenase terdiri atas dua sub unit protein, yakni kompleks protein pengoksidasi feredoksin (Fe protein) dan kompleks protein pereduksi N2 menjadi NH3 (MoFe protein). Protein Fe dan protein MoFe masing-masing disandikan oleh gen nifH dan nifD (Mevarech et al. 1980). Kedua gen tersebut dapat digunakan sebagai biomarker untuk analisis komunitas bakteri pemfiksasi nitrogen pendekatan non kultur. Bank data sekuen dari gen tersebut digunakan dalam identifikasi spesies bakteri pemfiksasi nitrogen secara metagenomik.
Salah satu teknik metagenom yang digunakan dalam analisis komunitas mikroorganisme pada suatu lingkungan ialah Denaturing Gradient Gel Electrophoresis (DGGE). Perilaku denaturasi DNA pada suhu tinggi menjadi dasar dari metode ini. Suhu denaturasi (melting point) fragmen DNA dipengaruhi oleh perbandingan komponen basa GC dan AT. Semakin banyak jumlah GC pada sekuen DNA, semakin tinggi suhu yang diperlukan untuk mendenaturasinya. Amplifikasi pada PCR-DGGE menggunakan primer yang mengandung GC-clamp (GC rich sequence) yang berfungsi sebagai clamp saat elektroforesis (Muyzer et al. 1993). Gradien konsentrasi denaturan sepanjang gel akrilamida berfungsi menaikkan suhu denaturasi saat elektroforesis. Oleh karena itu, amplikon hasil PCR-DGGE yang dielektroforesis pada gel bergradien denaturan akan terpisah berdasarkan urutan nukleotidanya. Teknik ini diharapkan mampu merepresentasikan komunitas bakteri pemfiksasi nitrogen pada tanah sawah di Sukabumi.
Tujuan
Penelitian ini bertujuan mengetahui keragaman bakteri pemfiksasi nitrogen pada tanah sawah di Sukabumi berdasarkan analisis PCR-DGGE gen nifH dan nifD.
METODE
Waktu dan Tempat
2
Alat dan Bahan
Bahan yang digunakan dalam penelitian ini antara lain: sampel tanah dari sawah percobaan di Sukabumi, kit isolasi DNA, kit PCR, serta alat dan bahan yang digunakan pada teknik DGGE. Sampel tanah diambil dari petak dengan tiga perlakuan, yaitu pemupukan dengan pupuk anorganik berupa NPK (nitrogen, fosfat, kalium) 15:15:15 sebanyak 250 kg/ha (takaran 100% pupuk normal yang dianjurkan) (K), 62.5 kg/ha (takaran 25% dari normal yang dianjurkan) dengan aplikasi pupuk hayati yang terdiri atas kombinasi isolat bakteri Metanotrof BGM1, BGM5, BGM9 dan SKM14 (Hapsari 2008), Ochrobactrum anthropi BL1 dan BL2 (Setyaningsih et al. 2010), Azotobacter dan Azospirillum koleksi Departemen Biologi FMIPA IPB (C); dan 62.5 kg/ha (takaran 25% dari normal yang dianjurkan) tanpa pupuk hayati (TC).
Prosedur Penelitian Ekstraksi DNA
Ekstraksi DNA dilakukan dengan menggunakan Ultra Clean Soil DNA Isolation Kit (MoBio Laboratories). Prosedur ekstraksi DNA yang dilakukan sesuai dengan protokol yang diberikan oleh produsennya. Ekstraksi DNA dari masing-masing sampel dilakukan tiga ulangan, kecuali sampel K yang dibuat dua ulangan. Sampel dengan konsentrasi DNA tertinggi digunakan sebagai template reaksi PCR.
Kuantifikasi DNA
DNA diukur konsentrasi dan kemurniannya dengan menggunakan Nanodrop.
Elektroforesis dan Visualisasi DNA
DNA hasil ekstraksi dan amplifikasi dicek dengan cara elektroforesis pada gel agarosa dalam Tris-Acetic acid-EDTA (TAE) 1X (1% agarosa untuk DNA genom; 1.25% agarosa untuk DNA amplikon). Elektroforesis dilakukan pada tegangan 80 volt selama 1 jam. Pita DNA pada gel agarosa diwarnai menggunakan 0.1% etidium bromida (EtBr) kemudian divisualisasi menggunakan UV transiluminator.
Amplifikasi Genom
3 Tabel 1 Primer yang digunakan untuk amplifikasi gen nifH dan nifD
Primer Sekuen
nifD F* 5‟-GYGGYTGCGCCTAYGCCGG-3‟
nifD R 5‟-TCCCANGARTGCATCTGRCGGA-3‟
nifH F* 5‟-TAYGGNAARGGNGGNATYGGNAARTC-3‟
nifH R 5‟-ADNGCCATCATYCTNCC -3‟
Y = T atau C; N = A, C, G, atau T; R = A atau G; and D = A, G, atau T
*sekuen GC-clamp (5‟- CGCCCGCCGCGCCCCGCGCCCGTCCCGCCGCCCCCGCCCG-„3) (Rosado et al. 1998) dihibridisasi pada ujung 5‟ primer forward nifH dan nifD
Tabel 2 Kondisi reaksi PCR untuk amplifikasi gen nifH dan nifD
Tahap reaksi PCR* NifH nifD
Polyacrylamide gel electrophoresis (PAGE) 40% dibuat dari akrilamida:bis-akrilamida dengan perbandingan 37.5:1. Konsentrasi PAGE yang digunakan pada penelitian ini ialah 6%. Larutan gel 100% denaturan dibuat dari larutan stok gel 40%, TAE 50X, urea 7M, formamida, dan akuades steril, sedangkan larutan gel 0% denaturan dibuat dari larutan stok, TAE 50X, dan akuades steril. Persentase denaturan dibuat menurut Muyzer et al. (1993). Gradien denaturan yang digunakan pada penelitian ini ialah 35-55% untuk gen nifH dan 40-55% untuk gen nifD. 20-50 µL DNA amplikon dimuat pada gel kemudian dielektroforesis dalam TAE 1X pada suhu 600C, 130 V selama 3 jam menggunakan alat DCodeTM Universal Mutation Detection System (BIO-RAD). PAGE diwarnai menggunakan EtBr 0.1% selama 15 menit, kemudian divisualisasi pada UV transiluminator. Nested PCR dan Sequencing Amplikon Gen nifH dan nifD
Pita DNA amplikon pada gel DGGE dipotong menggunakan scalpel steril. Potongan gel direndam dalam tabung mikro berisi 50 µL aqua bidestillata steril, dilanjutkan dengan inkubasi pada suhu 370C selama 2 jam, kemudian disimpan pada suhu 40C (Bodelier et al. 2005). Sebanyak 10 µL (30 ng) DNA hasil elusi digunakan sebagai template untuk reaksi nested PCR. Amplifikasi dilakukan menggunakan primer dan kondisi PCR seperti pada Tabel 2 tetapi tanpa GC-clamp. DNA amplikon kemudian dikirim ke perusahaan jasa sequencing.
Analisis BLAST
4
HASIL
Ekstraksi dan Kuantifikasi DNA
Hasil ekstraksi 8 sampel tanah menunjukkan konsentrasi yang berbeda-beda (Tabel 3). Perbedaan konsentrasi pada tiap sampel mengindikasikan perbedaan kelimpahan bakteri tiap sampel dan efektifitas ekstraksi. Konsentrasi DNA tertinggi diperoleh dari sampel TC 3 dengan konsentrasi 29.3 ng/µ L, sedangkan konsentrasi terendah diperoleh dari sampel C 3 dengan konsentrasi 7.4 ng/µL.
Tabel 3 Konsentrasi DNA total (ng/µ L) hasil ekstraksi No Kode Sampel Konsentrasi DNA [ng/µL]
1 K 1 16.7
2 K 2 7.8
3 C 1 14.8
4 C 2 18.3
5 C 3 7.4
6 TC 1 15.4
7 TC 2 14.0
8 TC 3 29.3
Amplifikasi Gen nifH dan nifD
Visualisasi produk PCR gen nifH dan nifD pada gel agarosa menunjukkan semua sampel menghasilkan amplikon. Pita DNA amplikon K1, C2 dan TC3 lebih tebal dibandingkan dengan amplikon sampel lainnya. Ketiga sampel tersebut dipilih sebagai template untuk amplifikasi gen nifH dan nifD menggunakan primer GC clamp. Ukuran amplikon gen nifH kurang dari 500 pb, sedangkan nifD lebih dari 100 pb (Gambar 1).
Gambar 1 Visualisasi amplikon menggunakan primer nifH dan nifD pada gel agarosa 1%. Keterangan lajur: marker 100 pb (2), nifH C (3), nifH K (4), nifH TC (5), nifD C (7), nifD K (8), nifD TC (9).
0.5 kb 1 kb
5 Running DGGE Gen nif
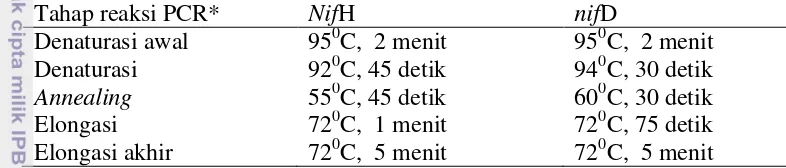
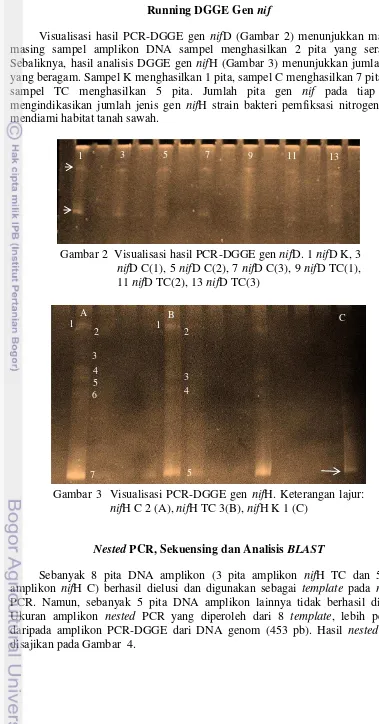
Visualisasi hasil PCR-DGGE gen nifD (Gambar 2) menunjukkan masing-masing sampel amplikon DNA sampel menghasilkan 2 pita yang seragam. Sebaliknya, hasil analisis DGGE gen nifH (Gambar 3) menunjukkan jumlah pita yang beragam. Sampel K menghasilkan 1 pita, sampel C menghasilkan 7 pita, dan sampel TC menghasilkan 5 pita. Jumlah pita gen nif pada tiap lajur mengindikasikan jumlah jenis gen nifH strain bakteri pemfiksasi nitrogen yang mendiami habitat tanah sawah.
Gambar 2 Visualisasi hasil PCR-DGGE gen nifD. 1 nifD K, 3 nifD C(1), 5 nifD C(2), 7 nifD C(3), 9 nifD TC(1), 11 nifD TC(2), 13 nifD TC(3)
Gambar 3 Visualisasi PCR-DGGE gen nifH. Keterangan lajur: nifH C 2 (A), nifH TC 3(B), nifH K 1 (C)
Nested PCR, Sekuensing dan Analisis BLAST
6
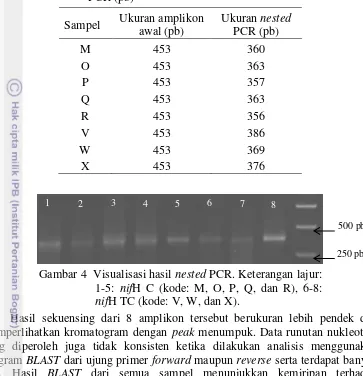
Gambar 4 Visualisasi hasil nested PCR. Keterangan lajur: 1-5: nifH C (kode: M, O, P, Q, dan R), 6-8: nifH TC (kode: V, W, dan X).
Hasil sekuensing dari 8 amplikon tersebut berukuran lebih pendek dan memperlihatkan kromatogram dengan peak menumpuk. Data runutan nukleotida yang diperoleh juga tidak konsisten ketika dilakukan analisis menggunakan program BLAST dari ujung primer forward maupun reverse serta terdapat banyak gap. Hasil BLAST dari semua sampel menunjukkan kemiripan terhadap uncultured nitrogen-fixing bacteria nifH gene.
PEMBAHASAN
DNA amplikon yang diperoleh menggunakan primer nifH berukuran di bawah 500 pb, dan lebih dari 1000 pb untuk gen nifD (Gambar 1). Panjang DNA amplikon yang diperoleh tidak berbeda nyata dengan hasil amplifikasi yang dilakukan oleh Dedysh et al. (2004) yang berukuran 453 (nifH) dan 1130 (nifD) pb. Penggunaan primer nifH reverse yang didesain oleh Zehr & McReynold (1989) serta primer nifH forward yang didesain oleh Boulygina et al. (2002) juga menghasilkan amplikon berukuran 453 pb (Dedysh et al. 2004).
PCR-DGGE gen nifD menghasilkan DNA amplikon lebih sedikit daripada gen nifH (Gambar 2). Jumlah amplikon yang dihasilkan juga seragam untuk semua sampel. Hal ini mengindikasikan tingkat kemiripan sekuen nifD yang sangat tinggi pada bakteri pemfiksasi nitrogen yang terdapat dalam delapan sampel yang dianalisis. Sebaliknya, pita DNA amplikon hasil PCR-DGGE gen
500 pb
250 pb
7 nifH (Gambar 3) lebih beragam jumlahnya. Keragaman gen nifH yang dimiliki oleh bakteri pemfiksasi nitrogen yang terdapat dalam ketiga jenis sampel tanah yang dianalisis lebih tinggi dari gen nifD-nya. Keseragaman pita DNA amplikon gen nifD menandakan bahwa gen tersebut tidak dapat digunakan sebagai markah dalam analisis molekular keragaman gen nitrogenase. Oleh karena itu, analisis keragaman bakteri pemfiksasi nitrogen lebih baik dilakukan berdasarkan gen nifH-nya. Berbagai laporan penelitian juga menyebutkan bahwa gen nifH sering digunakan sebagai bioprobe untuk analisis keragaman maupun identifikasi mikroorganisme pemfiksasi nitrogen (Zehr & McReynold 1989, Boulygina et al. 2002, Quok et al.2003, Dedysh et al. 2004).
Keragaman gen nifH pada sampel C lebih banyak dari sampel TC dan K. Ketiga sampel tersebut berturut-turut menghasilkan 7, 5 dan 1 amplikon. Hal ini mengindikasikan bahwa keragaman bakteri pemfiksasi nitrogen yang terdapat pada tanah yang diberi pupuk hayati dan NPK dosis rendah (25% takaran) lebih tinggi dibandingkan tanah yang hanya diberi NPK dengan dosis rendah (25%) dan tanah kontrol (K) yang diberi dosis lebih tinggi (100%). Sampel K hanya menampilkan satu pita DNA amplikon gen nifH, sehingga diduga bakteri-bakteri pemfiksasi nitrogen yang terdapat pada sampel tersebut hanya memiliki 1 jenis gen nifH yang sama. Pasokan pupuk nitrogen yang tinggi pada tanah akan menghambat aktivitas bakteri pemfiksasi nitrogen (Simanungkalit et al. 1995). Konsentrasi pupuk nitrogen yang tinggi di dalam tanah akan meningkatkan pertumbuhan bakteri lain (bukan pemfiksasi nitrogen), sehingga menjadi dominan di dalam komunitas tersebut. Dominasi bakteri non pemfiksasi nitrogen akan memengaruhi komposisi DNA hasil ekstraksi. Dominasi genom bakteri non pemfiksasi nitrogen mengakibatkan jumlah jenis gen nifH yang diperoleh juga sedikit.
Amplikon PCR-DGGE yang diperoleh dari sampel C lebih tinggi keragamannya dibandingkan TC. Hal ini diduga disebabkan oleh aplikasi bakteri pemfiksasi nitrogen (yang terkandung pada komponen pupuk hayati) ke lahan sawah yang sebelumnya tidak terdapat secara alami atau sudah menghilang dari lahan sawah tersebut. Berdasarkan jumlah DNA amplikon gen nifH yang tampak pada gel DGGE, sampel C diduga mengandung minimal 7 jenis bakteri pemfiksasi nitrogen yang memiliki 7 jenis gen nifH. Sampel TC diduga mengandung minimal 5 jenis bakteri pemfiksasi nitrogen yang memiliki 5 jenis gen nifH. Beberapa bakteri pemfiksasi nitrogen yang terdapat dalam sampel C diduga sama atau memiliki gen nifH yang sama dengan bakteri pemfiksasi nitrogen pada sampel TC. Jenis-jenis bakteri yang menghasilkan amplikon gen nifH sama pada kedua sampel tersebut diduga secara alami sudah ada di tanah sawah. Selain itu, pita tunggal gen nifH pada sampel K yang juga dimiliki oleh sampel TC dan C, berada pada posisi paling bawah pada gel DGGE. Posisi pita terbawah menunjukkan stabilitas migrasi amplikon pada saat elektroforesis suhu tinggi DGGE. Stabilitas amplikon pada suhu tinggi disebabkan konten GC yang tinggi pada runutan gennya. Pita tunggal gen nifH pada sampel K diduga merupakan bakteri tanah yang memiliki aktivitas nitrogenase, serta memiliki high GC content pada genomnya.
8
menunjukkan bahwa aplikasi bakteri pemfiksasi nitrogen yang telah teruji kemampuannya ke lahan pertanian dapat meningkatkan produktivitas tanaman padi. Selain itu, aplikasi pupuk hayati yang diimbangi dengan penurunan penggunaan pupuk dapat meningkatkan kembali keragaman bakteri pemfiksasi nitrogen di lahan sawah.
Hasil sekuensing amplikon pita-pita hasil elusi gel DGGE memperlihatkan peak menumpuk yang kemungkinan diakibatkan oleh jumlah basa degenerate pada primer yang digunakan. Ukuran amplikon lebih pendek kemungkinan diakibatkan oleh sebagian basa terdegradasi akibat paparan UV ketika divisualisasi. Di samping itu, hasil alignment sekuen dari arah forward dan reverse tidak konsisten dan memperlihatkan banyak gap. Analisis BLAST menunjukkan semua sekuen amplikon memiliki kemiripan dengan gen nifH dari uncultured nitrogen-fixing bacteria (Lampiran 1). Hasil analisis ini tidak dapat digunakan untuk menentukan jenis atau identitas bakteri pemfiksasi nitrogen yang terkandung pada sampel tanah.
Hasil sekuensing yang kurang baik dapat disebabkan oleh ketidakmurnian template, sehingga mengganggu proses sekuensing. Masalah ini dapat diatasi dengan melakukan kloning produk PCR yang akan disekuensing. Beberapa laporan penelitian mengenai keragaman gen nifH menggunakan teknik DGGE juga melakukan kloning amplikon DGGE (Tan 2011). Selain itu, penggunaan primer tanpa basa degenerate pada proses nested PCR juga dapat menjadi solusi (Bodelier et al. 2005).
SIMPULAN DAN SARAN
Simpulan
Keragaman bakteri pemfiksasi nitrogen pada sampel tanah dengan perlakuan NPK 25% takaran dengan aplikasi pupuk hayati (C) lebih tinggi daripada sampel TC (NPK 25% takaran) dan K (NPK 100% takaran). Sampel C diduga mengandung minimal 7 jenis bakteri pemfiksasi nitrogen yang memiliki 7 jenis gen nifH. Sampel TC diduga mengandung minimal 5 jenis bakteri pemfiksasi nitrogen yang memiliki 5 jenis gen nifH. Bakteri pemfiksasi nitrogen yang terdapat pada sampel K hanya memiliki 1 jenis gen nifH yang sama.
Saran
9
DAFTAR PUSTAKA
Auman AJ, Speake CC, Lidstrom ME. 2001. NifH sequences and nitrogen fixation in type I and type II methanotrophs. Appl.Environ.Microbiol 67(9): 4009–4016.doi: 10.1128/AEM.67.9.4009-4016.2001
Bodelier PLE, Franke MM, Zwart G, Laanbroek HJ.2005. New DGGE strategies for the analyses of methanotrophic microbial communities using different combinations of existing 16S rRNA-based primers. FEMS Microbiol. Ecol. 52: 163–174.
Boulygina ES, Kuznetsov BB, Marusina AI, Tourova TP, Kravchenko IK, Bykova SA, Kolganova TV, Galchenko VF.2002. A study of nucleotide sequences of nifH genes of some methanotrophic bacteria. Microbiology 71: 500–508.
Dedysh SN, Ricke P, Liesack W. 2004. NifH and nifD phylogenies: an evolutionary basis for understanding nitrogen fixation capabilities of methanotrophic bacteria.Microbiology 150: 1301–1313.
Hapsari W. 2008. Isolasi dan karakterisasi bakteri metanotrof asal sawah di Bogor dan Sukabumi [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Mevarech M, Rice D, Haselkorn R.1980. Nucleotide sequence of a cyanobacterial nifH gene coding for nitrogenase reductase. Proc. Natl. Acad. Sci 77(11):6476-6480.
Muyzer G, de Waal EC , Uitterlinden AG. 1993. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 59 (3): 695-700.
Pingak GMF. 2013. Efektivitas bakteri metanotrof dan Ochrobactrum anthropi sebagai pupuk hayati dan pereduksi emisi gas CH4 serta N2O di sawah anorganik [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Quok CC, Samian MR, Najimudin N. 2003. Phylogeny and characterization of three nifH-homologous genes from Paenibacillus azotofixans. Appl. Environ. Microbiol. 69: 3658-3662.
Rosado AS, Duarte GF, Seldin L, Elsas JD. 1998. Genetic diversity of nifH gene sequences in Paenibacillus azotofixans strains and soil samples analyzed by denaturing gradient gel electrophoresis of pcr-amplified gene fragments. Appl. Environ. Microbiol. 64: 2770–2779.
Setyaningsih R, Rusmana I, Setyanto P, Suwanto A. 2010. Physiological characterization and molecular identification of denitrifying bacteria possessing nitrous oxide high reduction activity isolated from rice soils. Microbiol. Indones. 4:75-78.
Simanungkalit RDM, Hastuti RD, Indrasumunar A, Pratiwi E, Roughley RJ.1995. Soybean response on nodulation to starter nitrogen and inoculation with Bradyrhizobium japonicum. J.Crop. Science 10: 25-32
10
11
LAMPIRAN
Lampiran 1 Hasil BLAST sekuen dari 8 sampel hasil nested PCR
No Sample Blast result Maximal
Score
Total Score
Query Cover
Identity Accession
1 M-Forward Uncultured bacterium clone hytp-77-11
dinitrogenase reductase (nifH) gene partial cds
110 110 37% 82% JQ928220.1
M-Reverse Uncultured nitrogen-fixing bacterium mRNA
for dinitrogenase reductase, partial cds
226 226 73% 83% AB208403.1
2 O-Forward Uncultured nitrogen-fixing bacterium mRNA
for dinitrogenase reductase, partial cds, clone Sp STR 16
143 143 95% 75% AB208894.1
O-Reverse Uncultured bacterium isolate DGGE gel band
N4 dinitrogenase reductas-like (nifH), partial sequences
165 165 64% 80% HM021144.1
3 P-Forward Uncultured nitrogen-fixing bacterium mRNA
for dinitrogenase reductase, partial cds, clone NE5U-ROR06
331 331 92% 80% AB208404.1
P-Reverse No significant similarity found - - - - -
4 Q-Forward Uncultured bacterium clone S2T4F423203
nitrogenase (nifH) gene, partial cds
73.1 73.1 24% 80% JN122957.1
12
No Sample Blast result Maximal
Score
Total Score
Query Cover
Identity Accession
5 R-Forward Uncultured bacterium clone clo A 42
dinitrogenase iron protein (nifH) gene, partial cds
399 399 83% 91% JX628272.1
R-Reverse No significant similarity found - - - - -
6 V-Forward Uncultured bacterium clone BN nif23
nitrogenase component 11 (nifH) gene, partial cds
340 340 86% 86% HQ190134.1
V-Reverse No significant similarity found - - - - -
7 W-Forward Geobacter daltonii FRC32 complete genome 233 233 82% 81% CP001390.1
W-Reverse No significant similarity found - - - - -
8 X-Forward Uncultured bacterium clone JPS 12-6
dinitrogenase reductase (nifH) gene, partial cds
368 368 90% 87% HM750483.1
13
RIWAYAT HIDUP
Penulis dilahirkan di Kediri, pada tanggal 30 Oktober 1991 dari Ayahanda Masduri dan Ibunda Siti Aminah. Penulis merupakan anak bungsu dari delapan bersaudara. Penulis lulus dari SMAN 1 Kediri pada tahun 2009. Pada tahun yang sama, penulis diterima sebagai mahasiswa program studi Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI). Pada tahun 2009-2010 penulis memperoleh beasiswa BBM, kemudian pada tahun 2010-2013 penulis memperoleh beasiswa Tanoto Foundation. Pada tahun 2012 penulis memperoleh kesempatan mengikuti program magang yang diadakan oleh Tanoto Foundation Internship Program di Asian Agri Sumatera.
Penulis aktif mengikuti beberapa kegiatan dalam masa studi seperti kegiatan Leadership Training of Dormitory dan menjadi ketua lorong salah satu gedung asrama, menjadi reporter dan penulis untuk majalah HIMABIO, Chepalos, Organisasi Mahasiswa Daerah (OMDA) Kediri, menjadi asisten praktikum untuk mata kuliah Genetika Dasar, Botani Umum, dan Genetika Molekular. Penulis juga aktif mengikuti kompetisi olimpiade Biologi untuk tingkat propinsi (OSN Pertamina) dan tingkat nasional (ON MIPA PT) berturut-turut pada tahun 2012 dan 2013.
Selama menjadi mahasiswa, penulis mengikuti beberapa kegiatan penelitian dan praktik kerja lapang. Penulis mengikuti kegiatan studi lapangan pada bulan Juli 2011 di Hutan Pendidikan Gunung Walat, Sukabumi, Jawa Barat dengan judul
laporan “Anatomi Daun Tumbuhan Herba yang Terpapar Cahaya dan yang Ternaung
di Hutan Pendidikan Gunung Walat”. Penulis juga mengikuti kegiatan Praktik
Lapang pada bulan Juni sampai Agustus tahun 2012 di Asian Agri bagian Researches & Development di Laboratorium Oil Palm Tissue Culture Kebun Buatan COPPU,
Pangkalan Kerinci, Kabupaten Pelalawan, Riau dengan judul laporan “Aplikasi