PERTAHANAN METABOLIK DAN ENZIM LITIK DALAM MEKANISME RESISTENSI TANAMAN TERHADAP SERANGAN PATOGEN
Dr. Ir. Edy Batara Mulya Siregar, MS
Program Ilmu Kehutanan Fakultas Pertanian Universitas Sumatera Utara
Pendahuluan
Secara umum tumbuhan akan memberikan respon terhadap serangan patogen dan respon tersebut akan bertanggung jawab terhadap resistensi tanaman terhadap patogen. Akibar adanya serangan patogen akan memberikan reaksi pertahanan untuk melindunginya.
Tanaman akan mempertahankan diri dengan dua cara, yaitu (i) adanya sifat-sifat struktural pada tanaman yang berfungsi sebagai penghalang fisik dan akan menghambat patogen untuk masuk dan menyebar di dalam tanaman, dan (ii) respon biokimia yang berupa reaksi-reaksi kimia yang terjadi di dalam sel dan jaringan tanaman sehingga patogen dapat mati atau terhambat pertumbuhannya.
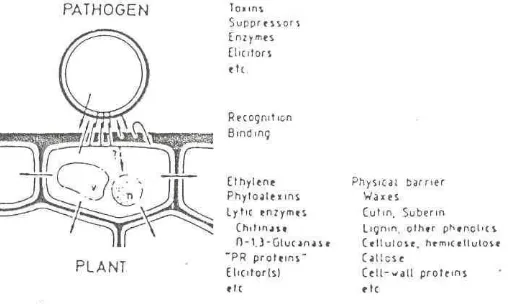
Tanaman akan memberikan respon terhadap patogen dengan cara-cara yang berbeda. Respon tersebut ada yang berinteraksi dan ada yang tidak berinteraksi. Pada kasus tertentu terjadi hubungan yang inkompatibel antara tanaman dan patogen (tanaman adalah resisten) atau hubungan yang kompafibel (tanaman adalah rentan). Namun interaksi yang terjadi antara tanaman dan patogen yang menyerangnya sangatlah kompleks dan banyak melibatkan reaksi-reaksi biokimia. Pada Gambar 1 ditunjukkan kompleksitas interaksi sel tanaman terhadap patogen yang menyerangnya.
Pada kondisi yang normal tanaman adalah resisten terhadap kebanyakan mikroorganisme patogen. Hanya relatif sedikit terdapat kombinasi hubungan patogen-inang yang terjadi. Kejadian biokimia yang terdapat pada interaksi tanaman inang dan bukan inang dengan suatu patogen adalah sama, tetapi intensitasnya dan bentuk penampilannya tergantung pada kondisi lingkungan dan fisiologinya.
Gambar 1. Kompleksitas Interaksi Sel Tanaman Terhadap Serangan Patogen
Pada Gambar 2 di bawah ditunjukkan ringkasan reaksi biokimia yang sering diamati setelah inokulasi tanaman inang dengan patogen. Belum semua reaksi terse but dipelajari secara jauh.
[image:2.612.188.444.531.683.2]1. Struktur Penghalang
Sebelum patogen dapat mencapai proses infeksi, patogen harus terlebih dahulu dapat mengatasi penghalang yang melindungi tanaman. Umumnya virus dan bakteri masuk ke dalam tanaman melalui luka atau dengan bantuan vektor. Cendawan patogenik dapat masuk ke dalam tanaman melalui luka atau bagian-bagian tanaman yang terbuka secara alamiah, seperti stomata atau secara aktif menetrasi lapisan permukaan tanaman.
Struktur penghalang ini termasuk di dalamnya adalah dinding sel yang biasanya disusun oleh lapisan lignin, kutikel, dan berhubungan dengan lapisan pektin dinding sel dan menutupi seluruh bagian permukaan aerial tanaman. Unsur pokok utama adalah kutin, suatu polimer asam hidroksifat yang tidak larutdan melekat di dalam lapisan jilin. Bagian di bawah permukaan dari tanaman dan luka pada bagian aerial tanaman dilindungi oleh suberin. Suberin merupakan suatu campuran polimer fenolik dan bahannya berasaf dari lipid yang diikiat kepada karbohidrat dinding sel.
1.1 Lapisan Lilin
Komponen utama dari lapisan lilin permukaan tanaman adalah C21-C37 n
alkana, C29 dan C9 lebih sering dijumpai dengan rantai panjang. Seringkali satu atau
beberapa alkana merupakan yang predominan. Unsur yang minor adalah monoester yang disusun oleh asam fatty dan rantai panjang alkohol, serta aldehid.
Komponen lilin yang ada pada permukaan atas tanaman tidak berperan secara tunggal dalam pertahanan tanaman melawan patogen. Karakter permukaan lapisan lilin yang hidrofobik juga mekanisme pertahanan tidak langsung terhadap patogen. Telah diketahui bahwa kelembaban adalah esensial untuk germinasi spora dan lapisan lilin yang hidrofobik tersebut akan sangat mengurangi kelembaban pada permukaan tanaman.
Produk yang lipofilik seperti furanocoumarin telah diidentifikasi merupakan unsur lilin yang terdapat pada permukaan daun wortel dan parstley yang secara langsung menghambat pertumbuhan beberapa patogen yang potensil.
Beberapa komponen yang menyusun lilin dapat menstimulir pertumbuhan cendawan, menginduksi sintesis dan ekskresi enzim degradatif seperti kutinase pada cendawan patogenik. Perubahan kompopsisi lapisan lilin yang berhubungan terhadap resistensi sebagai respon terhadap serangan patogen belum diteliti.
1.2 Kutin
Unsur penyusun utama kutin adalah asam fatty C16 dan C18 hidrolisasi. Asam
1O,16-dihidroxypalmitic dan isomernya adalah komponen utama pada grup C16.
Produk yang berasal dari senyawa ini melalui oksidasi dan reduksi merupakan komponen yang minor dan jarang dijumpai. Secara struktur kutin merupakan suatu polimer dari asam hidroksifatty.
Beberapa cendawan patogen tanaman mengeluarkan kutinase yang merupakan suatu enzim yang mengkatalisasi degradasi kutin. Sintesis enzim ini terinduksi oleh adanya kutin pada tanaman. Terjadinya penghambatan yang spesifik oleh enzim ini dapat melindungi tanaman dari infeksi cendawan.
Virulensi strain Fusarium solani f,sp. pisi yang berbeda pada tanaman pea berkorelasi dengan tingkat kutinase yang dihasilkan pada saat perkecambahan spora. Pada konteksi ini kutin merupakan penghalang utama pada tanaman terhadap serangan mikroorganisme fitopatogenik.
1.3 Suberin
Bagian bawah dari permukaan tanaman secara aerial terbentuk periderm yang ditutupi oleh suberin. Suberin adalah suatu campuran polimer yang tidak larut dari unsur-unsur alifatik dan aromatik yang terikat pada dinding sel.
Fraksi yang kava suberin pada umbi kentang sebagai contoh mempunyai komposisi 50% karbohidrat, 5-10%, komponen alifatik lain, dan 40-45% komponen fenolik. Komposisi monomerik dari suberin sedikit sekali yang diketahui.
Cendawan patogen mampu menetrasi suberin setelah proses depolimerisasi secara enzimatik. Cairan kultur F. Solani f.sp. pisi mengeluarkan I bagian alifatik yang sama seperti bagian alifatik pada suberin. Dua esterase ekstraseluler telah dimurnikan dari cendawan ini yang ditumbuhkan pada media yang mengandung suberin. Kedua esterase tersebut mendepolimerisasi komponen alifatik dari suberin dan sangat mirip dengan kutinase dari cendawan yang sama setelah ditumbuhkan pada media yang mengandung kutin.
Beberapa hal yang membingungkan tentang peranan induksi suberisasi dalam reaksi pertahanan tanaman adalah karena secara analisis tidak membedakan secara jelas perbedaan antara suberin dan lignin.
1.4 Lignin
Selama proses penebalan dinding sel yang kedua, dinding sel tanaman dan ruang interseluler diisi dengan polimer fenolik lignin. Lignifikasi terjadi secara enzimatik melalui dehidrogenasi dan selanjutnya diikuti kondensasi radikal coumaryl, coniferyl, dan sinapil alkohol. Pada spesies tanaman yang berbeda, proporsi relatif susunannya juga berbeda. Lignin secara kovalen terikat kepada hemiselulosa dan selulosa melalui ikatan ester dan eter, dan terhadap protein dinding sel seperti dengan glikoprotein yang kaya hidrksoprolin.
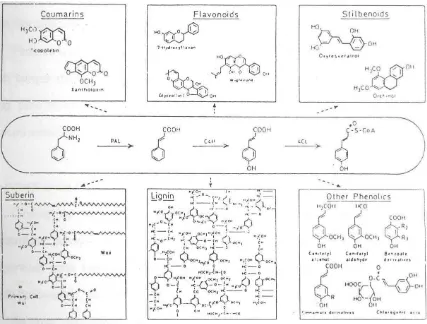
Pada gambar 3 terlihat bahwa prekursor berasal jalur umum phenylpropanoid. Banyak enzim yang terlibat dalam sintesisnya dan regulasinya telah dipelajari sehingga akan diketahui partisipasinya dalam reaksi pertahanan tanaman terhadap serangan patogen.
Seperti diilustrasikan pad a gambar 3 determinasi ini akan sangat sulit karena produk dari jalur phenylpropanoid adalah prekusor dari banyak poduk sekunder tanaman, termasuk diantaranya coumaryl, flavanoid, lignin, suberin dan feolik lainnya yang juga mungkin terlibat dalam resistensi penyakit.
Gambar 3. Illustrasi Biosintesis Berbagai Material Fenolik Pada Tanaman.
Peningkatan lignifikasi lokal disertai dengan peningkatan aktifitas enzim PAL dan peroksidase telah diajukan sebagai mekanisme penting untuk mengurangi penyebaran virus dalam jaringan tanaman. Beberapa enzim yang terlibat dalam biosintesis lignin dan regulasinya telah dipelajari dalam reaksi hipersensitif pada daun tembakau yang diinfeksi oleh TMV. Aktifitas PAL, cinnamic acid 4-hydroxlase, CoA liagase, O-methyl-transferase, dan peroksidse meningkat secara drastis pada area sekitar lesio nekrotik. Aplikasi AOA atau α-aminooxy-β-phe-nylpropionic acid (AOPP) pada daun ini meningkatkan ukuran lesio dan area yang mengandung virus.
Pada banyak kasus penempatan polimer fenolik setelah serangan patogen telah banyak dipelajari. Analisis kimia yang kurang dan kompleks dari material yang baru disintesis setelah infeksi patogen dan adanya kesamaan struktur diantara suberin dan lignin semakin menyulitkan dalam diterminasinya.
1.5 Polisakarida Dinding Sel
Dinding sel tanaman mengandung kira-kira 90% polikarida dan 10% glikoprotein sebagai tambahan dan methyl ester dan eter juga dijumpai. Tergantung pada kelarutannya. polisakarida dibagi ke dalam tiga kelas, yaitu pektik p.olisakarida, hemiselulosa,dan selulosa.
Selulosa merupakan struktur utama polisakarida dinding sel tanaman. Xyloglucan adalah kandungan utama hemiselulosa pada tanaman dikotil, dan xylam kandungan utama hemiselulosa pada tanaman monokotil.
Kalosa yang merupakan suatu glukan adalah unsur yang secara normal terdapat pada polen, pistil, dan plasmodesmata tanaman tingkat tinggi. Sel tanaman mampu mensintesis kalosa secara cepat sebagai respon terhadap stres, seperti pelukaan atau serangan patogen. Pembentukkan papil yang mengandung kalosa seringkali terlibat dalam pertahanan tanaman untuk melawan serangan cendawan dan virus. Akumulasi cepat dengan jumlah besar kalosa dalam papil berkorelasi dengan resistensi barley terhadap penyakit embun tepung. Namun apakah kalosa saja yang membuat tanaman menjadi resisten, masih belum dapat terjawab. Dalam kultur suspensi kacang kedelai, pembentukan kalosa bersamaan dengan adanya akumulasi fitoaleksin. Kejadian ini dielisitasi oleh kitosa, suatu polimer D-glucosamin dari beberapa dinding sel cendawan.
Tampaknya polisakarida dinding sel tanaman berperan sebagai suatu sumber yang disebut elisitor endogen dalam respon terhadap serangan patogen. Frakmen homogalakturosa yang dilepaskan dari dinding sel kacang kedelai mengelisitasi biosintesis fitoaleksin dalam kotiledon kacang kedelai.
1.6 Grikoprotein Dinding Sel
Protein yang berasosiasi dengan dinding sel tanaman umumnya adalah suatu glikoprotein. Glikoprotein dapat terikat secara kovalen atau terlarut dengan derajat yang beragam. Beberapa diantaranya bekerja sebagai enzim, yang lainnya dapat sebagai komponen struktur dinding sel tanaman.
Terdapat kira-kira 3 klas glikoprotein yang mengandung sejumlah besar hidroxiprolin, ekstensin, protein arabinogalaktan, dan lektin. Ekstensin dikeluarkan sebagai HRGP (hydroxiproline-rich glycoprotein) yang larut dan terikat pada dinding sel yang tidak dapat larut. Kira-kira 2/3 bagian dari bentuk yang dapat larut mengandung karbohidrat dan yang 1/3 mengandung protein. Protein arabinoga-laktan adalah HRGP yang dapat larut dan dijumpai ekstraseluler didalam sitoplasma. Perubahan dalam kandungan hidroxiprolin atau HRGP pada dinding sel tanaman terjadi dalam beberapa interaksi patogen-tanaman, tetapi belum diketahui apakah perubahan ini berhubungan dengan suatu resistensi. Pada dinding sel Cucumis melo jumlah HRGP meningkat sepuluh kali setelah infeksi dengan cendawan Collelotrichum lagenarium, bersamaan dengan peningkatan yang drastis produksi etilen oleh inang. Elisitor yang berasal dari miselium cendawan ini atau dari dinding sel melon menyebabkan reaksi yang sama. Perlakuan bibit dengan etilen sebelum infeksi meningkatkan kandungan HRGP dinding sel dan resistensi terhadap patogen. Penghambatan sintesis HRGP oleh 2 trans-hydoxiprolin menurunkan jumlah HRGP dinding sel dan meningkatkan kolonisasi patogen didalam inang.
Suatu elisitor yang berasal dari Phytophthora megasparma f.sp.Glycinea menstimulasi sintesis etilen dan HRGP dalam hipokotil kedelai.
Akumulasi HRGP dan RNA-nya dalam jaringan terinfeksi dan kultur sel yang diperlakukan dengan elisitor telah dipelajari dalam patosistem kacang panjang (Phaseolus vulgaris) dan Colletotrichum lindemuthianum. Dalam sistem ini kandungan HRGP dinding sel meningkat sebelum terjadi akumulasi fitoaleksin, sedangkan akumulasi putatif mRNA ekstensin turun dengan cepat.
Lektin dari solanaceae adalah klas ketiga HRGP ekstraseluler. Lektin terdapat secara ekstraseluler dan intraseluler. Kandungan proteinnya kira-kira 50-60% dengan asam amino utama terdiri dari hydroxiprolin, serin, glisin, dan sistem. Sejauh ini belum diketahui dengan jelas apakah lektin berperan dalam interaksi pato-gen- inang.
isoditirosin ekstensin. Peningkatan ektifitas peroksidase setelah serangan patogen telah didemonstrasikan. Glicosil hidrolase merupakan komponen umum dinding sel, dan bertanggung jawab untuk melepaskan elisitor dari cendawan atau dinding sel tanaman untuk inaktifasi elisitor.
Kesimpulan
Adanya sifat-sifat struktural pada tanaman yang berfungsi sebagai penghalang fisik dapat menghambat patogen untuk masuk dan menyebar di dalam tanaman.
Struktur penghalang yang terdapat di dalam tanaman antara lain adalah lapisan lilin, kutin, suberin, lignin, polisakarida dinding sel, dan glikoprotein dinding sel.
Bahan Referensi
Agrios, G.N. 1988. Plant Pathology 3nd Edit. Academic Press. New York. 803p.
Bell, A.A. 1988. Biochemical mechanisms of diseases resistance. Annu. Rev. Plant Physiol. 32:21-8.1
Hahlbrock, K. And D.Scheel. 1987. Biochemical Responses of Plants To Pathogen. In Chet, I. (edit.). Innovative P,pproaches Plant Diseases Control. John Wiley and Sons New York. 372p.
Ouchi, S. 1983. Induction of resistance of suceptibility. Annu.Rev. Phytopathol. 21 : 189-315.
Paxton, J.and J.Groth. 1994. Constrain on pathogen attacking Plant. Critical Rev.in PlantSciences.13(1): 77-95.
Sequiera, L. 1985. 1983. Machanism of induced resistance in Plant. Annu.Rev. Microbiol. 37 : 51-79.