Annals of Biomedical Engineering, Vol. 16, pp. 379-401, 1988 0090-6964/88 $3.00 + .00 Printed in the USA. All rights reserved. Copyright 9 1988 Pergamon Press plc
A MATHEMATICAL STUDY OF HUMAN
INTRACRANIAL HYDRODYNAMICS
PART I - T H E CEREBROSPINAL FLUID
PULSE PRESSURE
Mauro Ursino
Department of Electronics, Informatics and Systemistics University of Bologna
Italy
(Received 7/6/87; Revised 1/5/88)
A n original mathematical model o f human intracranial hydrodynamics is pro- posed. Equations able to mimic the behavior o f the intracranial arterial vascular bed, intracranial venous vascular bed, cerebrospinal fluid absorption and production pro- cesses, and the constancy o f overall intracranial volume are separately presented and discussed. The model parameters were given normal values computed using physio- logical considerations and recent anatomical data. In this paper the model is used to simulate the genesis and morphology o f the intracranial pressure pulse wave. In par- ticular, dependence o f the intracranial pressure pulse amplitude on mean intracranial pressure, obtained f r o m the model, shows excellent agreement with recent experimen- tal findings. The model explains the intracranial pressure pulse wave as the result o f the pulsating changes in cerebral blood volume (related to cerebrovascular compli- ance) which occur within a rigid space (i.e., the craniospinal compartmenO. A t low and medium values o f intracranial pressure, the intracranial pressure pulse amplitude mainly reflects the cerebral pressure-volume relationship. However, during severe intracranial hypertension, an abrupt increase in the cerebrovascular compliance becomes evident, which is reflected in an abrupt increase in the intracranial pressure pulse wave.
K e y w o r d s - - C S F dynamics, Intracranial pulsatility, Mathematical simulation.
I N T R O D U C T I O N
Knowledge of the major causes affecting the morphology and dynamics of the intracranial pressure (ICP) pulse wave is of the greatest value both for neurological practice and physiological research.
From a hemodynamical point of view the ICP directly affects both the transmural pressure and the perfusion pressure o f the cerebral vascular bed; its changes are com- parable to opposite changes in the systemic arterial pressure (SAP) (30). From a clin- ical point of view, examination of the ICP represents an essential condition for the
Address correspondence to M a u r o Ursino, Department o f Electronics, Informatics and Systemistics, Viale Risorgimento 2, 40136 Bologna, Italy.
comprehension and treatment of several brain disorders associated with intracranial hypertension. Finally, observation of ICP dynamics is particularly useful in deter- mining some parameters of clinical value, like intracranial compliance and cerebrospi- nal fluid (CSF) outflow resistance (21,25,26).
Many studies have appeared in recent years to clarify the origin and clinical sig- nificance of ICP dynamics. Most of these studies were experimental, providing interesting data both on man (2,3,4,11,24,33) and animals (3,9,10,24). A few math- ematical models of overall intracranial hydrodynamics have also been proposed in order to quantitatively describe the CSF pressure and volume changes in several clin- ical and experimental conditions (20,26,28). However, most authors who developed mathematical formulas for intracranial hydrodynamics focused their attention only on a particular aspect of it, like the CSF pressure-volume relationship (2,4,13) CSF production and absorption kinetics (16) or the collapsibility of the cerebral venous vascular bed (8).
In the following, an original mathematical model of overall human intracranial hydrodynamics is proposed in order to elucidate the role of different factors in determining the morphology and time pattern o f the CSF pulse pressure. Both the arterial and the venous, as well as the tissue compartments and the CSF production and absorption mechanisms are separately examined from a common mathematical point of view and included in the model. Autoregulatory adjustments of arterial- arteriolar resistance in order to change flow towards its control value have also been introduced.
The main original aspect of the present model is its ability to take a great num- ber of different features of intracranial dynamics simultaneously into account. To our knowledge this characteristic is unique, since no model proposed in the past has been able to combine so many specific properties of cerebral hydrodynamics. For instance, many studies do not incorporate the intracranial arterial and venous com- pliances; collapsibility o f the venous vascular bed is often left out also. The intro- duction of autoregulatory adjustments in cerebrovascular resistance is also new and can be considered an original feature of the present study with respect to similar pre- vious works.
The importance of having combined many aspects of intracranial dynamics is especially stressed at high values of ICP. In fact, in this condition, intracranial dynamics is the result of many causes (increase in venous pressure due to venous col- lapse, increase in arterial pulsatility, stiffening of the intracranial compartment, pro- gressive loss of autoregulation, changes in CSF production and absorption rates); all these aspects should be simultaneously taken into account if correct description of the phenomenon under study is to be presented.
In a subsequent paper the same model will be used to mimic the results of some relevant clinical tests concerning the intracranial hydrodynamics of normal man.
S T R U C T U R E OF T H E M O D E L
Human lntracranial Hydrodynamics 1 381
To take all these factors correctly into account, the general model of intracranial hydrodynamics has been divided into four submodels.
1. The Cerebral Arterial C o m p a r t m e n t
In the following, the expression "cerebral arterial compartment" is used to denote that portion of the cerebral vascular bed which extends from the point where the two vertebral and the two internal carotid arteries penetrate the dura mater, up to the level of the smallest intracerebral arterioles and capillaries.
Since the aim of the present model is not to describe propagation of pressure and blood flow along the cerebral arterial vascular bed, but only to synthesize the role played by arterial elasticity and arterial hydraulic resistance in the genesis and mor- phology of ICP, the following simplifications have been adopted: (a) compliance of the cerebral arterial vascular bed is mainly due to the large basal arteries. Conse- quently, a unique, lumped arterial compliance, Cai, has been included in the model. This compliance has been assumed equal to the sum of the compliances of all the large intracranial arteries; (b) small cerebral arteries downstream of the circle of Willis have been considered as purely resistive vessels.
With these simplifications the arterial intracranial compartment is described with the electric analog of Fig. 1. P~, Pc and Pic denote the arterial pressure at the point where the arteries penetrate the dura mater, the capillary pressure and the ICP, respectively. Cai is the intracranial arterial compliance, R~i the resistance of the intracranial arterial vascular bed and, finally, q is the cerebral blood flow (CBF).
In order to get a normal value for arterial resistance, Rai, it is necessary to assign normal values for arterial pressure, Pa, capillary pressure, Pc, and CBF, q. These values have been taken as
/3a, = 100 m m H g = 13.3 KPa
Pcn = 25 m m H g = 3.33 kPa
t~, = 12.5 cm3/sec (22) .
Q
---zz~- 9
Cai I
0
Ra i
0
7
o
Pc
382
M. Ursino
In the following, the subscript, n, is always used to denote a quantity in the nor- mal basal condition; the bar denotes mean value.
From these values one obtains
Rain
= 6 m m H g sec/cm 3.The value of Rai changes due to autoregulatory adjustments in the caliber of cere- bral arteries and arterioles. It has been demonstrated (see, among others, 18,23,29,30) that arterial resistance adjustments maintain cerebral blood flow rather close to nor- mal, despite changes in cerebral perfusion pressure, provided this last does not exceed a lower and an upper limit (about 60-70 and 130-140 mmHg, respectively).
This phenomenon has been simulated by making use of the following equations
dx
1
1 ( Pa-- Pv-- Pan + Pvn)
- x + - ~ = - P - 7 ~ ( 1 )
dt
r
r
Gai =
Gain( 1
7rl arctg(xTr))
(2)where Pv is the cerebral venous pressure and G~g the cerebral arterial conductance (i.e., Gag = l / g a i ).
According to differential Eq. 1, the autoregulatory action, x, depends on perfu- sion pressure percent changes. Moreover, its dynamics has been characterized by a a single real pole, with time constant r. In order to reproduce the time pattern of autoregulatory responses reported in Kontos
et al.
(23) r was given the value 2 sec. The arctan in Eq. 2 simulates the existence of an upper and a lower limit of autoregulation. When changes in perfusion pressure are very small, the result isGai = Gai n [
1
- x]that is, the percent changes in arterial conductance are equal to the percent changes in perfusion pressure. However, when perfusion pressure exceeds the lower or upper limit of autoregulation, Gai settles at a value 50~ greater or less than normal, respectively.
The dependence of arterial intracranial compliance, Cag, on pressure has been described by assuming a monoexponential relationship between arterial blood vol- ume, V~, and arterial transmural pressure,
Pa
- P i c (6,17); that isPa - Pi~ = Ploe K'(V'-V'~
(3)
where Plo, Va0 and K~ are constant parameters. According to Eq. 3 any increase in arterial blood volume causes a stiffening of the vessel wall. Equation 3 implies that arterial intracranial compliance is given by
dV~ 1
Cai - d(Pa - Pic) - Ka(Pa - Pic)"
(4)Human lntracranial Hydrodynamics 1 383
normal values of arterial and intracranial pressures and directly evaluating a normal value for arterial intracranial compliance, C~,. To this end the arterial intracranial compliance has been assumed equal to the sum of the compliances of all the large brain arteries. The following formula has been used (6,31)
C~i = ~ 4a'r2(1 - ~ + rk/hk)2
k Ek(1 + 2rk/hk)
t,
(5)
where rk, hk, Ik and Ek denote the inner radius, thickness, length and Young modu- lus of the generic kth vessel, respectively, o is the Poisson ratio, assumed equal to 0.5 for all the vessels.
The values of inner radius and the length o f basal human cerebral arteries in nor- mal conditions have been taken from recent literature (14,15,37,38,43).
Normal values of the other parameters (Young modulus and radius-thickness ratio) which appear in Eq. 5 have been taken from (46).
Starting from these data and by making use of Eq. 5 a compliance value of 2.5.10 -3 c m 3 / m m H g has been obtained. However, this value underestimates over- all intracranial arterial compliance, since the contribution of terminal branches of the anterior, posterior and middle cerebral arteries has not been included. Consequently, human intracranial arterial compliance in normal condition has been given the value
C a i n ~- 3.0" 10 -3 c m 3 / m m H g (6)
which is slightly higher than that computed above.
Finally, from Eqs. 6 and 4, and by making use of the normal values of Pa and P~c (Pie, = 9.5 mmHg) we have
1
Ka = = 3.68 cm -3.
Cal.(Pan - #it.)
2. The C S F Production and Absorption Mechanisms
As is well known, CSF shows a typical circulation from the place where it is pro- duced up to where it is reabsorbed. Even though minor routes also contribute to this process, CSF is mainly formed by the choroid plexi of the cerebral ventricles and is mainly reabsorbed by the arachnoid villi of the dural sinuses (especially of the sagit- tal sinus). Mathematical formulas able to mimic this process are proposed in the following.
C S F Production. It is thought (40) that CSF originates at the choroid plexi of the lateral ventricles through an active or energy requiring process. Nevertheless, evidence that the CSF production rate decreases by increasing the ICP (44) or decreasing the CBF (19) has been reported.
In the present model it is assumed that the CSF formation rate, q/, is propor- tional to the transmural pressure at the choroid capillary level; that is
Pc - P i c
(7)
where Rf denotes the resistance that the choroid capillary system offers to CSF secretion.
The assumption that qf linearly depends on transmural pressure represents a sim- plification of the active processes involved in CSF production. However, this assump- tion seems reasonable according to Starling's hypothesis of water transport in the periphery. Moreover, Eq. (7) allows the major results on CSF production to be well reproduced by the present model.
In fact, the venous vascular bed collapses during intracranial hypertension and thus, in accordance with the collapse mechanism in a flexible tube, CBF linearly depends on the difference between the capillary pressure, Pc, and the ICP, Pic (see below, especially Fig. 2 where Rp~ and R'~s are constant). Consequently, Eq. 7 implies that qf is proportional to CBF: when the ICP does not exceed 30-40 mmHg, ql remains rather constant due to autoregulation, as observed by Higashi et al. (19) and Guess et al. (16). However, at high values of ICP, the CSF production rate progressively declines, as observed by Sahar (44).
CSF Reabsorption. The process by which CSF is reabsorbed by the arachnoid villi
o f the sagittal sinus and other minor routes seems to be purely passive, depending on the difference between CSF pressure and dural sinus pressure and on the resistance that the arachnoid villi offer to liquid outflow. Consequently, the following relation has been adopted
P/c - Pus
q~ - (8)
Ro
where qa is the rate of CSF absorption, Pus the venous sinus pressure and Ro the resistance to CSF outflow.
Values for the CSF outflow resistance have recently been obtained, both in man and animals, using two different methods: the constant infusion technique (21) and the bolus injection technique (25).
Values of Ro for the normal man lie in the range 3-15 m m H g . min/ml (11,27,45). Accordingly, in the present simulation it has been assumed that
Ro = 8.75 m m H g min/ml.
The normal mean value of the venous sinus pressure has been taken as
P~s~ = 6.5 mmHg.
Since, in the normal basal condition, the CSF production rate must equalize the CSF absorption rate, one has
qa, = qf~ = 400 #l/min
Rf = 38.75 m m H g min/ml.
The values of CSF outflow resistance, Ro and CSF production resistance, R f,
Human Intracranial Hydrodynamics 1 385
sponding transmural pressure was positive. Where the corresponding transmural pres- sure was negative, the resistance value was assumed as infinite.
3. The Cerebral Venous Compartment
Pressure within the cerebral veins is determined by many factors, that is, the values of central venous pressure (Pc~), CBF and CSF pressure (the intracranial pressure). This last factor is especially influential in conditions associated with intracranial hypertension, when a collapse of the cerebral venous bed is expected to occur.
In normal conditions the value of dural sinus pressure is lower than that of CSF pressure; therefore, dural sinuses are maintained open only by virtue of the rigidity of their walls. However, pressure within the major cerebral veins is normally higher than CSF pressure and the cerebral veins do not collapse since they are subject to a positive transmural pressure. Accordingly, the following relations normally hold
The normal mean value of pressure in the major cerebral veins (P~) and value of normal central venous pressure (Pcv) have been assumed to be
/3on = 14.0 mmHg
/5c~ ~ = 4.0 mmHg.
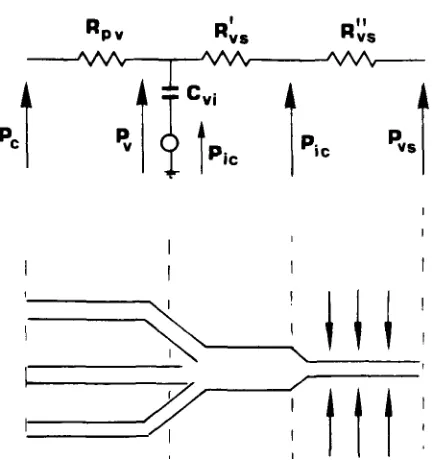
Since the transmural pressure in the cerebral venous vascular bed (Iv - P i c ) is very close to zero, collapse of these veins can easily occur. It is generally thought (8,30,41) that the cerebral venous vascular bed does collapse during intracranial hypertension, through a mechanism similar to that of the Starling resistor. This hypothesis is sup- ported by in vivo observations (32,39,42,47). In particular Yada et al. (47) showed that, in the case of intracranial hypertension, mean intraluminal pressure in the bridge veins and lateral lacunae is always a few mmHg higher than mean ICP, inde- pendently of the ICP value. At the same time, in accordance with the mechanism of the Starling resistor, an abrupt drop in intraluminal pressure does occur at a point 1-2 mm proximal to the junction between the lateral lacunae and the sagittal sinus. In order to mimic this behavior with the present mathematical model, the cerebral venous vascular bed has been subdivided into a series of two segments (see Fig. 2). It is assumed that the first segment (whose hydraulic resistance is denoted by the sym- bol Ruv) extends from the capillary venules up to and including the large cerebral veins. The second segment (whose hydraulic resistance is denoted by the symbol Rvs) extends from the final portion of the large cerebral veins up to the dural sinuses.
! I!
I:lpv R v s R v s
icv
I
I I
'II
i \
I
1 I
FIGURE 2. Electric analog of the cerebral venous vascular bed. Rpv: resistance of the proximal
intracranial venous vascular bed; R~,s: resistance of the distal intracranial venous vascular bed (por- tion upstream of the collapse); R~s: resistance of the distal intracranial venous vascular bed (por- tion downstream of the collapse); Cvi: intracranial venous compliance.
With notations of Fig. 2:
R~s = resistance o f the distal portion o f the cerebral venous vascular bed upstream o f the collapse;
R~;s = resistance of the distal portion of the venous vascular bed downstream of the collapse;
u
Ro~ = R'os + Ro,. (9)
By imposing that, at a point immediately upstream of the collapse, the intraluminal pressure equalizes the extravascular pressure, one obtains
Ro"~P~ + R ' ~ P ~
Pic = (10)
R;, + R~,
From Eqs. 9 and 10 one gets
P ~ - P~s
R v s - - - Rvs.
(ll)
P ~ - P~c
[image:8.504.144.360.77.308.2]H u m a n Intracranial Hydrodynamics 1 387
itself particularly useful in reproducing the relationship between cerebral venous and intracranial pressures in several conditions associated with intracranial hypertension. The values of the two constant parameters R~s and Rpo have been computed start- ing from the normal values of C1, Pc, P~, Pic a n d / 3 reported above.
The cerebral venous vascular bed is also characterized by a high value o f compli- ance which, being mainly due to the large veins, has been schematized in the pres- ent mathematical model with a unique, lumped parameter, C~, undergoing the transmural pressure P~ - Pic of the large cerebral veins. This simplification is simi- lar to that already used for the cerebral arterial vascular bed.
The relationship between cerebral venous volume and transmural pressure has been assumed to be of the monoexponential type with the addition o f a constant term; that is
P~ - Pic = Pvoe r~cv~-v~~ + P ~ (12) where Pro, Po1, K~ and V~o are constant parameters. Expression of the cerebral venous compliance thus becomes
dVo 1
C~i - d ( P o - Pie) - K ~ ( P ~ - Psc - P~I) (13) where Kv denotes the elastance coefficient o f the venous vascular bed, while the parameter Pvl represents the transmural pressure value at which the large cerebral veins would collapse and, therefore, venous compliance would tend to become infi- nite. Taking data reported in (31,36) the value of P~I has been assumed slightly negative; that is
Pv~ = - 2 . 5 mmHg.
The collapse mechanism of the venous vascular bed may also contribute to com- pliance. This problem has been extensively studied by Burattini et al. (7). The authors demonstrated that, when the inflow pressure in a tube is reduced below the external pressure, a complete collapse in the distal tube occurs during part of the cycle and, in this condition, compliance of the collapsed portion becomes extremely high. How- ever, this is not the case of the present simulation where the venous pressure always exceeds extravascular pressure, the venous flow never stops and the distal portion of the venous vascular bed is only partially collapsed. Compliance of the distal partially collapsed section is not easily identifiable in this case; the appropriate lengths of the collapsed (R~s) and noncollapsed portion (Rpv + R~s) should be known (7). Since Yada et al. (47) experimentally observed that collapse in the cerebral venous vascu- lar bed is limited to a very short segment (1-2 mm before entering the dural sinuses) its contribution to overall venous compliance can reasonably be assumed to be negligible.
I f a vascular bed is roughly approximated with the parallel o f n equal blood ves- sels of length l, Young modulus E, internal radius r and diameter thickness ratio d/h, the result is
2 7rnr31 Vd
C =- - (14)
Eh Eh
where a simplified thin-walled expression has been used for the compliance (obtain- able, for instance, f r o m Eq. 5 when h/r is low and ~ = 0) and V = 7rr2nl denotes the overall internal volume o f the vascular bed. Equation 14 shows that, in a first approximation, compliance of a vascular bed can be considered as directly propor- tional to the blood volume and the diameter-thickness ratio and inversely propor- tional to the Young modulus.
Consequently the ratio between the venous and arterial intracranial compliances is given by
C,,~
V,,
E~(d/h)oCai V~ Eo(d/h)~"
(15)
Since the cerebral venous blood volume in normal conditions is approximately 7 0 % - 8 0 % o f the total brain blood volume (1) it is:
Vo / V~ _~ 2.5.
The values o f the Young modulus and the diameter thickness-ratio for compliant cerebral arteries in normal condition have been taken f r o m (46) lEa = 8.10 6
dyn/cm2; (d/h)a = 8].
The values o f venous Young modulus and diameter-thickness ratio have been taken f r o m (36) lEo = 1.1-10 6 dyn/cm2; (d/h)v = 66].
By inserting the values reported above into Eq. 15 one obtains
Coin = 0.46 c m 3 / m m H g
Kv = 0.31 cm -3.
The validity of the venous compliance value obtained in this way is supported by the observation that, with such a value, the present mathematical model can well reproduce the I C P rise following an obstruction in the extracranial venous return (further publication, part 2).
Human Intracranial Hydrodynamics 1 389
4. The D y n a m i c s o f the Craniospinal C o m p a r t m e n t
Since the cranial cavity is a closed space, its volume must always remain constant. The volume of the cranial cavity is the sum of four major contributions: the arterial and the venous blood volumes (V~ and V~, respectively), the CSF volume (VcsF) and the volume of cerebral tissue (Vtiss). Constancy of the total intracranial volume is therefore expressed by the following differential equation, which reflects the origi- nal Monro-Kellie principle (29)
d V~ d Vo d Vcsr d V~i~
- - + + - - + - O . ( 1 6 )
dt dt dt dt
Since the CSF can be considered as an incompressible liquid, the time derivative of CSF volume is due only to the difference between the CSF entering and leaving the cranial space in unit time: that is
dVcsv
dt - qf - q~ + Ii. (17)
The term Ig denotes the rate at which saline is artificially injected into the CSF space. This term has been introduced into the model in order to simulate the results of some tests largely in use in neurological practice (see part 2).
CSF production rate, qy and CSF absorption rate, qo, have been given the expres- sion 7 and 8, respectively.
Since Cat and Cvg are the compliances of the intracranial arterial and venous vas- cular beds, respectively,
dV. =Cai d ~ (P~ - P,c) (18)
dVv d
d-~- = Cvi ~ (Po - Pie).
(19)
Both Cag and Cog are dependent on the local value of transmural pressure, according to Eqs. 4 and 13.
Finally, cerebral tissue volume can be assumed as a nonlinear function of the intracranial pressure: when ICP rises, the cerebral tissue is compressed and, there- fore, its volume is reduced. Several authors (4,25,26,35) have observed that the pressure-volume relationship of the craniospinal compartment, both in man and ani- mals, has approximately a monoexponential form, at least for a segment above and below the resting pressure, as revealed by the fact that elastance linearly increases with ICP. Particularly in man (12) the increase in craniospinal elastance is fairly lin- ear up to a CSF pressure of about 3-4 KPa (22.5-30 mmHg); thereafter, the elastance increase becomes steeper.
E.~s = Cti-~Is = ~ - g ~ Pi~ + \ eo~ / ]
(20)
where K e and PoL are constant parameters.
If ICP is low, the quadratic term in Eq. 20 is negligible and the dependence of tis- sue elastance on pressure is fairly linear, as in the traditional monoexponential model. However, when ICP rises, the quadratic term becomes relevant, which is reflected in a steeper increase in tissue elastance.
Equation 20 implies that
dVtiss d P ic
- - = -Ctiss - - (21)
dt tit
Values for Are and P01 have been given in order to best fit experimental data (see after). We have:
Ke = 0.26 cm -3 Po~ = 7.5 mmHg (1 KPa).
The value of K e is in the range of the elastance coefficients normally reported for man (2,34,45) and it is equal to the mean value found by Avezaat and van Eijnd~ hoven (2).
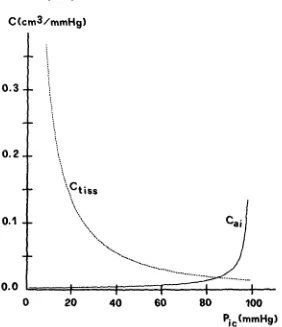
Figure 3 shows a comparison between the value of tissue compliance [Ctiss = E~s~], evaluated through formula 20 and that of arterial compliance (Cog) resulting from formula 4. The plot of C~i has been obtained by giving arterial pressure a nor- mal value (100 mmHg). As shown in Fig. 3, the model implies that the effect of arterial compliance is negligible at ICP values below 60-70 mmHg. On the contrary, when ICP rises above 60-70 mmHg, compliance of the arterial intracranial vascu- lar bed becomes relevant, thus producing an increase in CSF pulse pressure and a flattening in the volume pressure response, as experiment observed by Avezaat et al.
(3,4) on dogs and patients.
M O D E L S I M U L A T I O N
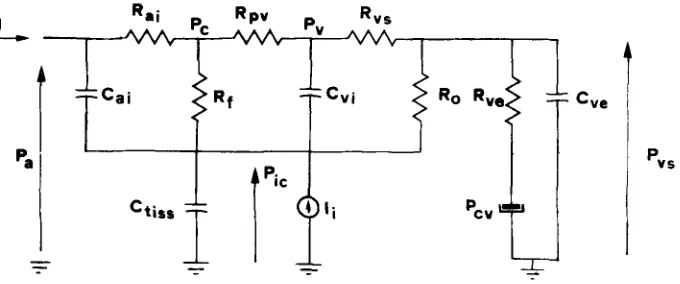
According to the general considerations developed above, human intracranial hydrodynamics is described by the electric analog of Fig. 4. The model is completed by Eqs. 1 and 2, which mimic autoregulation, Eqs. 4, 13 and 20, which represent the dependence of the arterial, venous and tissue compliance from pressure, and by Eq. 11, which reproduces the collapsibility of the venous vascular bed.
The model is of the fourth order, with the autoregulatory action (x), the cerebral venous, intracranial and dural sinus pressures as state variables. The input variables of the system are the arterial pressure and the mock CSF injection rate. The param- eters of the model were given the set of values reported in Table 1. Symbol G in Table 1 is used to denote hydraulic conductances (i.e., the inverse of the correspond- ing hydraulic resistances). The system has been numerically integrated on a VAX/VMS computer, by using the 4th order Runge-Kutta method.
Human lntracranial Hydrodynamics 1 391
C(cm3/mmHg)
0.3
0.2
_- i
i
i
\
Ctiss
j
0.1 ""\.,
Cai
0.0
~ i _I I 1 I I
0
20
40
60
80
100
Pic(mmHg)
FIGURE 3. Plots of the tissue cornpliance (C~;,s) and of the intracranlal arterial compliance (C,i) vs. ICP. computed for an arterial pressure value of 1 0 0 mrnHg.
paper (part 2) the model's capability in reproducing human hydrodynamics will be further tested in several conditions of clinical significance.
Arterial pressure has been given an unchanged pulsating waveform throughout the present simulation (diastolic pressure 75 mmHg, systolic pressure 125 mmHg, period 0.8 sec, Fig. 5), independently of the ICP mean value. This means that the system formed by the heart and the extracranial arterial vascular bed (extending from the aortic valve up to the point where arteries penetrate the dura mater) has been schema-
TABLE 1. Parameter values used during the present simulation.
G~i = 0 . 1 6 6 cm 3 sec -~ m m H g -'~ G~,= = 2 . 7 7 c r n 3 sec -~ mrnHg -~
G f = 0 . 4 2 . 1 0 - 3 cm 3 sec -~ m m H g -1 C ~ , = 2 . 3 ~ , c r n 3 m m H g - ~
K~ = 0 . 3 1 c r n - 3
Pvl = - 2 . 5 rnmHg
Gpv = 1 . 1 3 6 c m 3 sec -1 mmHg -1 Go = 1.90- 10 -3 cm 3 s e c - ~ m m H g -~ G ~ = 6 . 2 5 c r n 3 s e c - ~ m r n H g - 1
K= = 3 . 6 8 c r n - 3 KE = 0 . 2 6 c m - 3
[image:13.504.113.395.70.397.2] [image:13.504.74.430.580.655.2]Rai Pc Rpv Pv Rvs
7~
ai I ~ f - ~ ~ v i~ •
t"~ ''
"
Ro Rvel
Pcv "
Cve
FIGURE 4. Electric analog of overall human hydrodynamics, q: cerebral blood flow; Pa: arterial pressure; Pc: capillary pressure; Pv: cerebral venous pressure; Pic: intracranial pressure; Pv~; venous sinus pressure; Cai: intracranial arterial compliance; Ram: intracranial arterial resistance; Rp,,: resis- tance of the proximal venous vascular bed; Rv=: resistance of the distal venous vascular bed (lateral lacunae and dural sinuses); C~;: intracranial venous compliance; R( resistance to CSF formation; Ro: resistance to CSF outflow; Ct~: cerebral tissue compliance;/~: rate of saline infusion; Rve: resis- tance of the extracranial venous pathway; Cv,: extracranial venous compliance; P~: central venous pressure.
tized as an ideal oscillatory pressure source and that the Cushing response has not been included in the model. The consequences of these simplifications are to be discussed.
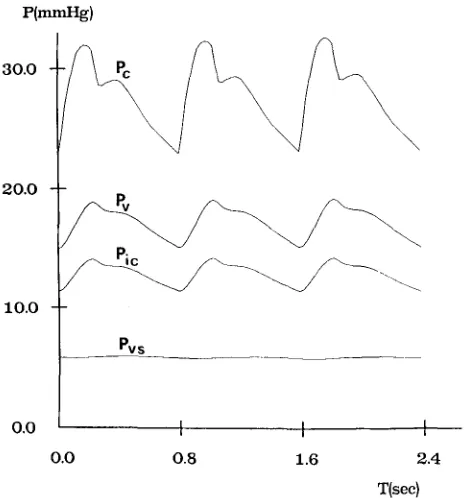
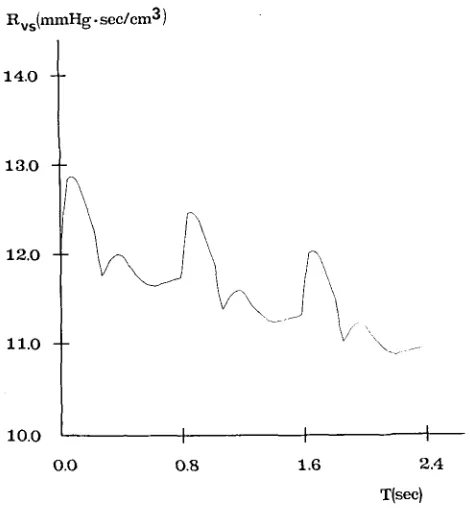
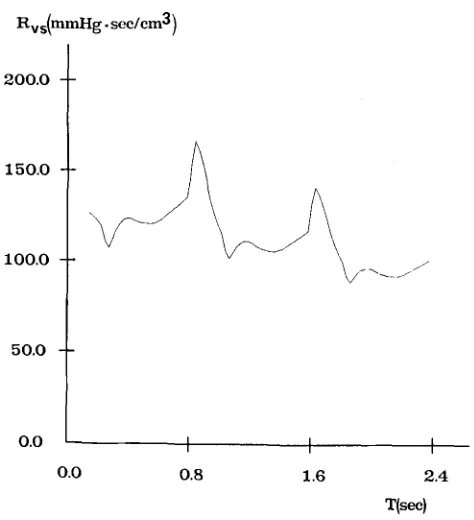
The ICP has been given several different values, ranging between 10 and 93 mmHg, by simulating the injection of various liquid boli into the CSF space. The time patterns of cerebral capillary pressure, cerebral venous pressure, intracranial pressure, venous sinus pressure and of lateral lacunae resistance (Rvs) have been computed over three cardiac periods after the injection. The most significant wave- forms among these quantities are reported in Figs. 6, 7 and 8, which refer to a mod- erate hypertension (diastolic ICP of about 11 mmHg), a mid-high hypertension (diastolic ICP of about 40 mmHg) and a very severe hypertension (diastolic ICP of about 75 mmHg), respectively.
From Figs. 7b and 8b it is evident that intracranial hypertension causes partial col- lapse in the distal cerebral venous vascular bed, which is reflected in a large increase in the resistance Rvs. The slow return of Rw towards baseline, shown in Figs. 7b and 8b, is especially a consequence of the progressive ICP reduction with time, due to CSF reabsorption. ICP time reduction is also evident from Fig. 8a.
Partial collapse of the terminal venous vascular bed makes the capillary pressure, Pc, and the venous pressure, Pv, increase in parallel with ICP, as shown in Figs. 6 and 7a. The difference between venous and intracranial pressure is always only a few m m H g , in accordance with data reported by Yada et aL (47). Venous sinus pressure is almost unaffected by intracranial hypertension (Figs. 6 and 7a).
[image:14.504.84.424.83.225.2]Human lntracranial Hydrodynamics 1
Pa(mmI-Ig)
393
125.0
100.0
75.0
50.0 I I I
0.0
0.8
1.6
2.4
T(sec)
FIGURE 5. Arterial pressure waveform used during the present simulation.
p(aunng)
30.0 ~
~
9 \
20.0
10.0
Pvs
o,o I I t
[image:15.504.135.370.79.336.2] [image:15.504.136.368.377.626.2]0.0
0.8
1.6
2.4
T(sec)
P(mmHg)
8 0 . 0
6 0 . 0
40.0
2 0 . 0
~ s
o.o I I
0.0
0.8
1.6
2.4T(sec)
FIGURE 7a. Waveforms of the capillary pressure (Pc), cerebral venous pressure (Pv), ICP (P;c) and the venous sinus pressure (Pvs) evaluated in the case o f a diastolic ICP o f 4 0 mmHg.
Rvs(mmHg. see/era 3 )
14.0 l
13,0
12.0
ii.0
1o.o I I
0 . 0 O.B
1.6
\ J-
I 2.4
T(sec)
[image:16.504.128.388.72.335.2] [image:16.504.134.369.379.633.2]Human lntracranial Hydrodynamics 1
Pic(ramHg)
395
125.O
I 0 0 . 0
75.0
50.0 I I
0,0 0.8 1.6 2.4
T(sec}
FIGURE 8a. ICP waveform evaluated in the case of a diastolic ICP of 75 mmHg.
Rv s(mmHg "sec/cm3 )
200.0
150.O
100.0
50.0
o.o I I I
0.0 0.8 1.6 2.4
T(sec)
[image:17.504.133.365.83.333.2] [image:17.504.132.369.371.631.2]sorption process, the following formulas have been used to compute I C P P A and M I C P :
Pic(tmax) + Pic(tmax + T )
APic
= -- Pic(tmin) 2Pic = [Pic(tmax) + Pic(tmax q- T ) ] / 2 + Pic(trnin)
where T is the cardiac period, tma x and tma• + T are the instant o f two successive
maximi in the I C P and tmin is the intermediate instant at which the I C P has a minimum.
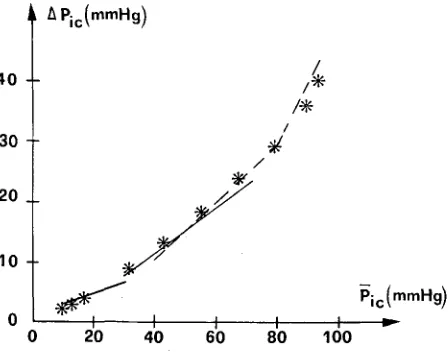
The resulting plot of I C P P A against the M I C P is shown in Fig. 9. In the same fig- ure the experimental curves obtained by Nornes et al. (33) and Avezaat and van Eijndhoven (2) on man are also reported. The agreement between model and exper- imental results is excellent.
The I C P P A shows a linear increase with mean intracranial pressure up to a break- point lying at about 60 m m H g ; above this point the I C P P A increases more rapidly.
D I S C U S S I O N
CSF pulse pressure is the product o f pulsating changes in cerebral blood volume, synchronous with the cardiac beat, which occur within a rigid space (i.e., the intracranial compartment). According to this, CSF pulse amplitude is mainly affected by two factors: the elastance o f the craniospinal system, as derived f r o m the pres- sure-volume relationship (Eq. 20) and the compliance o f the cerebral vascular bed. This last depends on both the intracranial arterial and venous compliances.
4 0
30
20
10
~ Pic(rnmHg)
/
/
f
I I I I
20 40 60 80
/.
Pic(mmHg) I ~- 100
FIGURE 9. Plot of ICP pulse amplitude (~PJc) vs. mean ICP (Pjc). Regression lines experimentally obtained in man by Nornes et al. (33) (dotted line) and by Avezaat and van Eijndhoven (2) (contin-
[image:18.504.138.362.445.623.2]Human Intracranial Hydrodynamics 1 397
The relative importance of arterial and venous pulsatility in producing CSF pulse pressure is still a matter of discussion. Generally, it is expected that in those cases in which the capability of the arterial wall to pulsate is minimal, the CSF pulse wave is mainly the result of cerebral venous pulsatility. On the contrary, when the capa- bility of the arterial wall to pulsate becomes maximal, the CSF pulse wave reflects the greater influence of the arterial waveform. This last situation is thought to occur, for instance, when the arterial transmural pressure is low (i.e., during arterial hypotension (3) or intracranial hypertension).
Figure 3 shows that, according to the present model, compliance of the intracra- nial arterial vascular bed is negligible at low and medium values of intracranial pres- sure. In these conditions, the ICP pulse amplitude appears to be mainly the result of oscillations in cerebral venous blood volume. However, when ICP is high and, con- sequently, arterial transmural pressure is reduced, the influence of arterial compli- ance becomes relevant. Such behavior is reflected both in the morphology of the ICP waveform and in the pattern of the graphic " I C P P A vs. M I C P . "
Figures 6, 7 and 8 show the morphology of ICP pulsations obtained from the model at various levels of ICP. When ICP is low (Fig. 6), the ICP pulse wave is largely different in shape from that of arterial pressure. On the contrary, the mor- phology of arterial and intracranial pressure pulse waves becomes similar when ICP tends to arterial pressure (Fig. 8).
A similar result is in agreement with the observations made by Dereymaker et al.
(10). These authors experimentally observed that the morphology of CSF pulsations is of the "venous type" at normal values of arterial pressure, whereas it is of the "arterial type" when the arterial pressure is reduced (that is, when the arterial trans- mural pressure is low).
Dependence of the ICPPA on MICP is also well simulated with the model. Several authors (2,3,4,33) observed that, both in the dog and man, the I C P P A moderately increases with CSF pressure up to a breakpoint lying at about 60-70 mmHg. Further increases in CSF pressure above breakpoint cause an abrupt, disproportional increase in CSF pulsatility.
The present mathematical model reproduces this pattern very well, as shown by the curves of Fig. 9. In accordance with the considerations developed above, the lin- ear increase in the I C P P A below breakpoint is explained by the model with the pro- gressive increase in craniospinal elastance, due to the exponential nature of the pressure-volume relationship (Eq. 20). The sharp increase in pulse amplitude above breakpoint is imputed to the sudden rise of cerebrovascular compliance, especially that of the arterial intracranial vascular bed (Eq. 4 and Fig. 3), consequent on the low value of transmural pressure.
Moreover, during severe intracranial hypertension the intracranial arterial compliance becomes very high (Fig. 3) so the i n p u t i m p e d a n c e o f the cerebral vascular bed becomes t h a t of a low-pass filter, with the effect o f a t t e n u a t i n g a n d s m o o t h i n g the arterial pressure w a v e f o r m ; (b) I C P pulsatility is also affected by the v a s o m o t o r t o n e o f the cerebral vascular bed which, in its t u r n , strictly depends o n the action o f mech- a n i s m s r e g u l a t i n g the cerebral b l o o d flow a n d cerebral b l o o d v o l u m e . It is expected that those actions which cause a relaxation in vascular muscle t o n e a n d v a s o d i l a t i o n are associated with a n increase in c e r e b r o v a s c u l a r pulsatility, a n increase in M I C P a n d , therefore, also a n increase in I C P P A . H o w e v e r , a n increase in vascular t o n e is associated with a decrease in c e r e b r o v a s c u l a r pulsatility, a n M I C P decrease a n d a r e d u c t i o n in I C P P A . In p a r t i c u l a r , h y p e r c a p n i a has b e e n o b s e r v e d to increase the C S F pulse pressure, whereas h y p e r o x i a has the opposite effect (41,42).
A m o r e detailed m a t h e m a t i c a l d e s c r i p t i o n o f r e g u l a t o r y m e c h a n i s m s w o r k i n g o n the cerebral vascular bed, a n d o f their effect o n vascular s m o o t h muscle t o n e a n d i n t r a c r a n i a l d y n a m i c s will be the subject o f s u b s e q u e n t models.
T h e present m a t h e m a t i c a l m o d e l m a y c o n s t i t u t e a first step t o w a r d a m o r e c o m - plete, quantitative description of h u m a n h y d r o d y n a m i c a l p h e n o m e n a , especially valu- able in clinical investigation. T o this end, the capability o f the m o d e l in r e p r o d u c i n g the results o f some typical clinical tests, used in n e u r o l o g i c a l practice, will be accu- rately checked in a s u b s e q u e n t paper.
R E F E R E N C E S
1. Auer, L.M.; MacKenzie, E.T. Physiology of the cerebral venous system. In: Kapp, J.P; Schmidek, eds. The cerebral venous system and its disorders. Orlando: Grune & Stratton; 1984: pp. 169-227. 2. Avezaat, C.J.J.; van Eijndhoven, J.H.M. The conflict between CSF pulse pressure and volume- pressure response during plateau waves. In: Ishii, S.; Nagai, M; Brock, M., eds. Intracranial pres- sure V. Berlin, Heidelberg; Springer-Verlag; 1983: pp. 326-332.
3. Avezaat, C.J.J.; van Eijndhoven, J.H.M. The role of the pulsating pressure variations in intracranial pressure monitoring. Neurosurg. Rev. 9:113-120; 1986.
4. Avezaat, C.J.J.; van Eijndhoven, J.H.M.; and Wyper, D.J. Cerebrospinal fluid pulse pressure and intracranial volume-pressure relationship. J. Neurol. Neurosurg. and Psych. 42: 687-700; 1979. 5. Beneken, J.E.W.; DeWit, B. A physical approach to hemodynamic aspects of the human cardiovas-
cular system. In: Reeve, E.B.; Guyton, A.C., eds. Physical bases of circulatory transport: Regula- tion and exchange. Philadelphia: Saunders Co.: 1967: pp. 1-45.
6. Bergel, D.H. The static elastic properties of the arterial wall. J. Physiol. 156: 458-469; 1961. 7. Burattini, R.; Sipkema, P.; van Huis, G.A.; Westerhof, N.; Identification of canine coronary resis-
tance and intramyocardial compliance on the basis of the waterfull model. Ann. Biomed. Eng. 13: 385-404; 1985.
8. Chopp, M.; Portnoy, H.D. Starling resistor as a model of the cerebrovascular bed. In: Ishii, I.; Nagai, H; and Brock, M. eds. Intracranial Pressure V. Berlin, Heidelberg: Springer-Verlag; 1983: pp.174-179. 9. Cuypers, J.; Matakas, F.; Potolicchio, S.J. Effect of central venous pressure on brain tissue pressure
and brain volume. J. Neurosurg. 45: 89-94; 1976.
10. DereymakeT, A.; Stevens, A.; Rombouts, J.J.; Lacheron, J.M.; Pierquin, A. Study on the influence of the arterial pressure upon the morphology of cisternal CSF pulsations. Europ. Neurol. 5:107-114; 1971.
11. Ekstedt, J. CSF hydrodynamic studies in man. 2: Normal hydrodynamic variables related to CSF pres- sure and flow. J. Neurol. Neurosurg. and Psych. 41: 345-353; 1978.
12. Friden, H.; Ekstedt, J. The CSF volume-pressure relationship in man. In: Ishii, S.; Nagai, H.; and Brock, M. eds. Intracranial pressure V. Springer-Verlag; Heidelberg: Berlin, 1983: pp: 252-260. 13. Gaab, M.R.; Haubitz, I.; Brawanski, A.; Faulstich, J.; Heissler, H.E. Pressure-volume diagram, pulse
Human Intracranial Hydrodynamics 1 399
14. Gibo, H.; Carver, C.C.; Rhoton, A.L.; Lenkey, C.; Mitchell, R.J. Microsurgical anatomy of the mid- dle cerebral artery: A microsurgical study. J. Neurosurg. 54: 151-169; 1981.
15. Gibo, H.; Lenkey, C.; Rhoton, A.L. Microsurgical anatomy of the supraclinoid portion of the internal carotid artery. J. Neurosurg. 55: 560-574; 1981.
16. Guess, H.A.; Charlton, J.D.; Johnson, R.N.; Mann, J.D. A nonlinear least square method for deter- mining cerebrospinal fluid formation and absorption kinetics in pseudotumor cerebri. Comp. & Biomed. Res. 18: 184-192; 1985.
17. Hayashi, K.; Handa, H.; Nagasawa, S.; Okumura, A.; Moritake, K. Stiffness and elastic behavior of human intracranial and extracranial arteries. J. Biomech. 13: 175-184; 1980.
18. Heistad, D.D.; Kontos, H.A. Cerebral circulation. In: Shepard, J.T.; Abboud, F.M. eds. Handbook of physiology: Peripheral circulation and organ blood flow, Sec. 2, Vol. III; American Physiologi- cal Society, Bethesda, MD 1983; pp. 137-182.
19. Higashi, K.; Katano, M.; lhara, K.; Katayama, S. Cerebral blood flow and production of cerebrospi- nal fluid. In: Gotoh, F.; Nagai, H.; Tazaki, Y. eds. Cerebral blood flow and metabolism. Acta Neurol. Sca., Suppl. 72; 1979; pp. 598-599.
20. Hoffman, O. CSF dynamics: Integration of pulsatory components and autoregulation into a math- ematical model. In: Ishii, S.; Nagai, H.; and Brock, M. eds. Intracranial Pressure V. Berlin, Heidel- berg, Springer-Verlag; 1983; 169-173.
21. Katzman, R.; Hussey, F. A simple constant infusion manometric test for measurement of CSF absorp- tion. I: Rationale and method. Neurology (Minneapolis) 20: 534-544; 1970.
22. Kety, S.S.; Schmidt, C.F. The effect of altered arterial tension of carbon dioxide and oxygen on cere- bral blood flow and cerebral oxygen consumption of normal young man. J. Clinic. Invest. 27: 484-492; 1948.
23. Kontos, H.A.; Wei, E.P.; Novari, R.M.; Levasseur, J.E.; Rosemblum, W.I.; Patterson, J.L. Responses of cerebral arteries and arterioles to acute hypotension and hypertension, Am. J. Physiol. 234 (4): H371-H383; 1978.
24. Mann, J.D.; Butler, A.B.; Rosenthal, J.E.; Maffeo, C.J.; Johnson, R.N.; Bass, N.H. Regulation of intracranial pressure in rat, dog and man. Ann. Nerol. 3: 156-165; 1978.
25. Marmarou, A.; Schulman, K.; LaMorgese, J. Compartmental analysis of compliance and outflow resistance of the cerebrospinal fluid system. J. Neurosurg. 43: 523-534; 1975.
26. Marmarou, A.; Schulman, K.; Rosende, R.M. A nonlinear analysis of the cerebrospinal fluid system and intracranial pressure dynamics. J. Neurosurg. 48: 332-344; 1978.
27. Martins, A.N. Resistance to drainage of cerebrospinal fluid: Clinical measurement and significance. J. Neurol. Neurosurg. Psych. 36: 313-318; 1973.
28. Matakas, F. The interrelationship between cerebral microcirculation and intracranial pressure. In: Baan, J.; Noordergraaf, A.; and Raines, J. eds. Cardiovascular system dynamics. Cambridge, MA: MIT press; 1978; pp. 224-230.
29. Mchedlishvili, G.I. Physiological mechanisms controlling cerebral blood flow. Stroke 11: 240-248;
1980.
30. Miller, J.D.; Stanek, A.; Langfitt, T.W. Concepts of cerebral perfusion pressure and vascular com- pression during intracranial hypertension. In: Meyer, J.S.; and Schade, J.P. eds. Progress in brain research: Cerebral blood flow. Amsterdam: Elsevier; 1972; pp. 411-432.
31. Milnor, W.R. Hemodynamics. Baltimore: William & Wilkins; 1982.
32. Nakagawa, Y.; Tsuru, M.; Yada, K. Site and mechanism for compression of the venous system during experimental intracranial hypertension. J. Neurosurg. 41: 427-434; 1974.
33. Nornes, H.; Aslid, R.; Lindegaard, K.F. Intracranial pulse pressure dynamics in patients with intra- cranial hypertension. Acta Neurochirurgica 38: 177-186; 1977.
34. Nyary, I.; Vajda, J. Relationship of cerebral blood volume changes and estimated intracranial com- pliance. In: Ishii, S.; Nagai, H.; and Brock, M. eds. Intracranial Pressure V, Berlin, Heidelberg; Springer-Verlag; 1983; pp. 316-319.
35. Paltsev, E.[.; Sirovsky, E.B. Intracranial physiology and biomechanics. J. Neurosurg. 57: 500-510; 1982.
36. Pedley, T.J. The fluid mechanics of large blood vessels. London: Cambridge University Press; 1980. 37. Perlmutter, D.; Rhoton, A.L. Microsurgical anatomy of the anterior cerebral-anterior communicating-
recurrent artery complex. J. Neurosurg. 45: 259-272; 1976.
39. Permutt, S.; Riley, R.L. Hemodynamics of collapsible vessels with tone; The vascular waterfull. J. Appl. Physiol. 18: 924-932; 1963.
40. Pollay, M. Formation of cerebrospinal fluid. In: Lundberg, N.; Ponten, U.; and Brock, M. eds. Intracranial Pressure II, Berlin Heidelberg: Springer-Verlag; 1975; 42-45.
41. Pormoy, H.D.; Chopp, M.; Branch, C.; Shannon, M. CSF pulse wave, ICP and autoregulation. In: Ishii, S.; Nagai, H.; and Brock, M. eds. Intracrania Pressure V, Berlin, Heidelberg: Springer-Verlag; 1983; pp. 180-185.
42. Portnoy, H.D.; Chopp, M.; Branch, C.; Shannon, M. Cerebrospinal fluid pulse waveform as an indi- cator of cerebral autoregulation. J. Neurosurg. 56: 666-678; 1982.
43. Saeki, N.; Rhoton, A.L. Microsurgical anatomy of the upper basilar artery and the posterior circle of Willis. J. Neurosurg. 46: 563-578; 1977.
44. Sahar, A. The effect of pressure on the production of cerebrospinal fluid by the choroid plexus. J. Neurol. Sci. 16: 49-58; 1972.
45. Shapiro, K.; Marmarou, A., Shulman, K. Characterization of clinical CSF dynamics and neural axis compliance using the pressure volume index: The normal pressure volume index. Ann. Neurol. 7: 508-514; 1980.
46. Westerhof, N.; Boosman, F.; DeVries, C.J.; Noordergraff, A. Analog studies of the human systemic arterial tree. J. Biomech. 2: 121-143; 1969.
47. Yada, K.; Nakagawa, Y.; Tsuru, M. Circulatory disturbances of the venous system during experimental intracranial hypertension. J. Neurosurg. 39: 723-729; 1973.
N O M E N C L A T U R E
Cai = i n t r a c r a n i a l a r t e r i a l c o m p l i a n c e Cti~ = c e r e b r a l t i s s u e c o m p l i a n c e
Cve = e x t r a c r a n i a l v e n o u s c o m p l i a n c e C~i = i n t r a c r a n i a l v e n o u s c o m p l i a n c e
d/h = d i a m e t e r t h i c k n e s s r a t i o E = v e s s e l Y o u n g ' s m o d u l u s Etiss = c e r e b r a l t i s s u e e l a s t a n c e h = v e s s e l t h i c k n e s s
Ka = e l a s t a n c e c o e f f i c i e n t o f t h e i n t r a c r a n i a l a r t e r i a l c o m p a r t m e n t
Ke = c e r e b r a l t i s s u e e l a s t a n c e c o e f f i c i e n t
Kv = e l a s t a n c e c o e f f i c i e n t o f t h e i n t r a c r a n i a l v e n o u s c o m p a r t m e n t l = v e s s e l l e n g t h
I,- = s a l i n e i n j e c t i o n r a t e P a = c e r e b r a l a r t e r i a l p r e s s u r e Pc = c e r e b r a l c a p i l l a r y p r e s s u r e /:'co = c e n t r a l v e n o u s p r e s s u r e
Pic = i n t r a c r a n i a l p r e s s u r e P~ = c e r e b r a l v e n o u s p r e s s u r e P~s = v e n o u s s i n u s p r e s s u r e
Pol = c o l l a p s e t r a n s m u r a l p r e s s u r e o f t h e c e r e b r a l v e n o u s v a s c u l a r b e d Poi = c o e f f i c i e n t o f t h e q u a d r a t i c t e r m in c e r e b r a l t i s s u e e l a s t a n c e q = c e r e b r a l b l o o d f l o w
qo = C S F a b s o r p t i o n r a t e
qy = C S F f o r m a t i o n r a t e r = v e s s e l r a d i u s
Rai = i n t r a c r a n i a l a r t e r i a l r e s i s t a n c e
R s = C S F f o r m a t i o n r e s i s t a n c e
Human Intracranial Hydrodynamics 1 401
R p u ~- R us
R r =
US
R " US -~- R u e
/min ~--- /max =
T =
O" v~ =
VCS F =
Vtiss
v~ =
A P i c = x
resistance o f the proximal venous vascular bed resistance o f the distal cerebral venous vascular bed
resistance of the distal cerebral venous vascular bed upstream o f the collapse resistance o f the distal cerebral venous vascular bed downstream o f the collapse
extracranial venous resistance
instant o f m i n i m u m intracranial pressure instant o f m a x i m u m intracranial pressure cardiac period
Poisson's ratio
intracranial arterial blood volume cerebrospinal fluid volume cerebral tissue volume intracranial venous volume
intracranial pressure pulse amplitude autoregulatory action