VARIASI MORFOLOGI DAUN JENIS PIONIR PULAI
(
Alstonia scholaris
R. Br.) DAN MACARANGA (
Macaranga
triloba
(Bl.) Muell. Arg.) DI HUTAN KARET JAMBI
INGGAR DAMAYANTI
DEPARTEMEN SILVIKULTUR FAKULTAS KEHUTANAN INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Variasi Morfologi Daun Jenis Pionir Pulai (Alstonia scholaris R. Br.) dan Macaranga (Macaranga triloba (Bl.) Muell. Arg.) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
INGGAR DAMAYANTI. Variasi Morfologi Daun Jenis Pionir Pulai (Alstonia scholaris R. Br.) dan Macaranga (Macaranga triloba (Bl.) Muell. Arg.) di Hutan Karet Jambi. Dibimbing oleh ISKANDAR Z. SIREGAR dan SRI RAHAYU.
Aktivitas manusia telah mengakibatkan pengurangan luasan hutan alami dan memicu transformasinya menjadi hutan karet (jungle rubber) seperti yang banyak dijumpai di Jambi. Pembentukan hutan karet melalui proses suksesi melibatkan pohon pionir penting seperti Alstonia scholaris dan Macaranga triloba yang keragaannya dipengaruhi oleh faktor tempat tumbuh, biodiversitas awal dan input perlakuan budidaya. Penelitian dilakukan untuk mengkaji variasi morfologi daun A. scholaris dan M. triloba sebagai informasi awal adanya perbedaan respon dan keragaan tumbuh pada dua lansekap yang berbeda yaitu di Taman Nasional Bukit Dua Belas dan Hutan Harapan. Selain itu variasi A. scholaris dan M. triloba juga dilihat berdasarkan keragaman genetiknya. Hasil sidik ragam menunjukkan bahwa perbedaan lansekap memberikan pengaruh nyata pada variabel morfologi daun NV (number of veination) untuk A. scholaris dan tidak memberikan pengaruh nyata terhadap semua variabel untuk M. triloba. Perbedaan plot memberikan pengaruh nyata pada variabel number of veination (NV) dan petiole ratio (PR) untuk A. scholaris dan variabel lamina length (LL), petiole length (PL), lobe width (LW), widest leaf to petiole (WP), number of veination (NV), lamina shape (LS), dan lobe width ratio (LWR) untuk M. triloba. Hasil analisis gerombol (Cluster Analysis) menunjukkan bahwa plot BJ2 pada A. scholaris digolongkan outgroup, sementara pada M. triloba satu plot yang outgroup yaitu plot BJ310. Hasil analisis sebaran variasi morfologi daun A.
scholaris dan M. triloba dari tiga analisis multivariat yang digunakan tidak menunjukkan pengelompokan berdasarkan perbedaan lansekap.
ABSTRACT
INGGAR DAMAYANTI. Leaf Morphological Variation of Pioner Trees Pulai (Alstonia scholaris R. Br.) and Macaranga (Macaranga triloba (Bl.) Muell. Arg.) in Jungle Rubber, Jambi. Supervised by ISKANDAR Z. SIREGAR and SRI RAHAYU.
Human activities have resulted of a reduction in natural forests and triggered their transformation into, for example jungle rubbers as often found in Jambi. The formation of jungle rubbers through successional processes involves important pioneer trees such as Alstonia scholaris and Macaranga triloba in which their performances are influenced by the site factors, the initial status of biodiversity, and inputs of cultivation treatments and management. The study was conducted to assess the leaf morphological variation of A. scholaris and M. triloba as the initial information in determining differences in response and growth performance in several CRC990 plots (50 m x 50 m) of the two distinct landscapes, namely: Bukit Dua Belas National Park and Harapan Rain Forest. In addition, the variation of A. scholaris and M. triloba also can be observed based on its genetic diversity. Results of analysis of variance showed that the differences in landscapes gave significant effects on one leaf morphology variable, i.e NV (number of veination) for A. scholaris, while no significant effect on all variables was observed for M. triloba. The plots gave significant effect on the number of veination (NV) and petiole ratio (PR) variables for A. scholaris and lamina length (LL), petiole length (PL), lobe width (LW), widest leaf to petiole (WP), number of veination (NV), lamina shape (LS), and lobe width ratio(LWR) variables for M. triloba. The results of Cluster Analysis indicated that in A. scholaris BJ2 plots were classified as outgroup, while in M. triloba the outgroup plot was BJ310 plot. The results of the distribution of leaf morphological variation
of A. scholaris and M. triloba based on three multivariate analyses showed no clustering due to the landscapes effects..
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
pada
Departemen Silvikultur
VARIASI MORFOLOGI DAUN JENIS PIONIR PULAI
(
Alstonia scholaris
R. Br.) DAN MACARANGA (
Macaranga
triloba
(Bl.) Muell. Arg.) DI HUTAN KARET JAMBI
DEPARTEMEN SILVIKULTUR FAKULTAS KEHUTANAN INSTITUT PERTANIAN BOGOR
Judul Skripsi : Variasi Morfologi Daun Jenis Pionir Pulai (Alstonia scholaris R. Br.) dan Macaranga (Macaranga triloba (Bl.) Muell. Arg.) di Hutan Karet Jambi
Nama : Inggar Damayanti NIM : E44100009
Disetujui oleh
Prof Dr Ir Iskandar Z Siregar, MForSc Pembimbing I
Dr Ir Sri Rahayu, MSi Pembimbing II
Diketahui oleh
Prof Dr Ir Nurheni Wijayanto, MS Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Februari 2014 ini ialah biodiversitas, dengan judul Variasi Morfologi Daun Jenis Pionir Pulai (Alstonia scholaris R. Br.) dan Macaranga (Macaranga triloba (Bl.) Muell. Arg.) di Hutan Karet Jambi.
Terima kasih penulis ucapkan kepada Bapak Prof Dr Ir Iskandar Z. Siregar, MForSc dan Ibu Dr Ir Sri Rahayu, MSi selaku dosen pembimbing, yang telah banyak memberi bimbingan dan saran sehingga tugas akhir ini dapat selesai, kepada Ms. Natalie Breidenbach atas kesediannya meminjamkan herbarium untuk proses pengambilan data. Ungkapan terima kasih juga disampaikan kepada ayah (Alm. Muhammad Toha), ibu (Summayah), adik (Anggi Rahmawati) serta seluruh keluarga, atas segala doa dan kasih sayangnya. Di samping itu penulis juga mengucapkan terimakasih kepada kakanda Laswi Irmayanti SHut yang telah memberikan bantuan dan arahan dalam penelitian ini. Terimakasih juga penulis sampaikan kepada rekan-rekan, Gumilar Adam, Lastiti, Fitria Nurmala, Rummi, Ayi, Dewi, Arie, Uci, Aji serta seluruh teman-teman Silvikultur 47 dan semua pihak yang tidak bisa disebutkan satu persatu yang telah membantu penulis dalam penelitian ini.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Manfaat Penelitian 2
Ruang Lingkup Penelitian 2
METODE PENELITIAN 3
Waktu dan Tempat Penelitian 3
Bahan 4
Alat 4
Prosedur Penelitian 5
Pengolahan Data 6
HASIL DAN PEMBAHASAN 7
Karakteristik Daun Jenis A. scholaris dan M. triloba 7 Pengaruh Perbedaan Lansekap dan Plot Pengambilan Sampel Terhadap Variasi
Morfologi Daun A. scholaris dan M. triloba 9
Variasi Morfologi Daun A. scholaris dan M. triloba Di Hutan Karet Harapan
dan Hutan Karet Bukit Duabelas 10
Sebaran Variasi Morfologi Daun A. scholaris dan M. triloba Terhadap
Perbedaan Lansekap (HJ dan BJ) 12
Keragaman Sekuen DNA A. scholaris dan M. triloba 17
SIMPULAN DAN SARAN 20
Simpulan 20
Saran 20
DAFTAR PUSTAKA 20
RIWAYAT HIDUP 23
LAMPIRAN 13
DAFTAR TABEL
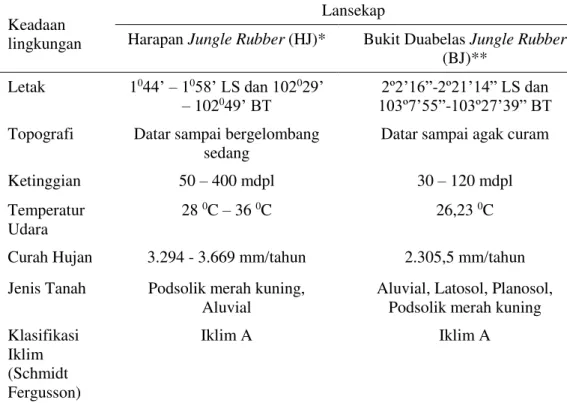
1 Keadaan lingkungan lansekap penelitian (HJ dan BJ) 3
2 Data herbarium 4
3 Karakteristik daun A. scholaris 7
4 Karakteristik daun M. triloba 8
5 Korelasi antara LL dengan variabel lain untuk daun A. scholaris dan
M. triloba 8
6 Hasil sidik ragam pengaruh lansekap dan plot terhadap variasi
morfologi daun A. scholaris dan M. triloba 10
7 Proporsi dari total varian yang dijelaskan oleh variabel sintesis pertama dan kedua dari analisis multivariat yang digunakan (CDA;PCA;MCA) pada A. scholaris dan M. triloba 12 8 Test of quality of group means A. scholaris 13 9 Test of quality of group means M. triloba 13 10 Korelasi seluruh variabel morfologi daun A. scholaris dan M. triloba
terhadap variabel sintesis 1 dan 2 dengan metode PCA 14 11 Korelasi semua variabel morfologi daun M. triloba dan A. scholaris
terhadap variabel sintesis 1 dan 2 dengan metode MCA 15 12 Hasil data mining untuk A. scholaris dan M. triloba (GenBank 2014) 17 13 Komposisi nukleotida dari 6 individu A. scholaris region matK 18 14 Jarak genetik nukleotida dan jarak lokasi pengambilan sampel pada A.
scholaris region matK 19
DAFTAR GAMBAR
1 Karakter morfologi variabel pengukuran A. scholaris (1) dan M.
triloba (2) 5
2 Daun A. scholaris (1), M. triloba (2) 7
3 Dendogram hasil CA untuk A. scholaris (skala menggunakan standar
deviasi) 11
4 Dendogram hasil CA untuk M. triloba (skala menggunakan standar
deviasi) 11
5 Distribusi dari variabel sintesis pertama dan kedua yang terbentuk dari dua analisis multivariat yang berbeda pada A. scholaris. (A) dan (C) adalah distribusi daun pada dua variabel sintesis yang terbentuk (variabel sintesis pertama sebagai X, variabel sintesis kedua sebagai Y); (B) dan (D) menjelaskan distribusi dari variabel sintesis pertama;
(A) dan (B): PCA;(C) dan (D): MCA. 15
6 Distribusi dari variabel sintesis pertama dan kedua yang terbentuk dari dua analisis multivariat yang berbeda pada M. triloba. (A) dan (C) adalah distribusi daun pada dua variabel sintesis yang terbentuk (variabel sintesis pertama sebagai X, variabel sintesis kedua sebagai Y); (B) dan (D) menjelaskan distribusi dari variabel sintesis pertama;
(A) dan (B): PCA; (C) dan (D): MCA. 16
PENDAHULUAN
Latar Belakang
Hutan dataran rendah Jambi merupakan hutan hujan tropika yang masih tersisa di Indonesia. Hutan hujan tropis memiliki keanekaragaman hayati yang sangat tinggi (Odum 1993), yang menjadi karakteristik khas dari hutan hujan tropis (Steege dan Hammond, 2001). Luas hutan hujan tropis di dunia hanya meliputi tujuh persen dari luas permukaan bumi, tetapi mengandung lebih dari 50 persen total jenis yang ada di seluruh dunia (Irwanto 2007). Ekosistem hutan hujan tropis dan seluruh keanekaragaman hayati di dalamnya memiliki nilai penting bagi kehidupan manusia dan lingkungan.
Kekayaan dan potensi ekonomi dari hutan hujan tropis seringkali menjadi ancaman yang dapat mengurangi luasan hutan. Ancaman tersebut berupa gangguan (disturbance) yang didefinisikan oleh Smith (1990) sebagai suatu aktivitas atau energi dari luar yang dapat memengaruhi ekosistem, komunitas, populasi, tanah, dan keanekaragaman hayati yang tersedia, dan dapat memfasilitasi masuknya jenis-jenis baru. Hutan dataran rendah primer di Indonesia telah lama terancam oleh deforestasi dan degradasi hutan yang menyebabkan hilangnya keanekaragaman hayati akibat penghancuran habitat hutan tropis yang unik (Margono 2014). Aktifitas manusia telah mengakibatkan pengurangan luasan hutan alami dan memunculkan beragam tipe lahan di Jambi, yang dapat didefinisikan sebagai sistem transformasi hutan (CRC 2012). Hasil dari pemetaan lansekap yang dilakukan oleh Dewi et al. (2008), terdapat 12 tipe lahan yang terbentuk di Jambi, salah satunya adalah hutan karet (Jungle rubber).
Terbentuknya hutan karet salah satunya diakibatkan oleh sistem yang telah sangat lama digunakan oleh para petani yaitu sistem perladangan berpindah (swidden agriculture). Hutan karet terbentuk karena masa pemberaan lahan (Ningsih 2009). Masa pemberaan lahan memberikan variasi umur pada setiap lahan. Umur lahan mempengaruhi proses perubahan alami dan terarah yang teramati dari komposisi vegetasi, yang dikenal dengan istilah suksesi (Barbour et al. 1999). Beberapa penelitian menunjukkan bahwa bila masa bera berlangsung cukup lama, maka struktur komunitas dan komposisi vegetasi yang terbentuk bisa mendekati stuktur dan komposisi hutan alami.
2
lingkungan ini tentunya akan menunjukkan pola keragaman baik secara fenotipe maupun genetiknya yang membentuk tingkat keragaman biodiversitas. Oleh karena itu perlu diamati sejauh mana perubahan penggunaan lahan di Jambi mempengaruhi pola platisitas tersebut.
Keragaman fenotipe suatu makhluk hidup ditentukan oleh faktor genetika dan lingkungan. Sifat fenotipe dapat diamati secara kasat mata baik bentuk, warna maupun ukuran. Dalam penelitian ini akan dikaji lebih dalam mengenai perbedaan fenotipe antara jenis Alstonia scholaris dan Macaranga triloba yang tumbuh pada dua lansekap (bentang lahan) yang berbeda ditinjau dari morfologi daun. Indentifikasi morfologi daun dipilih karena mudah diamati, serta merupakan oragan vital bagi keberlangsungan hidup tumbuhan. Selain dilihat dari sisi perbedaan fenotipe, variasi Alstonia scholaris dan Macaranga triloba juga dilihat berdasarkan keragaman nukleotidanya berdasarkan sekuen DNA Barcode atau marka lainnya. Gambaran variasi morfologi daun dari A. scholaris dan M.triloba diharapkan dapat memberikan informasi sejauh mana respon morfologi daun kedua jenis tersebut terhadap lingkungan.
Perumusan Masalah
Berdasarkan uraian tersebut di atas, maka penulis membuat rumusan penelitian sebagai berikut:
1. Adakah perbedaan respon morfologi yang ditunjukkan oleh karakter morfologi daun antara kedua jenis di dua lansekap yang berbeda?
2. Bagaimana status keragaman A. scholaris dan M. triloba berdasarkan DNA barcode yang diperoleh dari pangkalan data molekuler?
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk:
1. Memberikan informasi mengenai perbedaan morfologi A. scholaris dan M. triloba yang tumbuh pada dua lansekap yang berbeda.
2. Mengetahui keragaman genetik A. scholaris dan M. triloba berdasarkan DNA barcode dari bank data DNA.
Manfaat Penelitian
Manfaat yang diharapkan dari penelitian ini adalah untuk:
1. Memberikan penjelasan mengenai respon tumbuh A. scholaris dan M. triloba pada dua lansekap yang berbeda berdasarkan karakter morfologi daun yang diamati.
2. Mengetahui keragaman genetik A. scholaris dan M. triloba berdasarkan DNA barcode.
Ruang Lingkup Penelitian
3 scholaris dan M. triloba berdasarkan kesamaan sekuen DNA barcode dari data sekunder. Variabel pengukuran yang digunakan adalah panjang daun, panjang petiole, lebar daun, jarak daun terlebar ke petiole, besar sudut pertulangan daun, dan jumlah tulang daun (Kremer et al. 2001). Variasi di dalam dan antar lansekap dari morfologi daun dijelaskan melalui analisis statistik multivariat. Analisis DNA mengenai hubungan keragaman genetik dijelaskan melalui keragaman nukleotidanya.
METODE PENELITIAN
Waktu dan Tempat Penelitian
Penelitian dilakukan di Laboratorium Genetika Hutan, Fakultas Kehutanan, Institut Pertanian Bogor yang dilakukan selama 6 bulan dari bulan Januari sampai dengan Juni 2014. Sampel daun yang digunakan berasal dari dari sub- proyek B03 CRC (Collaborative Research Center) 990/EFForTS (http://www.uni-goettingen.do/efforts.com) yang dilakukan di dua lansekap yaitu hutan karet hutan Harapan/Harapan Jungle Rubber (HJ) dan hutan karet Taman Nasional Bukit Duabelas/Bukit Duabelas Jungle Rubber (BJ) di Jambi. Kondisi lingkungan dari kedua lansekap disajikan pada Tabel 1.
Tabel 1 Keadaan lingkungan lansekap penelitian (HJ dan BJ)
Keadaan lingkungan
Lansekap
Harapan Jungle Rubber (HJ)* Bukit Duabelas Jungle Rubber
(BJ)**
Letak 1044’ – 1058’ LS dan 102029’
– 102049’ BT
2º2’16”-2º21’14” LS dan
103º7’55”-103º27’39” BT Topografi Datar sampai bergelombang
sedang
Datar sampai agak curam
Ketinggian 50 – 400 mdpl 30 – 120 mdpl
Temperatur Udara
28 0C – 36 0C 26,23 0C
Curah Hujan 3.294 - 3.669 mm/tahun 2.305,5 mm/tahun
Jenis Tanah Podsolik merah kuning, Aluvial
Aluvial, Latosol, Planosol, Podsolik merah kuning
Klasifikasi Iklim (Schmidt Fergusson)
Iklim A Iklim A
4
Bahan
Contoh daun yang berasal dari dua lansekap tersebut di atas berupa herbarium/voucher A. scholaris dan M. triloba yang sudah dewasa/berkembang penuh (full developed) yang diambil secara acak. Voucher yang digunakan secara lengkap tersaji dalam Tabel 2.
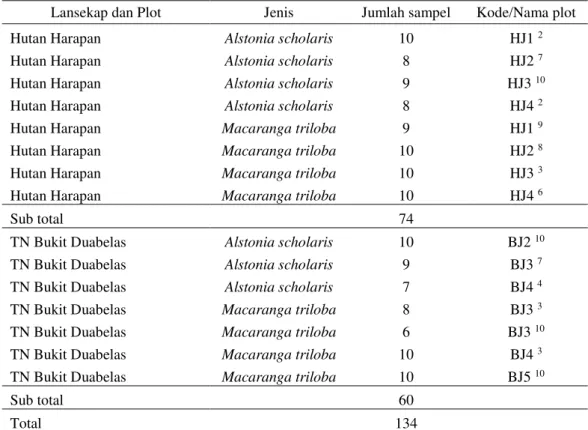
Tabel 2 Data herbarium
Lansekap dan Plot Jenis Jumlah sampel Kode/Nama plot
Hutan Harapan Alstonia scholaris 10 HJ1 2
Hutan Harapan Alstonia scholaris 8 HJ2 7
Hutan Harapan Alstonia scholaris 9 HJ3 10
Hutan Harapan Alstonia scholaris 8 HJ4 2
Hutan Harapan Macaranga triloba 9 HJ1 9
Hutan Harapan Macaranga triloba 10 HJ2 8
Hutan Harapan Macaranga triloba 10 HJ3 3
Hutan Harapan Macaranga triloba 10 HJ4 6
Sub total 74
TN Bukit Duabelas Alstonia scholaris 10 BJ2 10
TN Bukit Duabelas Alstonia scholaris 9 BJ3 7
TN Bukit Duabelas Alstonia scholaris 7 BJ4 4
TN Bukit Duabelas Macaranga triloba 8 BJ3 3
TN Bukit Duabelas Macaranga triloba 6 BJ3 10
TN Bukit Duabelas Macaranga triloba 10 BJ4 3
TN Bukit Duabelas Macaranga triloba 10 BJ5 10
Sub total 60
Total 134
Ket: HJ= Harapan Jungle rubber; BJ= Bukit Dua Belas Jungle rubber; 1= nomor plot; 2= urutan
pohon dominan dalam plot
Alat
5
Prosedur Penelitian
Identifikasi morfologi daun
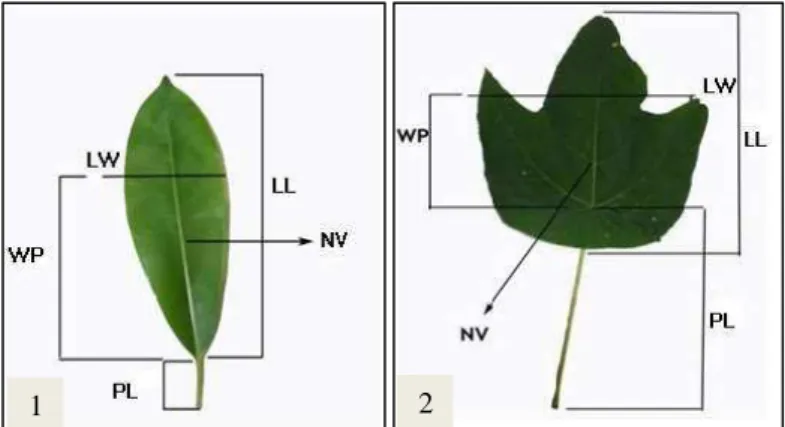
Metode penilaian morfologi daun merujuk pada Kremer et al. (2001) dengan beberapa modifikasi yang bertujuan untuk menyederhanakan prosedur. Adapun variabel yang diukur dan diamati untuk setiap daun dibagi menjadi 3 penilaian yaitu:
1. Empat karakter dimensional
Panjang Daun (LL), Panjang Petiole (PL), Lebar Daun (LW), jarak daun terlebar ke petiole (WP).
2. Variabel yang dihitung
Jumlah tulang daun (NV). Tulang daun yang dihitung dalam penelitian ini adalah tulang daun primer dan sekunder yang terlihat jelas pada daun.
Gambar 1 Karakter morfologi variabel pengukuran A. scholaris (1) dan M. triloba (2)
3. Variabel yang dikonversi
Lamina shape (LS) = 100 ×WP / LL
Petiole ratio (PR) = 100 × PL / (LL + PL) Lobe width ratio (LWR) = 100 × LW / LL
Rancangan Percobaan
Rancangan percobaan yang digunakan dalam penelitian ini adalah rancangan tersarang (Nested Design) dengan dua faktor yaitu lansekap dan plot. Data yang diperoleh berdasarkan pengukuran kemudian dianalisis menggunakan model linier:
� = � + � + + �
dimana:
Yijk = Nilai pengamatan level ke-j yang bersarang dalam level ke-i pada ulangan ke-k
µ = Nilai rataan umum
αi = Pengaruh faktor A (lansekap) pada level ke-i
j(i) = Prngaruh faktor B (plot) pada level ke-j yang bersarang pada faktor A (lansekap) level ke-i
� (ij)k = Nilai galat akibat level ke-j yang bersarang pada level ke-i pada ulangan ke-k
i = Level-level faktor A (lansekap)
6
j = Level-level faktor B (plot) yang bersarang di tiap level A k = Jumlah ulangan
Analisis DNA
Pengambilan data penelitian pada bagian ini merupakan data sekunder. Data yang diambil berupa sekuens DNA barcode A. scholaris pada region matK dan M. triloba pada region rbcL. Metode pengambilan data sekunder yang dilakukan pada penelitian ini adalah data mining. Pengumpulan data sekuens DNA menggunakan metode data mining tidak dilakukan melalui prosedur kerja pada laboratorium. Data ini diambil dari database GenBank dalam situs National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.g-ov/genbank/) untuk A. scholaris pada region matK dan M. triloba pada region rbcL.
Pengolahan Data
Analisis data morfologi daun
1. Pengaruh Perbedaan Lansekap dan Plot Terhadap Variasi Morfologi Daun Pengaruh perbedaan lansekap terhadap variasi morfologi daun dianalisis menggunakan metode Analysis of Variance (ANNOVA). Data-data karakteristik morfologi daun pada perbedaan lansekap diolah dengan software Ms.Excel dan SAS 9.1 portable.
2. Variasi Morfologi Daun A. scholaris dan M. triloba di HJ dan BJ.
Variasi morfologi daun A. scholaris dan M. triloba dianalisis dengan Cluster Analisis (CA) dan dilakukan dengan software SPSS 13.0. Hasil dari CA akan ditampilkan dalam bentuk dendogram. Cluster Analysis (CA) digunakan untuk mengelompokkan sampel A. scholaris dan M. triloba berdasarkan variasi morfologi daunnya. CA akan mengelompokkan obyek yang memiliki kemiripan ke dalam satu kelompok (group) (Henderson 2006). 3. Sebaran Variasi morfologi A. scholaris dan M. triloba terhadap perbedaan
lansekap (HJ dan BJ).
7
Analisis sekuens DNA
Sekuens DNA A. scholaris dan M. triloba dirunutkan dengan menggunakan program perangkat lunak Clustal X2 (http://www.clustal.org), BioEdit (http://www.mbio.ncsu.edu/bioedit.html), dan TreeViewX (http://www.treeview.n et/tv/download.asp). Sekuens DNA yang telah dirunutkan kemudian dianalisis keragaman basa nukleotidanya menggunakan software MEGA6.0 (http://www.megasoftware.net).
HASIL DAN PEMBAHASAN
Karakteristik Daun Jenis A. scholaris dan M. triloba
Daun termasuk organ pokok pada tubuh tumbuhan. Pada umumnya tiap tumbuhan mempunyai sejumlah besar daun. Daun A. scholaris tersusun melingkar dari 4-8 di axils atas, lamina atau badan daun berbentuk obovate hingga elips atau elips-lanset, berbulu gundul atau jarang berbulu. Daun meruncing ke arah dasar. Permukaan atas daun berwarna hijau tua, ujung daun bulat meruncing ke arah basis. Daun M. triloba memiliki petiole silinder, daun berbentuk pisau bulat telur ditutupi oleh rambut-rambut tegak berwarna keperakan. Daun berwarna hijau dan memiliki 3 lobus. Karakteristik daun M. triloba adalah lebar dibagian bawahnya dan menyempit bertahap hingga ujung daun. Keragaan daun A. scholaris dan M. triloba disajikan pada Gambar 2.
Gambar 2 Daun A. scholaris (1), M. triloba (2)
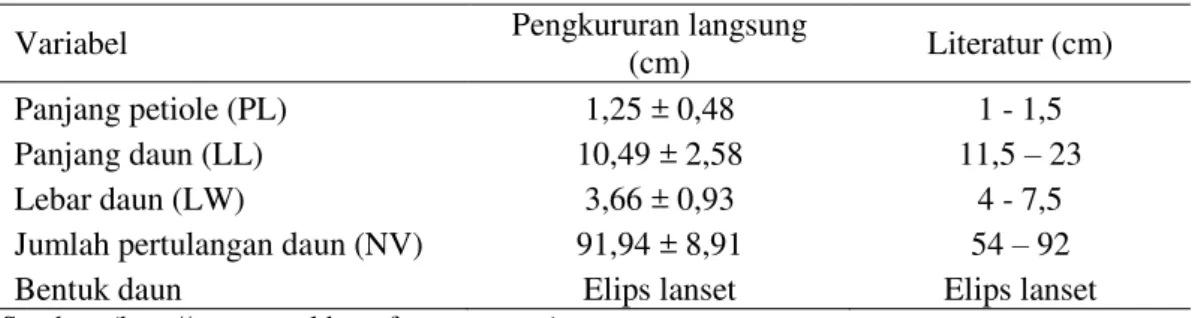
Tabel 3 Karakteristik daun A. scholaris Variabel Pengkururan langsung
(cm) Literatur (cm) Panjang petiole (PL) 1,25 ± 0,48 1 - 1,5 Panjang daun (LL) 10,49 ± 2,58 11,5 – 23 Lebar daun (LW) 3,66 ± 0,93 4 - 7,5 Jumlah pertulangan daun (NV) 91,94 ± 8,91 54 – 92 Bentuk daun Elips lanset Elips lanset
Sumber: (http://www.worldagroforesstry.com)
8
Tabel 4 Karakteristik daun M. triloba Variabel Pengkururan langsung
(cm) Literatur (cm) Panjang petiole (PL) 18,51±7,66 10 – 25 Panjang daun (LL) 22,60 ± 8,06 15 – 35 Lebar daun (LW 20,68 ± 7,25 12 – 24
Jumlah lobus 3 3
Bentuk daun Pisau bulat telur Pisau bulat telur
Sumber: Norfaizal et al. (2012); (http://www.worldagroforesstry.com)
Tabel 3 dan 4 menyajikan karakteristik daun A. scholaris dan M. triloba berdasarkan beberapa karakter morfologinya berdasarkan literatur dan pengukuran langsung. Data pada kedua tabel menunjukkan kesesuaian antara literatur dan pengukuran langsung beberapa karakter morfologi daun A. scholaris dan M. triloba.
Data karakteristik morfologi daun yang diambil berasal dari tegakan-tegakan A. scholaris dan M. triloba yang luas, sehingga diduga bahwa variabel pengukuran mungkin dipengaruhi oleh ukuran daun (Kramer et al. 2001). Oleh karena itu, korelasi antara panjang daun (LL) dan semua variabel lain yang diukur dikomputasi atas seluruh kumpulan data A. scholaris dan M. triloba.
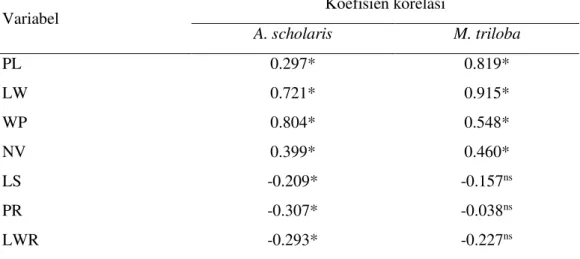
Tabel 5 Korelasi antara LL dengan variabel lain untuk daun A. scholaris dan M. triloba
Variabel Koefisien korelasi
A. scholaris M. triloba
PL 0.297* 0.819*
LW 0.721* 0.915*
WP 0.804* 0.548*
NV 0.399* 0.460*
LS -0.209* -0.157ns
PR -0.307* -0.038ns
LWR -0.293* -0.227ns
Ket: *= signifikan pada α=5%; ns= tidak signifikan pada α=5%
9 jenis, pada M. triloba 3 variabel transformasi memiliki korelasi yang tidak signifikan terhadap LL, sementara 3 variabel transformasi pada A. scholaris menunjukkan korelasi yang signifikan.
Variabel ciri-ciri dimensional pada A. scholaris yang memiliki korelasi paling kuat diantara yang lainnya adalah LW dan WP, dengan nilai masing-masing 0,721 dan 0,804. Variabel dimensional lainnya seperti PL dan NV memiliki korelasi yang lemah dengan LL. Artinya perubahan nilai PL dan NV tidak cukup kuat untuk mempengaruhi perubahan nilai LL. Hal ini juga terlihat dari pengamatan saat pengukuran sampel, yaitu daun yang memiliki luasan sama, seringkali memiliki panjang petiole dan jumlah tulang daun yang berbeda.
Terjadi variasi nilai korelasi LL dengan variabel ciri-ciri dimensional seperti PL, LW, WP, dan NV untuk M. triloba. Nilai korelasi antara LL dan LW sebesar 0,915. Nilai ini merupakan nilai korelasi terbesar dari variabel lainnya. Variabel dimensional lain yang memiliki korelasi sangat kuat adalah PL (panjang petiole). Nilai korelasi LL dengan PL sebesar 0,819. Korelasi yang kuat antara LL dengan LW dan PL juga terlihat saat pengukuran sampel daun, dimana daun M. triloba yang diamati memiliki bentuk pisau bulat telur, karakteristik daun M. triloba adalah lebar dibagian bawahnya dan menyempit bertahap hingga ujung daun, sehingga kenaikan panjang daun akan menyebabkan kenaikan lebar daun dan panjang petiole (Noofaizal et al. 2012). Variabel ciri-ciri dimensional lain seperti WP dan NV hanya berkorelasi cukup kuat atau sedang. Artinya perubahan nilai WP dan NV berkorelasi dengan perubahan nilai LL namun pengarunya tidak begitu kuat.
Pengaruh Perbedaan Lansekap dan Plot Pengambilan Sampel Terhadap
Variasi Morfologi Daun A. scholaris dan M. triloba
10
Tabel 6 Hasil sidik ragam pengaruh lansekap dan plot terhadap variasi morfologi daun A. scholaris dan M. triloba
Jenis Perlakuan Variabel
LL PL LW WP NV LS PR LWR
A. scholaris
Lansekap 0,5753tn 0,3843tn 0,4961tn 0,4961tn 0,0038* 0,8250tn 0,3224tn 0,7780tn
Plot 0,6891tn 0,0583tn 0,6792tn 0,6795tn 0,0273* 0,6318 0,0267* 0,1495
M. triloba
Lansekap 0,2985tn 0,0924tn 0,01426tn 0,8458tn 0,7713tn 0,6279tn 0,1244tn 0,0591tn
Plot 0,0001** 0,0080** 0,0002** 0,5663tn 0,0064** 0,0185* 0,6499tn 0,0028**
Ket: **= Perlakuan berpengaruh nyata pada α 1%; *= Perlakuan berpengaruh nyata pada α=5%; tn= Perlakuan tidak berpengaruh nyata pada α=5%.
Variasi Morfologi Daun A. scholaris dan M. triloba Di Hutan Karet Harapan
dan Hutan Karet Bukit Duabelas
Variasi yang terjadi karena adanya kondisi lingkungan menunjukkan bahwa suatu tumbuhan melakukan adaptasi (Jones dan Luchsinger 1987). Sebagai bentuk adaptasi terhadap kondisi alam atau tekanan lingkungan, tumbuhan dapat mengalami plastisitas fenotipe, yaitu kemampuan suatu individu untuk memodifikasi beberapa sifat khusus selama masa perkembangannya (Jones dan Wilkins 1974). Tumbuhan menyediakan banyak bukti mengenai perubahan lingkungan tempat tumbuh, salah satunya melalui organ daun. Daun merupakan salah satu organ yang berkembang dengan cukup cepat dan tergolong sensitif. Cox dan Moore (1980) menyebutkan bahwa terdapat korelasi antara iklim dan karakter daun. Ukuran daun dan tepi daun dapat menyediakan informasi bagi proses adaptasi tumbuhan terhadap rata-rata curah hujan dan suhu.
Alstonia scholaris
11
Gambar 3 Dendogram hasil CA untuk A. scholaris (skala menggunakan standar deviasi)
Macaranga triloba
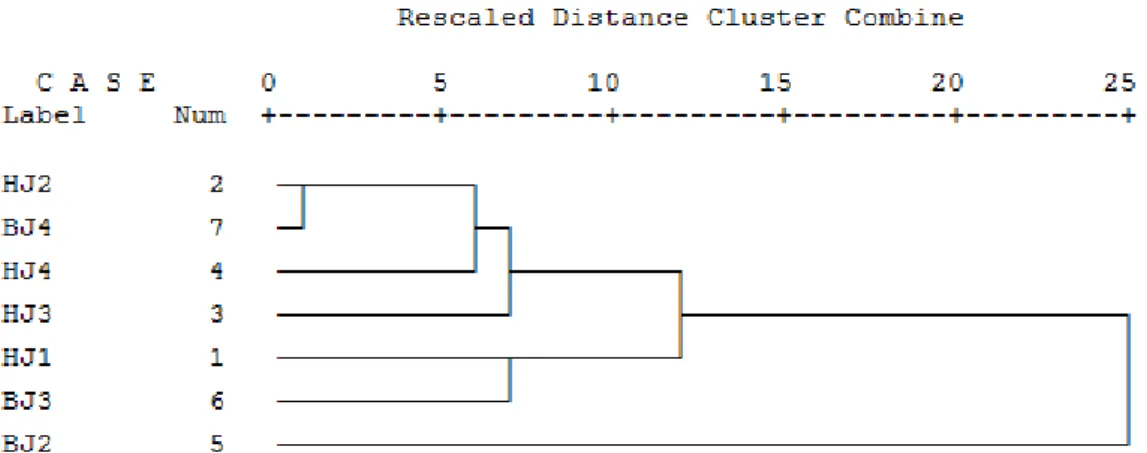
Hasil Cluster Analysis (CA) untuk M. triloba pada 8 plot yang diamati menunjukkan adanya 2 kelompok (group) berdasarkan variasi morfologi daunnya. Dendogram hasil CA disajikan pada Gambar 4. Berdasarkan dendogram terlihat bahwa berdasarkan variasi morfologi daunnya plot HJ2, BJ4, HJ1, HJ4, HJ3, dan BJ5 memiliki banyak kesamaan jika dibandingkan dengan plot BJ3 3. Hal tersebut ditandai dengan koefisien jarak (distance coefficient) antar plot HJ2, BJ4, HJ1, HJ4, HJ3, dan BJ5 lebih dekat jika dibandingkan dengan plot BJ3 3. Koefisien jarak menunjukkan besarnya nilai perbedaan antar kelompok sehingga semakin besar nilai koefisien jarak maka semakin besar pula perbedaan antar kedua kelompok (McGarigal et al 2000). Koefisien jarak yang terbesar diantara 8 plot yang diamati adalah pada plot BJ310, plot ini digolongkan sebagai outgroup, hal ini berarti berdasarkan variasi morfologi daun plot BJ3 10 sangat berbeda dengan 7 plot lainnya.
12
Sebaran Variasi Morfologi Daun A. scholaris dan M. triloba Terhadap
Perbedaan Lansekap (HJ dan BJ)
Tiga metode analisis multivariat digunakan pada keseluruhan data sampel daun A. scholaris dan M. triloba. Variabel sintetis pertama pada masing-masing metode yang digunakan berkontribusi paling tinggi terhadap total varian seperti yang diharapkan, dilanjutkan oleh variabel sintesis kedua. Nilai pada variabel sintesis menunjukkan keragaman yang dapat diwakilkan dari masing-masing metode.
Tabel 7 Proporsi dari total varian yang dijelaskan oleh variabel sintesis pertama dan kedua dari analisis multivariat yang digunakan (CDA;PCA;MCA) pada A. scholaris dan M. triloba
A. scholaris M. triloba
CDA MCA PCA CDA MCA PCA Variabel
Sintesis 1 100% 43,18% 36,20% 100% 53,65% 43,30% Variabel
Sintesis 2 0% 38,79% 26,00% 0% 46,87% 22,00% Total 100% 81,97% 62,20% 100% 99,42% 65,30%
Tabel 7 menunjukkan kecenderungan yang sama pada hasil analisis multivariat pada A. scholaris dan M. triloba. Tiga analisis multivariat yang digunakan mampu menjelaskan lebih dari 50% total varian yang ada, sehingga representatif untuk menunjukkan sebaran variasi morfologi daun A. scholaris dan M. triloba pada dua lansekap yang berbeda. Variabel sintesis pertama pada CDA untuk kedua jenis memberikan sumbangan varian sebesar 100%, artinya total varian dari data variasi morfologi daun A. scholaris dapat dijelaskan oleh variabel sintesis pertama CDA. Analisis lebih lanjut dilakukan untuk mengetahui variabel apa saja yang berperan dalam pembentukan variabel sintesis pertama dalam CDA pada kedua jenis.
13 Tabel 8 Test of quality of group means A. scholaris
Variabel Wilks' Lambda F df1 df2 Sig. Ket
LL .982 4.251 1 235 .040 s
PL .985 3.468 1 235 .064 ns
LW .995 1.300 1 235 .255 ns
WP .985 3.587 1 235 .059 ns
NV .885 30.596 1 235 .000 s
LS .998 .446 1 235 .505 ns
PR .975 5.994 1 235 .015 s
LWR .996 .982 1 235 .323 ns
Ket: s= signifikan pada α=5%; ns= tidak signifikan pada α=5%
Tabel 9 Test of quality of group means M. triloba
Variabel Wilks' Lambda F Df1 Df2 Sig. Ket
LL .978 1.619 1 71 .207 ns
PL .952 3.562 1 71 .063 ns
LW .971 2.133 1 71 .149 ns
WP .998 .117 1 71 .733 ns
NV .991 .661 1 71 .419 ns
LS 1.000 .024 1 71 .877 ns
PR .970 2.177 1 71 .145 ns
LWR .982 1.274 1 71 .263 ns
Ket: ns= tidak signifikan pada α=5%
Berdasarkan nilai F pada Tabel 9, tiga variabel yang memberikan pengaruh besar pada variabel sintesis pertama adalah PL, PR, dan LW, namun ketiga variabel tersebut tidak signifikan mampu mengelompokkan variabel kedalam dua lansekap. Hal ini terlihat dari nilai signifikansi yang lebih besar dari nilai α. Kesimpulan tersebut didukung oleh hasil dari Wilks’ Lamda yang tinggi. Nilai
Wilks’ Lamda berada dalam interval 0-1 dimana semakin nilainya mendekati 0
menunjukkan semakin signifikan variabel tersebut membedakan kelompok lokasi. Sebaliknya jika nilainya mendekati 1 maka variabel tersebut semakin tidak mampu untuk membedakan kelompok lokasi.
14
26,0% terhadap total varian. Variabel morfologi daun yang memberikan pengaruh besar terhadap variabel sintesis pertama A. scholaris adalah PL, LL, LW, sedangkan untuk variabel sintesis kedua adalah LS, LWR, dan PR. Analisis menggunakan PCA untuk M. triloba, menunjukkan variabel sintesis pertama memberikan sumbangan 43,3% terhadap seluruh varian, variabel sintesis kedua memberikan sumbangan sebesar 22,0% terhadap seluruh varian. Variabel morfologi daun yang memberikan pengaruh besar terhadap variabel sintesis pertama adalah LL, LW, dan WP, sedangkan variabel morfologi daun yang berpengaruh pada variabel sintesis kedua adalah PR, PL, dan NV. Selengkapnya disajikan pada Tabel 10.
Tabel 10 Korelasi seluruh variabel morfologi daun A. scholaris dan M. triloba terhadap variabel sintesis 1 dan 2 dengan metode PCA
Variabel A. scholaris M. triloba
VS1 VS2 VS1 VS2
LL -0,486 -0,274 -0,520 -0,246
PL -0,490 -0,018 -0,344 0,521
LW -0,485 -0,088 -0,485 -0,106
WP -0,411 0,346 -0,480 -0,278
NV -0,303 -0,180 -0,381 0,308
LS -0,087 0,660 -0,017 -0,068
PR -0,138 0,367 -0,024 0,668
LWR 0,025 0,441 0,007 0,181
Ket: VS= Variabel sintesis
15 Tabel 11 Korelasi semua variabel morfologi daun A. scholaris dan M. triloba
terhadap variabel sintesis 1 dan 2 dengan metode MCA
Variabel
Ket: VS= Variabel sintesis
16
Gambar 6 Distribusi dari variabel sintesis pertama dan kedua yang terbentuk dari dua analisis multivariat yang berbeda pada M. triloba. (A) dan (C) adalah distribusi daun pada dua variabel sintesis yang terbentuk (variabel sintesis pertama sebagai X, variabel sintesis kedua sebagai Y); (B) dan (D) menjelaskan distribusi dari variabel sintesis pertama; (A) dan (B): PCA; (C) dan (D): MCA.
Distribusi dari variabel sintesis pertama dan kedua pada dua metode analisis multivariat yang digunakan, yaitu PCA dan MCA, disajikan dalam diagram pencar, sementara distribusi dari variabel sintesis pertama terhadap jumlah spesiemen yang diamati disajikan dalam bentuk histogram (Gambar 5 dan 6). Hasil analisis menggunakan CDA tidak ditampilkan karena hasilnya tidak dapat digunakan untuk analisis selanjutnya. Diagram pencar dan histogram yang dihasilkan oleh kedua analisis multivariat yang berbeda menunjukkan persebaran yang acak dengan pola yang berbeda yang mengindikasikan perbedaan rentang adaptasi dan plastisitas. Secara umum daun A. scholaris dan M. triloba yang diamati tidak menunjukkan pengelompokan berdasarkan lansekap.
Kesamaan hasil pada diagram pencar dan histogram berdasarkan PCA dan MCA pada A. scholaris dan M. triloba ini menunjukkan bahwa berdasarkan dua metode tersebut, delapan variabel yang diukur dari masing-masing daun A. scholaris dan M. triloba, tidak mampu mengklasifikasikan kedua jenis tersebut ke dalam dua lansekap yang berbeda. Hal tersebut diduga dapat disebabkan adanya variasi kontinu pada karakter morfologi A. scholaris dan M. triloba karena karakter morfologi yang digunakan termasuk dalam karakter metrik. Briggs dan Walters (1984) menyebutkan bahwa penggunaan karakter metrik dalam identifikasi jenis berdasarkan morfologi menyebabkan adanya variasi yang bersifat kontinu sehingga tidak dapat menunjukkan batas variasi morfologi yang jelas. Selain itu sebaran acak pada variasi morfologi A. scholaris dan M. triloba diduga karena kedua spesies ini merupakan spesies yang memiliki plastisitas
17 tinggi. Davies dan Ashton (1999) menyatakan bahwa M. triloba adalah jenis yang memiliki salah satu tumbuhan pionir yang beradaptasi tinggi untuk tumbuh di lahan terbuka pada kondisi ekologi yang beragam. Hal ini mengakibatkan perbedaan lingkungan dapat menimbulkan respon beragam pada morfologi daun.
Keragaman Sekuen DNA A. scholaris dan M. triloba
Hasil yang didapat pada bab sebelumnya menyatakan bahwa keragaman A.scholaris dan M. triloba berdasarkan morfologi daunnya sangat tinggi. Kajian lebih lanjut dilakukan untuk mengetahui apakah kergaman A.scholaris dan M. triloba berdasarkan keragaman genetik juga tinggi. Keragaman genetik mempunyai arti penting dalam stabilitas dan ketahanan populasi, kehilangan keragaman genetik akan mengurangi kemampuan spesies tersebut untuk beradaptasi terhadap perubahan lingkungan. Keragaman genetik juga mempunyai dampak secara langsung maupun tidak terhadap populasi, komunitas, dan ekosistem (Hughes et al 2008). Salah satu cara mengetahui keragaman genetik suatu jenis tanaman adalah dengan melihat keragaman sekuen DNAnya. DNA pada suatu tanaman dapat berasal dari kloroplas dan nucleus. DNA yang berasal dari kloroplas cenderung konservatif atau tidak mudah berubah, dan telah disepakati sebagai untuk digunakan dalam barcode (Kolondam et al. 2012). Dua region pada DNA kloroplas diamati dalam penelitian ini, yaitu region matK untuk jenis A. scholaris dan region rbcL untuk jenis M. triloba. Keragaman sekuens DNA A. scholaris dan M. triloba yang didapat dari data mining pada database GenBank dalam situs National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.g-ov/genbank/) disajikan pada Tabel 12.
Tabel 12 Hasil data mining untuk A. scholaris dan M. triloba (GenBank 2014)
Spesies Region Sumber Genbank Accession
Asal Panjang Basa (Bp)
A. scholaris matK Mahadani et al. (2011) JN228931 India 825
A. scholaris matK Toyama et al. (2014) AB925226 Kamboja 765
A. scholaris matK Chen et al. (2010) GQ434101 China 811
A. scholaris matK Bremer et al. (2002) AJ429321 - 1672
A. scholaris matK Endress et al. (1996) Z70189 - 1518
A. scholaris matK Kong et al. (2014) KJ510993 China 819
M. triloba rbcL Banfer et al. (2006) DQ358346
Asia
Selatan 792
M. triloba rbcL Banfer et al. (2006) DQ358318
Asia
Selatan 818
M. triloba rbcL Banfer et al. (2006) DQ358316
Asia
Selatan 818
M. triloba rbcL Banfer et al. (2006) DQ35810
Asia
Selatan 820
M. triloba rbcL Banfer et al. (2006) DQ358305
Asia
18
Berdasarkan pengambilan data menggunakan metode data mining, terdapat 6 sekuens DNA dari 6 individu berbeda untuk jenis A. scholaris. Sementara itu, untuk jenis M. triloba, didapatkan 6 sekuens DNA dari 1 individu. Perbedaan sekuen tersebut hanya didasarkan pada perbedaan panjang basa. Data sekuens yang didapat untuk jenis M. triloba dirasa tidak mewakili tujuan penelitian, yaitu untuk melihat keragaman genetik M. triloba pada perbedaan lingkungan, karena berasal dari individu yang sama di tempat yang sama, oleh karena itu analisis keragaman genetik hanya dilakukan untuk jenis A. scholaris saja.
Hasil sekuen yang didapat untuk jenis A. scholaris di lakukan alignment dengan menggunakan software ClustalX2 kemudian sekuen DNA dirunutkan melalui program BioEdit. Hasil runutan sekuen DNA untuk 6 individu A. scholaris menunjukkan rata-rata nukleotida yang banyak ditemukan adalah nukleotida T(U) sebesar 35,9%, diikuti dengan A sebesar 29,8% dan C sebesar 17,9%. Nukleotida yang paling sedikit ditemukan adalah G dengan rata-rata sebesar 16,4%. Secara lengkap komposisi dari nukleotida yang terdapat pada masing-masing sekuen disajikan pada Tabel 13.
Tabel 13 Komposisi nukleotida dari 6 individu A. scholaris region matK Spesies T(U) (%) C (%) A (%) G (%) Sumber
JN228931 35,9 18,3 29,5 16,4 Mahadani et.al
AB925226 36,1 17,8 29,9 16,2 Toyama et al.
GQ434101 36,0 19,4 28,5 16,2 Chen et al.
AJ429321 35,5 16,9 30,7 17,0 Bremer et al.
Z70189 35,9 17,8 29,9 16,4 Endress et.al
KJ510933 36,5 18,1 29,3 16,1 Kong et al.
Rata-rata 35,9 17,9 29,8 16,4
Perunutan DNA ini dapat membantu dalam pendeteksian perbedaan nukleotida atau nukleotida yang bermutasi (Sambrook dan Russel 2001). Keragaman genetik dapat muncul karena adanya mutasi, aliran gen, migrasi, dan proses seleksi (Finkeldey 2005). Hasil pengamatan dari runutan sekuen DNA menunjukkan bahwa keragaman nukleotida dalam spesies (intraspesies) pada jenis A. scholaris beragam. Hal ini dipengaruhi oleh panjang basa yang beragam pula.
19 Tabel 14 Jarak genetik nukleotida dan jarak lokasi pengambilan sampel pada A.
scholaris region matK Spesies
(Asal)
KJ510933
(China)
AB925226
(Kamboja) Z70189
JN228931
(India) AJ429321
GQ434101
(China)
KJ510933
(China) - 2454.10 - 2665.60 - 0.00
AB925226
(Kamboja) 0.0028 - - 3033.87 - 2454.10
Z70189 0.0028 0.0000 - - -
-JN228931
(India) 0.0028 0.0000 0.0000 - - 2665.60
AJ429321 0.0028 0.0000 0.0000 0.0000 - -GQ434101
(China) 0.0028 0.0000 0.0000 0.0000 0.0000 -
Jarak genetik adalah perselisihan genetik antara spesies atau antara populasi dalam satu spesies tertentu. Jarak genetik yang kecil menunjukkan hubungan genetik yang dekat dan sebaliknya, jarak genetik yang besar menunjukkan hubungan genetik yang jauh (Finkeldey 2005). Tabel 14 menyajikan jarak genetik keenam individu A. scholaris yang diamati (di bawah diagonal) serta jarak lokasi asal sampel (di atas diagonal) dalam satuan km. Jarak lansekap asal sampel dianalisis menggunakan bantuan google map (https://maps.google.com/). Berdasarkan data pada tabel dapat dilihat bahwa spesies dengan kode KJ510933 memiliki hubungan genetik yang berbeda dari ke 5 spesies lainnya. Hal ini terlihat dari jarak genetik KJ510933 terhadap ke 5 spesies lainnya yaitu sebesar 0,0028. Sementara, dari data tabel dapat dilihat bahwa 5 spesies A. scholaris lainnya yang diamati yaitu: AB925226; Z70189; JN228931; AJ429321; dan GQ434101,menunjukkan hubungan genetik yang erat karena jarak genetiknya sebesar 0.0000.
A. scholaris menyebar di Australia, Bangladesh, Brunei, Cambodia, China, India, Indonesia, Laos, Malaysia, Myanmar, Nepal, Papua New Guinea, Philippines, Solomon Islands, Sri Lanka, Thailand, dan Vietnam (Orwa et al. 2009). Berdasarkan Tabel 14 dilihat bahwa jarak lokasi pengambilan sampel tidak mempengaruhi jarak genetik dari sampel tersebut. Artinya keragaman sekuen DNA pada A.scholaris beragam baik pada lokasi yang sama, misalnya antara individu KJ5100933 yang berasal dari China dan individu GQ434101 yang juga bersaal dari China, maupun dari lokasi yang berbeda. Hal ini diduga disebabkan lokasi pengambilan sampel masih dalam wilayah penyebaran A. scholaris, sehingga perbedaan lokasi tidak terlalu mempengaruhi keragaman genetiknya.
20
Gambar 7 Pohon filogeni 6 spesies A. scholaris yang diamati pada region matK
SIMPULAN DAN SARAN
Simpulan
Berdasarkan hasil penelitian didapatkan kesimpulan bahwa:
1. Ada pengaruh perbedaan plot di dalam lansekap untuk beberapa variabel morfologi daun A. scholaris dan M. triloba. Perlu verifikasi kualitas tanah dan iklim mikro untuk mengkaji pengaruh tersebut secara detail.
2. Hasil analisis sebaran variasi morfologi daun A. scholaris dan M. triloba dari tiga analisis multivariat yang digunakan menunjukkan adanya perbedaan daya plastisitas kedua jenis, dimana M. triloba memiliki rentang plastisitas yang lebih lebar ditinjau dari hasil pada diagram pencar.
3. Keragaman sekuen DNA pada A.scholaris beragam baik pada lokasi yang sama maupun dari lokasi yang berbeda.
Saran
Perlu dilakukan penelitian lanjutan mengenai variasi morfologi daun dengan metode lain agar perbedaan tempat tumbuh dapat dikaji lebih dalam.
DAFTAR PUSTAKA
[CRC] Collaborative Research Center. 2012. Collaborative research center 990: ecological and socioeconomic function of tropical lowland rainforest transformation system (Sumatra, Indonesia). [Internet]. [diunduh Jun 18]. Tersedia pada: www.uni-goettingen.de/en/310995.html.
[Kemenhut] Kementrian Kehutanan. 2006. 50 Taman Nasional Indonesia. Bogor (ID): Kementrian Kehutanan.
Banfer G, Moog U, Fiala B, Mohamed M, Weising K, Blattner FR. 2006. A. chloroplast genealogy of myrmecophytic Macaranga spesies (Euphorbiaceae) in Shoutest Asia reveals hybridization, vicariance and long distance dispersals. J. Mol. Ecol. 15(14):4409-4424.
AJ429321
GQ434101
JN228931
AB925226
Z70189
KJ510933
21 Barbour MG, Burk JH, Pitts WD, William FS. 1999. Terrestrial Plant Ecology.
Third Edition. California (US): Addision Wesley Longman, Inc.
Bremer B, Bremer K, Heidari N, Erixon P, Olmstead RG, Anderberg AA, Kallersjo M, Barkhordarian E. 2002. Mol. Phylogenet. Evol. 24(2):274-301. Briggs D, Walters SM. 1884. Plant Variation And Evolution. 2nd ed. Cambridge
(UK): Cambridge University Press.
Cassie RM. Multivariate analysis in ecology. Proceeding of the New Zealand ecological society. 16:53-57.
Chen S, Yao H, Han J, Liu C, Song J, Shi L, Zhu Y, Gao T, Pang X, et al. 2010. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant spesies. J. Plos. ONE. 5(1):E8613.
Cox CB, Moore PD. 1980. Biogeography: An ecological and evolutionaru approach “3rd ed”. New York (US): John Willey & Sons Inc.
Davies SJ, Ashton PS. 1999. Phenology and fecundity in 11 sympatric pioneer species of Macaranga (euphorbiaceae) in borneo. American Journal of Botany. 86(12): 1786 – 1795.
Dewi S, Ekadinata A, Nugroho DK. 2008. Land cover changes in different forest transition stages in Indonesia: East Kalimantan, Jambi and Lampung. Bogor (ID): World Agroforestry Centre (ICRAF).
Endress ME, Sennblad B, Nilsson S, Civeyrel L, Chase MW, Huysmans S, Grafstroem E, Bremer B. 1996. A phylogenetic analysis of Apocynaceae s.str. and some related taxa in Gentiales, A multidiciplinary approach. 1996. Opera Bot. Belg. 7:59-102.
Finkeldey R. 2005. Pengantar Genetika Hutan Tropis. Djamhuri E, Siregar IZ, Siregar UJ, Kertadikara AW, penerjemah. Goettingen: Institut of Forest Genetics and Forest Tree Breeding Georg-August-University-Goettingen. Terjemahan dari: An Introduction to Tropical Forest Genetics.
Henderson A. 2006. Traditional morphometrics in plant systematic and its role in palms systematic. Botanical Journal of the Linneas Society. 151:103-111. Hughes AR, Inouye BD, Jhonson J, Underwood N, Vellend M. 2008. Ecological
consequences of genetic diversity. Ecology Letters. 11:609-623.
Irwanto. 2007. Analisis vegetasi untuk pengelolaan kawasan hutan lindung marsegu, Kabupaten Seran bagian barat, Provinsi Maluku [Tesis]. Yogyakarta: Sekolah Pascasarjana Universitas Gadjah Mada.
Jones SB, Luchsinger AE. 1986. Plant systematics, Second Edition. New York (US): McGraw-Hill Book Company.
Jones SB, Luchsinger AE. 1987. Plant Systematic 2nd edition. New York (US): McGraw-Hill.
Koch K, Hartman KD, Schreiber L, Barthlott W, Neinhuis C. 2006. Influences of air humidity during the cultivation of plants on wax chemical composition, morphology and leaf surface wettability. Env. Exp. Bot. 56:1-9.
Kolondam BJ, Lengkong E, Polii-Mandang J, Pinaria A, Rutunuwu S. 2012. Barcode dna berdasarkan gen rbcL dan matK Anggrek Payus Limondok (Phaius tancarvilleae). Bioslogos. 2(2):58-62.
22
Kremer A et al.. 2001. Leaf morphological differentiation between Quercus robur and Querqus petraea in stable across Western European mixed oak stands. For. Science: 59(777-787).
Mahadani P, Sharma GD, Ghosh SK. 2011. Evaluation of matK sequence for species level DNA passport in medicinal Rauvolfioidae (Apocynaceae) plants from northeast India. [Internet]. [diunduh 2014 Jun 18]. Tersedia pada: http://www.ncbi.nlm.nih.gov/nuccore/AB925226.1.
Manan S. 1979. Masalah Pembinaan Kelestarian Ekosistem Hutan. Bogor (ID): Departemen Manajemen Hutan, Fakultas Kehutanan IPB.
Margono BA, Potapov PV, Turubanova S, Stolle F, Hansen MC. Primary forest cover loss in Indonesia over 2000-2012. [internet]. [diunduh 2014 Jul 13]. Tersedia pada:http://www.uni-goettingen.de.sci-hub.org/document/downloa d/e6ee0fcaedcae51098a62d8df2e38aaa.pdf.
McGarigal K, Cushman S, Stafford S. 2000. Multivariate statistic for wildlife and ecology research. New York (US): Spinger-Verlag.
Norfaizal GM, Khatijah H, Ruzi ARM. 2012. Leaf anatomical study of five Macaranga spesies (Euphorbiaceae). J. Trop. Agric. And Fd. Sc. 40(2):289-296.
Odum EP. 1993. Dasar-Dasar Ekologi. Edisi ketiga. Yogyakarta: Gajah Mada University Press.
Orwa C, Mutua A, Kindt R, Jamnadass R, Anthony S. 2009. Agroforestree database: a tree reference and selection guide version 4.0. world agroforestry center, Kenya. [Internet]. [diunduh 2014 Jun 12]. Tersedia pada: http://www.worldagroforestry.org/treedb2/AFTPDFS/Alstonia_schola ris.pdf.
REKI PT. 2009. Rencana Kerja Usaha Pemanfaatan Hasil Hutan Kayu Restorasi Ekosistem (RKUPHHK) dalam Hutan Alam pada Hutan Produksi Periode Tahun 2008 – 2017 Kabupaten Musi Banyuasin Provinsi Sumatera Selatan. Tidak Dipublikasikan.
Smith RL. 1990. Ecology and Field Biology. New York (US): Harper and Row. Supranto. 2010. Analisis Multivariat: Arti dan Interpretasi. Jakarta (ID): PT Asdi
Mahasatya.
23
RIWAYAT HIDUP
Penulis dilahirkan di Lampung Tengah pada tanggal 21 April 1992 dari Ayah Alm. Muhammad Toha dan ibu Summayah. Penulis adalah putri pertama dari dua bersaudara. Tahun 2010 penulis lulus dari SMA Negeri 1 Kotagajah dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB, diterima di Departemen Silvikultur, Fakultas Kehutanan dan mendapatkan beasiswa Bidikmisi.
Selama mengikuti perkuliahan, penulis aktif di berbagai organisasi yaitu sebagai anggota lembaga kemahasiswaan IFSA LC IPB pada divisi Public Relation (PR) pada tahun 2011, anggota seedling group Himpunan profesi Tree Grower Community pada tahun 2011, anggota Scientific Improvement (SI) Himpunan profesi Tree Grower Community pada tahun 2013. Selain itu penulis juga aktif di kepanitiaan yakni sebagai panitia Forester Cup 2011, panitia SEAFYM (South East Asia Forest Youth Meeting) 2011, panitia Belantara 2012, sekretaris EKSFLORASI 2013, panitia TGC in action 2013, dan panitia Bina Corps Rimbawan 2013. Penulis pernah menjadi asisten praktikum dalam mata kuliah Silvika pada tahun 2013. Penulis melaksanakan Praktek Pengenalan Ekosistem Hutan (PPEH) di TN Gunung Ciremai - Indramayu. Penulis melakukan Praktek Pembinaan Hutan (PPH) di Hutan Pendidikan Gunung Walat (HPGW) Sukabumi tahun 2013. Penulis juga telah melaksanakan Praktek Kerja Profesi (PKP) di Persemaian Permanen BPDAS Citarum – Ciliwung pada bulan Februari sampai dengan April 2014. Untuk memperoleh gelar Sarjana Kehutanan IPB, penulis menyelesaikan skripsi dengan judul “Variasi Morfologi Daun Jenis Pionir Pulai (Alstonia scholaris R. Br.) dan Macaranga (Macaranga triloba (Bl.) Muell.
Arg.) di Hutan Karet Jambi” di bawah bimbingan Prof Dr Ir Iskandar Z. Siregar,