IDENTIFIKASI
Trichoderma
sp. MELALUI SEKUEN
INTERNAL
TRANSCRIBED SPACER
(ITS) DAN UJI INHIBISINYA
TERHADAP
Colletotrichum
sp. DAN
Rhizoctonia
sp.
LISA KARIS
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Identifikasi
Trichoderma sp. Melalui Sekuen Internal Transcribed Spacer (ITS) dan Uji Inhibisinya terhadap Colletotrichum sp. dan Rhizoctonia sp. adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
Lisa Karis
ABSTRAK
LISA KARIS. Identifikasi Trichoderma sp. Melalui Sekuen Internal Transcribed Spacer (ITS) dan Uji Inhibisinya terhadap Colletotrichum sp. dan Rhizoctonia sp. Dibimbing oleh MARIA BINTANG dan PUJI LESTARI.
Rhizoctonia sp. dan Colletotrichum sp. merupakan patogen yang banyak menyerang tanaman. Penggunaan agen biokontrol dalam melawan patogen tanaman menjadi salah satu alternatif terhadap penggunaan pestisida sintetik. Penelitian ini bertujuan mengidentifikasi Trichoderma sp. secara morfologi dan molekuler serta menyeleksi isolat Trichoderma sp. yang memiliki penghambatan terbesar terhadap patogen secara in vitro. Metode identifikasi dilakukan dengan mikroskop cahaya dan amplifikasi daerah Internal Transcribed Spacer (ITS) sedangkan seleksi dilakukan dengan metode uji berganda secara bertahap. Secara umum, Trichoderma sp. memiliki permukaan rata, tepian regular, berwarna kuning sampai hijau, serta konidiofor bercabang. DNA hasil amplifikasi menggunakan primer ITS1 dan ITS4 berukuran 600-700 bp. Hasil identifikasi dari 17 isolat didapatkan sebanyak 13 isolat merupakan Trichoderma asperellum, 1 isolat Trichoderma gamsii, 1 isolat Trichoderma koningiopsis, dan 2 sisanya
Trichoderma harzianum. Nilai penghambatan > 80% yang lebih banyak menunjukkan isolat Trichoderma sp. lebih berpotensi menghambat Rhizoctonia sp. dibandingkan Colletotrichum sp.
Kata kunci: antagonis, Colletotrichum sp., ITS, Rhizoctonia sp., Trichoderma sp.
ABSTRACT
LISA KARIS. Identification Trichoderma sp. through Internal Transcribed Spacer (ITS) and Inhibition Assay against Colletotrichum sp. and Rhizoctonia sp. Supervised by MARIA BINTANG and PUJI LESTARI.
Rhizoctonia sp. and Colletotrichum sp. are pathogen that can attack many plants. The use of biocontrol agents against plant pathogens become an alternative to synthetic pesticides. This study aims to identify the Trichoderma sp. through morphological and molecular characterizations, and to select isolates Trichoderma
sp. which has the largest inhibition against both pathogens via in vitro assay. Identification method performed by light microscopy and amplification of Internal Transcribed Spacer (ITS) region, while the selection was done with multiple test methods gradually. In general, Trichoderma sp. has a flat surface, regular edges, yellow to green color, as well as branched conidiophores. DNA amplification using primers ITS1 and ITS4 sized in a range of 600-700 bp. Results of identification of 17 isolates were obtained 13 isolates that are Trichoderma asperellum, 1 isolate Trichoderma gamsii, 1 isolate Trichoderma koningiopsis, and rest of it are Trichoderma harzianum. Trichoderma sp. more potent as inhibitors to Rhizoctonia sp. compared Colletotrichum sp. based on inhibition values > 80 % which more on Rhizoctonia sp.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biokimia
IDENTIFIKASI
Trichoderma
sp. MELALUI SEKUEN
INTERNAL
TRANSCRIBED SPACER
(ITS) DAN UJI INHIBISINYA
TERHADAP
Colletotrichum
sp. DAN
Rhizoctonia
sp.
LISA KARIS
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas segala karunia-Nya sehingga karya ilmiah ini dapat diselesaikan dengan baik. Judul penelitian yang dilaksanakan sejak bulan Januari 2015 ini adalah Identifikasi
Trichoderma sp. Melalui Sekuen Internal Transcribed Spacer (ITS) dan Uji Inhibisinya Terhadap Colletotrichum sp. dan Rhizoctonia sp.
Terima kasih penulis ucapkan kepada Prof Dr drh Maria Bintang, MS dan Ibu Puji Lestari Sp, MSi, PhD selaku pembimbing yang telah banyak memberikan pengarahan dan saran. Di samping itu, penghargaan penulis sampaikan kepada Bapak Jajang, Bapak Ughi dan Ibu Aminah beserta seluruh staf Laboratorium Mikrobiologi Konservasi Mikroorganisme dan staf Laboratorium Genom BB-Biogen yang telah membantu selama pengumpulan data penelitian. Ungkapan terima kasih juga disampaikan kepada Ayah, Ibu, Kakak, Adik, sahabat dekat (Cynthia, Ken, Caroline, Desi, Kathi, Liya, dan Chelsea), serta teman-teman Biokimia 48 untuk segala doa, motivasi, inspirasi, kasih sayang dan dukungannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2015
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
METODE 2
Alat dan Bahan 2
Prosedur Penelitian 2
HASIL 5
Morfologi Isolat Trichoderma sp. 4
DNA Total 17 Isolat Trichoderma sp. dan Amplifikasi Sekuen ITS 5 Homologi dan Filogenetik Isolat Trichoderma sp. 6 Kemampuan Antagonis Trichoderma sp. terhadap Colletotrichum sp. dan
Rhizoctonia sp. 8
PEMBAHASAN 10
Morfologi Isolat Trichoderma sp. 10
DNA Total 17 Isolat Trichoderma sp. dan Amplifikasi Sekuen ITS 11 Homologi dan Filogenetik Isolat Trichoderma sp. 11 Kemampuan Antagonis Trichoderma sp. terhadap Colletotrichum sp. dan
Rhizoctonia sp. 12
SIMPULAN DAN SARAN 13
Simpulan 13
Saran 14
DAFTAR PUSTAKA 14
LAMPIRAN 18
DAFTAR TABEL
1 Kuantitatif 17 isolat DNA Trichoderma sp. 6
2 Identifikasi 17 isolat Trichoderma sp. 7
3 Daya hambat 17 isolat Trichoderma sp. terhadap cendawan patogen 8 4 Daya hambat 3 isolat Trichoderma sp. terseleksi terhadap
Colletotrichum sp. 9
5 Daya hambat 3 isolat Trichoderma sp. terseleksi terhadap Rhizoctonia
sp. 9
DAFTAR GAMBAR
1 Morfologi isolat Gam4 (A) dan PB2 (B) 5
2 DNA Trichoderma sp. hasil isolasi 5
3 Amplikon hasil PCR 17 isolat Trichoderma sp. 6 4 Pohon filogenetik antar isolat Trichoderma sp. yang dibandingkan
dengan database NCBI serta sekuen outgroup 7
5 Kontrol Colletorichum sp. (A) dan inhibisi isolat Pan23.1 terhadap
Colletotrichum sp. (B) 9
6 Kontrol Rhizoctonia sp. (A) dan inhibisi isolat Pan23.2 terhadap
Rhizoctonia sp. (B) 9
7 Inhibisi isolat R.Pan23.2 (A) dan C.TR3 (B) 10
DAFTAR LAMPIRAN
1 Diagram alir penelitian 18
2 Identitas 17 isolat Trichoderma sp. 19
3 Contoh hasil sekuensing Trichoderma sp. 20
PENDAHULUAN
Penurunan hasil panen berbagai tanaman pangan banyak disebabkan oleh penyakit tanaman. Salah satu yang menyebabkan penyakit tanaman yaitu cendawan patogen, diantaranya Colletotrichum sp. dan Rhizoctonia sp. Kedua patogen tersebut dapat menyerang tanaman saat pertumbuhan maupun pasca panen. Menurut Herwidyarti et al. (2013), penyakit yang dapat disebabkan oleh
Colletotrichum sp., yaitu busuk batang (Colletotrichum graminearum), antraknosa, dan gugur daun yang menyerang papaya, pisang, cabai, tomat, kentang, kakao, mentimun, dan kacang. Sementara itu, Rhizoctonia sp. dapat menyebabkan penyakit batang kawat, busuk pelepah, busuk rimpang, penyakit semai (Rhizoctonia solani), dan mati pucuk (Yu et al. 2002; Athul dan Jisha 2013).
Tindakan preventif yang saat ini cenderung dilakukan oleh petani Indonesia yaitu menggunakan pestisida sintetik. Penerapan yang lebih mudah dan harga yang relatif murah menjadi alasan utama penggunaan pestisida sintetik. Namun, penggunaan senyawa kimia ini dalam jangka waktu yang lama dan kadar berlebihan dapat menyebabkan beberapa pengaruh negatif. Pestisida dapat berdampak merugikan pada kesehatan manusia dan lingkungan karena mencemari lahan pertanian dan menyebabkan produk pertanian tidak aman dikonsumsi (Yuliar 2008). Oleh karena itu, sangat dibutuhkan adanya agen pengendali hayati yang dapat digunakan sebagai jalur alternatif.
Penggunaan mikroorganisme sebagai agen biokontrol merupakan alternatif yang berpotensi tinggi dalam menangani penyakit tanaman (Kulkarni et al. 2007).
Trichoderma sp. merupakan salah satu mikroorganisme yang dapat digunakan sebagai agen biokontrol. Beberapa cendawan fitopatogen yang dapat dikendalikan oleh Trichoderma sp. antara lain Fusarium spp., Lentinus Lepidus, Phytium spp., Botrytis cinerea, Gloeosporium gloeosporoides, Rigidoporus lignosis, Sclerotium roflsii, dan Phytopthora palminovora (Benitez et al. 2004; Sunantapongsuk et al.
2006; Zeilinger dan Omann 2007). Penggunaan Trichoderma sp. sebagai agen biokontrol memiliki beberapa kelebihan, yaitu pertumbuhannya cepat, kapasitas reproduksi yang tinggi, spektrum penghambatan yang luas terhadap cendawan patogen, keragaman mekanisme penghambatan, kemampuan bertahan hidup yang tinggi dalam kondisi kurang nutrisi, dan menginduksi pertumbuhan tanaman (Benitez et al. 2004; Vinale et al. 2006).
Trichoderma sp. merupakan kapang yang bersifat kosmopolitan, saprofit, dan memiliki keragaman genetik yang tinggi. Berdasarkan penelitian sebelumnya,
Trichoderma sp. dari sampel tanah telah berhasil diisolasi oleh Peneliti di BALITRO. Selain itu, penelitian di BB-BIOGEN menunjukkan bahwa beberapa dari isolat tersebut menghasilkan selulase yang berpotensi mendegradasi limbah pertanian (Lestari et al. 2014). Anggota dari genus ini memiliki keragaman potensi yang bergantung pada genetik atau jenis spesiesnya. Penggunaan
2
Penelitian ini bertujuan mendapatkan isolat Trichoderma sp. yang dapat melawan patogen tanaman Colletotrichum sp. dan Rhizoctonia sp. serta mengidentifikasi isolat-isolat tersebut melalui analisis sekuen Internal TranscribedSpacer (ITS). Manfaat penelitian adalah isolat Trichoderma sp. yang memiliki kemampuan antagonis terhadap kedua patogen dapat digunakan sebagai agen biokontrol untuk mengatasi penyakit tanaman.
METODE
Alat dan Bahan
Alat-alat yang digunakan dalam penelitian ini adalah gelas piala, sudip, gelas ukur, labu Erlenmeyer (100, 250, 500, dan 500 mL), pipet mikro (10, 200, dan 1000 L), cawan Petri (3 dan 9 cm), saringan kasa, jarum ose, tabung Eppendorf, kertas saring Whatmann no.1, pengaduk, ruang kabinet (laminar air flow cabinet), dan penangas Bunsen. Selain itu, alat berupa mesin terdiri atas oven, kulkas, perangkat autoklaf, mesin vorteks, pH meter, mesin sentrifugasi, penangas air, termometer, neraca analitik, spektrofotometer nanodrop, mikroskop cahaya, dan mesin PCR.
Bahan-bahan yang digunakan yaitu isolat kapang, bahan media, reagen, dan marker. Bahan isolat kapang terdiri atas 17 isolat Trichoderma sp. yang merupakan koleksi BALITRO (Lampiran 2) dan isolat patogen (Colletotrichum sp.
dan Rhizoctonia sp.) yang merupakan koleksi BB BIOGEN. Bahan media terdiri atas kentang, D-glukosa, agar, dH2O, ddH2O, dekstrosa, dan serbuk agarosa. Reagen terdiri atas bufer ekstraksi (2 % CTAB; 20 mM EDTA; 0.1 M Tris-HCL; 1.4 M NaCl), 2-merkaptoetanol, bufer TE, fenol, kloroform, isoamil alkohol, iso-propanol, etil alkohol 70%, bufer TAE 0.5X, loading dye 1X, larutan
Etidium-Sebanyak 1 ose isolat cendawan dari kultur koleksi dipindahkan ke dalam media PDA (potatoes dextrose agar) baru. Peremajaan ini dilakukan secara steril dalam kabinet laminar air flow. Isolat yang telah dipindahkan kemudian dibiarkan tumbuh selama 3-5 hari pada suhu ruang.
Pengamatan Morfologi Trichoderma sp.
3 Identifikasi Molekuler berdasarkan Sekuen ITS
Isolasi DNA (Modifikasi Chakraborty et al. 2010). Pemecahan sel dilakukan secara mekanik menggunakan mortar dengan bantuan nitrogen cair.
Serbuk miselium dipindahkan ke dalam tabung Eppendorf dan diberi 500 L
bufer ekstraksi (2% CTAB; 20 mM EDTA; 0.1 M Tris-HCL; 1.4 M NaCl (pH 8.2); CTAB bufer : 2-merkaptoetanol = 100:1 (1% volume) (CTAB bufer 20 mL, 2-merkaptoetanol 200 L). Campuran dikocok dan diinkubasi pada suhu 65 oC selama 30 menit. Larutan kloroform : isoamil alkohol (24:1) ditambahkan sebanyak 400 L dan divorteks. Campuran disentrifugasi pada kecepatan 12.000 rpm selama 10 menit. Fase cair (bagian atas) dipindahkan ke tabung Eppendorf baru dan ditambahkan 400 L larutan kloroform : isopropanol (24:1) dingin dan dikocok perlahan. Campuran disentrifugasi dengan kecepatan 10.000 rpm selama 10 menit. Lapisan atas yang terbentuk dipindahkan ke tabung Eppendorf baru dan ditambahkan 500 L isopropanol dingin. Larutan diinkubasi pada suhu – 20 oC selama 1 jam, kemudian disentrifugasi pada 10.000 rpm selama 2 menit.
Supernatan dibuang dan pelet dikeringudarakan. Bufer TE 1X sebanyak
600 L dan campuran fenolμkloroformμisoamilalkohol (25μ24μ1) sebanyak 300 L
ditambahkan ke dalam tabung lalu dikocok menggunakan vorteks sebelum disentrifugasi pada 13.000 rpm selama 10 menit. Lapisan atas yang terbentuk
dipindahkan kembali ke tabung Eppendorf baru dan ditambahkan 500 L
isopropanol dingin. Larutan dikocok perlahan lalu diinkubasi pada -20 oC selama 1 jam. Setelah itu, larutan disentrifugasi pada 10.000 rpm selama 5 menit dan pelet hasil sentrifugasi dicuci dengan etanol 70 %. Larutan kemudian disentrifugasi pada 12.000 rpm selama 5 menit. Etanol dibuang dan pelet DNA dikering udarakan. Pelet DNA dilarutkan dengan bufer TE 1X lalu disimpan pada suhu –20 oC.
Analisis Kuantitatif DNA (Thermo Fisher Scientific 2009). DNA
Trichoderma sp. hasil isolasi diukur konsentrasi dan kemurniannya menggunakan spektrofotometer nanodrop. Pengaturan program dilakukan dan lubang optik dibersihkan menggunakan kertas tisu.Sebanyak 2 L bufer TE digunakan sebagai blanko dan dipipet ke dalam lubang optik spektrofotometer. Tombol read blank
pada komputer ditekan. Bufer TE yang tersisa di lubang optik dibersihkan dengan kertas tisu. Sebanyak 2 L sampel DNA dipipet ke dalam lubang optik lalu dibaca konsentrasinya dalam satuan ng/ L. Kemurnian DNA dapat dilihat berdasarkan
perbandingan nilai absorbansi sampel pada 260 dan 280 nm.
Analisis Kualitatif DNA (Sambrook dan Russel 2001). DNA hasil isolasi diseparasi menggunakan gel agarosa 1 % di dalam bufer TAE (Tris HCl-asam asetat-EDTA) 0.5 X. Sebanyak 2 L Loading dye dicampur dengan 3 L DNA, lalu diinjeksikan ke dalam sumur gel agarosa. Proses elektroforesis dilakukan selama 30 menit dengan menggunakan arus 50 mA dan voltase 60 V. Gel agarosa diangkat dan direndam dalam larutan EtBr selama 10-15 menit untuk proses pewarnaan lalu direndam kembali dalam akuades selama 5 menit. Gel agarosa diletakan ke dalam ruang UV-transluminator lalu diamati menggunakan ChemDoc.
Amplifikasi DNA Menggunakan PCR (Modifikasi Fu et al. 2012).
4
12.5 L DNA Taq Polimerase (Kappa 2G Fast Ready Mix), 1 L primer ITS1 (forward), 1 L primer ITS4 (reverse), 8.5 L ddH2O, dan 2 L cetakan DNA. Reaksi dengan mesin PCR dijalankan dengan program sebagai berikut, denaturasi awal pada 95 oC selama 5 menit; diikuti dengan 35 siklus untuk 1 menit denaturasi pada 95 oC, 35 detik penempelan primer pada 55 oC, dan 30 detik elongasi pada 72 oC; dan elongasi akhir selama 7 menit pada 72 oC. DNA hasil amplifikasi dianalisis melalui elektroforesis gel agarosa 1 % dalam bufer TAE 0.5 X, diwarnai dengan etidium bromida (EtBr), dan divisualisasi dengan bantuan sinar UV menggunakan ChemiDoc.
Analisis Homologi dan Filogenetik DNA Hasil Amplifikasi. Pengurutan basa nukleotida DNA dilakukan dengan mengirim sampel ke PT. Genetika Science, Jakarta. Hasilnya lalu dicocokkan dengan database yang berada di
GenBank dan dianalis menggunakan BLAST di website NCBI. Proses blast
dengan query nukleotida dipilih nr/nt (non-redundant nucleotide) (Zhang et al. 2000). Hubungan kekerabatan antar sekuen dan sekuen outgroup dianalisis menggunakan pohon filogenetik yang dibuat menggunakan program MEGA 5 (Tamura et al. 2011).
Uji Daya Hambat Isolat Trichoderma sp. terhadap Cendawan Patogen
Kemampuan antagonis Trichoderma sp. melawan Colletotrichum sp. dan
Rhizoctonia sp. secara in vitro dilakukan menggunakan metode uji berganda (Zivkovic et al. 2010). Tiga isolat Trichoderma sp. yang memiliki nilai PIRG tertinggi terhadap masing-masing patogen (Colletotrichum sp. dan Rhizoctonia
sp.) kemudian diuji inhibisi lanjut dengan perlakuan waktu inokulasi dan ulangan sebanyak tiga kali.
Perlakuan I dilakukan dengan menumbuhkan patogen terlebih dahulu lalu disusul dengan Trichoderma sp. setelah 24 jam. Perlakuan II dilakukan dengan menumbuhkan Trichoderma sp. terlebih dahulu lalu disusul dengan patogen setelah 24 jam. Hasilnya dianalisis menggunakan rancangan acak lengkap (RAL). Persentase penghambatan pertumbuhan patogen (PIRG) diukur dengan membandingkan jari-jari patogen perlakuan (R1) dan jari-jari patogen kontrol (R2) menggunakan rumus yang dikembangkan oleh Skidmore dan Dickinson (1976) sebagai berikut:
PIRG = 2- 1
2 x 100%
PIRG = percentage inhibition radius growth (%) R1= jari-jari patogen perlakuan (mm)
R2= jari-jari patogen kontrol (mm)
HASIL
Morfologi Isolat Trichoderma sp.
Pengamatan menggunakan mikroskop cahaya terhadap dua isolat
5
Gambar 1 Morfologi isolat Gam4 (A) dan PB2 (B)
berbentuk bulat seperti anggur berwarna hijau hingga keabuan. Banyaknya spora pada isolat berusia tiga hari menandakan tingkat pertumbuhan isolat yang relatif cepat. Jumlah spora antar masing-masing isolat kurang lebih sama menunjukkan kecepatan tumbuh yang seragam.
Morfologi makroskopik 17 isolat Trichoderma sp. secara umum memiliki permukaan rata menyerupai wol dengan tepian berbentuk regular. Sementara itu, dari 17 isolat Trichoderma sp., terdapat dua isolat berwarna putih, tujuh isolat berwarna kuning, dan 8 isolat berwarna hijau. Warna yang semakin hijau menandakan jumlah spora yang semakin banyak. Rerata pertumbuhan tiap isolat yaitu 8-9 cm dalam tiga hari.
DNA Total 17 isolat Trichoderma sp. dan Amplifikasi Sekuen ITS
Analisis kualitatif DNA hasil isolasi menggunakan elektroforesis gel agarosa 1 % dapat dilihat pada Gambar 2. Marker yang digunakan berada pada lajur paling kiri (M) yaitu marker ladder 1 Kb. Pita yang terdapat pada lajur no 1-17 menunjukkan DNA tiap isolat. Semua DNA berukuran lebih dari 10 000 kb dapat dilihat dari pita yang berada di atas marker. Kebanyakan DNA masih terkontaminasi oleh RNA atau pengotor polisakarida lainnya dapat dilihat dari adanya smear pada DNA.
Konsentrasi dan tingkat kemurnian DNA hasil isolasi yang diperoleh dari uji menggunakan spektrofotometer nanodrop dapat dilihat pada Tabel 1. Isolat
Pan23.1 memiliki konsentrasi terendah sebesar 215.7 ng/ L dan isolat T 4
memiliki konsentrasi tertinggi sebesar 827λ.2 ng/ L. Tingkat kemurnian DNA dapat dilihat dari perbandingan absorbansi pada 260 dan 280 nm. Isolat N34 memiliki tingkat kemurnian terendah sebesar 1.18. Kebanyakan isolat lainnya memiliki nilai A260/280 yang berada pada rentang 1.8-2.0. Hasil kuantitatif dan kualitatif tidak selalu menunjukkan hubungan yang positif jika dilihat dari Gambar 2 dan Tabel 1. Isolat TR4 memiliki nilai konsentrasi tertinggi namun pita hasil elektroforesis yang sangat tipis.
Gambar 2 DNA Trichoderma sp. hasil isolasi (M= Marker ladder 1 Kb; 1-17= O5, Pan23.1, Kun4, I4, Gam4, OA9, D4, Bo17, Pan23.2, PC5, TR4, RR9, PB2, G09, P03, N34, dan TR3)
A B
6
Tabel 1 Kuantitatif 17 isolat DNA Trichoderma sp.
No Isolat [asam nukleat] (ng/ L) A260/280 Pan23.2, PC5, TR4, RR9, PB2, G09, P03, N34, dan TR3)
Amplifikasi terhadap DNA hasil isolasi menggunakan primer ITS1 dan ITS 4 dapat dilihat pada Gambar 3. Lajur paling kanan merupakan marker 100 bp yang digunakan untuk menentukan ukuran dari tiap amplikon. Semua isolat
Trichoderma sp. yang diamplifikasi menghasilkan pita berukuran 600-700 bp dengan tingkat ketebalan yang cukup bervariasi. Ukuran pita yang seragam karena proses amplifikasi dilakukan pada daerah yang sama dengan ukuran pasang basa yang relatif sama.
Homologi dan Filogenetik Isolat Trichoderma sp.
Hasil penjajaran DNA yang telah diurutkan basa nukleotidanya dengan database di web NCBI dapat dilihat pada Tabel 2. Sebanyak 13 dari 17 isolat merupakan Trichoderma asperellum, sebanyak satu isolat merupakan
Trichoderma gamsii, satu isolat merupakan Trichoderma koningiopsis, dan dua sisanya merupakan Trichoderma harzianum. Kemiripan hasil sekuen semua isolat memiliki nilai 99-100 % dengan nilai E 0.0. Isolat Pan23.1, Pan23.2, dan Kun4 memiliki nilai kemiripan 100 % dan nilai E sebesar 0.0 dengan Trichoderma asperellum T8. Hal tersebut mengindikasikan tingkat kecocokan yang lebih baik dibandingkan hasil pencocokan yang lainnya.
7
Trichoderma virens, dan Trichoderma reesei. Ketiga anggota sekuen outgroup
berada pada cabang yang berbeda dan terletak cukup jauh dari 17 isolat
Trichoderma sp. asal Sukabumi. Sementara itu, 17 isolat Trichoderma sp. hasil sekuensing berada dalam satu cabang yang sama menunjukkan hubungan kekerabatan yang dekat antar 17 isolat Trichoderma sp.
Tabel 2 Identifikasi 17 isolat Trichoderma sp.
No Isolat Deskripsi Nilai
E
Kemiripan
(%) No. Aksesi
1 O5 Trichoderma asperellum isolat T8 0.0 99 JX422014.1
2 Pan23.1 Trichoderma asperellum isolatT8 0.0 100 JX422014.1
3 Kun4 Trichoderma asperellum isolatT8 0.0 100 JX422014.1
4 I4 Trichoderma asperellum isolatM29 0.0 99 JX422010.1
5 Gam4 Trichoderma asperellum UTP-16 0.0 99 FJ640576.1
6 OA9 Trichoderma asperellum isolat M29 0.0 99 JX422010.1
7 D4 Trichoderma asperellum TR11 0.0 99 KC859432.1
8 Bo17 Trichoderma asperellum TR11 0.0 99 KC859432.1
9 Pan23.2 Trichoderma asperellum T8 0.0 100 KC859432.1
10 PC5 Trichoderma asperellum TR11 0.0 100 KC859432.1
11 TR4 Trichoderma gamsii UNISS 11-62 0.0 99 EF488141.1
12 RR9 Trichoderma asperellum UTP-16 0.0 99 FJ640576.1
13 PB2 Trichoderma harzianum isolat A1S4-2 0.0 99 KJ767087.1
14 G09 Trichoderma asperellum isolatM29 0.0 99 JX422010.1
15 P03 Trichoderma koningiopsis F-2-46 0.0 99 KF751670.1
16 N34 Trichoderma harzianum LIPIMC0572 0.0 99 KC847182.1
17 TR3 Trichoderma asperellum TR11 0.0 99 KC859432.1
8
Kemampuan Antagonis Trichoderma sp. terhadap Colletotrichum sp. dan
Rhizoctonia sp. secara in vitro
Hasil uji daya hambat 17 isolat Trichoderma sp. terhadap kedua patogen dapat dilihat pada Tabel 3. Nilai persentase penghambatan Trichoderma sp.
terhadap Colletotrichum sp. terendah yaitu 51.72 % ditunjukkan oleh isolat I4, Bo17, dan PB2. Sementara itu, Pan23.1 merupakan satu-satunya isolat yang memiliki nilai penghambatan > 80 % dan memiliki daya hambat tertinggi yaitu 82.76 %. Nilai penghambatan Trichoderma sp. terhadap Rhizoctonia sp. yang terendah sebesar 60.00 % (isolat OA9) dan tertinggi sebesar 92.50 % (isolat TR3). Isolat Trichoderma sp. yang memiliki nilai penghambatan > 80 % terhadap
Rhizoctonia sp. yaitu Kun4, Pan23.2, RR9, N34, dan TR3. Nilai penghambatan >80 % menunjukkan isolat yang berpotensial sebagai biokontrol melawan patogen.
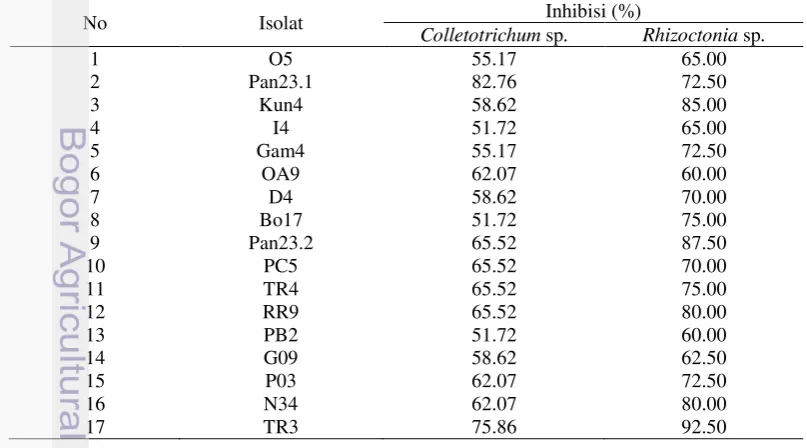
Jari-jari patogen kontrol (Gambar 5.A) jauh lebih besar dibandingkan dengan jari-jari di cawan perlakuan (Gambar 5.B). Selain itu, isolat Pan23.1 dapat menutupi seluruh permukaan Colletotrichum sp. yang tumbuh pada cawan perlakuan. Hal tersebut menunjukkan patogen Colletotrichum sp. terhambat oleh kehadiran Trichoderma sp. (isolat Pan23.1). Penghambatan pertumbuhan
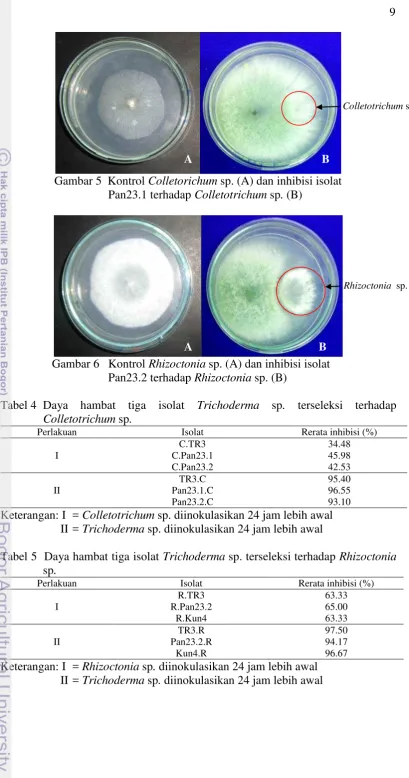
Rhizoctonia sp. juga terjadi saat kehadiran Trichoderma sp. isolat Pan23.2 (Gambar 6).
Tiga isolat Trichoderma sp. yang memiliki nilai hambat tertinggi terhadap
Colletotrichum sp. yaitu TR3, Pan23.1, dan Pan23.2. Ketiga isolat tersebut kemudian diuji lebih lanjut yang hasilnya dapat dilihat pada Tabel 4. Isolat C.TR3 memiliki nilai rerata inhibisi terendah sebesar 34.48 % dan isolat Pan23.1.C memiliki nilai rerata inhibisi tertinggi sebesar 96.55 %. Sementara itu, isolat TR3, Pan23.2, dan Kun4 merupakan tiga isolat yang memiliki nilai penghambatan tertinggi terhadap Rhizoctonia sp. Nilai hambat terendah ditunjukkan oleh isolat R.Kun4 dan R.TR3 sebesar 63.33 % sedangkan nilai hambat tertinggi ditunjukkan oleh isolat TR3.R sebesar 97.50 % (Tabel 5).
Tabel 3 Daya hambat 17 isolat Trichoderma sp. terhadap cendawan patogen
9
Gambar 5 Kontrol Colletorichum sp. (A) dan inhibisi isolat Pan23.1 terhadap Colletotrichum sp. (B)
Gambar 6 Kontrol Rhizoctonia sp. (A) dan inhibisi isolat Pan23.2 terhadap Rhizoctonia sp. (B)
Tabel 4 Daya hambat tiga isolat Trichoderma sp. terseleksi terhadap
Colletotrichum sp.
Perlakuan Isolat Rerata inhibisi (%)
C.TR3 34.48
I C.Pan23.1 45.98
C.Pan23.2 42.53
TR3.C 95.40
II Pan23.1.C 96.55
Pan23.2.C 93.10
Keterangan: I = Colletotrichum sp. diinokulasikan 24 jam lebih awal II = Trichoderma sp. diinokulasikan 24 jam lebih awal
Tabel 5 Daya hambat tiga isolat Trichoderma sp. terseleksi terhadap Rhizoctonia
sp.
Perlakuan Isolat Rerata inhibisi (%)
R.TR3 63.33
I R.Pan23.2 65.00
R.Kun4 63.33
TR3.R 97.50
II Pan23.2.R 94.17
Kun4.R 96.67
Keterangan: I = Rhizoctonia sp. diinokulasikan 24 jam lebih awal II = Trichoderma sp. diinokulasikan 24 jam lebih awal
A B
Colletotrichum sp.
Rhizoctonia sp.
10
Gambar 7 Inhibisi isolat R.Pan23.2 (A) dan C.TR3 (B)
Terdapat perbedaan yang signifikan antara perlakuan I dan II untuk kedua patogen. Persentase penghambatan Trichoderma sp. jauh lebih rendah bila patogen diinokulasikan terlebih dahulu. Berdasarkan uji Anova (P<0.05), perbedaan waktu inokulasi memberi pengaruh yang nyata terhadap nilai penghambatan patogen. Perubahan warna pada patogen (Gambar 7) mengindikasikan adanya metabolit sekunder yang dihasilkan baik oleh
Trichoderma sp. dalam melakukan aksi penghambatan maupun oleh patogen sebagai pertahanan terhadap serangan Trichoderma sp.
PEMBAHASAN
Morfologi Isolat Trichoderma sp.
Morfologi makroskopik yang dilakukan terhadap 17 isolat Trichoderma sp.
pada usia tiga hari berupa bentuk tepian dan elevasi serta warna isolat. Semua isolat memiliki tepian yang berwarna putih dengan elevasi rata dan tepian regular. Pengamatan belum bisa digunakan untuk mengidentifikasi isolat karena hasilnya tidak jauh berbeda antar masing-masing isolat. Hasil penelitian Gomez et al.
(1997) juga menunjukkan hasil yang sangat mirip terhadap pengamatan makroskopik Trichoderma sp., meliputi laju pertumbuhan, warna spora, dan pola penyebaran spora. Sifat morfologi dan fisiologi dari Trichodema sp. adalah sama (Soesanto et al. 2011). Beberapa isolat dapat menghasilkan pigmen warna kuning yang berdifusi ke dalam media agar, yaitu PB2, Pan23.1, N34, dan Kun4. Hasil penelitian sebelumnya (Khang et al. 2013) menunjukkan Trichoderma sp. memiliki permukaan yang rata dengan struktur lembut seperti wol, spora hijau yang menyebar, dan pigmen kuning-hijau yang berdifusi ke dalam media tumbuh. Bentuk spora, bentuk konidiofor, dan tingkat percabangan dari konidiofor diamati menggunakan mikroskop cahaya. Hampir semua isolat menunjukkan percabangan konidiofor yang cukup tinggi dan berbentuk seperti piramida. Hal tersebut sesuai dengan hasil morfologi pertama yang diklasifikasikan oleh Rifai (1969) bahwa percabangan konidiofor Trichoderma sp. sangat tinggi dengan pola menyerupai piramida atau konikal. Spora yang teramati berwarna hijau hingga keabuan berbentuk bulat dan menyebar di sekitar konidiofor. Athul dan Jisha (2013) juga menyatakan bahwa konidia Trichoderma sp. berbentuk bulat hingga elips, uniselular, dan dihasilkan dari konidiofor yang bercabang seperti susunan piramida.
11 DNA Total 17 Isolat Trichoderma sp. dan Amplifikasi Sekuen ITS
Isolasi DNA dilakukan menggunakan metode Cetyl Trimethyl Ammonium Bromide (CTAB) yang dimodifikasi dengan penambahan 2-merkaptoetanol. Pemilihan tersebut didasarkan pada kemampuan CTAB sebagai salah satu metode umum dalam mengekstraksi DNA yang banyak mengandung polisakarida dan senyawa polifenol (Jose dan Usha 2000). Bufer ekstraksi CTAB berfungsi melisiskan membran sel dan fosfolipid bilayer (Syafaruddin dan Santoso 2011), kandungan garam di dalamnya juga dapat memisahkan polisakarida dari dinding sel (Surzycki 2000; Santoso 2005).
Tingkat kemurnian DNA isolat Kun4, I4, Gam4, OA9, D4, PC5, RR9, PB2, G09, dan P03 dapat dikatakan murni karena rasio A260/280 berada pada rentang 1.8-2.0 (Sambrook dan Russel 2001). Nilai tersebut dapat diperoleh karena metode ekstraksi berulang (dua kali) sehingga presipitasi DNA lebih maksimal (Mulyani et al. 2012). Namun, isolat O5, Pan23.1, Bo17, Pan23.2, dan N34 memiliki nilai rasio A260/280 < 1.8 yang menunjukkan DNA tidak murni. Hal serupa juga terjadi pada isolat TR4 dan TR3 memiliki nilai A260/280 > 2.0. Hampir seluruh isolat memiliki pita DNA yang kurang baik atau smear (Gambar 3). Selain itu, terdapat beberapa pita yang sangat tipis (lajur no 5, 10, 11, 14, dan 15). Hal tersebut diduga karena ekstraksi DNA tidak menggunakan enzim ribonuklease (RNAse) sehingga masih terdapat kontaminan, misalnya RNA masih mengkontaminasi DNA dan menyebabkan hasil yang kurang murni (Syafaruddin dan Santoso 2011).
Teknik yang mendasari proses amplifikasi ini adalah Polymerase Chain Reaction (PCR) pada daerah Internal Transcribed Spacer (ITS). Daerah ITS meliputi ITS1 dan ITS2 yang dipisahkan oleh gen 5.8S, terletak di antara gen 18S dan 28S (Bellemain et al. 2010). Pemilihan PCR-ITS disebabkan sekuen tersebut memiliki spektrum yang luas dalam mengidentifikasi berbagai cendawan. Proses amplifikasi DNA menggunakan primer ITS1 dan ITS4 yang bekerja pada tiga daerah, yaitu ITS1, ITS2, dan gen 5.8S. Sekuen DNA pada ketiga daerah tersebut memiliki tingkat variasi yang tinggi dan dapat digunakan untuk mengidentifikasi inter- dan intra-spesies. ITS1 dan ITS4 merupakan primer universal yang dapat mengamplifikasi dan mengidentifikasi banyak fungi, salah satunya yaitu
Trichoderma sp. (Singh et al. 2014; Chakraborty et al. 2010).
Pita DNA hasil amplifikasi memiliki ketebalan yang bervariasi namun ukurannya berada pada rentang 600-700 bp untuk semua isolat Trichoderma sp.
Hal tersebut sesuai dengan beberapa penelitian yang telah dilakukan sebelumnya (Chakraborty et al. 2010; Chakraborty et al. 2011; Sagar et al. 2011; Fu et al.
2012) bahwa DNA Trichoderma sp. yang diamplifikasi dengan primer ITS1 dan ITS4 menghasilkan amplikon berukuran 400-650 bp. Rentang ukuran amplikon bergantung pada komposisi basa nukleotida yang dapat berbeda pada tiap spesies
Trichoderma sp.
Homologi dan Filogenetik Trichoderma sp.
12
dengan database untuk Trichoderma asperellum terbagi menjadi empat jenis yaitu galur TR11, galur UTP-16, isolat T8, dan isolat M29. Sementara itu, Trichoderma harzianum terbagi menjadi 2 jenis yaitu isolat A1S4-2 dan galur LIPIMC0572. Hasil yang diperoleh dapat dikatakan baik karena memiliki nilai E 0.0 untuk 17 isolat. Semakin rendah nilai E menunjukkan semakin tinggi tingkat homologi antara kedua sekuen. Nilai E merupakan nilai dugaan terhadap kedua sekuen berdasarkan perhitungan statistika (Claverie dan Notredame 2003).
Sebanyak 76.47 % dari 17 total isolat yang diidentifikasi merupakan
Trichoderma asperellum dan 11.76 % merupakan Trichoderma harzianum.
Sisanya, masing-masing sebesar 5.8 % merupakan Trichoderma gamsii galur UNISS 11-62 dan Trichoderma koningiopsis galur F-2-46. Hal ini sesuai dengan penelitian yang telah dilakukan sebelumnya oleh Druzhinina et al. (2006) bahwa
Trichoderma asperellum merupakan isolat terbanyak yang ditemukan pada daerah tropikal. Hoyos-Carvajal et al. (2009) juga menyatakan bahwa spesies umum yang ditemukan pada daerah neotropikal adalah Trichoderma asperellum diikuti dengan Trichoderma harzianum.
Hubungan kekerabatan antara 17 isolat Trichoderma sp. yang telah disejajarkan dengan database di NCBI dianalisis menggunakan pohon filogenetik. Tiga sekuen outgroup yang dipilih merupakan jenis dari Trichoderma sp. yang berpotensi sebagai agen biokontrol. Hal tersebut bertujuan membandingkan hubungan kekerabatan jenis spesies yang telah teridentifikasi dengan agen biokontrol lainnya. Isolat Kun4, Pan23.1, O5, dan Pan23.2 memiliki hubungan kekerabatan yang sangat dekat dengan T. asperelllum isolat T8 (India) dapat dilihat pada Gambar 4. Isolat I4 memiliki kedekatan dengan T. asperellum isolat M29 (India) dan P03 memiliki kedekatan dengan T. koniongiopsis (China). Sementara sisanya, memiliki kedekatan antar sesama isolat Trichoderma sp. yang berasal dari Sukabumi. Menurut Schuster dan Schmoll (2010), Trichoderma sp. merupakan fungi soilborne yang dapat ditemukan di seluruh dunia sehingga memungkinkan isolat antar negara memiliki hubungan kekerabatan yang dekat.
Kemampuan Antagonis Trichoderma sp. terhadap Colletotrichum sp. dan
Rhizoctonia sp.
Kemampuan antagonis dianalisis menggunakan metode uji berganda dengan mengukur persentase inhibisi berdasarkan perbandingan jari-jari patogen perlakuan dan kontrol. Nilai hambat tersebut diukur untuk mengetahui pengaruh penghambatan cendawan antagonis Trichoderma sp. terhadap pertumbuhan koloni patogen. Pertumbuhan Colletotrichum sp. dan Rhizoctonia sp. mulai terlihat terhambat pada hari kedua setelah diinokulasi. Luas koloni Colletotrichum sp. dan
Rhizoctonia sp. lebih rendah dibandingkan luas koloni Trichoderma sp. Hal tersebut disebabkan laju pertumbuhan Trichoderma sp. yang relatif cepat (>8-9 cm setelah 3 hari). Consolo et al. (2012) juga menyatakan bahwa kebanyakan anggota dari Trichoderma sp. tumbuh cepat dalam media kultur dan memproduksi banyak konidia putih ataupun hijau.
13 lebih tinggi dibandingkan dengan perlakuan I. Kehadiran Trichoderma sp. yang lebih awal memungkinkan Trichoderma sp. memproduksi senyawa-senyawa yang dapat menyerang cendawan patogen. Mereka dapat memproduksi beberapa enzim
ektstraselular seperti kitinase, β-1,3 glukanase, protease (Chet dan Chernin 2002; Viterbo et al. 2002), β-1,4 glukanase dan lipase (Benitez et al. 2004; Diby et al.
2005; Vinale et al. 2008). Hal tersebut memperkuat peranan Trichoderma sp.
sebagai agen biokontrol karena kitin dan β-1,3 glukan merupakan komponen utama penyusun dinding sel patogen (Kullnig et al. 2000; Kubicek et al. 2001). Selain itu, protease dan lipase yang dihasilkan juga dapat melisis protein dan lipid yang terdapat dapat dinding sel patogen.
Perubahan warna patogen Rhizoctonia sp. dan Colletotrichum sp. dari putih menjadi kekuningan (Gambar 7) diduga akibat adanya produksi metabolit sekunder dari Trichoderma sp. sebagai bagian dari mekanisme pertahanan. Menurut Mukherjee (2011), Trichoderma sp. ketika berada berdekatan dengan patogen dapat menimbulkan respon mikoparasitik yang diawali dengan penarikan (attraction), pelekatan (attachment), pelilitan (coiling), dan penghancuran hifa patogen yang berujung pada kematian patogen. Proses tersebut dapat memicu sekresi enzim-enzim yang mendegradasi dinding sel patogen (Kubicek et al. 2011). Selain enzim hidrolitik, metabolit sekunder lain dihasilkan oleh
Trichoderma sp. yaitu antibiotik antifungi (Barea et al. 2005; Vinale et al. 2008), keduanya merupakan senjata utama dalam membunuh patogen.
Nilai penghambatan Trichoderma sp. terhadap Rhizoctonia sp. lebih tinggi dibandingkan Colletotrichum sp. pada perlakuan waktu inokulasi maupun tidak. Penghambatan yang dapat dilakukan oleh Trichoderma sp. yaitu kompetisi hara dan ruang (Lo 1998; Benitez et al. 2004) serta mikoparasitisme (Howell 2003).
Colletotrichum sp. termasuk ke dalam kelas Deuteromycetes sedangkan
Rhizoctonia sp. merupakan anggota Basidiomycetes. Menurut Monte (2001) dan Benitez et al. (2004), Trichoderma spp. dapat mengontrol pertumbuhan patogen yang termasuk ke dalam Basidiomycetes. Hal tersebut mendukung daya hambatnya yang lebih tinggi terhadap Rhizoctonia sp.
SIMPULAN DAN SARAN
Simpulan
Seluruh isolat memiliki karakteristik morfologi yang sama dengan permukaan rata seperti wol, elevasi regular, dan warna konidia putih hingga hijau. Bentuk konidiofor yang teramati melalui mikroskop cahaya menyerupai piramid dan memiliki percabangan yang cukup tinggi sedangkan konidia berbentuk bulat. Hasil identifikasi terhadap 17 isolat menunjukkan tingkat kemiripan yang sangat tinggi terhadap Trichoderma asperellum (13 isolat), Trichoderma harzianum (2 isolat), Trichoderma gamsii (1 isolat), dan Trichoderma koningiopsis (1 isolat). Secara keseluruhan, Trichoderma sp. lebih berpotensi sebagai anti fitopatogen terhadap Rhizoctonia sp. dibandingkan Colletotrichum sp. Isolat Trichoderma sp.
14
Saran
Pengujian sinergis antar isolat Trichoderma sp. yang memiliki penghambatan terhadap kedua patogen. Selain itu, perlu dilakukan analisis dan identifikasi lebih lanjut terhadap metabolit sekunder yang dihasilkan oleh masing-masing isolat Trichoderma sp. dan uji in vivo menggunakan tanaman pangan.
DAFTAR PUSTAKA
Athul SR, Jisha MS. 2013. Screening of Trichoderma spp and Pseudomonas spp. for their biocontrol potential against phytopathogens of Vanilla. Int. J. of. Agriculture, Environment & Biotechnology 6: 799-806.
Barea JM, Poso MJ, Azcon R, Aguilar CA. 2005. Microbial co-operation in the rhizosphere. Journal of Experimental Botany: 1-18.
Benıtez T, inconAM, Limon MC, Codon AC. 2004. Biocontrol mechanisms of
Trichoderma strains. International Microbiology 7: 249–260.
Chakraborty BN, Chakraborty U, Saha A, Sunar K, Dey PL. 2010. Molecular characterization of Trichoderma viride and Trichoderma harzianum
isolated from soils of North Bengal based on rDNA markers and analysis of their PCR-RAPD profiles. Global Journal of Biotechnology and Biochemistry 5(1): 55-61.
Chakraborty BN, Chakraborty U, Sunar K, Dey PL. 2011. RAPD profiles and rDNA sequence analysis of Talaromyces flavus and Trichoderma sp.
Indian Journal of Biotechnology 10: 487-495.
Chet I, Chernin L. 2002. Biocontrol microbial agents in soil. Bitton G (ed) Encyclopedia of Environmental Microbiology 1: 450–465
Claverie JM, Notredame C. 2003. Bioinformatics for Dummies. Indianapolis (USA): Wiley Publishing.
Consolo VF, Monaco CI, Cordo CA, Salerno GL. 2012. Characterization of novel
Trichoderma spp. isolates for effective biocontrollers of fungal diseases of economic important crops in Argentina. World J Mivrobiol Biotechnol 28: 1389-1398.
Diby P, Sju KA, Jisha PJ, Sarma YR, Kumar A, Anandaraj M. 2005. Mycolytic enzyme produced by Pseudomonas fluorescens and Trichoderma spp. against Phytophthora capsici, the foot rot pathogen of black pepper (Piper ningrum, L.). Annals of Microbiology 55(2) : 129-133.
Druzhinina IS, Kopchinskiy AG, Kubicek CP. 2006. The first one hundred of
Trichoderma species characterized by molecular data. Mycoscience 47: 55– 64.
15 Gams W, Bissett J. 1998. Morphology and identification of Trichoderma. In Trichoderma and Gliocladium (eds. Kubicek CP and Harman GE). London (USA): Taylor and Francis.
Gomez I, Chet I, Herrera-Estrella A. 1997. Genetic diversity and vegetative compatibility among Trichoderma harzianum isolates. Mol. Gen. Genet. 256: 127-135.
Herwidyarti KH, Ratih S, Sembodo DRJ. 2013. Keparahan penyakit antraknosa pada cabai (Capsicum annuum L) dan berbagai jenis gulma. J. Agrotek Tropika 1(1): 102-106.
Howell CR. 2003. Mechanisms employed by Trichoderma species in the biological control of plant diseases : the history and evolution of current concepts. Plant Disease 87(1): 4-9.
Hoyos-Carvajal L, Orduz S, Bissett J. 2009. Genetic and metabolic biodiversity of
Trichoderma from Colombia and adjacent neotropic regions. J. Fungal Genetics and BiologyV 46: 615–631.
Jose J, Usha R. 2000. Extraction of geminiviral DNA from a highly mucilaginous plant (Abelmoschus esculentus). Plant Mol. Biol. Rep. 18: 349 -355.
Kubicek CP, Mach RL, Peterbauer CK, Lorito M. 2001. Trichoderma: from genes to biocontrol. J Plant Pathol 83:11–23
Kubicek CP, Herrera-Estrella A, Seidl-Seiboth V, Martinez DA. 2011. Comparative genome sequence analysis underscores mycoparasitism as the ancestral life style of Trichoderma. Genome Biol 12: R40.
Kulkarni M, Chaudhari R, Chaudhari A. 2007. Novel tension-active microbial compounds for biocontrol aplicatins. In: General Concepts in Integrated Pest and Disease Management (eds. Ciancio A and Mukerji KG). Springer:
295-304.
Khang VT, Anh NTM, Tu PM, Tham NTH. 2013. Isolation and selection of
Trichoderma spp. exhibiting high antifungal activities against major pathogens in Mekong Delta. Omonrice 19: 159-171.
Lo CT. 1998. General mechanisms of action of microbial biocontrol agents. Plant Pathology Bulletin 7:155-166.
Mulyani Y, Purwanto A, Nurruhwati I. 2012. Perbandingan beberapa metode isolasi DNA untuk deteksi dini Koi Herpes Virus (KHV) pada ikan mas (Cyprinus carpio L.). Jurnal FPIK, siap terbit.
Mukherjee PK. 2011. Genomics of biological control-whole genome sequencing of two mycoparasitic Trichoderma spp. Curr Sci 101:268.
Monte E. 2001. Understanding Trichoderma: between biotechnology and microbial ecology. Int Microbiol 4: 1–4.
16
Sagar MSI, Meah MB, Rahman MM, Ghose AK. 2011. Determination of genetic variations among different Trichoderma isolates using RAPD marker in Bangladesh. J. Bangladesh Agric. Univ. 9(1):9-20
Sambrook J, Russel DW. 2001. Molecular Cloning: A Laboratory Manual 3rd Edition. New York (USA): Cold-Spring Harbor Laboratory Press.
Santoso PJ. 2005. Modified CTAB-based DNA isolation procedure for fruit crops.
Jurnal Stigma XIV(1):1-4.
Schuster A, Schmoll M. 2010. Biology and biotechnology of Trichoderma. Appl. Microbiol Biotechnol 87: 787-799.
Singh A, Shahid M, Srivastava M. 2014. Genetic relationship of Trichoderma asperellum Tasp/8940 using Internal Transcribed Spacer (ITS) sequences.
International Journal of Advanced Research 2(3): 979-986.
Shahid M, Srivastava M, Sharma A, Kumar V, Pandey S, Singh A. 2013. Morphological, molecular indentification and SSR marker analysis of a potential strain of Trichoderma/Hypocrea for production of bioformulation.
JPlant Pathol Microb (4)10: 204-211.
Skidmore AM, Dickinson CH. 1976. Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi. Trans. Brit. Mycol. Soc. 66:57-64.
Soesanto L, Utami DS, Rahayuniati RF. 2011. Morphological characteristics of four Trichoderma isolates and two endophytic Fusarium isolates. Can J Sci. and Industrial Res. 2(8): 294-304.
Sunantapongsuk V, Nakapraves P, Piriyaprin Sm Manoch L. 2006. Protease production and phosphate solubilisation from potential biological control agents Trichoderma viride and Azomonas agilis from Vetiver rhizosphere.
J. Land Development Department Thailand: 1-4.
Surzycki S. 2000. Basic Techniques in Molecular Biology. New York (USA): Springer-Verlag.
Syafaruddin, Santoso TJ. 2011. Optimasi teknik isolasi dan purifikasi DNA yang efisien dan efektif pada kemiri sunan (Reutalis trisperma (Blanco) Airy Shaw). Jurnal Litri 17(1): 11-17.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol.
28: 2731-2739.
Thermo Fisher Scientific. 2009. Nanodrop 2000/200c Spectrophotometer V1.0 User Manual. Wilmington (USA): Thermo Fischer Scientific.
Vinale F, Marra R, Scala F, Ghisalbert EL, Lorito M, Sivasithamparam K. 2006. Major secondary metabolites produced by two comercial Trichoderma
17 Vinale F, Sivasithamparam K, Ghisalberti EL, Marra R, Woo SL, Lorito M. 2008.
Trichoderma– plant–pathogen interactions. Review Article. Soil Biology & Biochemistry 40: 1–10.
Viterbo A, Ramot O, Chermin LY, Chet I. 2002. Significance of lytic enzymes from Trichoderma spp. in the biocontrol of fungal plant pathogens. Anton Leeuw Int J G 81:549–556
Yu GY, Sinclair JB, Hartman GL,Bertagnolli BL. 2002. Production of iturin A by
Bacillus amyloliquefaciens suppressing Rhizoctonia solani. Soil Biology and Biochemistry 34: 955-963.
Yuliar. 2008. Skrining bioantagonistik bakteri untuk agen biokontrol Rhizoctonia solani dan kemampuannya dalam menghasilkan surfaktin. Biodiversitas
9(2): 83-86.
Zeilinger S, Omann M. 2007. Trichoderma biocontrol: signal transduction pathways involved in host sensing and mycoparasitism. Gene Regulation and Systems Biology 1: 227-234.
Zhang Z, Schwartz S, Wagner L, Miller W. 2000. A greedy algorithm for aligning DNA sequences. J Comput Biol. 7: 201-214.
Zivkovic S, Stojanovic S, Ivanovic Z, Gavrilovic V, Popovic T, Balaz J. 2010. Screening of antagonistic activity of microorganisms against
18
Lampiran 1 Diagram alir penelitian
Peremajaan isolat Trichoderma sp.
Isolasi DNA genom
Uji kuantitatif dan kualitatif DNA genomik: spektrofotometer nanodrop dan elektroforesis
Elektroforesis hasil amplifikasi Amplifikasi DNA sekuen ITS
Pengurutan basa nukleotida DNA
Pensejajaran dengan program blastn pada web NCBI.
Uji daya hambat Trichoderma sp. terhadap
Colletotrichum sp. dan Rhizoctonia sp.
Pengamatan morfologi makroskopik dan mikroskopik
19 Lampiran 2 Identitas 17 isolat Trichoderma sp.
No Kode Isolat Asal
1 O5 Tanah Sukabumi
2 Pan23.1 Tanah Sukabumi
3 Kun4 Tanah Sukabumi
4 I4 Tanah Sukabumi
5 Gam4 Tanah Sukabumi
6 OA9 Tanah Sukabumi
7 D4 Tanah Sukabumi
8 Bo17 Tanah Sukabumi
9 Pan23.2 Tanah Sukabumi
10 PC5 Tanah Sukabumi
11 TR4 Tanah Sukabumi
12 RR9 Tanah Sukabumi
13 PB2 Tanah Sukabumi
14 G09 Tanah Sukabumi
15 P03 Tanah Sukabumi
16 N34 Tanah Sukabumi
20
Lampiran 3 Contoh hasil pengurutan basa nukleotida Trichoderma sp.
Isolat D4
21 Lampiran 4 Hasil uji daya hambat lanjut isolat Trichoderma sp. terseleksi
Trichoderma sp. terhadap Colletotrichum sp. dengan perlakuan waktu inokulasi
Isolat Ulangan Inhibisi (%) Rerata inhibisi (%)
C. TR 3 1 37.93
Trichoderma sp. terhadap Rhizoctonia sp. dengan perlakuan waktu inokulasi
Isolat Ulangan Inhibisi (%) Rerata inhibisi (%)
22