KERAGAAN SEGREGAN F2 PADI (
Oryza sativa
L.) DI LAHAN
KERING
BENNI SITUMORANG
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
*Dengan ini saya menyatakan bahwa skripsi berjudul “Keragaman Segregan
F2 Padi (Oryza sativa L.) di Lahan Kering” adalah benar karya saya dengan arahan dari pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada institut Pertanian Bogor.
Bogor, Desember 2015
Benni Situmorang NIM A24110033
________________________
ABSTRAK
BENNI SITUMORANG. Keragaman Segregan F2 Padi (Oryza sativa L.) di Lahan Kering. Dibimbing oleh DESTA WIRNAS dan HAJRIAL ASWIDINNOOR.
Penyediaan pangan dalam jumlah yang cukup dan terjangkau harus menjadi prioritas pembangunan nasional. Hal ini disebabkan laju pertumbuhan penduduk yang sangat cepat dengan laju konsumsi beras melebihi rata-rata tingkat konsumsi dunia. Produksi beras nasional mengalami penurunan sebesar 0.45 juta ton pada tahun 2014 dari tahun sebelumnya sehingga diperlukan strategi untuk meningkatkan produksi beras nasional yang masih berfluktuasi. Varietas unggul yang berdaya hasil tinggi hasil pemuliaan diharapkan mampu menjadi teknologi kunci untuk meningkatkan produktivitas padi nasional. Penelitian ini bertujuan untuk menduga keragaman genetik, skewness dan kurtosis, nilai heritabilitas arti luas, koefisien keragaman genetik, dan koefisien korelasi antar karakter dua populasi F2 padi. Penelitian dilaksanakan di Kebun Percobaan Leuwikopo IPB pada November 2014 hingga April 2015. Percobaan dilakukan pada lahan kering dengan menanam dua populasi F2 hasil persilangan IR64 x Situ Patenggang dan IR64 x Mekongga. Nilai duga heritabilitas dan koefisien keragaman genetik memiliki nilai yang berbeda-beda pada kedua persilangan. Jumlah anakan 45 HST, jumlah gabah bernas malai-1, jumlah gabah hampa malai-1, bobot 100 butir dan
jumlah gabah total tanaman-1 memiliki nilai KKG dan heritabilitas yang luas hingga
sedang. Deteksi segregan transgresif dilakukan dengan seleksi langsung berdasarkan satu karakter seleksi, yaitu jumlah gabah bernas malai-1. Terdapat 128
individu hasil seleksi berdasarkan satu karakter pada persilangan IR64 x Situ Patenggang dan 132 individu pada persilangan IR64 x Mekongga.
ABSTRACT
BENNI SITUMORANG. Segregant Diversity of F2 Rice (Oryza sativa L) on Dry Land. Supervised by DESTA WIRNAS and HAJRIAL ASWIDINNOOR.
Food sufficiency is a priority of national development. This is due to the population growth is very high with the rate of rice consumption exceeds the average world consumption. Indonesian rice production actually decreased by 0.45 million tonnes in 2014. It is needed strategies to increase national rice production. High-yielding varieties are expected to become a key of technology to increase national rice production. This study was aimed to estimate genes action, broad sense heritability, coefficient of genetic variability, and the correlation coefficient of two F2 rice populations. The experiment was conducted at the experimental Leuwikopo IPB in November 2014 until April 2015. Experiments conducted on dry land by planting two F2 populations from crosses IR64 x Situ Patenggang and IR64 x Mekongga. There are differences in heritability estimates and coefficient of genetic variability in both crosses. Number of tillers 45 HST, the number of pithy grain panicle-1, the number of empty grain panicle-1, weight of 100 grains and grain total
number plant-1 have a wide to moderate of KKG and heritability. Segregant
transgressive detection is done by direct selection based on one characters, that is the number of pithy grain panicle-1. There are 128 individuals results of the
selection based on one characters from IR64 x Situ Patenggang and there are 132 indiviuals from IR64 x Mekongga.
KERAGAMAN SEGREGAN F2 PADI (
Oryza sativa
L.) DI
LAHAN KERING
BENNI SITUMORANG
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Agronomi dan Hortikultura
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Skripsi : Keragaman Segregan F2 Padi (Oryza sativa L.) di Lahan Kering
Nama : Benni Situmorang
NIM : A24110033
Disetujui oleh
Dr Desta Wirnas, SP, MSi Dr Ir Hajrial Aswidinnoor, MSc
Pembimbing I Pembimbing II
Diketahui oleh
Dr Ir Sugiyanta, MSi Ketua Departemen
PRAKATA
Puji serta syukur penulis panjatkan kepada Tuhan Yang Maha Kuasa atas limpahan rahmat serta karunia-Nya sehingga penulis dapat menyelesaikan skripsi
yang berjudul “Keragaman Segregan F2 Padi (Oryza Sativa) di Lahan Kering”. Tujuan dilakukannya penelitian ini adalah untuk menduga keragaman genetik beberapa persilangan padi. Penelitian ini berlangsung sejak bulan November 2014 hingga bulan April 2015 di Kebun Percobaan Leuwikopo IPB, Dramaga, Bogor.
Penulis mengucapkan terima kasih kepada Ibu Dr Desta Wirnas, SP MSi selaku dosen pembimbing akademik sekaligus pembimbing skripsi dan Bapak Dr Ir Hajrial Aswidinnoor, MSc juga selaku dosen pembimbing skripsi yang telah memberikan masukan dan saran selama kegiatan penelitian dan penyusunan skripsi ini. Ungkapan terima kasih juga penulis sampaikan kepada ayah, ibu, serta seluruh keluarga atas dukungannya baik dalam bentuk moral, materi, serta doa. Penulis juga mengucapkan terima kasih kepada teman-teman Agronomi dan Hortikultura yang telah memberi saran, semangat dan motivasi kepada penulis dan terima kasih kepada semua pihak yang telah memberi dukungan penuh selama penyusunan skripsi ini. Harapannya, hasil dari penelitian ini dapat bermanfaat bagi masyarakat.
Bogor, Desember 2015
DAFTAR ISI
DAFTAR TABEL viiii
DAFTAR LAMPIRAN viiii
PENDAHULUAN 1
Latar Belakang 1
Tujuan 2
TINJAUAN PUSTAKA 2
Tanaman Padi 2
Pemuliaan Tanaman Padi 3
Segregan Transgresif 4
Pendugaan Parameter Genetik 4
METODE PENELITIAN 5
Tempat dan Waktu Penelitian 5
Bahan dan Alat 5
Prosedur Percobaan 5
Analisis Data 6
HASIL DAN PEMBAHASAN 7
Kondisi Umum 7
Nilai Tengah antar Karakter 7
Pendugaan Aksi Gen Berdasarkan Persebaran Populasi F2 9
Heritabilitas 10
Analisis Korelasi antar Karakter 13
Seleksi 15
KESIMPULAN DAN SARAN 19
Kesimpulan 19
Saran 19
DAFTAR PUSTAKA 20
LAMPIRAN 24
RIWAYAT HIDUP 27
DAFTAR TABEL
1 Nilai tengah dan simpangan baku setiap karakter persilangan IR64 x Situ
Patenggang dan IR64 x Mekongga 8
2 Pendugaan aksi gen masing-masing karakter kuantitatif persilangan IR64 x Situ Patenggang melalui analisis skewness dan kurtosis 9 3 Pendugaan aksi gen masing-masing karakter kuantitatif persilangan IR64 x
Mekongga melalui analisis skewness dan kurtosis 10 4 Nilai duga komponen ragam, KKG, heritabilitas arti luas pada persilangan
IR64 x Situ Patenggang 12
5 Nilai duga komponen ragam, KKG, heritabilitas arti luas pada persilangan
IR64 x Mekongga 12
6 Koefisien korelasi linier antar karakter pada persilangan IR64 x Situ
Patenggang 14
7 Koefisien korelasi linier antar karakter pada persilangan IR64 x Mekongga 14 8 Karakteristik jumlah gabah bernas malai-1 dua populasi F2 persilangan padi 16
9 Individu F2 yang diduga segregan transgresif persilangan IR64 x Situ
Patenggang berdasarkan satu karakter 17
10Individu F2 yang diduga segregan transgresif persilangan IR64 x Mekongga
berdasarkan satu karakter 18
11 Diferensial seleksi dua populasi persilangan padi berdasarkan karakter
jumlah gabah bernas malai-1 19
DAFTAR LAMPIRAN
1 Data varietas tanaman padi 24
2 Data iklim bulanan bulan November 2014 – Maret 2015 26 3 Alur pendugaan aksi gen dengan analisis skewness dan kurtosis (Jambormias
PENDAHULUAN
Latar Belakang
Masalah ketahanan pangan saat ini menjadi isu global dan menjadi agenda utama di seluruh negara sebagai akibat adanya penyusutan lahan pertanian, perubahan iklim global, dan pertambahan penduduk. Menurut data BPS (2010), jumlah penduduk indonesia mencapai 237,6 juta jiwa dan akan terus meningkat hingga tahun 2025 yang diproyeksikan mencapai 300 juta jiwa sehingga penyediaan pangan, terutama beras, dalam jumlah yang cukup dan terjangkau harus menjadi prioritas utama pembangunan nasional. Hal ini disebabkan beras adalah makanan pokok lebih dari 95% penduduk indonesia dengan laju konsumsi sebesar 136 kg kapita-1 tahun-1 melebihi rata-rata tingkat konsumsi dunia 60 kg kapita-1
tahun-1. Produksi padi Indonesia pada tahun 2014 mengalami penurunan sebesar
0,45 juta ton atau 0,63% dari tahun 2013, yaitu dari 71,28 juta ton gabah kering giling (GKG) menjadi 70,83 juta ton GKG (BPS 2015).
Pemuliaan tanaman merupakan suatu metode sistematik yang dilakukan untuk merakit keragaman genetik menjadi suatu bentuk yang bermanfaat bagi kehidupan manusia. Varietas unggul hasil pemuliaan tanaman diharapkan mampu menjadi salah satu teknologi kunci dalam peningkatan produktivitas padi (Sadimantara et al. 2013). Keragaman genetik dapat diperluas dengan persilangan atau hibridisasi, yaitu menggabungkan karakter-karakter yang diinginkan dari para tetua sehingga diperoleh populasi baru sebagai bahan seleksi dalam program perakitan varietas unggul baru (Biswal et al. 2008). Seleksi dilakukan secara visual dengan mengamati fenotipe tanaman untuk memisahkan genotipe-genotipe yang unggul dari genotipe yang tidak diharapkan. Genotipe-genotipe yang dikehendaki dapat diperoleh dengan mempertimbangkan besaran beberapa parameter genetik. Parameter genetik yang diduga dalam penelitian ini adalah aksi gen, nilai heritabilitas, koefisien keragaman genetik, dan koefisien korelasi.
Tingkat segregasi tertinggi pada tanaman menyerbuk sendiri terjadi pada generasi F2 yang tergambar melalui sebaran frekuensi genotipenya (Welsh 1991).
Sebaran frekuensi tersebut dapat digunakan sebagai penduga pola pewarisan sifat dan jumlah gen yang terlibat dalam pengendalian suatu sifat sehingga bila tidak ada pengaruh lingkungan yang besar, suatu segregan transgresif telah ada pada generasi F2 atau pada generasi seleksi S0 (Christiana 1996).
2
Tujuan
Tujuan dari penelitian ini adalah untuk menduga parameter genetik dua persilangan padi dan mendapatkan karakter-karakter yang dapat dijadikan kriteria seleksi pada populasi F2 hasil dua persilangan padi.
TINJAUAN PUSTAKA
Tanaman Padi
Padi (Oryza sativa L.) merupakan tanaman rumput berumpun yang budidayanya sudah ada sejak sebelum Masehi. Berdasarkan bukti sejarah, padi pertama kali dibudidayakan sebagai makanan pokok orang Asia (Wasim 2002). Penanaman padi sudah dimulai sejak 3 000 tahun Sebelum Masehi di Zhejiang, Cina. Bukti lainnya yaitu ditemukannya fosil butir padi dan gabah di Hanstinapur Uttar Pradesh India sekitar 100−800 Sebelum Masehi (Purwono dan Purnamawati 2007). Bukti-bukti sejarah tersebut tidak dapat membuktikan secara pasti dari negara mana padi berasal, namun yang pasti bahwa padi berasal dari Asia Tenggara dan menyebar ke Asia Utara (Wasim 2002).
Negara Indonesia merupakan negara yang 95% penduduknya mengonsumsi beras. Hal ini disebabkan tanaman pangan yang menyebar hampir secara merata di seluruh wilayah Indonesia namun beberapa daerah tertentu saja yang merupakan sentra beberapa jenis tanaman pangan karena kesesuaian lahan dan kultur masyarakat dalam mengembangkan jenis tanaman pangan berbeda-beda setiap daerah (Purwono dan Purnamawati 2007).
Berdasarkan sistematika tanaman (taksonomi), tanaman padi diklasifikasikan ke dalam divisio Spermatophytae,subdivisio Angiospermae, kelas Monocotyledonae, dengan ordo Poales, famili Graminae, serta genus Oryza Linn dan dengan nama spesies Oryza sativa L (Grist 1960). Tanaman padi memiliki morfologi yang terdiri dari organ-organ vegetatif dan generatif. Organ-organ tanaman padi ini sangat berpengaruh terhadap produktivitasnya, misalnya suhu mikro tajuk tanaman, efektivitas menangkap radiasi surya, dan ketersediaan air bagi tanaman (Makarim dan Suhartatik 2009).
Organ vegetatif tanaman padi terdiri dari akar, batang, dan daun. Tanaman padi memiliki akar primer dan akar sekunder. Akar primer adalah akar yang tumbuh bersama akar-akar lain yang muncul dari embrio dekat bagian buku skutellum saat berkecambah. Akar ini disebut akar seminal, jumlahnya antara 1−7 (Chang dan Bardenas 1976). Sedangkan akar sekunder adalah akar akar yang tumbuh dari bagian buku terbawah batang menggantikan akar seminal. Akar ini disebut akar adventif, yaitu akar yang tumbuh dari bagian tanaman bukan embrio atau bukan dari akar yang tumbuh sebelumnya (Makarim dan Suhartatik 2009).
3
terjadi akibat dua faktor, yaitu: 1) meningkatnya jumlah anakan, 2) meningkatnya luas daun itu sendiri (Murata dan Matsushima 1978). Luas daun total pada tiap satuan luas lahan disebut Indeks Luas Daun (ILD). Padi memiliki ILD optimal berkisar antara 4-7 dan kira kira akan mencapai maksimal saat sebelum berbunga (Yoshida 1981).
Batang tanaman padi terdiri atas beberapa ruas yang dibatasi oleh buku yang berfungsi sebagai penopang tanaman dan penyalur senyawa-senyawa kimia dan air dalam tanaman. Permukaan stadia tumbuh batang terdiri atas pelepah-pelepah daun dan ruas-ruas yang tertumpuk padat dan setelah tanaman memasuki stadia reproduktif (perpanjangan ruas), ruas-ruas tersebut memanjang dan berongga (Yoshida 1981).
Bunga secara keseluruhan dalam satu tanaman padi disebut malai. Malai yang tersusun dari tiap unit bunga disebut spikelet. Spikelet terdiri atas tangkai, bakal buah, lemma, palea, putik, dan benang sari serta beberapa organ lainnya yang bersifat inferior. Spikelet terletak pada cabang-cabang bulir yang terdiri atas cabang primer dan sekunder (Siregar 1981). Tiap unit bunga pada padi hanya terdiri dari satu bunga atau disebut floret yang terdiri dari satu organ betina (pistil) dan 6 organ jantan (stamens) (Gould 1968).
Pemuliaan Tanaman Padi
Program pemuliaan tanaman memegang peranan penting dalam meningkatkan produktivitas tanaman. Kegiatan persilangan dan seleksi merupakan bagian dari proses pemuliaan tanaman yang bertujuan untuk menghasilkan suatu varietas baru (Jambormias 2014). Persilangan padi di Indonesia dimulai pada tahun 1920-an dengan memanfaatkan gene pool yang dibangun melalui introduksi tanaman (Harahap et al. 1972). Pemuliaan padi kemudian diarahkan pada lahan dengan pemupukan yang rendah atau tanaman kurang responsif terhadap pemupukan sampai tahun 1960-an dan pada tahun 1943 dilakukan pelepasan varietas pertama padi tipe Begawan yang kurang responsif terhadap pupuk.
Padi tipe PB5 dilepaskan pada tahun 1967 dengan umur genjah dan responsif terhadap pemupukan. Kemudian pada tahun 1977 dan 1986 dilepaskan varietas yang tahan terhadap hama dan penyakit dan cekaman biotik dan penambahan keunggulan rasa nasi yang enak yaitu padi tipe IRxx dan IR64 (Susanto et al. 2003). Pelepasan varietas-varietas tersebut ternyata belum mampu memenuhi kebutuhan beras penduduk yang terus meningkat, sehingga dikembangkan padi hibrida dan padi tipe baru. Teknologi padi hibrida potensial untuk memenuhi kebutuhan pangan di Asia Selatan dan Asia Tenggara pada tahun 2020 yang diprediksikan sebesar 800 juta ton (IRRI 2001). Padi tipe baru juga diharapkan dapat memacu peningkatan produksi padi di Indonesia (Susanto et al. 2003).
Pemuliaan tanaman padi bersifat dinamis. Varietas-varietas baru terbentuk sepanjang waktu diikuti dengan rata-rata peningkatan produktivitas secara nasional. Sampai pada tahun 2012, diperkenalkan padi tipe baru yaitu padi varietas IPB 3S dan IPB 4S dengan potensi hasil mencapai 11.23 ton GKG ha-1 dan 10.56 ton GKG
4
Segregan Transgresif
Pelaksanaan persilangan bertujuan untuk merakit kombinasi gen-gen dari sifat-sifat penting yang berada pada dua atau lebih tipe berbeda. Zuriat pertama (F1) dari suatu hasil persilangan umumnya homogen dan heterozigot. Heterozigotas pada tanaman F1 dapat ditemukan pada semua lokus. Hasil selfing tanaman F1 menghasilkan tanaman F2 yang merupakan populasi bersegregasi yang heterogen dengan campuran individu yang mengandung genotipe homozigot, kombinasi homozigot dan heterozigot, dan genotipe heterozigot. Hasil segregasi yang bersifat transgresif dapat ditemukan pada genotipe yang heterogen ini (Poehlman dan Sleper 1996).
Segregan transgresif adalah zuriat pada generasi awal yang memiliki keragaan fenotipe atau rata-rata penampilan fenotipik yang tinggi, di luar sebaran fenotipik kedua tetuanya. Individu-individu hasil segregasi transgresif yang memiliki keragaan di luar rentang keragaan tetuanya ditandai oleh nilai tengah yang tinggi dan ragam dalam populasi yang kecil (Jambormias dan Riry 2009). Segregan transgresif dapat diprediksi pada generasi F1, yaitu genotipe terbaik dari dua varietas dengan daya gabung umum tinggi, dan dapat diamati pada generasi awal persilangan, yaitu pada generasi F2, F3, dan F4, dengan akurasi terbaik pada generasi F3 (Chahota et al. 2007). Secara teoritis suatu segregan transgresif telah ada pada generasi segregasi F2 atau pada generasi seleksi S0 apabila tidak ada pengaruh lingkungan yang besar.
Pendugaan Parameter Genetik
Pendugaan nilai heritabilitas, koefisien keragaman genetik, aksi gen pengendali, dan koefisien korelasi penting sebagai tolak ukur kemajuan genetik yang dapat diharapkan dalam suatu proses seleksi. Nilai duga heritabilitas arti luas adalah perbandingan antara ragam genotipe total terhadap ragam fenotipe. Nilai duga heritabilitas menunjukkan apakah sesuatu karakter dikendalikan oleh faktor genetik atau faktor lingkungan, sehingga dapat diketahui sejauh mana karakter tersebut dapat diturunkan ke keturunan selanjutnya (Lestari et al. 2006). Herawati et al. (2009) menyebutkan bahwa seleksi akan lebih efektif jika karakter yang menjadi target seleksi memiliki nilai heritabilitas yang tinggi.
Koefisien keragaman genetik digunakan untuk menduga luas atau tidaknya keragaman genetik yang dimiliki masing-masing karakter. Bila tingkat keragaman genetik sempit maka keragaman antar individu dalam populasi relatif seragam, sehingga seleksi untuk perbaikan sifat menjadi kurang efektif (Puspitasari 2011). Sebaliknya, apabila keragaman genetik semakin luas, maka peluang keberhasilan seleksi dalam meningkatkan frekuensi gen yang diinginkan semakin besar pula (Allard 1960).
5
karakter yang diamati dan sebagai petunjuk bagi karakter yang lain yang lebih penting sehingga dapat melengkapi kriteria seleksi.
METODE PENELITIAN
Tempat dan Waktu Penelitian
Penelitiaan dilaksanakan pada bulan November 2014 hingga April 2015 di Kebun Percobaan Leuwikopo Institut Pertanian Bogor, Kecamatan Dramaga, Kabupaten Bogor, Jawa Barat. Pengeringan, perontokan dan penimbangan padi dilakukan di Laboratorium Pemuliaan Tanaman, Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor.
Bahan dan Alat
Bahan tanam yang digunakan dalam penelitian ini adalah tiga populasi tetua dan dua populasi zuriat hasil persilangan. Tetua yang digunakan yaitu IR64, Situ Patenggang, dan Mekongga. Pupuk yang digunakan adalah pupuk urea 250 kg ha -1, sp 36 200 kg ha-1, KCL 100 kg ha-1. Hama dan penyakit tanaman dikendalikan
menggunakan pestisida dan jaring. Alat yang digunakan adalah alat-alat pertanian secara umum, label, jaring, meteran, penggaris, counter, gunting, timbangan digital, kamera, amplop, dan alat tulis.
Prosedur Percobaan
Tanaman yang digunakan dalam penelitian ini adalah tiga populasi tetua (IR64, Situ Patenggang, dan Mekongga) dan persilangannya (IR64 x Situ Patenggang dan IR64 x Mekongga) yang terdiri dari populasi P1, P2, dan F2.
Masing-masing tetua (P1 dan P2) ditanam sebanyak 20 tanaman. Populasi F2
ditanam sebanyak 250 tanaman, sehingga tiap seri persilangan yang ditanam yaitu sebanyak 290 tanaman. Seluruh bahan tanam ini ditanam di lahan kering.
Persiapan lahan dilakukan satu minggu sebelum tanam yang meliputi pembersihan gulma, penggemburan menggunakan cangkul, dan pembuatan lubang tanam menggunakan tugal. Benih padi ditanam sebanyak satu benih tiap lubang secara langsung tanpa dilakukan penyemaian dengan jarak tanam 35 cm x 15 cm. Pemupukan dilakukan di awal tanam dan saat 2 MST dengan cara ditabur pada alur yang telah dibuat diantara barisan tanaman. Pemupukan pertama yaitu pupuk Urea 125 kg ha-1, SP-36 200 kg ha-1 dan KCl 100 kg ha-1. Pemupukan kedua yaitu Urea
dengan dosis 125 kg ha-1.
6
hingga 14 MST. Pengairan yaitu dengan sistem tadah hujan dan menggunakan sprinkle.
Pemanenan dilakukan secara bertahap, dimulai saat tanaman berumur 100
– 130 hari. Pemanenan menggunakan gunting yaitu dengan cara memotong batang padi bagian atas lalu masing-masing rumpun dipisahkan dalam satu amplop. Pengeringan dilakukan di dalam oven dengan suhu 35 oC selama ± 3 hari.
Pengamatan karakter kuantitatif dilakukan pada masing-masing tanaman yang meliputi:
1. Tinggi tanaman vegetatif, diukur dari pangkal batang sampai daun tertinggi pada saat tanaman berumur 45 HST.
2. Tinggi tanaman generatif, diukur dari pangkal batang sampai daun tertinggi pada saat tanaman berumur 90 HST.
3. Jumlah anakan vegetatif, total jumlah anakan setiap tanaman saat tanaman berumur 45 HST.
4. Jumlah anakan generatif, total jumlah anakan setiap tanaman saat tanaman berumur 90 HST.
5. Jumlah anakan produktif, jumlah anakan yang memiliki malai dari setiap tanaman.
6. Panjang malai, diukur dari leher malai sampai ujung malai pada saat panen. 7. Jumlah gabah bernas dan hampa malai-1, yaitu jumlah gabah bernas dan
hampa setiap malai dari masing-masing tanaman.
8. Bobot 100 butir, bobot 100 gabah bernas dari masing-masing tanaman. 9. Bobot gabah bernas tanaman-1, total bobot gabah bernas dari masing-masing
tanaman.
Analisis Data
Analisis data yang dilakukan dalam penelitian ini adalah pendugaan komponen ragam, pendugaan nilai heritabilitas arti luas, koefisien keragaman genetik, pendugaan aksi gen, dan koefisien korelasi. Pendugaan komponen ragam diperoleh berdasarkan Mahmud dan Kramer (1951):
Ragam fenotipe (�2
Koefisien keragaman genetik (KKG) = √��
2
�̅ x 100%; �̅ = nilai tengah populasi
Kriteria nilai KKG menurut Kight (1979) yaitu: sempit (0−10%), sedang
(10−20%), dan luas (> 20%)
Nilai duga heritabilitas arti luas (HBS) dihitung berdasarkan nilai duga
komponen ragam. Berdasarkan Allard (1960), HBS dihitung berdasarkan rumus
berikut:
HBS =
��2
��2 x 100%
7
50% ≤ H ˂ 100% = tinggi 20% ≤ H ˂ 50% = sedang
0 ≤ H ˂ 20% = rendah
Nilai koefisien korelasi dihitung berdasarkan Walpole (1992):
r = ∑ −
pengamatan; x dan y masing-masing berupa peubah bebas
Pendugaan aksi gen dilakukan dengan menggunakan analisis kemenjuluran kurva (skewness) dan keruncingan kurva (kurtosis). Nilai skewness (S), kurtosis (K), galat baku skewness (SES), dan galat baku kurtosis (SEK) dihitung
menggunakan perangkat lunak STAR IRRI. Statistik uji untuk kedua parameter tersebut mengikuti sebaran normal baku sebagai berikut:
ZS = �
���; ZK =
� ���
nilai kritikal untuk pengujian dua arah yaitu Z0.05/2 = 1.96 dan Z0.01/2 = 2.57.
HASIL DAN PEMBAHASAN
Kondisi Umum
Penilitian ini dilaksanakan di Kebun Percobaan Leuwikopo, Institut Pertanian Bogor, yang terletak ±201 m dpl. sejak bulan November 2014 hingga April 2015. Curah hujan rata-rata pada bulan November hingga Maret yaitu sebesar 371.74 mm, suhu rata-rata dan kelembaban rata-rata bulan-1 adalah 25.68 oC dan
73.8% (BMKG 2015). Gulma yang tumbuh pada pertanaman yaitu gulma dari golongan teki, rumput, dan daun lebar diantaranya adalah cyperus rotundus, Setaria plicata, Eleusine indica, dan Asystasia intrusa, sehingga diperlukan pengendalian secara manual yang intensif sebab pertumbuhan gulma cukup pesat. Hama yang menyerang berupa wereng coklat (Nilaparvata lugens), kepik hijau (Nezara viridula), walang sangit (Leptocorisa acuta), dan burung gereja (Passer montanus). Penyemprotaan pestisida dilakukan mulai 6 MST hingga 14 MST dilakukan tiap minggu sekali.
Penyakit yang menyerang tanaman padi yaitu hawar daun (Xanthomonas campestris), bercak coklat (Helminthosporium oryzae), dan gosong palsu (Ustilagonoidea virens). Selain hama, penyakit, dan gulma, faktor lain yang menyebabkan kerusakan pada tanaman padi adalah adanya hujan dan angin yang menyebabkan padi rebah. Kerebahan ini terutama terjadi pada padi persilangan IR64 x Mekongga.
Nilai Tengah antar Karakter
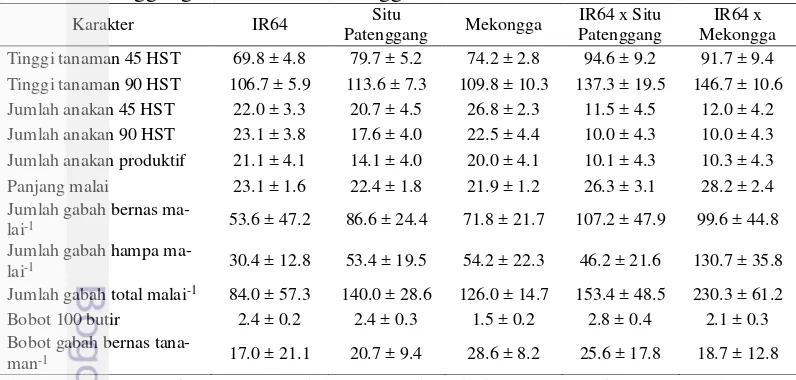
8
jumlah anakan 45 HST, jumlah anakan 90 HST, dan jumlah anakan produktif memiliki nilai tengah di bawah nilai tengah kedua tetuanya dan hanya karakter jumlah gabah hampa malai-1 yang berada di anatara nilai tengah kedua tetuanya.
Sementara hampir semua simpangan baku karakter lebih besar dari pada simpangan baku kedua tetuanya kecuali karakter bobot gabah bernas tanaman-1. Hasil
perhitungan nilai tengah dan simpangan baku tersebut menunjukkan besarnya ragam genetik pada populasi bersegregasi hasil persilangan IR64 x Situ Patenggang. Karakter tinggi tanaman 45 HST dan 90 HST, panjang malai, jumlah gabah bernas malai-1, jumlah gabah hampa malai-1, dan jumlah gabah total malai-1
pada persilangan IR64 x Mekongga memiliki nilai tengah yang lebih tinggi dari nilai tengah kedua tetuanya. Jumlah anakan 45 HST, jumlah anakan 90 HST, dan jumlah anakan produktif memiliki nilai tengah yang lebih kecil dari nilai tengah kedua tetuanya dan hanya bobot 100 butir yang memiliki nilai tengah di antara kedua tetuanya. Simpangan baku pada persilangan IR64 x Mekongga juga memiliki nilai yang lebih tinggi dari pada kedua tetuanya kecuali jumlah anakan 90 HST, jumlah anakan produktif, dan bobot gabah bernas tanaman-1.
Karakter-karakter pada kedua populasi zuriat hasil persilangan yang memiliki nilai tengah yang lebih tinggi dari pada nilai tengah kedua tetuanya adalah karakter tinggi tanaman 45 HST, tinggi tanaman 90 HST, panjang malai, jumlah gabah bernas malai-1, dan jumlah gabah total malai-1. Simpangan baku pada kedua
persilangan menunjukkan besarnya kisaran nilai tengah. Semakin tinggi simpangan baku maka kisaran nilai tengah juga semakin besar. Tingginya simpangan baku atau kisaran nilai tengah ini menandakan adanya keragaman genetik yang besar pada kedua populasi zuriat hasil persilangan. Hampir seluruh karakter kuantitatif pada kedua persilangan memiliki simpangan baku yang lebih tinggi dari kedua tetuanya. Berdasarkan deskripsi nilai tengah dan simpangan baku ini secara umum menggambarkan adanya peluang untuk memperbaiki keragaan karakter dari kedua populasi melalui seleksi (Jambormias 2014).
Tabel 1 Nilai tengah dan simpangan baku setiap karakter persilangan IR64 x Situ Patenggang dan IR64 x Mekongga
9
Pendugaan Aksi Gen Berdasarkan Persebaran Populasi F2
Populasi generasi awal (F2) persilangan IR64 x Situ Patenggang dan IR64 x Mekongga merupakan populasi bersegregasi yang terdiri individu-individu yang beragam dan ragam menyebar tak normal untuk hampir semua karakter kuantitatif (Tabel 3 dan Tabel 4). Karakter-karakter kuantitatif pada persilangan IR64 x Situ Patenggang, seperti jumlah anakan 45 HST, jumlah anakan 90 HST, jumlah anakan produktif, panjang malai, jumlah gabah hampa malai-1, bobot 100 butir, dan bobot
gabah bernas tanaman-1 menyebar platikurtik atau banyak gen aditif yang terlibat
dalam mengendalikan suatu sifat. Sementara untuk karakter tinggi tanaman 45 HST, tinggi tanaman 90 HST, jumlah gabah bernas malai-1, dan jumlah gabah total
malai-1 menyebar leptokurtik atau sedikit segregasi gen aditif yang terlibat.
Analisis skewness dan kurtosis berperan penting dalam menentukan terjadi atau tidaknya epistasis pada individu F2 hasil persilangan (Jambormias 2014). Analisis skewness dan kurtosis juga akan memberikan informasi tentang sifat dasar aksi gen (Fisher et al. 1932) dan menentukan suatu karakter dikendalikan oleh gen mayor atau gen minor (Robson 1956). Karakter tinggi tanaman 45 HST dan jumlah gabah total malai-1 terjadi sebaran mesokurtik dengan skewness positif
mengindikasikan bahwa aksi gen yang terjadi adalah aksi gen aditif tanpa pengaruh gen dominan maupun epistasis. Karakter tinggi tanaman 90 HST memiliki sebaran mesokurtik dengan skewness negatif dan bobot gabah bernas tanaman-1 juga
memiliki sebaran mesokurtik dengan skewness positif mengindikasikan bahwa aksi gen yang terjadi adalah epistasis duplikat tanpa pengaruh aditif. Hampir keseluruhan karakter memiliki sebaran leptokurtik atau kurva skewness yang menjulur ke kanan dengan aksi gen epistasis koplementer.
Tabel 2 Pendugaan aksi gen masing-masing karakter kuantitatif persilangan IR64 x Situ Patenggang melalui analisis skewness dan kurtosis
Karakter S Zs K Zk Jlh. Gen dan
Keterangan: S: skewness; Zs: statistik uji skewness; K: kurtosis; Zk: statistik uji kurtosis; *: statistik
uji nyata pada taraf nyata 0.05; **: statistik uji sangat nyata pada taraf nyata 0.01; tn:
10
Tabel 3 Pendugaan aksi gen masing-masing karakter kuantitatif persilangan IR64 x Mekongga melalui analisis skewness dan kurtosis
Karakter S Zs K Zk Jlh. Gen dan
Keterangan: S: skewness; Zs: statistik uji skewness; K: kurtosis; Zk: statistik uji kurtosis; *: statistik
uji nyata pada taraf nyata 0.05; **: statistik uji sangat nyata pada taraf nyata 0.01; tn:
statistik uji tidak nyata; S: dikendalikan sedikit gen; B: dikendalikan banyak gen; Ad: hanya aditif; Dm: dominansi; EK: epistasis komplementer; EA: epistasis aditif; ED: epistasis duplikat
Persilangan IR64 x Mekongga memiliki karakter-karakter kuantitatif yang memiliki sebaran leptokurtik, kecuali karakter jumlah anakan 45 HST dan jumlah gabah bernas malai-1 yang memiliki sebaran platikurtik. Karakter tinggi tanaman
45 HST, tinggi tanaman 90 HST, panjang malai dan bobot 100 butir memiliki nilai skewness negatif yang mengindikasikan bahwa aksi gen yang terlibat adalah epistasis duplikat. Karakter jumlah anakan 90 HST, jumlah anakan produktif, dan bobot gabah bernas tanaman-1 memiliki nilai skewness positif, hal ini diduga bahwa
gen yang mengendalikan adalah epistasis komplementer. Sementara pada karakter jumlah anakan 45 HST, jumlah gabah bernas malai-1, jumlah gabah hampa malai-1,
dan jumlah gabah total malai-1 mengikuti sebaran mesokurtik yang
mengindika-sikan bahwa aksi gen yang terlibat adalah aksi gen aditif tanpa pengaruh dominan maupun epistasis.
Karakter-karakter tanaman yang berpotensi untuk dijadikan kriteria seleksi adalah karakter yang sepenuhnya dikendalikan oleh gen aditif. Hayward (1990) mengatakan bahwa pengaruh aditif dapat diwariskan, sementara pengaruh bukan gen aditif tidak diwariskan. Pengaruh tindak gen bukan aditif akan lenyap semasa seleksi sehingga sifat-sifat yang dikendalikan oleh gen-gen bukan aditif menyebabkan kemajuan genetik yang rendah.
Heritabilitas
11
Nilai duga heritabilitas arti luas adalah perbandingan antara ragam genotipe total terhadap ragam fenotipe (Borojevic 1990). Nilai heritabilitas dikategorikan tinggi
apabila 50% ≤ H ˂ 100%, sedang apabila 20% ≤ H ˂ 50%, dan rendah apabila 0% ≤ H ˂ 20% (Stanfield 1983). Roy (2000) menyatakan jika nilai duga heritabilitas tinggi maka seleksi dilakukan pada generasi awal karena kemajuan seleksinya akan besar. Sebaliknya, jika heritabilitasnya rendah hingga sedang maka karakter tersebut perlu difiksasi melalui seleksi.
Persilangan IR64 x Situ Patenggang menunjukkan bahwa tinggi tanaman 45 HST, tinggi tanaman 90 HST, panjang malai, jumlah gabah bernas malai-1, dan
bobot 100 butir memiliki nilai heritabilitas yang tinggi dengan nilai heritabilitas masing-masing sebesar 70%, 89%, 70%, 50%, dan 60% . Karakter jumlah anakan 45 HST, jumlah gabah hampa malai-1, jumlah gabah total malai-1, dan bobot gabah
bernas tanaman-1 memiliki nilai heritabilitas yang sedang dengan nilai heritabilitas
masing-masing sebesar 27%, 46%, 30%, dan 37%. Sementara untuk karakter yang memiliki nilai heritabilitas yang rendah adalah jumlah anakan 90 HST dan jumlah anakan produktif dengan nilai heritabilitas sebesar 18% dan 13%.
Selain nilai duga heritabilitas dalam arti luas, koefisien keragaman genetik (KKG) juga menjadi salah satu parameter penentu apakah suatu karakter dapat dijadikan kriteria seleksi atau tidak (Yunianti 2010). Menurut Puspitasari (2011) koefisien keragaman genetik digunakan untuk menduga luas atau tidaknya keragaman genetik yang dimiliki masing-masing karakter. Bila tingkat keragaman genetik sempit maka keragaman antar individu dalam populasi relatif seragam, sehingga seleksi untuk perbaikan sifat menjadi kurang efektif. Sebaliknya, apabila keragaman genetik semakin luas, maka peluang keberhasilan seleksi dalam meningkatkan frekuensi gen yang diinginkan semakin besar pula (Allard 1960). Nilai KKG sempit (0−10%), sedang (10−20%) dan luas (> 20%) (Knight 1979).
Populasi dasar dengan keragaman genetik yang tinggi merupakan bahan pemuliaan yang penting untuk perakitan varietas unggul. Seleksi pada populasi dasar yang memiliki keragaman genetik yang tinggi akan memberikan respon yang baik karena variasi genetik yang tinggi akan memberikan peluang besar untuk mendapatkan kombinasi persilangan yang tepat dengan gabungan sifat-sifat yang baik (Suprapto dan Kairudin 2007).
Berdasarkan perhitungan nilai KKG (Tabel 5), jumlah anakan 45 HST, jumlah gabah bernas malai-1, jumlah gabah hampa malai-1, dan bobot gabah bernas
tanaman-1 memiliki nilai KKG yang luas. Tinggi tanaman 90 HST, jumlah anakan
90 HST, jumlah anakan produktif, panjang malai, jumlah gabah total malai-1, dan
12
Tabel 4 Nilai duga komponen ragam, KKG, heritabilitas arti luas pada persilangan IR64 x Situ Patenggang
keragaman genetik; HBS: heritabilitas arti luas
Tabel 5 Nilai duga komponen ragam, KKG, heritabilitas arti luas pada persilangan IR64 x Mekongga
keragaman genetik; HBS: heritabilitas arti luas
Karakter-karakter pada persilangan IR64 x Mekongga yang memiliki nilai heritabilitas yang tinggi adalah tinggi tanaman 45 HST, jumlah anakan 45 HST, panjang malai, jumlah gabah hampa malai-1, jumlah gabah total malai-1, dan bobot
100 butir dengan nilai heritabilitas masing-masing sebesar 85%, 56%, 67%, 78%, 81%, dan 58%. Karakter yang memiliki nilai heritabilitas sedang adalah tinggi tanaman 90 HST dan jumlah gabah bernas malai-1 dengan nilai heritabilitas
masing-masing sebesar 46% dan 49%. Sementara untuk karakter jumlah anakan 90 HST, jumlah anakan produktif, dan bobot gabah bernas tanaman-1 memiliki nilai
heritabilitas yang tergolong rendah dengan nilai heritabilitas masing-masing sebesar 8%, 10%, dan 5%. Karakter jumlah anakan 45 HST, jumlah gabah bernas malai-1, jumlah gabah hampa malai-1, dan jumlah gabah total malai-1 memiliki nilai
KKG yang luas. Jumlah anakan 90 HST, jumlah anakan produktif, bobot 100 butir, dan bobot gabah bernas tanaman-1 memiliki nilai KKG sedang dan terdapat tiga
13
Hasil pendugaan nilai heritabilitas pada kedua populasi persilangan tersebut terlihat bahwa terdapat perbedaan nilai duga heritabilitas arti luas pada karakter yang sama pada kedua populasi persilangan padi. Natawijaya (2012) menyebutkan hal ini disebabkan oleh adanya perbedaan nilai genotipe tetua-tetuanya. Nilai heritabilitas suatu karakter nilainya tidak tetap karena banyak faktor yang dapat mempengaruhi nilai heritabilitas, yaitu populasi yang digunakan, metode estimasi, adanya pautan gen, pelaksanaan percobaan, generasi populasi yang diuji, dan kondisi lingkungan (Puspitasari 2011).
Berdasarkan nilai heritabilitas arti luas dan KKG, kedua populasi ini memiliki karakter-karakter yang dapat dijadikan kriteria seleksi. Karakter yang memiliki KKG yang luas dan heritabilitas sedang hingga tinggi pada persilangan IR64 x Situ Patenggang adalah jumlah anakan 45 HST, jumlah gabah bernas malai -1, jumlah gabah hampa malai-1, dan bobot gabah bernas tanaman-1.
Karakter-karakter yang memiliki nilai KKG yang luas dan heritabilitas yang sedang hingga tinggi pada persilangan IR64 x Mekongga adalah jumlah anakan 45 HST, jumlah gabah bernas malai-1, jumlah gabah hampa malai-1, dan jumlah gabah total malai-1.
Karakter yang memiliki nilai heritabilitas yang tinggi menunjukkan bahwa pengaruh faktor genetik lebih besar terhadap penampilan fenotipik dibandingkan dengan pengaruh lingkungan. Sebaliknya, bahwa karakter yang memiliki nilai heritabilitas rendah disebabkan karena pengaruh lingkungan yang lebih dominan terhadap fenotipiknya dari pada pengaruh genetiknya, sehingga seleksi akan lebih efektif jika dilakukan terhadap karakter yang memiliki duga heritabilitas tinggi daripada karekter yang memiliki heritabilitas yang rendah. Berkaitan dengan nilai KKG, maka seleksi akan efisien dan efektif jika dilakukan pada karakter yang memiliki nilai KKG yang luas dan nilai heritabilitas yang tinggi (Sutjahjo et al. 2007).
Analisis Korelasi antar Karakter
Daya hasil merupakan karakter kuantitatif yang sangat dipengaruhi oleh karakter komponen hasil maupun karakter agronomi lain yang terkait dengan daya hasil. Keeratan hubungan antara karakter daya hasil dengan karakter lain yang mempengaruhi daya hasil dapat diduga dengan menghitung nilai koefisien korelasi antara kedua karakter. Aminasih (2009) juga menyebutkan bahwa koefisien korelasi merupakan suatu ukuran keeratan hubungan antara dua karakter yang diamati dan sebagai petunjuk bagi karakter yang lain yang lebih penting sehingga dapat melengkapi kriteria seleksi.
Koefisien korelasi antar karakter masing-masing persilangan disajikan pada Tabel 7 dan Tabel 8. Hasil analisis korelasi pada persilangan IR64 x Situ Patenggang menunjukkan bahwa tinggi tanaman 45 HST, tinggi tanaman 90 HST, jumlah anakan 45 HST, jumlah anakan 90 HST, jumlah anakan produktif, panjang malai, bobot gabah bernas malai-1, bobot gabah total malai-1, dan bobot 100 butir
berkorelasi positif dan sangat nyata terhadap bobot bernas total tanaman-1. Hal ini
menunjukkan bahwa apabila karakter komponen hasil mengalami peningkatan makan bobot bernas total tanaman-1 juga meningkat (Tabel 7).
14
Berdasarkan nilai koefisien korelasi bahwa jumlah gabah hampa malai-1 berkorelasi
nyata namun berkebalikan yang ditandai dengan nilai negatif. Hal ini menunjukkan bahwa penurunan jumlah gabah hampa malai-1 akan meningkatkan bobot bernas
total tanaman-1. Karakter tinggi tanaman 45 HST, tinggi tanaman 90 HST, jumlah
anakan 90 HST, jumlah anakan produktif, panjang malai, jumlah gabah total tanaman-1, bobot 100 butir, dan bobot gabah bernas total tanaman-1 juga berkorelasi
positif dan sangat nyata terhadap jumlah gabah bernas total malai-1, kecuali jumlah
gabah hampa malai-1 yang juga berkorelasi negatif dan nyata terhadap jumlah gabah
bernas malai-1.
Tabel 6 Koefisien korelasi linier antar karakter pada persilangan IR64 x Situ Patenggang
Keterangan: TT45: tinggi tanaman 45 HST; TT90: tinggi tanaman 90 HST; JA45: jumlah anakan 45 HST; JA90: jumlah anakan 90 HST; JAP: jumlah anakan produktif; PM: panjang malai; GB: jumlah gabah bernas malai-1; GH: jumlah gabah hampa malai-1; GT: jumlah
gabah total malai-1; 100BTR: bobot 100 butir; BBT: bobot gabah bernas total tanaman -1; *: berkorelasi nyata pada taraf 5%; **: berkorelasi nyata pada taraf 1%; tn: tidak
berkorelasi nyata
Tabel 7 Koefisien korelasi linier antar karakter pada persilangan IR64 x Mekongga
TT45 TT90 JA45 JA90 JAP PM GB GH GT 100BTR
Keterangan: TT45: tinggi tanaman 45 HST; TT90: tinggi tanaman 90 HST; JA45: jumlah anakan 45 HST; JA90: jumlah anakan 90 HST; JAP: jumlah anakan produktif; PM: panjang malai; GB: jumlah gabah bernas malai-1; GH: jumlah gabah hampa malai-1; GT: jumlah
gabah total malai-1; 100BTR: bobot 100 butir; BBT: bobot gabah bernas total tanaman -1; *: berkorelasi nyata pada taraf 5%; **: berkorelasi nyata pada taraf 1%; tn: tidak
berkorelasi nyata
15
terjadi berlawanan (Falconer 1989). Apabila terdapat dua sifat yang diamati menunjukkan korelasi yang positif, maka dapat dijelaskan bahwa seiring bertambah besar atau bertambah banyaknya suatu sifat akan selalu diikuti oleh bertambah besar atau bertambah banyaknya sifat yang lain. Karakter-karakter komponen hasil yang berkorelasi positif dan sangat nyata pada kedua persilangan dapat dijadikan sebagai kriteria seleksi.
Hampir semua karakter komponen hasil yang diamati pada persilangan IR64 x Mekongga berkorelasi positif dan sangat nyata terhadap bobot bernas total tanaman-1, kecuali jumlah gabah hampa malai-1 yang berkorelasi tidak nyata
terhadap bobot bernas total tanaman-1. Jumlah gabah bernas malai-1 juga berkorelasi
positif dan sangat nyata terhadap semua karakter yang diamati, artinya setiap peningkatan komponen-komponen hasil tersebut akan meningkatkan bobot bernas malai-1. Hal ini sejalan dengan penilitian yang dilaksanakan oleh Rachmawati
(2014) bahwa beberapa karakter yang diamati memiliki korelasi positif terhadap daya hasil, seperti jumlah malai dan jumlah anakan. Jumlah daun berhubungan langsung dengan jumlah anakan, jadi semakin banyak jumlah anakan maka semakin bertambah pula jumlah daunnya. Daun berfungsi penting terhadap penerimaan dan penyerapan cahaya untuk proses fotosintesis. Semakin tinggi kegiatan fotosintesis maka semakin tinggi fotosintat yang akan menambah bobot atau mutu hasil (Rachmawati 2014). Akhtar et al. (2011) juga menyebutkan dalam penelitiannya bahwa jumlah gabah malai-1 dan bobot 100 butir berkorelasi positif dan nyata
terhadap hasil.
Seleksi
Kegiatan seleksi memegang peranan penting dalam program pemuliaan tanaman, khususnya dalam perbaikan genetik tanaman untuk menghasilkan varietas baru. Seleksi berlangsung efektif saat sejumlah individu-individu segregan transgresif berhasil difiksasi pada generasi awal. Keberhasilan fiksasi segregasi transgresif memungkinkan pengembangan metode seleksi generasi awal yang dapat mempercepat perolehan galur-galur harapan baru.
Strategi pemuliaan untuk mendapatkan varietas unggul pada tanaman menyerbuk sendiri adalah efisiensi seleksi. Seleksi akan akan optimal bila menggunakan karakter seleksi yang tepat. Seleksi berdasarkan daya hasil biasanya kurang memberikan hasil optimal bila tidak didukung oleh karakter seleksi lain berupa komponen pertumbuhan dan komponen hasil yang berkorelasi positif terhadap daya hasil (Limbongan 2008). Karakter hasil yang baik untuk dijadikan kriteria seleksi adalah karakter yang memiliki nilai KKG yang luas dan memiliki nilai heritabilitas arti luas yang tinggi (Kasno 1983). Perakitan varietas berdaya hasil tinggi dapat dilakukan melalui seleksi secara langsung terhadap daya hasil atau tidak langsung melalui beberapa karakter lain yang terkait dengan daya hasil (Falconer dan Mackay 1996).
16
tersebut memiliki nilai KKG yang luas dan nilai heritabilitas yang tinggi pada kedua persilangan. Karakter terpilih itu juga berkorelasi sangat nyata dan positif terhadap bobot 100 butir dan bobot bernas tanaman-1 berdasarkan analisis koefisien korelasi.
Karakter jumlah gabah bernas malai-1 yang pada persilangan IR64 x Mekongga
dikendalikan oleh banyak gen dengan aksi gen aditif, artinya bahwa karakter ini akan diwariskan kepada generasi berikutnya.
Nilai tengah tetua pada kedua persilangan dapat dilihat pada Tabel 9. Nilai tengah IR64, Situ Patenggang, dan Mekongga yaitu masing-masing sebesar 53.6, 86.6, dan 71.8. Seleksi segregan transgresif berdasarkan karakter jumlah gabah bernas malai-1 dilakukan dengan memilih tanaman yang memiliki nilai tengah yang lebih tinggi dari pada nilai tengah kedua tetuanya. Individu-individu F3 terpilih yang terduga segregan transgresif tersebut perlu diverifikasi kebenarannya melalui tanam baris untuk menentukan nilai tengah dan ragamnya.
Segregan transgresif dapat diprediksi dan diamati pada zuriat suatu generasi persilangan awal. Periode seleksi yang panjang dapat diperpendek dengan mendeteksi segregan transgresif di generasi awal. Oleh karena itu, mendeteksi segregan transgresif di generasi awal juga dapat meningkatkan efisiensi seleksi dalam kegiatan pemuliaan tanaman.
Keragaan generasi F2 yang lebih baik dari kedua tetuanya serta keragaman dan heritabilitas yang tinggi menunjukkan keunggulan komponen hasil generasi F2 terhadap kedua tetua yang mengindikasikan daya hasil yang tinggi pada generasi F2. Keragaman dan heritabilitas yang tinggi juga mengindikasikan masih adanya peluang untuk meningkatkan daya hasil dalam program seleksi.
Berdasarkan hasil seleksi satu karakter pada persilangan IR64 x Situ Patenggang terdapat 128 tanaman yang diduga sebagai segregan transgresif. Segregasi transgresif adalah segregasi gen pada sifat-sifat kuantitatif dari individu hasil persilangan yang memiliki jangkauan sebaran yang melampaui jangkauan sebaran kedua tetuanya (Poehlman dan Sleper 1996). Sementara pada persilangan IR64 x Mekongga terdapat 132 individu yang memiliki sebaran di atas sebaran tetuanya.
Tujuan pelaksanaan seleksi setelah persilangan untuk pemuliaan galur adalah untuk meningkatkan frekuensi genotipe segregan transgresif yang dikehendaki dari dalam populasi homozigositas dan heterozigositas pada setiap generasi sehingga diperoleh genotipe segregan transgresif homozigot untuk semua gen yang telah mengalami fiksasi. Segregasi transgresif membentuk dua gugus segregan transgresif dalam spektrum sebaran, yaitu lebih kecil dari sebaran tetua dengan keragaan rendah dan lebih besar dari sebaran tetua dengan keragaan tinggi (Jambormias dan Riry 2009).
Tabel 8 Karakteristik jumlah gabah bernas malai-1 dua populasi F2 persilangan
padi
Jumlah gabah bernas malai-1
IR64 x Situ Patenggang IR64 x Mekongga
...(butir malai-1) ...
Nilai tengah P1 53.56 53.56
Nilai tengah P2 86.56 71.80
Nilai tengah F2 107.24 99.62
HBS (%) 50 49
17
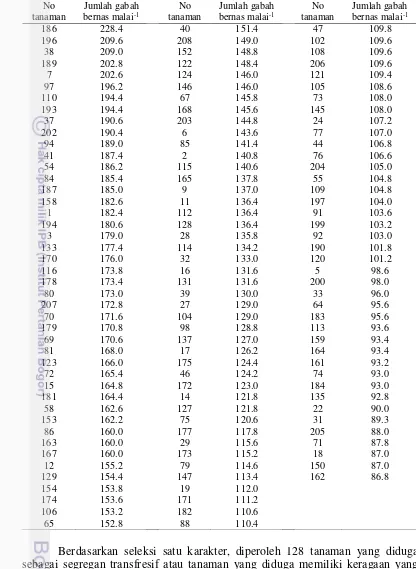
Tabel 9 Individu F2 yang diduga segregan transgresif persilangan IR64 x Situ Patenggang berdasarkan satu karakter
Berdasarkan seleksi satu karakter, diperoleh 128 tanaman yang diduga sebagai segregan transfresif atau tanaman yang diduga memiliki keragaan yang lebih baik dari pada kedua tetuanya pada persilangan IR64 x Situ Patenggang. Tanaman nomor 186 memiliki jumlah gabah bernas malai-1 yang lebih banyak,
sementara tanaman nomor 162 memiliki jumlah gabah bernas malai-1 paling sedikit
18
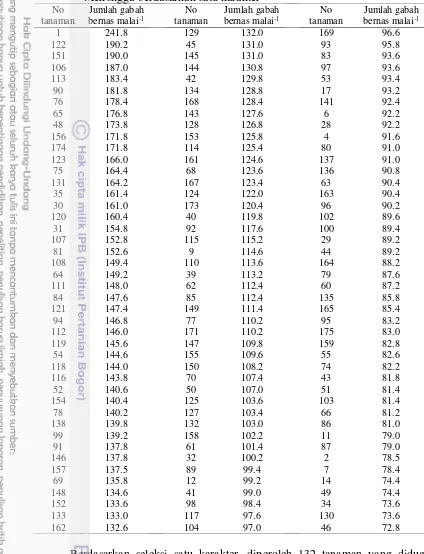
Tabel 10 Individu F2 yang diduga segregan transgresif persilangan IR64 x Mekongga berdasarkan satu karakter
Berdasarkan seleksi satu karakter, diperoleh 132 tanaman yang diduga sebagai segregan transfresif atau tanaman yang diduga memiliki keragaan yang lebih baik dari pada kedua tetuanya pada persilangan IR64 x Mekongga. Tanaman nomor 1 memiliki jumlah gabah bernas malai-1 yang lebih banyak, sementara
tanaman nomor 46 memiliki jumlah gabah bernas malai-1 paling sedikit diantara
19
DiferensialSeleksi
Efektifitas seleksi juga dapat ditentukan dari nilai diferensial seleksi. Diferensial seleksi merupakan selisih rerata individu-individu terseleksi dari rerata populasi. Diferensial seleksi menunjukkan keunggulan individu-individu terpilih dibandingkan populasi awalnya, semakin besar nilai diferensial seleksi maka kemajuan genetik pada populasi terseleksi akan semakin besar.
Tabel 11 Diferensial seleksi dua populasi persilangan padi berdasarkan karakter jumlah gabah bernas malai-1
Jumlah gabah bernas malai-1
Rata-rata F2 awal Rata-rata F2 terseleksi
Diferensial sele-ksi ...(butir malai-1) ...
IR64 x Situ Patenggang 107.24 137.07 29.83
IR64 x Mekongga 99.62 118.81 19.19
Tabel 12 menunjukkan nilai diferensial seleksi pada persilangan IR64 x Situ Patenggang dan IR64 x Mekongga berdasarkan karakter jumlah gabah bernas malai-1. Seleksi pada persilangan IR64 x Situ Patenggang menunjukkan
peningkatan jumlah gabah bernas malai-1 yang ditandai dengan nilai diferensial
seleksi sebesar 29.83 gram. Seleksi pada persilangan IR64 x Mekongga juga menunjukkan peningkatan jumlah gabah bernas malai-1 pada populasi terseleksi
yang ditandai dengan nilai diferensial seleksi sebesar 19.19 gram. Berdasarkan nilai diferensial tersebut terdapat peluang terjadinya peningkatan jumlah gabah bernas malai-1 pada generasi berikutnya (tanaman F3).
KESIMPULAN DAN SARAN
Kesimpulan
Berdasarkan analisis skewness dan kurtosis, hampir semua karakter kedua persilangan dikendalikan oleh sedikit gen aditif dengan aksi gen yang berbeda -beda. Jumlah gabah bernas malai-1 memiliki nilai tengah yang lebih baik dari pada
kedua tetuanya dengan nilai heritabilitas dan koefisien keragaman genetik yang luas, sehingga karakter ini dapat dijadikan kriteria seleksi. Terdapat 128 tanaman yang diduga sebagai segregan transgresif berdasarkan seleksi satu karakter pada persilangan IR64 x Situ Patenggang, sementara hasil seleksi pada persilangan IR64 x Mekongga terdapat 132 tanaman yang diduga segregan transgresif.
Saran
20
hara dan tingkat kehomogenan lahan. Hal ini dilakukan untuk mengurangi pengaruh lingkungan agar tidak terlalu besar. Pengendalian gulma dan hama pada masa pemeliharaan sebaiknya dilakukan secara lebih intensif karena dapat mempengaruhi hasil yang diperoleh.
DAFTAR PUSTAKA
Akhtar N, Nazir MF, Rabnawaz A, Mahmood T, Safdar ME, Asif M, Rehman A. 2011. Estimation of heritability, correlation, and path coeficient analysis in fine grain rice (Oryza sativa L.). The Journal of Animal and Plant Science. 21(4):660-664.
Allard RW. 1960. Principles of Plant Breeding. New York (US): J Wiley.
Aminasih N. 2009. Penentuan kriteria seleksi 45 galur terigu (Triticum aestivum L.) introduksi di Dempo Selatan, Pagar Alam, Sumatera Selatan. Jurnal Penelitian Sains. 12(1):1-6.
[BMKG] Badan Meteorologi Klimatologi dan Geofisika. 2015. Data iklim stasiun Dramaga. Bogor (ID): BMKG.
[BPS] Badan Pusat Statistik. 2010. Jumlah dan distribusi penduduk [Internet]. [diunduh 2015 Jul 09]. Tersedia pada: http://sp2010.bps.go.id/.
__________________________. 2015. Produksi padi, jagung, dan kedelai [Internet]. [diunduh 2015 Jul 09]. Tersedia pada: http://www.bps.go.id/brs/view/id/1122.
Biswal MK, Mondal MAA, Hossain M, Islam R. 2008. Persilangan untuk merakit varietas unggul baru kentang [Internet]. [diunduh 2015 Jul 10]. Tersedia pada:
http://balitsa.litbang.pertanian.go.id/ind/images/Iptek%20Sayuran/04.pdf. Borojevic S. 1990. Daya warisdan harapan kemajuan seleksi karakter agronomi kedelai generasi F2 hasil persilangan antara Yellow Bean dan Taichung. J Agrotek Tropika. 1(1):20-24.
Chahota RK, Kishore N, Dhiman KC, Sharma TR, Sharma SK. 2007. Predicting transgressive segregants in early generation using single seed descent method-derived micro-macrosperma genepool of lentil (Lens culinaris Medikus). Euphytica. 156(3): 305-310.
Chang Te-Tzu, Bardenas EA. 1976. Morfologi dan fisiologi tanaman padi. [Internet]. [diunduh 2015 Jul 09]. Tersedia pada: http://www.litbang.deptan.go.id/special/padi/bbpadi_2009_itkp_11.pdf. Christiana AL. 1996. Pola segregasi karakter agronomi tanaman kedelai (Glycine
max L. Merrill) generasi F2 hasil persilangan wilis x B3570. J Agrotek Tropika. 1(1):8-13.
Diennazola R. 2012. Liputan khusus: yang baru untuk tingkatkan produksi. [Internet]. [diunduh 2015 Jul 09]. Tersedia pada: http://agrina-online.com/redesign2.php?rid=19&aid=3940.
21
Fisher RA, Immer FR, Tedin O. 1932. The genetical interpretation of statistics of the third degree in the study of quantitative inheritance. Genetics. 17(2):107-124.
Gould FW. 1968. Morfologi dan fisiologi tanaman padi. [Internet]. [diunduh 2015 Jul 09]. Tersedia pada:http://www.litbang.deptan.go.id/special/padi/ bbpadi_2009_itkp_11.pdf.
Grist DH. 1960. Rice Formerly Agricultural Economist, Colonial Agricultural Service, Malaya. London (GB): Longmans Green and Co Ltd.
Harahap Z, Siregar H, Siwi BH. 1972. Rice Breeding. Filipina (PH): International Rice Research Institute.
Hayward G. 1990. Applied genetics. Hampshire (GB): Macmillan Education Ltd. Herawati R, Purwoko BS, Dewi IS. 2009. Keragaman genetik dan karakter
agronomi galur haploid ganda padi gogo dengan sifat-sifat tipe baru hasil kultur antera. J Agron Indonesia. 37(2):87-94.
[IRRI] International Rice Research Institute. 2001. Sekilas Kerja Sama Indonesia-IRRI, Dampak dan Tantangan ke Depan. Filipina (PH): IRRI.
Jambormias E. 2014. Analisis genetik dan segregasi transgresif berbasis kekerabatan untuk potensi hasil dan panen serempak kacang hijau [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Jambormias E, Riry J. 2009. Penyesuaian data dan penggunaan informasi kekerabatan untuk mendeteksi segregan transgresif sifat kuantitatif pada tanaman menyerbuk sendiri (suatu pendekatan dalam seleksi). J Budidaya Pertanian. 5(1):11-18.
Kasno A. 1983. Pendugaan parameter genetik sifat-sifat kuantitatif kacang tanah (Arachis hypogeae L. Merr) pada beberapa lingkungan tumbuh dan penggunaannya dalam seleksi [tesis]. Bogor (ID): Institut Pertanian Bogor. Knight R. 1979. Quantitive Genetic Statistics and Plant Breeding. Brisbane (AU):
Vice-Chancellors Committee.
Lestari AD, Dewi W, Qosim WA, Rahardja M, Rostini N, Setiamihardja R. 2006. Keragaman dan heritabilitas 10 genotip pada cabai besar (Capsicum annuum L.). Jurnal Produksi Tanaman. 2(4):301-307.
Limbongan YL, Aswidinnoor H, Purwoko BS, Trikoesoemaningtyas. 2008. Pewarisan sifat toleransi padi sawah (Oryza sativa L.) terhadap cekaman suhu rendah. Bul Agron. 36(2):111-117.
Mahmud I, Kramer HH. 1951. Segregation for yield, height, and maturity folowing a soybean cross. J Agron. 43(12):605-609.
Makarim AK, Suhartatik E. 2009. Morfologi dan fisiologi tanaman padi. [Internet]. [diunduh 2015 Jul 09]. Tersedia pada: http://www.litbang.deptan.go.id/ special/padi/bbpadi_2009_itkp_11.pdf.
Mahendra W. 2010. Pola segregasi karakter agronomi tanaman kedelai (Glycine max L. Merrill) generasi F2 hasil persilangan wilis x B3570. J Agrotek Tropika. 1(1):8-13.
Murata Y. 1969. Morfologi dan fisiologi tanaman padi. [Internet]. [diunduh 2015 Jul 09]. Tersedia pada: http://www.litbang.deptan.go.id/special/padi/
bbpadi_2009_itkp_11.pdf.
22
Poelhman JM, Sleper DA. 1995. Pola pewarisan adaptasi kedelai (Glycine max L. Merrill) terhadap cekaman naungan berdasarkan karakter morfo-fisiologi daun. Bul Agron. 36(1):1-7.
_____________________. 1996. Penyuaian data dan penggunaan informasi kekerabatan untuk mendeteksi segregan transgresif sifat kuantitatif pada tanaman menyerbuk sendiri. Jurnal Budidaya Pertanian. 5(1):11-18. Purwono, Purnamawati H. 2007. Budidaya 8 Jenis Tanaman Pangan Unggul.
Jakarta (ID): Penebar Swadaya.
Puspitasari W. 2011. Pendugaan parameter genetik dan seleksi karakter agronomi dan kualitas sorgum di lahan masam [tesis]. Bogor (ID): Institut Pertanian Bogor.
Rachmawati R, Kuswanto, Purnamaningsih SL. 2014. Uji kesegaman dan analisis sidik lintas antara karakter agronomis dengan hasil pada tujuh genotip Padi Hibrida Javonica. Jurnal Produksi Tanaman. 2(4):292-300.
Robson DS. 1956. Application of K4 statistic to genetic variance component
analyses. Biometrics. 12(4):433-444.
Roy D. 2000. Plant Breeding: Analysis and Exploitation of Variation. Calcutta (IN): Narosa Publishing House.
Sadimantara GR, Widarsih A, Muhidin. 2013. Seleksi beberapa progeni hasil persilangan padi gogo (Oryza sativa L.) berdasarkan karakter pertumbuhan tanaman. Jurnal Agroteknos. 3(1):48-52.
Siregar H. 1981. Morfologi dan fisiologi tanaman padi. [Internet]. [diunduh 2015 Jul 09]. Tersedia pada: http://www.litbang.deptan.go.id/special/padi/ bbpadi_2009_itkp_11.pdf.
Stanfield WD. 1983. Theory And Problems of Genetics 2nd Schain’s Outline Series. New Delhi (IN): Mc Grow thill Book Co.
Suprapto, Kairudin MD. 2007. Variasi genetik, heritabilitas, tindak gen, dan kemajan genetik kedelai (Glycine max Merrill) pada ultisol. JIPI. 9(2):183-190.
Susanto U, Daradjat AA, Suprihatno B. 2003. Perkembangan pemuliaan padi sawah di Indonesia. Jurnal Litbang Pertanian. 22(3):125-131.
Sutjahjo SH, Rustikawati, Sandhi SG AW. 2007. Kajian genetik dan seleksi genotipe S5 kacang hijau (Vigna radiata) menuju kultivar berdaya hasil tinggi dan serempak panen. Agrin. 11(1):10-18.
Walpole RE. 1992. Pengantar Statistika Edisi ke-3. Jakarta (ID): PT Gramedia Pustaka Utama.
Wasim MP. 2002. A study of rice in the major growing countries of the world their growth instability and world share. Pakistan Economic and Social Review. 40(2):153-183.
Welsh JR. 1991. Dasar-dasar Genetika dan Pemuliaan Tanaman. Mogea JP, penerjemah. Jakarta (ID): Erlangga. Terjemahan dari: Fundamental of Plant Genetics and Breeding.
Wirnas D, Sobir, Surahman R. 2005. Pengembangan kriteria seleksi pada pisang (Musa sp.) berdasarkan analisis lintas. Bul Agron. 33(3):48-54.
Wirnas D, Widodo I, Sobir, Trikoesoemaningtyas, Sopandie D. 2006. Pemilihan karakter agronomi untuk menyusun indeks seleksi pada 11 populasi kedelai generasi F6. Bul Agron. 34(1):19-24.
23
Jul 09]. Tersedia pada: http://www.litbang.deptan.go.id/special/padi/ bbpadi_2009_itkp_11.pdf.
24
LAMPIRAN
Lampiran 1 Data varietas tanaman padi
Nama varietas : IR64
Nomor seleksi : IRI8384-36-3-3 Tahun dilepas : 1986
SK Menteri Pertanian : 449/Kpts/TP.240/07/1986 tanggal 17 Juli 1986 Asal seleksi : Pesilangan IR5657/IR2061
Golongan : Cere
Umur tanaman : 110-120 hari Bentuk tanaman : Tegak Tinggi tanaman : 85 cm
Anakan produktif : 20-35 batang Warna kaki : Hijau
Nama varietas : Situ Patenggang Nomor seleksi : BP1153-C-9-12 Tahun dilepas : 2003
SK Menteri Pertanian : 385/Kpts/SR. 120/7/2003 Tetua : Kartuna/TB47H-MR-10
Rata-Rata hasil : 3.6-5.6 ton/ha GKG Golongan : Cere
Umur tanaman : 110-120 hari setelah tanam Bentuk tanaman : Tegak
Tinggi tanaman : 100-110 cm
Daun bendera : Menyudut 35-50 (derajat) terhadap batang Bentuk gabah : Agak gemuk
25
Kerontokan : Sedang Kerebahan : Tahan Tekstur nasi : Sedang Kadar amilosa : 23,93% Indeks glikemik : 53.7
Bobot 1000 butir : 26.5-27.5 gram (KA 14%) Ketahanan terhadap
Penyakit : Tahan terhadap blas Diferensial
Sifat khusus : Aromatik, respon terhadap pemupukan sehingga mampu dikembangkan di sawah
Anjuran tanam : Lahan kering musim hujan, tumpangsari, lahan tipe tanah Aluvial dan Podsolik ketinggian tidak lebih dari 300 m dpl Pemulia : Ismail BP, Atiti D.S, Yamin S., Z. A. Simanullang, dan Aan
A. Daradjat
Nama varietas : Mekongga
Nomor seleksi : S4663-5d-Kn-5-3-3 Tahun dilepas : 2004
SK Menteri Pertanian : 374/kpts/LB.420/6/2004 Asal seleksi : A2790/2*IR64
Umur tanaman : 116-125 hari setelah tanam Bentuk tanaman : Sedang
Tinggi tanaman : 91-106 cm Daun bendera : Tegak
Bentuk gabah : Ramping panjang Warna gabah : Kuning bersih Kerontokan : Sedang Tekstur nasi : Pulen Kadar amilosa : 23% Indeks glikemik : 88
Bobot 1000 butir : 27-28 gram Potensi hasil : 6 ton/ha GKG Ketahanan terhadap
hama : Agak peka terhadap wereng coklat biotipe 2 dan 3 Ketahanan terhadap
penyakit : Agak peka terhadap hawar daun bakteri strain IV Sifat khusus :
Anjuran tanam : Baik ditanam di sawah dataran rendah sampai ketinggian 500 m dpl
Pemulia : Z. A. Simanullang, Idris Hadade, Aan A. Daradjat, dan Sahardi
26
Lampiran 2 Data iklim bulanan bulan November 2014 – Maret 2015 Bulan Suhu rata-rata (oC) Curah hujan
(mm bulan-1)
Kelembaban (%)
November 2014 26.3 673.2 64
Desember 2014 26.3 209.5 45
Januari 2015 25.2 251 87
Februari 2015 25.0 351 88
Maret 2015 25.6 374 85
Rata-rata 25.7 371.7 73.8
Sumber: Badan Meteorologi Klimatologi dan Geofisika (BMKG) Stasiun Klimatologi Darmaga, Bogor
27
RIWAYAT HIDUP
Penulis dilahirkan di Perdagangan, Sumatera Utara pada tanggal 06 Maret 1993. Penulis merupakan anak pertama dari lima bersaudara dari pasangan Hotlan Situmorang dengan Sumiatik. Tahun 2005 penulis lulus dari SD Negeri 091695 Afdeling III Mayang, kemudian pada tahun 2008, penulis menyelesaikan pendidikan di SMP Swasta PTPN IV Kebun Mayang, dan pada tahun 2011 penulis lulus dari SMA Negeri 1 Pematangsiantar dan pada tahun yang sama penulis lulus seleksi SNMPTN Undangan di Institut Pertanian Bogor di Departemen Agronomi dan Hortikultura.
Selama mengikuti perkuliahan, penulis aktif mengikuti berbagai kegiatan organisasi kemahasiswaan. Pada tahun 2011, penulis aktif menjadi anggota PASKIBRA IPB dan pada tahun yang sama, penulis juga aktif menjadi staff
Lembaga Pengajaran Qur’an (LPQ) dan Bina Desa (Bindes) IPB. Kemudian pada tahun 2012-2013, penulis aktif menjadi staff Islamic Student Center (ISC) IPB, tahun 2013-2014 aktif juga menjadi Kepala Departemen Pengembangan Sumber Daya Mahasiswa (PSDM) Himpunan Mahasiswa Agronomi (HIMAGRON).