NURHAIDAH IRIANY SINAGA
DEPARTMENT OF BIOLOGY
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
Guinea is my own work and effort directed by supervisory commission and has not been submitted in any forms to any other universities for my award. The information sources from the published or unpublished references of other authors have been acknowledged and written in the references at the end of this dissertation.
Bogor, March 2011
Prihardhyanto Keim).
FreycinetiaGaud. in West New Guinea had been investigated. For about 4.000 specimen collections were studied using morphological, ecological and molecular data of cp DNA sequence trnL-F, atpB-rbcL and trn-L . The result showed that in West New Guinea 92 species occur. All species can be divided into 4 groups namely F. macrostachya, F. funicularis, F. oblanceolata and F. angustissima groups. The F. macrostachya group have 36 species and 12 species are prepare as a new species namely F. aurihasteta Sinaga, F. brevipedicelata Sinaga, F. clavata Sinaga & Utteridge, F . circuita Sinaga, F.concordia Sinaga,F. frutaspiralica Sinaga,F. frutonumerataSinaga, F. mandacana Sinaga, F.megaauriculata Sinaga & Utteridge, F. stigmabeliata Sinaga, F. ultrapedicelata Sinaga and F. yapenina Sinaga. The F. funicularis group have 17 species and 9 species are prepare as a new species namely F. ayawasa Sinaga, F. fusiformaSinaga & A.P. Keim,F. hasteta Sinaga,F. imbristigma Sinaga & A.P. Keim, F. magnoareola Sinaga,F.millikenus Sinaga & A.P. Keim,F. nervoauricula Sinaga, F. spiroaxilla Sinaga & A.P. Keim and F. simpliaxillata. The F. oblanceolata group have 31 species and 9 species are considerated as a new species namely F. albaauria Sinaga,F. batantaensis Sinaga, F. decendataSinaga, F. folitenella Sinaga, F. frutasolla Sinaga, F. broccoareola Sinaga, F. iriana Sinaga,F. manokwarianaSinaga and F. verstegiiSinaga. The last group is F. angustissima group that have 8 species but only one as a new species namely F. pauciberria Sinaga. The F. macrostachya and F. oblanceolata groups living from forest beach to 1700 m asl butF. macrostachya group are found alone in secondary forest. TheF. funicularis andF. angustissimagroups prefer living on the mountain area and both of them are found on 3000 m asl. 35 species has limited distribution it is only found in Papua but 52 species occurs both in Papua &PNG and 5 species are widely distribution throughout Malesia and Australia. Analysis phylogenetic through PAUP program using both morphological and molecular data support the groups with CI (Coefficient Index) = 0.7, HI (Homoflashy Index) = 0.3 for morphology characters and CI =0.930; 0.868; 0.890 and HI = 0.07; 0.132; 0.110 for molecular data bytrnL-F; atpB- rbcLand trn-L sequencing of CP DNA. According to morphological data F. macrostachya group is a primitive one, It is followed by F. funicularisgroup,F. oblanceolata andF. angustissima groups as an advance one. But in the trnL-F sequencing cpDNA data F. macrostahya group as a primitive one separate from 3 other derivate groups and this result are supported by ecological situation. On the other hands, atpB–rbcL sequencing strongly support the groups but advance one is not belong to theF. angustissimagroup anymore butF. funicularisgroup, The same way is found in the trn-L sequencing data. Therefore trnL-F sequencing had given more conservative characters butatpB-rbcLshowed both conservative and advance characters whiletrn-Lhad given more advance characters.
Freycinetia Gaud. di Nugini Barat telah diteliti. Sebanyak kurang lebih 4000 lembar spesimen herbarium telah dipelajari, menggunakan data morfologi, ekologi dan molekular dengan sekuensing kloroplast DNA yakni sekuens trnL-F; atpB-rbcL ; dan trn-L .Hasil penelitian menunjukan bahwa di Nugini Barat terdapat sebanyak 92 jenis Freycinetia . Jenis-jenis Freycinetia ini terdiri atas 4 kelompok yakni kelompok F. macrostachya, F. funicularis, F. oblanceolatadan kelompok F. angustissima. Kelompok F. macrostachya memiliki 36 jenis dengan 13 jenis baru yaitu F. aculeata Sinaga, F. aurihastata Sinaga, F. brevipedicelata Sinaga, F. clavata Sinaga & Utteridge, F .circuita Sinaga, F.concordia Sinaga, F.frutaspiralica Sinaga, F. frutonumerata Sinaga, F. mandacana Sinaga, F.megaauriculata Sinaga & Utteridge, F. stigmabeliata Sinaga, F. ultrapedicelata Sinaga dan F. yapenina Sinaga. Kelompok F. funicularis memiliki 17 jenis dengan 9 jenis baru yaitu F. ayawasa Sinaga, F. fusiforma Sinaga & A.P.Keim, F. hastatus Sinaga, F. imbristigma Sinaga & A.P. Keim , F. magnoareola Sinaga, F. millikenus Sinaga & A.P. Keim, F. nervoauricula Sinaga, F. spiroaxilla Sinaga & A.P. Keim dan F. simpliaxillata. Kelompok F. oblanceolata memiliki 31 jenis dengan 10 jenis baru yakni F. albaauria Sinaga, F. batantaensis Sinaga, F. decendata Sinaga, F. folitenella Sinaga, F. frutasolla Sinaga, F .gunungmejensis Sinaga, F. broccoareola Sinaga, F. iriana Sinaga, F. manokwariana Sinaga dan F.verstegii Sinaga. Kelompok F. angustissima memiliki 8 jenis dengan 1 jenis baru yaitu F. pauciberria Sinaga. KelompokF. macrostachya danF. oblanceolata hidup di hutan pantai hingga ketinggian 1700 m dpl. Sementara kelompok F. funicularis dan F. angustissima menyenangi tempat tinggi, hingga ketinggian 3000 m dpl. Sebanyak 35 jenis hanya ditemukan di Nugini Barat, namun terdapat 52 jenis yang juga hidup di PNG dan 5 jenis memiliki persebaran luas hingga ke Malesia bahkan Australia. Keempat kelompok ini didukung hasil analisis filogenetik dengan program PAUP berdasarkan data morfologi dan molekular, dengan nilai CI (Coefficient Index)= 0.7, HI (Homoflashy Index) = 0.3 untuk data morfologi dan nilai CI =0.930; 0.868; 0.890 and HI = 0.07; 0.132; 0.110 untuk data molekular sekuensing DNA kloroplast berturut-turut trnL-F; atpB-rbcL; dan trn-L. Berdasarkan data morfologi kelompok F. macrostahyaadalah kelompok primitif, dikuti kelompokF. funicularis,, F. oblanceolata dan F. angustissima. Tetapi sekuensing trnL-F lebih memisahkan kelompok primitif dari ketiga turunannya dan data sekuensing ini sangat didukung oleh data ekologis. Di sisi lain, sekuensingatpB-rbcL mendukung keempat kelompok ini, namun kelompokF. angustissima tidak lagi sebagai yang lebih maju melainkan kelompok F. funicularis, keadaan yang sama terjadi dalam sekuensing trn-L. Dengan demikian trnL-F lebih banyak menggambarkan karater yang konservatif sementara atpB-rbcL menggambarkan keduanya baik sifat konservatif maupun sifat yang lebih maju dan trn-Lsekuensing lebih menggambarkan sifat yang lebih maju.
at least 60 species have been recognized (Stone 1976; 1983). This number is much higher compare to Borneo (24) and the Malay Peninsula (8). The majority of herbarium specimens of Freycinetia from New Guinea are deposited only in few Herbaria such as Herbarium Bogoriense (BO) in Bogor-Indonesia, Herbarium Manokwariense (MAN) in Manokwari-Indonesia, Herbarium Lae in Lae-Papua New Guinea, the National Herbarium of the Netherlands in Leiden (L)-the Netherlands, and Herbarium Kewensis in Kew-UK. Unfortunately, due to the incomplete nature of the specimens most are labelled as unknown species. From time to time deposit specimens increase in numbers but many of them are still unidentified. Therefore, it need taxonomical study to find out how many species of New GuineaFreycinetiaespecially in the west area.
First serious molecular study of the family was proceeded by Callmander et al. (2003). Unfortunately, the infrageneric classification of the genus is still unresolved. So the aims of this study were to unravel the diversity of Freycinetia in the western part of New Guinea, revision of the genus particularly the infrageneric classification and knowing the species distribution in western part of New Guinea in relation with New observation in the field and morphological observation in herbarium B0 (2006, 2008), MAN (2006), LAE (2006), L (2008) and K (2009). Molecular work using sequence non coding cpDNA: trnL-F, trn-L and atpB – rbcL was done at Plant Biology and Biotechnology Laboratorium IPB (Bogor Agricultural University) on August to October 2008 and van der Klauw Molecular Laboratorium in Leiden on November 2008 until January 2009.
The result showed that 92 species of Freycinetiaoccur in the area. All species can be divided into 4 groups namely F. macrostachya, F. funicularis, F. oblanceolata and F. angustissimagroups.
The F. macrostachya group have robust habit and are high climbers. Stem robust, hard, rarely branched, 2-4 cm diameter. Leaf imbricate, linear or lanceolate, margin with sharp and hard spines, 20-120 cm long, 1.5-7 cm wide; cauline leave colorfull in common, basically leaf with yellow, orange, red or mixed color, consist of 2-3 whorls; Infructescence terminal, ternate, sometimes with 4, 8 to 12 cephalia, spirally to umbelly arranged. Prophyll bracts never found. Cephalium large, 3-20 cm long, 2-6 cm wide. Berries numerous, thin, stout; stigmatic remains 1-3 or 5-6, rarely 8 or 12, one species with 32. Seeds linear or ellipse, usually longitudinally arranged.
species with marginal stigmatic remains. Seed usually flat, transversally arranged.
The F. oblanceolata group possess slender stem, 0.5-2 cm diameter, numerously branching, branch short, less than 50 cm long. Leaf non imbricate, elliptical to oblong and oblanceolate, margin with spines on terminal and basal parts only, 2-15 cm long, 0.5-4 cm wide; Cauline leaves usually green; Prophyll only in both terminalia and axillary inflorescence, if present only consist of 2 whorls; Infructescence usually terminal, rarely both terminal and axillary. Cephalium minute, 2-7 cm long, 1-3 cm wide. Berry numerous or few, some species with flat tips, fusion or self-changing areola present; stigmatic remains 1-3 or 5-6. Seed linear or oblong-flat, usually arranged transversally, in some species longitudinally
The F. angustissima group have slender stem, less than 0.5 cm in diameter, usually numerously branched, branch short, less than 20 cm long. Leaves pseudo-rosette in appearance, each linear or lanceolate, in some members observed elliptical, margin with spines on terminal and basal parts, 2-8 cm long, 0.2-1 cm wide. Prophyll bracts never found. Infructescence terminal, usually binate, rarely single or ternate, arranged in umbel. Cephalium small, 2-4 cm long, 1-2 cm wide. Berry usually few in numbers, some species with flat-tipped berries; stigmatic remains 1 or 2, rarely 3, self-changing areola present giving variation in stigmatic areola. Seeds linear, transversally or longitudinally arranged.
The F. macrostachyagroup have 36 species. The previous species have 23 species. These are F. aculeata Sinaga, F. andajensis Martelli, F. arfakiana Martelli, F. archboldiana Merr. & Perry,F. bicolor B.C.Stone, F .excelsa F. Muell., F. fibrosa Martelli, F. formosula Huynh, F. impundens B.C. Stone, F. Klosii Ridl.,F. lacinulata Kanehira, F. macrostachyaMartelli, F. marginata Blume, F. palida Huynh, F. plana Huynh, F. percostata Merr. & Perry, F. pseudoinsignis Warb., F. rosellana Huynh, F. solomonensis B.C. Stone, F. trachypoda Merr. & Perry, F. tessellata Merr. & Perry, F. undulate Merr. & Perry, and F. whitmorei B.C. Stone. Twelve species are regarded as new species namely F. aurihasteta Sinaga, F. brevipedicelata Sinaga, F. clavata Sinaga & Utteridge,F. circuitaSinaga, F. concordia Sinaga,F. frutaspiralica Sinaga, F. frutonumerata Sinaga, F. mandacana Sinaga, F. megaauriculata Sinaga & Utteridge,F. stigmabeliata Sinaga,F. ultrapedicelata Sinaga and F. yapenina Sinaga.
TheF. funicularisgroup have 17 species. The previous 8 species areF. cryptocarpa Kanehira, F. erythospatha Merr. & Perry, F. funicularis Merr.,, F.gibbsiae Rendle, F. lateriflora Ridll., F. pleurantha Merr. & Perry, F. rhodospatha Ridll., and F. sterrophyllaMerr. & Perry. Nine species are regarded as new species namelyF. ayawasa Sinaga,F. fusiformaSinaga & A.P. Keim, F. hastata Sinaga, F. imbristigma Sinaga & A.P. Keim, F. magnoareola Sinaga, F.millikenus Sinaga & A.P. Keim, F. nervoauricula Sinaga, F. spiroaxilla Sinaga & A.P. Keim and F. simpliaxillata Sinaga.
F. tenuisSolms. Nine species are regarded as new species namely F. albaauriaSinaga, F. batantaensisSinaga, F. decendataSinaga,F. folitenella Sinaga,F. frutasolla Sinaga, F. broccoareola Sinaga, F. iriana Sinaga, F. manokwariana Sinaga and F. verstegii Sinaga.
The F. angustissima group have 8 species and 1 species are regarded as a new species namely F. pauciberriaSinaga. The previous species here areF. angusta Huynh, F. angustissima Ridl., F. brachyclada Huynh, F. linearis Merr. & Perry, F. polyclada Merr. & Perry, F. pseudoangustisimma Huynh and F. stenophyllaWarb.
F. macrostachya & F. oblanceolata groups living from forest beach to 1700 m above sea level. But F. macrostachya group are found alone in secondary forest. on the other hand, theF. funicularis and F. angustissima groups prefer living on mountain area and both of them are found up to 3000 m asl. Despite share with F. angustissima group, theF. funicularis group also share habitat with F. oblanceolata group, especially in the forest beach where F. angustissima group never is found, but this group has ability living on the wet area. West New guinea Freycinetia have 35 species that are limited distribution, because these species are only occur in the area. But West New Guinea Freycinetia share 52 species with PNG, and 5 species remains are widely distribution species that spread through out Malesia region until Australia.
1. It is prohibited to cite all or part of this dissertation without referring to and mentioning the source.
a. Citation only permitted for the sake of education, research, scientific writing, critical writing or reviewing scientific problem .
b. Citation doesn’t inflict the name and honor of Bogor Agricultural University 2. It is prohibited to republish and reproduce all or part of this dissertation without the
NURHAIDAH IRIANY SINAGA
Dissertation
Submitted as a part of requirement for the fulfillment of Doctor Degree in Biology
DEPARTMENT OF BIOLOGY
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
Examiners in comprehensive examination: Dr. Nunik Sri Aryanti, M.Si
Department of Biology, Faculty of Mathematics and Natural Science, Bogor Agricultural University, Bogor, Indonesia
Dr. Ruliyana Susanti
Herbarium Bogoroense, Botany Division, Research Centre for Biology
Institute of Science, Cibinong, Bogor, Indonesia
Examiners in final examination: Dr. Tatik Chikmawati, M,Si
Department of Biology, Faculty of Mathematics and Natural Science, Bogor Agricultural University, Bogor, Indonesia
Dr. Rugayah
Herbarium Bogoroense, Botany Division, Research Centre for Biology
The main object of the research conducted at November 2005 to January 2009 is Freycinetiain West New Guinea. This study emphasis four different topics namely the morphological characters, ecological and distribution, chloroplast DNA sequencing and description of 92 species of New Guinean Freycinetia. A part of morphological approach i.e. The Stigma and Areola in the Western New Guinean Species of Freycinetia Gaud. (Pandanaceae; Freycinetoideae) submmited to the 8 th International Flora Malesiana Symposium in Singapore and Two New Species of Freycinetia (Pandanacea) from Manokwari, West Papua have been published in Reinwardtia 13:2 (2010) 183 - 187. Part two: The Ecology and Distribution of Freycinetia Gaud. (Pandanaceae; Freycinetoideae) in The Indonesian New Guinean have also been published in Reinwardtia 13:2 (2010) 189 -197.
I am extremely grateful to the advisory committees who directed, advised and read critically the manuscript, Dr. Rita Megia, DEA (Department Biology IPB), Prof. Alex Hartana (Department Biology IPB), Prof. Mien A. Rifai (Herbarium Bogoriense, LIPI) and Dr. Ary .P. Keim (Herbarium Bogoriense, LIPI).
Profound thanks and appreciation to Dr. Eko B. Waluyo and all staff of Herbarium Bogoriense especially Dr. Rugayah and Prof. Dr. Elizabeth Widjaja; Prof. Dr. Peter van Welzen, Dr. Barbara Gravendell, Marcel Eurings and Dr. Michael Stech (Leiden University, NHN Herbarium, Nederland); Dr. Rogier de Kok, Dr. William Bekker and Dr. Timothy Utteridge (Kew Herbarium, UK); Roy Banka (Lae Herbarium, PNG); Ir. Jack Wanggai, M.Si and staff (Herbarium Manokwariense) and Dr. Fenny Ismoyo (Papua University); Ir. Pratita Puradyatmika, M.M (PT. Freeport Indonesia) for any kind of help during my research.
I gratefully acknowledge the BPPS Scholarship, Sandwich Program and Litdas from Director General of Higher Education (Dikti), Departement of National Education Republic Indonesia. I thanks Ir. J.P. Karafir M,Sc (Rector Unipa); Dr. Hasyim (Dean Math and Science Faculty of IPB) and Dr. Deddy Duryadi DEA (Head of Program Study Biology Post Graduate School IPB).
I am indebted to all staff and my colleagues in Papua University, my field assistant: Gasper Taudufu, Christhopere Diaz and Tinus Iwanggin and my friends at Subprogram Plant Taxonomy IPB: Dr. Iris Rengganis, Dr. Nursahara Pasaribu, Dr. Sri Endarti, Dr. Puji Widodo, Dr. Fitmawati, Dr. Himah Rustiami, also my brothers: Dr. Soaloon Sinaga, Kristian Sinaga, SKM, Benjamin Sinaga, S.Tp and my sisters: Sesniwati Sinaga, SE and Bertha Sinaga, S.Sos with their families. Finally, I give my honor to my beloved parent: Kasiaman Sinaga and Johana Taudufu.
Nurhaidah Iriany Sinaga was born on 6 January 1969, in Karubaga Papua, as the second daughter of Kasiaman Sinaga and Johana Taudufu. She graduated from
Page
Relationship and Evolution of the Freycinetia...
27 - 38
E. DESCRIPTION OF THE FREYCINETIA IN WEST NEW GUINEA Key to Groups
TheF. funicularis Group ... Key to Species of F. macrostachya Group
Species ofF. macrostachya Group...
F. undulata Merr. & Perry……….
Species ofF. funicularis Group ...
F. pleurantha Merr. & Perry………..
F. rhodospatha Ridll………
Key to Species of F. oblanceolata Group 152
F. obtusiacuminataHuynh………
Key to Species of F. angustissima Group 195
Species of F. angustissima Group... 196
page
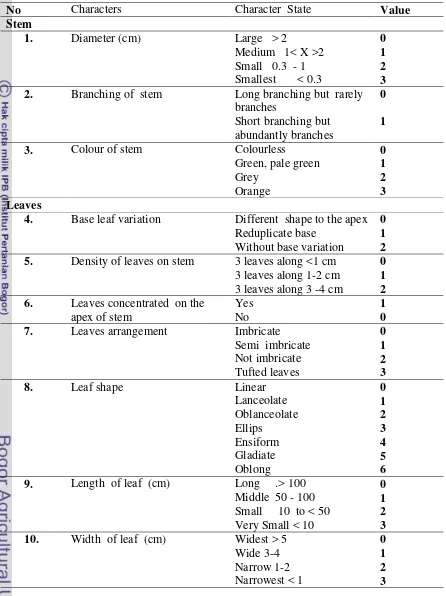
1. Primer Using for Sequensing………..…….5
2. Material for Sequencing DNA...6
3. Morphological characters for PAUPanalysis………. ………..…. 32
4. Distribution of the members of the infrageneric groups suggested ………40
in this current study (A = secondary forest; B = primary beach forest; C = wet land area)
5. Species distribution ofFreycinetiain West New Guinea according to the three ...45 diffrences location
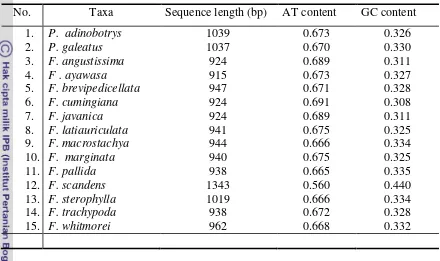
6.Variation of sequence length, AT, GC content oftrnL-Fintergenic spacer
InFreycinetia... 50
7. Deletion, insertion and subtitution on DNA sequence of oftrnL-Fintergenic spacer some Freycinetia...51
8.Variation of sequence length, AT, GC content ofatpB-rbcL intergenic spacer
inFreycinetia...55
9. Deletion, insertion and subtitution on DNA sequence ofatpB-rbcL intergenic spacer some Freycinetia...56
10.Variation of Sequence length, AT, GC content oftrn-Lintergenic spacer
inFreycinetia...59 11.Deletion, insertion and subtitution on DNA sequence of oftrn-Lintergenic spacer
Page
1. The fourth kinds of leaves arrangement of the Freycinetia………..
2. The surface of the lamina ofFreycinetia………
3. The shape and colour of the prophyll leaves……….
4. The variation of the auricle………
5. The variation of cauline leaves………..
6.The prophyll and variation of the bracts………
7. The staminate flowers……... ……….……….
8. The pistillate flowers……….………
9. The variation of the stigmas………..
10. The variation of the areola……….………
11. The variation and density of the berry………..
12. The variation of the seed………
13. Phylogenetic tree show four groups of West New Guinean Freycinetia…
14. Phylogenetic tree show four groups of West New Guinean Freycinetia…
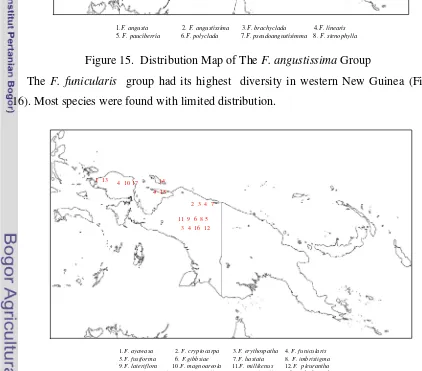
15. Distribution Map of theF. angustissima Group……….
16. Distribution Map of theF. funicularis Group………...
17. Distribution Map of the F. oblanceolata Group……….
18. Distribution Map of theF. macrostachya Group ………...
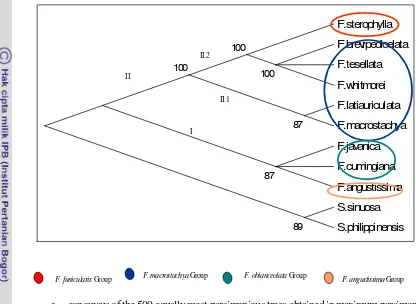
19. Strict consensus of the 500 equally most parsimonioustrees obtained in ……
maximum parsimony analysis of thetrnL-Fsequences data.
20. Strict consensus of the 500 equally most parsimonious trees obtained in ……
21. Strict consensus of the 500 equally most parsimonious trees obtained in……
maximum parsimony analysis of the trn -Lsequences data.
22. New Species of F. macrostachya Group
1.F. aurihastataSinaga………….…………..………
2.F. brevipedicelataSinaga…………. ………….……….
3.F. clavata Sinaga & Utterdige…………..………
4.F. circuita Sinaga………….………
5.F. concordia Sinaga………….……….
6.F. frutaspiralica Sinaga…………..………..
7.F. frutonumerata Sinaga…………...………..
8.F. mandacana Sinaga………….………
9.F. megaauriculata Sinaga & Utteridge…………....………..
10.F. stigmabeliata Sinaga……….………
11.F .ultrapedicelataSinaga…………...………
12.F. yapenina Sinaga…………...………
23. New Species ofF. funicularis Group
1.F. ayawasa Sinaga…………..………
2.F. fusiforma Sinaga & A.P.Keim…………...………
3.F. hastata Sinaga………….………. .
4.F. imbristigma Sinaga & A.P. Keim...…..…………..…………
5.F. magnoareola Sinaga…………..………
6.F. millikenus Sinaga & A.P. Keim…………..………...
7.F. nervoauriculata Sinaga…………....……….
8.F. spiroaxillaSinaga & A.P. Keim…………..……….
9.F. simpliaxillata Sinaga…………. ………...
24. New Species of F. oblanceolata Group
1.F. albaauria Sinaga, ………….………
25. New Species of F. angustissima Group
Name : Nurhaidah Iriany Sinaga Registration Number : G361040051
Program Study : Biology
Approved by
1. Advisory Committees
Dr. Rita Megia, DEA (Chairman)
Prof. Dr. Ir. Alex Hartana Prof. Dr. Mien A. Rifai Dr. Ary P. Keim (Member) (Member) (Member)
2. Program Study of Biology 3. Post Graduate School
Dr. Dedy Duryadi Solihin, DEA Dr. Ir. Dahrul Syah, MSc. Agr (Head) (Dean)
Martelidendron. Freycinetia was described in 1824 by the French botanist Gaudichaud. Until now approximately 200 species present in the world. But this genus is not very widespread and the real home of Freycinetia is in Malaysia, Indonesia, Philippines, Formosa, Okinawa, New Guinea and surrounding islands, Australia and and New Zealand. The centre of Freycinetia distribution is certainly from the Philippines to New Guinea (Stone, 1970). The New Guinea islands according to Stone (1976, 1983) may be have 60 species, while in comparison Malaya has 8 species and Borneo has 24 species.
The genus are climbing pandan except F. arborea in Hawai . Freycinetia prefer living near rivers or in humid areas especially under canopy. Freycinetia could be found from the sea level to 2400 m asl . It is restricted in their habitat. For example F. elegantula and F. pseudoangustisimma that are found only in mountains area but only few species could be found both in the low land and mountain area for example F. funicularis. The limitation area of occupation may related to the agent of distribution. Unlike Pandanuswith used win as a distribution agent,Freycinetiaseeds are spread by mammals and birds . That is why most species have colourfull beauty bracts to attract animals to visit the flower.
species from Papua and 25 species were new species.
Many specimen from Papua are collected and put in Herbarium Manokwariense (Man) in Manokwari, Bogoriense (BO) in Bogor, Lae in Morobe PNG, NHN (L) in Leiden, Nederland and Kew (K) in England . Most of them were put as unknown species. For example in BO, from 200 specimens only 42 specimens are known species. How many species exactly occur in Papua need a taxonomical work to find out.
Inter species classification of Freycinetia were published first by Warburg in 1900 which about 2 sections and it was followed by Stone (1968) who mentioned about 15 sections. Warburg worked based on number of stigma namely Oligostigma section with 1-3 stigmas and Pleiostigma with 3 or more stigmas. Stone mentioned section based on raceme and umbel inflorescence and the umbel one were divided for terminal versus lateral or both lateral and terminal inflorescence. Next the terminal were divided again based on pseudopetiole, cephalia, berries, stigma and auricle. However, some specimens have possibilities to put under 2 section. likeF. archboldiana
Other taxonomy problems are sinonim name of Freycinetia. Kanehira and Hatusima in 1940 mentioned F. inouei as a new species from Dalman in Nabire, West New Guinea. They mentioned that the species have closely related toF. archboldianabut different species. Stone in 1972 put theF. inoueias sinonim ofF. archboldiana.
Some botanist like Martelli in 1910; Merril & Perry in 1940; Kanehira in 1941 and Stone in 1967 said that F.angustissima were sinonim to F. stenophylla Warb. and F. polyclada Merr.& Perry. But, Huynh in 1999 mentioned the three species above were different species. He used both morphological and anatomical characters. Huynh said that in morphological characters F. angustissima and F. polyclada were showed their differences. He also said that the stigma of F. angustisimma is 2 0r 3 rarely 1 or 4 but stigma and areola is distinctly girdled and F. stenophylla has 2 stigmas with sitgma and areola is un-distinctly girdled. Actually, our preliminary study showed that stigma and areola in west New GuineanFreycinetiahave many variation and many specimens with unidentified label have the variation.
morphological approached but only in generic level. Further, Callmander et. al (2003) produced phylogenetic trees based on cp DNA data on generic levels too.
Some Freycinetia species without flowers and good auricle looks similar like F. palida and closely unknown species. For this species information from molecular data will guide to understanding what species is, is it same or different species. However, for build a good phylogenetic classification it need comprehensive approached.
According to FWI (Forest Watch Indonesia) report in 2004, deforestation happen in Indonesia, with acceleration of degradation 3 times foot ball area in every minutes. Consequently some species include Freycinetia may disappeared without ever identified .
Freycinetiahave much utilities . All parts of this plant could be used, for example hanging root for nail; stem for lance, traditional rain clothes; leaves for roof, and handicraft like basket and bag, also for perfume; spadix and cephalia as foods; and bracts for cooling drink and juice. To get more benefit for human life, more researches need start from basic study. Therefore the taxonomic research were done using main morphological information and molecular data as a second approach.
Purposed of the Study
The purposed of the West New GuineanFreycinetiastudy were: 1. To know the diversity ofFreycinetiain West New Guinea 2. To obtain delimitation of the genus and species concepts
3. To build phylogenetic classification ofFreycinetiabased on morphological and molecular data.
The Pandanaceae has four genera namely Pandanus, Freycinetia, Sararanga and Martelidendron. They all occur in the old tropicals, from Africa to India, Indo-China to Japan, Malaysia to Sumatera until New Guinea area include some islands surroundings like Salomon islands and also present in Australia. Two genera namely .Martelidendron andSararanga have limited in both species number and distribution. Martelidendrohas 6 species found only in Madagaskar and some surrounding islands. Sararanga has 2 species, one species only found in Philippines and the other one in New Guinea and Salomon Islands.
Freycinetia was described in 1824 by the French botanist Gaudichaud. Until now approximately 200 species present in the world. But this genus is not very widespread and the real home ofFreycinetia is in Malaysia, Sumatera, Java, Borneo, Lesser Sunda Islands, Celebes, Philippines, Formosa, Okinawa, New Guinea, The Solomon Islands, New Hebrides (Vanuatu), The Marianas and Carolines, Hawaii, Society Islands, New Caledonia, Queensland and New Zealand. The centre of Freycinetia distribution is certainly from the Philippines to New Guinea (Stone, 1970). The New Guinea islands according to Stone (1976, 1983) may be have 60 species, while in comparison Malaya has 8 species and Borneo has 24 species.
The genus are climbing Pandan except F. arborea in Hawai . It could be found from the sea level to 2400 m asl . It is restricted in their place. For example F. elegantula and F. pseudoangustisimma that are found only in mountains area. Some species could be found both in the low land and mountain because It is brought by the water flood for example F. funicularis. Many species of Freycinetia are found abundantly only near rivers or in humid areas especially under canopy.
island.
Some botanists have been published Freycinetia species from West New Guinea. Solms et.al (1883) mentioned 3 species, of which 2 have been previous published and one new species. They published Beccari works in Andai Manokwari. Martelli (1910) mentioned about four new species, three species from Andai - Manokwari and one from Korido - Biak. Followed by Rendle (1917), who published two new species from Arfak mountain, Manokwari. Ridley ( 1916-1922) published four new species from Tembagapura. Next, Merril and Perry (1940) published 4 new species, 2 species from Tembagapura and 2 species from Habema, Wamena. In 1941 Kanehira & Hatusima along their journey in Manokwari and Nabire mentioned 9 new species and in 1976 Schumann & Lauterbach mentioned 1 species from Jayapura, followed by Huynh (2002) who published 1 new species from Tembagapura . Therefore they all found totally 36 species from Papua and 25 species of them were mentioned as a new species.
From east part of New Guinea, more species were published by Warburg (1900), Marteli (1910), Merril & Perry (1940), Stone (1963) and Huynh (1996 - 2003). Huynh mentioned about 87 new species into 8 parts journals of New Guinea Freycinetia . Few of the species are also found in west area. Huynh and Stone also published many species from Salomon island and few of them present in Papua too like F. salomonensis Stone andF. planaHuynh.
According to International Plant Nomenclature Index for about 135 names of Freycinetiapresent in New Guinea. Perhaps some are synonymous name, but it show that many species occurs in the area. How many species exactly occur in Papua need a taxonomical work to find out. Indeed, many specimen from Papua are collected and put in Herbarium Manokwariense (Man) in Manokwari, Bogoriense (BO) in Bogor, Lae in Morobe PNG, Rijks (L) in Leiden Nederland and Kew (K) in England . Most of them were put as unknown species. For example in BO, from 200 specimens only 42 specimens are known species.
and it was followed by Stone (1968 ) who mentioned about 17 sections. Warburg worked based on number of stigma namely Oligostigma section with 1-3 numbers of stigma and Pleiostigma with 3 or more stigma. Stone mentioned section based on raceme and umbel inflorescence and the umbel one were divided for terminal versus lateral or both lateral and terminal inflorescence. Next the terminal were divided again based on pseudopetiole, cephalia, berries, stigma and auricle. But, in fact some specimens have slightly umbel or closely spirally arrangement. This species could be put on both raceme and umbel section. Actually, raceme and umbel inflorescences have different kind of bracts but without seen the living collection, the kinds of spirally bracts is difficult to understand so it is not easy for their classification. All section here were never analyzed by phylogenetic or phenetic approached to make sure about the sections. Therefore, some species from New Guinea like F. archboldiana could be put on 2 sections.
unidentified label have the variation.
Solving all problems above need taxonomy work also phylogenetic analysis to make sure about the sections. In Freycinetia species phylogenetic analysis even used morphological approach never been done. Cox already made cladistic analysis based on morphological approached but only in generic level. Further, Callmander et. al (2003) produced phylogenetic trees based on cp DNA data on generic levels too. Some Freycinetia species without flowers and good auricle looks similar like F. palida and closely unknown species. For this species information from molecular data will guide to understanding what species is, is it same or different species. However, for build a good phylogenetic classification it need comprehensive approached.
Morphological Work
The research was carried out from Jully 2005 until Maret 2009. Taking and collecting specimens were done at several area in west New Guinea such as Timika (2005), Manokwari (2006), Sarmi (2006) and Jayapura (2006, 2008). Followed by ecological observation in the field and morphological observation in herbarium Bogoriense in Bogor( 2006, 2008), herbarium Manokwariense in Manokwari (2006) , LAE herbarium in Lae, Morobe Province – PNG (2006), Leiden Herbarium in Leiden Netherland (2008) and Kew herbarium in Kew, England (2009).
Terminology used follows Harris and Harris (1954), Strearn (1966) Radford, et. al (1976), and Radford (1986). Step activities followed those as described by Rifai (1976), Radford (1986) and Vogel (1987). The work previously done from a fundamental for further study, therefore the literatures pertinent to the treatment have been carefully studied and characters have been studied throughout the entire range of the taxon to determine any inter-gradation between two or more taxa. Finally, phylogenetic analysis using PAUP program version 32 were done with Sararanga as an out group.
Molecular Work
Primers and Material for Sequencing
The primers that are used for sequencing show on table 1.
Table 1. Primers using for sequencing on Taxon 52 (4), 747-762 (2003) Primer Sequence (5’ to 3’)
PrimeratpB GAAGTAGTAGGATTGATTCTC
PrimerrbcL TACAGTTGTCCATGTACCAG
Primer c CGAAACTGGTAGACGCTACG
Primer d GGGGATAGAGGGACTTGAAC
Primer e GGTTCAAGTCCCTCTATCCC
Primer f ATTTGAACTGGTGACACGAG
Samples were obtained from both living plants and herbarium specimens that are collected from several area. The out group was obtain and chosen from Gen Bank by Callmander et. all. All material sequencing are shown on table 2.
DNA Extraction, Amplification and Sequencing
Total DNA was extracted from leaves preserved in silica gel , material herbarium and fresh material from about 40 species. Extraction was performed using the CTAB procedure modified of Doyle & Doyle (1987). Double -stranded DNA was directly amplified by PCR for all loci. Reaction volumes were 25 µl and contained MilliQ = 17,2, template =2 µl, Buffer 10 x = 2,5µl, dNTP’s 2,5 mM = 1,0 µl, Primer foward
2,5 mM = 0,1 µl, Primer rivers 2,5 mM = 0,1 µl, MgCl2 25mM = 1,5 µl, TAQPOL 5 U/ µl = 0,1 µl, for trnL-F andtrn-L. Primer that are used for trn-L are primer c and primer d and for trnL-F are primer e and primer f. Reaction volumes for atpB – rbcL have different from primer, that are primer f and primer r, for 1 µl and volume of template is 5 µl and TAQPOL 5 U/ µl is 0,3 µl.
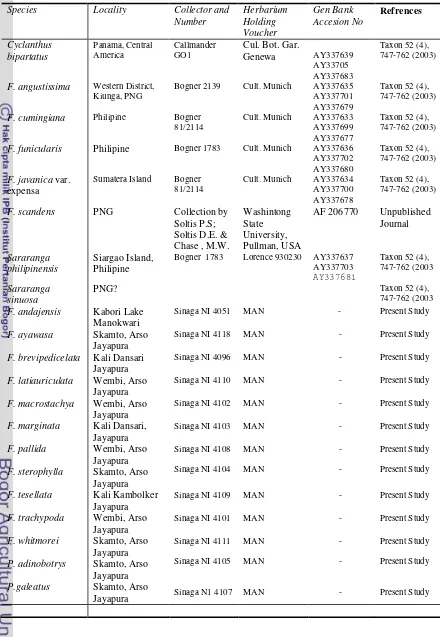
Species Locality Collector and
F. funicularis Philipine Bogner 1783 Cult. Munich AY337636 AY337702
Sinaga NI 4051 MAN - Present Study
F. ayawasa Skamto, Arso Jayapura
Sinaga NI 4118 MAN - Present Study
F. brevipedicelata Kali Dansari Jayapura
Sinaga NI 4096 MAN - Present Study
F. latiauriculata Wembi, Arso Jayapura
Sinaga NI 4110 MAN - Present Study
F. macrostachya Wembi, Arso Jayapura
Sinaga NI 4102 MAN - Present Study
F. marginata Kali Dansari, Jayapura
Sinaga NI 4103 MAN - Present Study
F. pallida
Sinaga NI 4109 MAN - Present Study
F. trachypoda Wembi, Arso Jayapura
Sinaga NI 4101 MAN - Present Study
promega wizard S V gel and PCR clean up system. Cycle sequencing was performed according to the dideoxy chain termination method using promega system. Cycling parameter were 25 cycles of 20 sec at 96 º C for denaturation, 10 sec at 54 º C for primer anneling and 4 min at 60 º C for primer extension. The cycle sequencing products were cleaned by ethanol precipitation and applied to Promega system. All cycle sequencing process were done in Korea by sending PCR product.
Sequence Alignment and Phylogenetic Analysis.
Sequences were aligned using Mega program after do blast and editing sequencing data by sequencer program. Editing were done by compilation data from two kinds of primers with making contig , for trn L-F data, primer eas a forward and primer f as a rivers ; for trn Lwith primer c as a forward and primer d as a rivers and foratpB- rbcL with primer f as a forward and primer r as a rivers. If the data can’t contig , one good
Based on morphological characters western New Guinea Freycinetia has 92 species that are grouped into 4 groups namelyF. macrostachya, F. funicularis, F. oblanceolata and F. angustissima groups. The variation could be seen in many plants organs like stem, leaves, inflorescences, infructusence, fruit and seed.
Stem
Stems of West New GuineanFreycinetiavary in size, colour, and nodes. In term of size, the F. macrostachya group includes the largest and most robust members of the genus. On the other hand, the F. angustissima group includes the smallest and most slender species. The situation is rather different in colour and nodes. So far the distinctive stem colouration commonly found in the F. oblanceolata group, in which the stem colour is orange. Other groups possess more colourfull stem such as green, grey, light yellow, and light green . Almost all species have cylindrical nodes except few species within the group of species with F. oblanceolata group such as F. marantifolia and F. ellipsoidalis that possess obovoid nodes. Other species, such as F. gunungmejensis have dots around the internodes. Nothing is known about this morphological structure. It is suggested here that the dots support the connection
between leaf and stem.
There are variations in the leaves arrangement, shapes, sizes, apex, and laminar surfaces. The F. macrostachya group possess continuous overlapping arrangement of leaves (Fig. 1A). If the species possess less continuous overlapping imbricate leaves arrangement, and imbricate leaves in the middle part till the apex of the stem, and leafless in the basal half part of their stem they are included in the other group called the F. funicularis group (Fig. 1B). If the leaves of species are entirely composed of non imbricate, they are included in the F. oblanceolatagroup (Fig.1C). The F. angustissima group are obviously consists of species with slender-very narrow leaves in the pseudo rossete (Fig.1D).
B C D
A
1 . 5 c m
1 . 5 c m
4 c m 6 c m
Fig. 1. The fourth kinds of leaves arrangement of the Freycinetia: A. The imbricate leaves ; B. The semi imbricate leaves; C. The non imbricate leaves; D. The tufted leave with pseudo rosette.
ellipsoidal, oblong, ovate, obovate to oblanceolate.
The nature of the leaf apexs also varies. Caudate and cuspidate leaf apexs are commonly found in theF. macrostachyagroup. Mucronate leaf apex is unique to the F. oblanceolatagroup. TheF. funicularis and F. angustissima group usually found with acuminate leaf apexs. Acuminate leaf apexs are rarely seen in the other groups.
The longitudinal nerves are more commonly observed in the F. macrostachya group except inF. ellipsoidalis, which is a member of theF. oblanceolata group. Freycinetia ellipsoidalis possesses 15 longitudinal nerves in half part of its leaf. The number of nerves, both true or pseudo-nerves in adaxial and abaxial surfaces of leaves is unique for each species, thus it is a good character for species identification (Fig. 2). Other unique character is the lines longitudinally stretched between main nerves and leaf margins. This character is usually found in the members of Pandanus, but similar structure (i.e. pseudo longitudinal lines) presence in F. iriana, a species endemic to Western New Guinea.
The leaves early grow in stem axis then spread on axis branches. In the F. angustissima group the leaves are found densely arranged on the terminal parts of the branches only, thus forming a rosette. In the F. macrostachya group the leaves are mostly imbricate spread along the stem axis and possess few branches.
A B C
2 m m 2 m m 1 m m
differ from the group of species with F. funicularis group in the nature of the stem and branches. In the last group the basal parts of the stems and branches are leafless.
Leaf growing mechanism is also unique for each group. The terminal leaves in theF. macrostachya group start growing from the apex after anthesis or fruit maturation. In the F. oblanceolata and F angustissima group the young leaves are not growing from the apexs but from the older leaf axils (i.e. axillary growing). In contrast to these groups, in F. funicularis group the young leaves grow on the lateral stem and this phenomenon makes the members of this group spread lateral stems.
Spines
In species ofFreycinetia from West New Guinea the spines can be found on terminal part (apex) of the leaf, leaf margin and throughout the length of the leaf. Spines can also be found on the middle nerves on abaxial surface of the leaf. Robust spines are commonly found in the members of the F. macrostachyagroup. Minute spines usually seen in the members of theF. oblanceolata group in which the spines are rarely seen in leaf margin and usually absent in the nerves.
The shape of spines vary from pyramidal to triangular, with the size vary from minute to robust and the colour vary from yellow to red or purple. Triangular shape of spines is more common than pyramidal, which is unique to F. macrostachya. In F. aculeata the spines can be both minute and obvious, in which they are regularly arranged throughout leaf margin. The possession of purple spines is unique toF. plana.
Prophyll Leaves
membranaceous, green prophylls leaves that is arranged in 1 tritichous (Fig. 3B) and F. megaauriculata has oblong, greenish red prophylls (Fig. 3C) .
A
C B
3 cm 1 cm
1 cm
Fig. 3. The shape and colour of the prophyll leaves: A. The triangular purple of the prophyll leaves of F. gunungmejensis; B. The lanceolate green prophyll leaves of theF. inermis; C. The oblong greenish red prophylls leaves of theF. megaauriculata.
A
B
D C
3 c m
1 c m
3 c m
2 c m
Auricle is a morphological character almost exclusively owned by species of Freycinetia in Pandanaceae. So far there is only one species from the genus Pandanus that possesses this character, e.i P. pectinatus.
Thus, auricle can be regarded as a good field identification character for Freycinetia. Auricle can be easily observed in fresh materials or immature leaves. The shape, size, texture, nature of margin, nerves (both longitudinal and transversal), and colours of auricle vary between species.
The auricle shape can be tapered, truncated, triangular, rounded, half circular or like a lance. A half circular auricle present inF. megaauriculata( Fig. 4A), while lance auricle present in F. hastata (Fig. 4B). The auricle can be less than 1 cm in F. angustissima group to 20 cm long in F. macrostachya group . The texture can either be membranaceous or fibres. Fibres one present in F. plana( Fig. 4D). The margin can be with minute or fairly obvious spines. The nerves may present in some species but absent in other species. Species with distinctly longitudinal nerves of auricle is F. nervoauriculata. If present the number varies greatly from 2 to 15. Few species has shortly longitudinal nerves, likeF. andajensis while other species has longer transversal nerves. The colour also varies greatly from green (Fig. 4C), pale yellow, orange, red to purple, even mixed.
Cauline leaves are small leaves that situated on the apical leaves under bracts. Mostly species with terminalia inflorescences have cauline leaves or they could be found in speces with both terminalia and axillary inflorescences. Cauline leaves consist of 2 types. First is cauline leaves with widely base leaves that is usually ovate rarely lanceolate and the apex is a true leaves which are lanceolate or linear. The second cauline leaves have never widely basic leaves.
The first type of cauline leaves is belong to the F. macrostachya group and the cauline leaves here mostly colorfull and consist of 2 or 3 rarely 4 whorls (Fig. 5A).
A B C
1 c m
3 c m
3 c m
Fig. 5. The variation of cauline leaves: A. The coloring of the cauline leaves; B. The mix colour of cauline leaves ofF. macrostachya; C. The green small cauline leaves of theF.oblanceolata group.
Freycinetia macrostachya has more white mixed with orange cauline leaves on the lamina and green especially on the apex (Fig. 5B). The cauline leaves in F. macrostachya group is easy dry and firstly falling down, it is followed by bracts.
This cauline leaves usually green or pale green with different or similar shape to the leaves, usually consist of 1 to 2 whorls (Fig. 5C ). It is present even after the bracts is falling down.
Prophyll Bracts
The propyll bracts is the characters belong to the axilary species or species with both axillary and terminalia inflorescences. As consequent the character are found in all F. funicularis group and rarely in F. oblanceolata group. In species belong to F. funicularis group prophyll bracts usually has similar colour to the bracts but harder and smaller than bracts (Fig. 6A). The prophyll bracts is arranged from the smallest one to the biggest on the apex and consist of 3 to more than 8 whorls. Base and apical prophyll bracts could be different or similar in shape or surface of the lamina. Prohyll bracts in species of F. oblanceolata group is less in number, usually consist of 2 whorls; and they are also different in colour to the bracts as example inF. gunungmejensis.
Bracts
This current study recognises 2 types of bracts: Proper and a half leaf bracts. The proper bracts are the true bracts (Fig. 6B). They are covering the inflorescence and usually formed by the three levels of tristichously arranged bracts. The interior bract covers the inflorescence, while the other two namely exterior and middle bracts protect the interior bract and the inflorescence inside. Apart from flowers protection the proper bracts also give rewards to possible pollinators such as insects through fleshy structure and aroma. The aroma of mature bracts is fairly similar to the aroma of sugar (i.e. fructose).
involute. Some species have lanceolate or triangular bracts. In proper bracts the interior bracts are usually lanceolate but variation in shape and size observed in the exterior and middle bracts, especially in the lamina and margin parts.
B
E C
D
A
2 c m
1 m m
2 m m
1 m m
2 c m
Fig. 6. The prophyll and variation of the bracts: A. The prophyll bracts (black arrow); B. The white interior bracts ofF. macrostachya (black arrow); C. The sharp spines on the margin bractsF. broccoareola; D. The terraces of outer exterior bract; E. The distinctly longitudinal nerves of the outer middle bract.
The inner and outer laminar bracts vary in apex, margin, lamina and the colour. The apex of bracts are observed caudate or acuminate, rarely lobed. The lobed one could be
In Freycinetia the peduncle is a morphological structure that essentially still part of stem which in peduncle not only bracts attached, but also prophyll. Despite being the most basal part of inflorescence or infructescence, peduncle only indirectly supports the flowering or fruiting parts. The peduncle is usually straight and short. The members of theF. macrostachya group have the largest peduncles in common 2 cm in diam., while the smallest are commonly observed in the F. angustissima group, for about 0.5 cm in diam or less. But, the longest one more than 3 cm usually are found in the species of F. funicularisgroup.
Pedicel
The pedicel is a morphological structure that directly supports the spikes in inflorescence and cephalium in infructescence. It is the second most basal part of inflorescence or infructescence and immediately attached to peduncle. Usually pedicel is longer than peduncle. Most of species in F. macrostachya and F. funicularis group have robust pedicel. On the contrary, the members of the groups of species with F. oblanceolata andF. angustissima have slender pedicel. The pedicels are usually semi terete, but terete often seen. In some species the pedicels are observed as both terete and square-shaped, such as inF. ultrapedicelata.
The surface of pedicels can be covered by spines, nerves or indumentums. The covering can be dense, sparse or glabrous. Glabrous pedicels are rare. The surface of staminate and pistillate pedicels can also be different, both in the presence of coverings or the way the coverings are arranged.
Staminate Flower
A B C
1 c m 1 m m ½ m m
Fig. 7. The staminate flowers : A. The staminate flower of the F. marginata which has longest oblong of spadix; B. The oblong anther that longer than filament belong to F.simpliaxillata; C. The ovoid anther that is shorter than filament belong to F. megaauriculata.
B
A C
2 c m
1 m m
2 m m
one present in F. aculeata and also in F. funicularis, for about 2 cm. The shape of spadix can be oblong, cylindrical, globose or falcate. Globose spadix is very rare. Each species has its own shape, for example F. trachypoda has cylindrical spadix, while F. gunungmejensis has globose one. Filament is more or less uniform throughout the species. Anthers are rounded , oblong (Fig 7B) or ovoid (Fig. 7C).
The length of anther varies from equal to the five times longer of the filament. The androecium is monaldelphous or tetradynamous with distal appendage. Anthesis in Freycinetia takes 1 to 3 days. Prior to anthesis the laminar bract is transformed into fleshy structure and produces specific aroma to attract possible pollinators. Shortly afterward, bracts open. After anthesis bracts usually fall .
Pistillate Flower
Cephalia usually oblong (Fig. 8A) or cylindrical but it is globose in F. gunungmejensisand falcate inF. fusiforma These two species have few ovaries. On the other hand, F. megaauriculata, F. marginata, F. macrostachya, and F. flavicep have numerous. Some ovaries are unclose each other like what present in the F. milikenus (Fig. 8B) andF. fusiforma but so close in F. gibbsiae(Fig. 8C). The first three species are robust members of the F. macrostachya group, while F. flavicepis a member of the F. oblanceolatagroup. Cephalia is usually uniform before and after fruiting, except for F. aculeata. In this species cephalia is oblong before fruiting and turns to cylindrical after fruiting. Approximately the same situations are found in stigma and areola. As the consequence the structure of areola and number of stigma persist through maturity, thus considerated as good identification characters. On the contrary, the structure of style might change after fruiting as can be seen in F. oreophilaandF. broccoareola. In these two species the remains of styles are not observed in immature fruits but become obvious in mature fruits.
Stigma and Areola
number of stigma might vary within the species, for example F. aculeata has 1, 2 and 4 stigmas . Freycinetia funicularisis also known to have a variation in number of stigma that is 5 or 6. In this current study the variation is suggested to have a correlation with the fusion of berries, in which the fusion automatically changes the structure of stigma. This can be observed in F. imbristigma. This species usually has 6 stigmas but in some berries the number is increased to 12.
The shape of stigma might be linear, circular, semi terete, falcate, button-like, discoid, perforate or diffused. Linear stigma is found in F. cryptocarpa and F. yapenina (Fig. 9B). Circular stigma is in F. circulata. Semi terete and falcate stigmas are in F. aculeata . Discoid stigma is observed in F. decentata; however, in this species the button-like stigma can also be seen. Perforate stigma is found in F. oreophila, while diffused stigma inF. pauciberria.
Stigma is usually found on the terminal part of ovary but can be found olso on the transversal side. In this case the stigmatic remains on the transversal side too. In fused berries, as can be seen inF. funicularisthe areolas are united in the central of the ovary or berry, and as the consequence stigma moves to the margin side.
In few cases areola is lifted to the centre and stigma is moved into the longitudinal side of ovary or berry as can be observed in F. decipiens or into radial side as in F. millikenus.(Fig. 9C). In common, one stigma is surrounded by thin areola even in many numbers of stigmas,Freycinetiakeep their position , for example inF. salomonensis, but some species has different like inF. fusiforma where the stigmas moved to the marginal side change position with areola (Fig. 9D).
widely distributed marginal areola as can be seen in F. stenophylla. Some species, such as F. lenifolia have wide longitudinal areola that covers the style. In F. descendeta the areola even descent cunneately so it appears like corolla (Fig. 10C). Development of the areola from thin to the wide areola could be seen inF. frutospiralica.
The result of this current study shows that species that possess high number of stigmas also have wide variations in their stigmas and areolas. In contrast, the species with few numbers of stigmas have less variation. Furthermore, the arrangements of stigma and areola are not independent, but in an association. The stigma can be surrounded by areola or vice versa to form complex structures such as broccoli-like, imbricate, linear, or circular.
Broccoli-like stigma-areola structure is observed inF. megaauriculata (Fig. 9A). The imbricate structure is found in F. imbristigma. Linear structure with 2 lines is observed inF. solomonensis, while more than 2 lines can be seen in F. pleurantha. Circular is the most common structure found in West New Guinean species and it is found in F. fusiforma, F. gibsea, and F. sterophylla. The central structure, in which areola is encircling stigma is found in F. clavata, where as the central structure, in which the stigma is encircling the areola is observed inF. marantifoliaandF. inermis.
A
B
C D
¼ m m
¼ m m
½ m m ½ m m
A B C
¼ m m ¼ m m
¼ m m
Fig. 10.The variation of the areola: A. The commonly narrowest areola inF.trachypoda; B. The broccoli areola ofF.broccoareola; C. Widely descendent longitudinal areola ofF. desendeta.
Cephalia
Some of the important distinctive morphological characters for species level in Freycinetia are in their cephalia (singular, cephalium). These include the position relative to stems or shoots (terminal or axillary), numbers, shape, colours, and surface. The position of a cephalium might be on the terminal part of the stem or appears from the lateral side of the stem or shoots.
Species with terminal cephalia are found in the F. macrostachyaand F. angustissima groups. Axillary cephalia are found in the members of the F. funicularis group. In the F. oblanceolata group both terminal and axillary cephalia are found such as in F. gunungmejensis.
usually arrangement in umbel, while species that have more than 4 cephalia arrangement in spirally with exception in F. frutaspiralica, where the 4 cephalia are spirally arranged. The colour of mature cephalia varies from yellowish green, yellow, orange, crimson, bright red to blackish red.
Cephalium can be globose (rounded), oblong, cylindrical, and falcate. Generally cephalia are uniform in shape; however, few species have variety such as in F. frutonumerata, in which two shapes are present, falcate and cylindrical. The colour of immature cephalia is usually green but change through maturity. Mature cephalia might be yellow as inF. marantifolia, orange inF. iriana, and red in F. gibsae orF.aculeata. The surface of a cephalium can be flat or not flat. As in the case of colour, the surface of cephalia varies through maturity. All species have flat surface in their young cephalia . Some species (all members of the F. oblanceolata group) keep this flat surface through out maturity, some other only half flat. Freycinetia scandens and F. inermis are examples of species that keep the flat surface throughout maturity, while in F. manokwarianaandF. marantifoliaonly persist half part of their original flat surface .
Berries
Berry is the simple or basic structure of fruit in all species of Freycinetia and it is always fleshy. Unlike in Martellidendron, Pandanus, and Sararanga, a berry contains many fused ovules, thus a multiovulate fruit, while in the other three genera the simple fruit is always uniovulate. Assemblage of berries forms a cephalium, a complex structure of fruit. In this circumstance a cephalium can be regarded as a complex fruit and thus equals to cephalium found in the other three genera.
pyramidal berry. Thus, caution is essential when observing the shape of a berry.
Berry usually has more than one stigma. The increasing number of stigmas per berry is made through the fusion of berries. This fusion can be seen mainly in the F. funicularis group. The presence of perforation in the middle part of a berry is the evidence of such fusion. Thus, in this present study a robust berry with more than one stigma might have been the result of evolution.
Robust berries usually low in number and density per cephalium due to their size (Fig.11D). Low density of berries per cephalium is found mostly in the members of the F. funicularisgroup such as F. funicularis, F. imbristigma,andF. millikenus. UnlikeF. macrostahya group, in theF. angustissima group the relatively large sizes of berries are not caused by fusion of berries.
B
A C
D E
2 m m
½ m m 2 m m
2 m m
cephalium. Numerous and high density of berries are common in the members of the F. macrostahya group, for example F. flaviceps (Fig. 11E). Despite possess non fused berries members of this group have high density of berries. Apparently this is due to relatively large size of berry just like in the members of theF. angustissima group.
TheF. oblanceolata group is more diverse than the other groups. Species with low and high densities together with numerous and few numbers of berries are found in this group.
Freycinetia flaviceps and F. lagenicarpa are two examples of species with numerous berries, while F. biroi, F. folitenella, F. marantifolia, F. scandens, and F. tenuis are species with few numbers of berries .
The berry variation also includes the shapes of segments, wings (Fig.11C), and the number and size of segments. Freycinetia macrostachya is one example of species that possesses the non segmented berry. Freycinetia batantaensisandF. simpliaxilla are the example of species with winged-segmented berry. The species with numerous segmented berries can be seen inF. megaauriculata, while F. clavata has the few numbers of such berries. Freycinetia clavata has fairly large berries.
One berry normally has one pileus. In some species, such as in F. macrostachyaa berry has more than one pileus, in which it has two pilei Despite being a multiovulate fruit, one ovary is still indicated by one stigma; however, a berry with one stigma is extremely rare. So far it is only observed in F. angustissima, F. scandens and F. gunungmejensis.
Seed
A&B), falcate, oblong to linear with range of length 1 to 3 mm (Fig.12 C).
B
A C
1 m m ½ m m
The phylogenetic analysis carried out in this current study uses PAUP version. 4.1. Sixteen characters from 48 species and 17 out of 60 characters were selected with Pandanus tectorius andP. dubius were chosen as outgroups, change Sararanga as first out group. The 17 selected characters from the original 60 characters are character numbers 1, 2, 5, 6, 7, 8, 9, 23, 25, 28, 30, 31, 35, 39, 41, and 52 (Table 1.) with one addition character, which was not part of the 60 original characters, the upright laminar character.
Two species with mixed infrageneric grouping characters such as F. erythospatha (possessing characters of small organs, presence of lateral inforescences, absence of pseudo-rosette, absence of half leaf-like bracts) and F. charcatea (possessing character of slender habit and caducous auricle) were included to establish their position in the phylogeny.
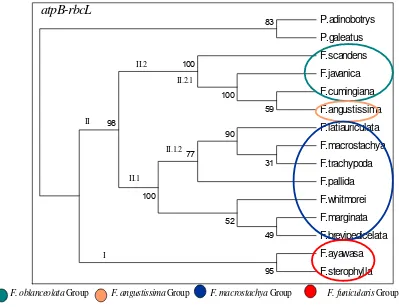
The result of this study shows that the West New Guinean species ofFreycinetiaare firmly divided into four infrageneric groups: The group of F. macrostachya, The group of F. funicularis,The group of F. oblanceolata and The group of F. angustissima (Fig. 14.). This infrageneric grouping are supported by the Consistency Index (CI) = 0.6977, Homoplasy Index (HI)= 0.3023, Retention Index (RI) = 0.857, and Rescaled Consistency Index (RC) = 0.5980.
Species inF. funicularis, F. oblanceolataand F. angustissima groups are arranged close each other and separated from the species inF. macrostachya group supported by 71 % value. The clade that includes the species ofF. macrostachya group is supported by 63 % value, the species ofF. funicularisgroup by 80 % value, while the species in F. angustissimagroup by 97 % value. It could be interpreted that the three groups are distinct one another and they are as a large group are distinct to the group of species with F. macrostachya group.
cephalia, and auricles.
Difference in the four infrageneric groups could be seen immediately by the habits, nature of the stems, and leaves attached to the stems. The F. macrostahya group is identified by the high climbing habit, rarely branched robust stem, and the presence of imbricate leaves throughout the stem, from basal to apical parts.
Species in F. funicularis group has semi imbricate leaves on the apical part of the stems, thus not throughout the stems. In the majority members of this group the stems on proximal half part are almost always naked. They also have more lateral branching, and the inflorescences often appear from the naked stems, thus possessing lateral inflorescences. The possession of lateral inflorescence is unique to this group.
Species inF. oblanceolataand F. angustissimagroup are often found as shrubs due to their slender and highly branched stems. Despite sharing these two characters, their leaves arrangements are different. The members of the group of species in F. angustissima have their leaves mostly on the apical part of the stem and the leaves are small, linear, and narrow; thus grass-like. Whereas the members of the group of species in F.oblanceolata have ellipsoidal, oblong, ensiform, oblanceolate, and other types of leaves. Furthermore, this group has their auricles easy to fall or disintegrated, thus make the leaves as having no leafsheaths.