i
BIOLOGI DAN STATISTIK DEMOGRAFI
Thrips parvispinus
KARNY
(THYSANOPTERA: THRIPIDAE) PADA
TANAMAN CABAI
RUDI TOMSON HUTASOIT
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
iii
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Biologi dan Statistik Demografi Thrips parvispinus Karny (Thysanoptera: Thripidae) pada Tanaman Cabai adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, September 2016
iv
RINGKASAN
RUDI TOMSON HUTASOIT. Biologi dan Statistik Demografi Thrips parvispinus Karny (Thysanoptera: Thripidae) pada Tanaman Cabai. Dibimbing oleh HERMANU TRIWIDODO dan RULY ANWAR.
Trips merupakan hama utama pada pertanaman cabai. Informasi mengenai luas serangan, kelimpahan, spesies, serta biologi dan statistik demografitrips pada pertanaman cabai masih terbatas. Penelitian ini bertujuan untuk mempelajari dan mendapatkan informasi mengenai luas serangan, kelimpahan, spesies trips yang menyerang pertanaman cabai, dan biologi serta statistik demografi Thrips parvispinus Karny sebagai spesies yang ditemukan dominan menyerang per-tanaman cabai di lapangan. Penelitian ini berlangsung dari bulan juni 2015 sampai Maret 2016.
Pengamatan trips dilakukan pada tanaman cabai besar dan rawit. Pengamatan dilakukan pada 14 petakan yang tersebar di empat kecamatan di Kabupaten Bogor yaitu, Dramaga, Cibungbulang, Tenjolaya, dan Cisarua. Trips dikumpulkan dari bagian bunga dan daun pada 10 tanaman sampel yang ditentukan secara acak pada setiap petakan pengamatan. Trips yang telah terkumpul diidentifikasi dan dihitung kelimpahannya.
Pengamatan biologi dan statistik demografi T. parvispinus dilakukan dengan memelihara 50 individu nimfa instar-1 dengan umur kohort pada helaian daun cabai dan diamati setiap hari untuk dicatat perkembangan dan keturunan yang diletakkan. Data yang didapat digunakan untuk memperoleh informasi biologi seperti stadia setiap instar, periode praoviposisi, periode oviposisi, siklus hidup, lama hidup imago, dan keperidian. Data tersebut kemudian digunakan untuk menyusun tabel neraca hayati untuk menghitung statistik demografi dengan menggunakan metode jackknife.
v
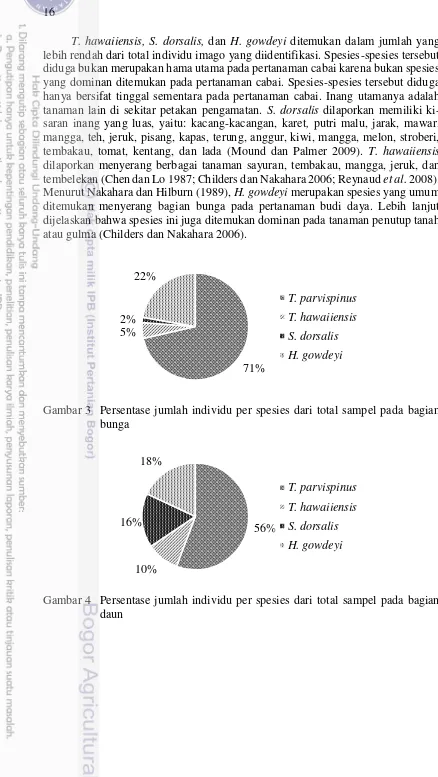
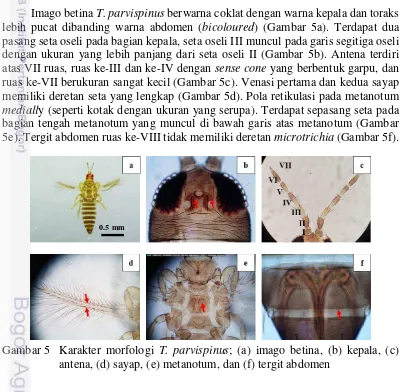
spesies yang paling dominan ditemukan menyerang bunga dan daun pada kedua jenis cabai sebesar 71 % dan 56 %.
Serangga pradewasa T. parvispinus terdiri atas lima fase, yaitu telur, nimfa instar-1, nimfa instar-2, prapupa dan pupa. Stadium telur berlangsung selama 4.79 hari, nimfa instar-1 selama 1.36 hari, nimfa instar-2 selama 3.54 hari, prapupa se-lama 1.08 hari, dan pupa sese-lama 1.96 hari. Siklus hidup berlangsung sese-lama 13.68 hari dengan stadium praoviposisi selama 1.11 hari. Lama hidup imago betina ber-langsung selama 8.55 hari, sedangkan jantan berber-langsung selama 6.00 hari. Rataan keperidian sebanyak 15.33 telur per imago betina sepanjang generasi. Tipe perkembangan populasi T. parvispinus termasuk dalam kurva sintasan tipe III. La-ju reproduksi kotor (GRR) T. parvispinus sebanyak 25.60 individu per generasi, laju reproduksi bersih (Ro) sebanyak 5.71 individu per induk per generasi, laju pertambahan intrinstik (r) sebesar 0.15 individu per induk per hari, rata-rata masa generasi (T) selama 11.49 hari dan waktu berlipat ganda selama 4.57 hari.
vi
SUMMARY
RUDI TOMSON HUTASOIT. Biology and Demographic Statistic of Thrips parvispinus Karny (Thysanoptera: Thripidae) on chili peppers. Supervised by HERMANU TRIWIDODO and RULY ANWAR.
Thrips is one of the major pests on chili. Information of the percentage of infested plants, abundance, species, and also the biology and demographic statistic has been still limited especially on chili peppers. This research was aimed to study about percentage of plants infested, abundance, species, biology and demographic statistic of Thrips parvispinus Karny was found as the dominant species attack on chili in the field. This research was conducted in June 2015-March 2016.
Observation of thrips was conducted on chili and cayenne in fourteen sites in four different locations in Bogor that is Dramaga, Cibungbulang, Tenjolaya, and Cisarua. Thrips were collected from leaves and flowers from 10 plant samples selected randomly. The thrips were identified and the number of thrips were cal-culated.
Studies of biology and demoghrapic statistic of T. parvispinus were conducted by rearing fifty first instar with the kohort on leaf of chili peppers in-dividually and observed daily to note the development and number of offsprings laid. The collected data were used to obtain information about the biology such as stadia of each instar, pre-oviposition period, oviposition period, life cycle, adult longevity, and fecundity. The data were also used to construct the life table for demoghrapic statistic calculation using jackknife method.
vii
The pre-adult phase of T. parvispinus consisted of egg, 1st instar nymph, 2nd instar nymph, pre-pupa, and pupa. The stadia of egg, 1st instar nymph, 2nd instar nymph, pre-pupa, and pupa was 4.79, 1.36, 3.54, 1.08, and 1.96 days respectively. The life cycle lasted for 13.68 days with the pre-oviposition periods for 1.11 days. Female longevity was 8.85 days, and male was 6.00 days. The mean of fecundity was 15.33 eggs/female. The population development type of T. parvispinus was type III. The gross reproductive rate (GRR) as much as 25.60 thrips per generation, net reproductive rate (Ro) was 5.71 offsprings per parent per generation, the intrinsic rate of increase (r) was 0.15 thrips per parents per day, the average period of generation (T) was 11.49 day and the doubling time was 4.57 days.
viii
© Hak Cipta Milik IPB, Tahun 2016 Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
ix Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Entomologi
BIOLOGI DAN STATISTIK DEMOGRAFI
Thrips parvispinus
KARNY
(THYSANOPTERA: THRIPIDAE) PADA
TANAMAN CABAI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
x
xii
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas segala karunia-Nya sehingga penulis dapat menyelesaikan penelitian dan penulisan tesis
dengan judul “Biologi dan Statistik Demografi Thrips parvispinus Karny (Thysanoptera: Thripidae) Pada Tanaman Cabai”. Penelitian ini dilaksanakan dari bulan Juni 2015 sampai Maret 2016 di beberapa kecamatan di Kabupaten Bogor, Laboratorium WiSH Batuhulung Bogor, dan Laboratorium Biosistematika Se-rangga Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor.
Penghargaan dan terima kasih sebesar-besarnya penulis sampaikan kepada Dr Ir Hermanu Triwidodo, MSc dan Dr Ir Ruly Anwar, MSi selaku komisi pem-bimbing. Dr Ir Nina Maryana, MSi selaku penguji luar komisi yang telah mem-berikan masukan serta perbaikan pada penelitian dan naskah tesis saya. Penulis juga menyampaikan terima kasih kepada Direktorat Jenderal Pendidikan Tinggi yang telah memberikan beasiswa kepada penulis untuk menyelesaikan studi melalui Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN).
Ungkapan terima kasih penulis sampaikan kepada Bapak Mangasa Hutasoit, Ibu Marintan Siburian, Purnama Pasaribu, dan seluruh keluarga atas doa, motivasi, dan dukungan yang selalu diberikan. Di samping itu, Penulis juga menyampaikan terima kasih kepada keluarga Entomologi-Fitopatologi 2013, Laboratorium WiSH, dan seluruh civitas akademika Departemen Proteksi Tanaman yang telah mem-berikan bantuan dalam penyelesaian tugas akhir ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, September 2016
xiii
Hama Penting Tanaman Cabai 5
Trips(Thysanoptera: Thripidae) 5
Morfologi Trips 5
Biologi Trips 6
Tanaman Inang dan Gejala Serangan 7
Trips pada Tanaman Cabai 7
Statistik Demografi 8
METODE 9
Waktu dan Tempat Penelitian 9
Metode Penelitian 9
Pengamatan Trips pada Tanaman Cabai Besar dan Cabai Rawit 9
Pengamatan Biologi T. parvispinus 11
Analisis Data 12
HASIL DAN PEMBAHASAN 13
Kondisi Umum Petakan Penelitian 13
Luas SerangandanKelimpahan Trips 13
Spesies Trips pada Tanaman Cabai Besar dan Rawit 15
Nisbah Kelamin 17
Karakter Morfologi 17
Biologi T. parvispinus 19
Sintasan dan Keperidian T. parvispinus 22
Statistik Demografi T. parvispinus 23
xvi
DAFTAR TABEL
1 Hama-hama penting tanaman cabai 5
2 Distribusi geografi spesies trips yang berasosiasi dengan tanaman cabai 8 3 Tanaman sekitar petakan pengamatan cabai besar 13 4 Tanaman sekitar petakan pengamatan cabai rawit 13 5 Luas serangan dan kelimpahan trips pada bagian bunga cabai besar dan
rawit 14
6 Luas serangan dan kelimpahan trips pada bagian daun cabai besar dan
rawit 14
7 Spesies dan jenis kelamin trips pada tanaman cabai besar dan rawit 15 8 Karakter biologi T. parvispinus pada tanaman cabai 21
9 Karakteristik demografi T. parvispinus 23
DAFTAR GAMBAR
1 Wadah koleksi trips 9
2 Kurungan pemeliharaan trips 11
3 Persentase jumlah individu per spesies dari total sampel pada bagian bunga 16 4 Persentase jumlah individu per spesies dari total sampel pada bagian daun 16
5 Karakter morfologi T. parvispinus 17
6 Karakter morfologi T. hawaiiensis 18
7 Karakter morfologi S. dorsalis 18
8 Karakter morfologi H. gowdeyi 19
9 Fase pradewasa T. parvispinus 20
10 Fase prapupa dan pupa T. parvispinus 20
11 Fase dewasa T. parvispinus 20
12 Kurva sintasan (lx) dan keperidian (mx) T. parvispinus 22
DAFTAR LAMPIRAN
1 Petakan pengamatan trips pada pertanaman cabai besar 31 2 Petakan pengamatan trips pada pertanaman cabai rawit 32 3 Data iklim (CH, HH, Suhu udara dan RH) Kabupaten Bogor
November 2015 33
4 Biologi T. parvispinus pada tanaman cabai 34
1
PENDAHULUAN
Latar Belakang
Cabai merupakan komoditas sayuran yang memiliki nilai ekonomi tinggi dan banyak dibudidayakan oleh petani di dataran rendah sampai dataran tinggi (Moekasan et al. 2014). Jenis cabai yang umum dibudidayakan di Indonesia, yaitu cabai besar (Capsicum annuum L.) dan cabai rawit (Capsicum frutescens L.). Buah cabai yang pedas, sangat populer di masyarakat sebagai penguat rasa makanan. Ekstrak bubuk cabai digunakan sebagai pengganti lada untuk membangkitkan selera makan dan penyedap masakan. Selain itu, cabai juga digunakan sebagai ramuan obat-obatan dalam industri farmasi, industri pewarna makanan, dan penghasil minyak atsiri (Cahyono 2003).
Luas panen produksi cabai besar di Indonesia mencapai 128.734 ha, dengan tingkat produksi sebesar 1.074.611 ton, dan produktivitas sebesar 8.35 ton/ha cabai basah pada tahun 2014 (BPS 2014). Cabai rawit memiliki luas panen produksi mencapai 134.882 ha, dengan tingkat produksi sebesar 800.484 ton, dan produktivitas sebesar 5.93 ton/ha (BPS 2014). Data tersebut menunjukkan masih terdapat kesenjangan antara produktivitas riil ditingkat usahatani dan produktivitas potensial cabai besar yang dapat mencapai 12-15 ton/ha (Duriat dan Sastrosiswojo 1994) dan cabai rawit dapat mencapai 12-20 ton/ha (Sujitno dan Dianawati 2015). Produktivitas produksi cabai yang rendah salah satunya disebabkan oleh adanya serangan organisme pengganggu tanaman (OPT). Saat ini ada 14 jenis hama penting yang dilaporkan menyerang tanaman cabai di lapangan, salah satunya adalah trips (Sumarni dan Muharam 2005). Kehilangan hasil akibat serangan trips pada pertanaman cabai besar dilaporkan mencapai 23% (Vos et al. 1991).
Trips merupakan serangga berukuran kecil dengan panjang tubuh 0.5-5 mm. Panjang tubuh beberapa spesies di daerah tropika dapat mencapai 14 mm (Borror et al. 1996; Antonelli 2003). Trips umumnya ditemukan pada bagian bunga dan daun tanaman (Mound dan Collins 2000). Peranan trips sebagai hama pada tanaman disebabkan oleh aktivitas makannya (meraut-menghisap). Gejala kerusakan yang ditimbulkan berupa bercak keperakan yang menjadi kecoklatan pada daun yang dapat mengganggu proses fotosintesis, sehingga daun mengeriting dan tunas terminal yang terserang menjadi kerdil (Kirk 2001).
Trips pada tanaman juga dapat berperan sebagai vektor virus (Rezende et al. 1997; Riley et al. 2011). Beberapa jenis virus yang ditularkan oleh serangga ini diantaranya Tomatto spotted wilt virus (TSWV), Lettuce spotted wilt virus (LSWF), Pineapple yellolv spotted virus (PYSV), Tip chlorosis, Kromneck diseases, dan Tobacco mosaic virus (TMV) (Klose et al. 1996; Sartiami 2008).
2
Phlaeothripidae), T. parvispinus, dan T. hawaiiensis (Terebrantia: Thripidae) ditemukan berasosiasi dengan tanaman cabai.
Keperidian yang tinggi dan siklus hidup yang singkat merupakan faktor penting yang menyebabkan terjadinya kolonisasi dan perkembangan populasi trips yang besar di lapangan. Imago betina mampu menghasilkan 30-300 telur tergan-tung spesies dan kualitas nutrisinya (Lewis 1973; Lewis 1997). Perkembangbiakan trips dapat mencapai 12-15 generasi setiap tahunnya pada daerah tropis atau di rumah kaca (Lewis 1973; Mound 2006).
Trips mampu menguasai suatu habitat dalam rentang waktu yang singkat secara optimal. Trips merupakan serangga oportunis dengan tipe seleksi r, waktu generasi pendek, toleransi terhadap kisaran inang yang luas, cenderung par-thenogenesis, dan struktur perkembangbiakan yang kompetitif, sehingga me-nyebabkan terjadinya agregasi (Funderburk 2001). Menurut Sastrosiswojo (1991), meskipun permasalahan trips di Indonesia dinilai cukup penting, namun tidak ba-nyak penelitian yang telah dilakukan, sehingga informasi mengenai serangga ini masih sangat terbatas.
Mengingat pentingnya peran serangga ini sebagai hama utama dan vektor penyebaran penyakit pada pertanaman cabai. Penelitian mengenai luas serangan, kelimpahan, inventarisasi spesies, serta biologi dan statistik demografi serangga hama ini pada pertanaman cabai perlu dilakukan. Statistik demografi merupakan salah satu langkah awal dalam mempelajari pertumbuhan populasi serangga. Informasi biologi dan demografi didapatkan dengan merancang neraca kehidupan (life table). Neraca kehidupan merupakan tabel data kesintasan dan fekunditas setiap individu dalam suatu populasi (Rockwood 2006). Neraca kehidupan dapat memberikan informasi secara terperinci mengenai kelahiran, perkembangan, re-produksi, dan kematian setiap individu dalam suatu populasi atau dengan kata lain memberi gambaran mengenai laju pertumbuhan suatu populasi.
Neraca kehidupan dapat menghasilkan ringkasan statistik sederhana termasuk harapan hidup individu. Selain itu, neraca kehidupan memiliki bentuk dasar yang dapat dimodifikasi untuk berbagai macam analisis data seperti mortalitas yang disebabkan oleh beragam faktor (Carey 2001). Informasi mengenai luas serangan, kelimpahan, spesies, dan biologi serta statistik demografi trips yang diperoleh, dapat menjadi bahan pertimbangan dalam pengambilan keputusan serta tindakan dalam upaya pengendalian serangga hama ini.
Perumusan Masalah
3 Tujuan Penelitian
Tujuan penelitian ini adalah:
1. Mendapatkan informasi mengenai luas serangan, kelimpahan, dan spesies trips yang menyerang pertanaman cabai besar dan cabai rawit di lapangan.
2. Mempelajari dan mendapatkan informasi mengenai biologi dan statistik demografi T.parvispinus sebagai spesies yang ditemukan paling dominan me-nyerang pertanaman cabai besar dan cabai rawit.
Manfaat Penelitian
4
TINJAUAN PUSTAKA
Tanaman Cabai (Capsicum spp.)
Cabai (Capsicum spp.) termasuk ke dalam famili Solanaceae. Terdapat seki-tar 20 spesies yang termasuk dalam genus Capsicum. Secara umum cabai dike-lompokkan dalam dua jenis, yaitu cabai besar atau cabai merah (C. annuum) dan cabai kecil atau cabai rawit (C. frutescens). Cabai rawit berukuran lebih kecil dibanding cabai besar dan memiliki rasa yang lebih pedas (Setiadi 1996).
Tanaman cabai berasal dari Amerika Tropik, ditemukan oleh Columbus dan disebarkan ke Amerika Tengah menuju Amerika Serikat Bagian Selatan. Penye-baran terus meluas hingga ke daerah tropis dan subtropis. C. annuum, C. frutescens, Capsicum baccatum, Capsicum pubescens, dan Capsicum chinense merupakan spesies yang sudah dikenal oleh masyarakat secara umum, namun informasi ketiga spesies terakhir masih kurang (Setiadi 1996).
Cabai besar merupakan tanaman perdu, berdiri tegak, bertajuk lebar, dan ber-cabang banyak. Tinggi tanaman mencapai 30 cm-2.5 m. Cabai besar memiliki daun berbentuk lonjong dengan panjang mencapai 8-12 cm dan lebar 3-5 cm. Bunga cabai berbentuk terompet kecil, umumnya berwarna putih, tetapi ada juga yang berwarna ungu (Setiadi 1996). Bunga cabai termasuk berkelamin dua, karena pada satu bunga terdapat kepala sari dan kepala putik. Tangkai putik dan mahkota ber-warna putih dengan 5-6 helai cuping. Kepala putik berber-warna kuning kehijauan, sedangkan kepala sari berwarna ungu (Samadi 1997).
Cabai rawit memiliki ukuran buah yang kecil dan pendek serta memiliki rasa yang lebih pedas di antara cabai lainnya, sehingga sering disebut cabai kecil. Tinggi tanaman cabai rawit dapat mencapai 1.5 m. Batangnya berbuku-buku dan bersudut. Daun berbentuk bulat telur, bagian ujung meruncing, pangkal menyempit, dan berwarna hijau. Panjang daun mencapai 1.5-10 cm dan lebar 0.5-5 cm. Bunga ber-ukuran kecil dengan mahkota berbentuk bintang, berwarna putih kekuningan, kadang berwarna kehijauan atau ungu. Bunga keluar dari ketiak daun dalam jumlah tunggal atau berkelompok sebanyak 2-3 bunga. Bunga terdiri atas 5 sampai 6 helai cuping. Putik berwarna kuning kehijauan dan kadang berwarna ungu. Kepala putik berwarna kehijauan dan kepala sari berwarna hijau kebiruan. Buah berbentuk bulat telur atau jorong dengan bagian ujung menyempit. Buah memiliki ukuran yang be-ragam, dengan panjang dapat mencapai 7.5 cm dan diameter 1-3 cm. Buah muda berwarna hijau tua, putih kehijauan atau putih, dan buah yang masak berwarna merah terang (Setiadi 1994; Tindall 1983).
5 Hama Penting Tanaman Cabai
Sampai saat ini ada 14 jenis hama penting yang dilaporkan menyerang tanaman cabai di lapangan. Kehilangan hasil karena serangan OPT tersebut dapat mencapai 20-100%. Jenis hama penting pada setiap fase pertumbuhan tanaman cabai disajikan pada Tabel 1.
Tabel 1 Hama-hama penting tanaman cabai
Fase pertumbuhan Hama
Vegetatif Trips (T. parvispinus)
Kutu daun persik (Myzus persicae)
Tungau teh kuning(Polyphagotarsonemus latus) Ulat tanah (Agrotis ipsilon)
Gangsir (Brachytrypes portentus) Uret (Phyllophaga spp.)
Ulat bawang (Spodoptera exigua) Ulat grayak (Spodoptera litura) Kutu kebul (Bemisia tabaci) Wereng kapas (Empoasca lybica)
Lalat pengorok daun (Lyriomiza huidobrensis) Anjing tanah (Gryllotalpa africanal)
Generatif Trips (T. parvispinus) Ulat bawang (S. exigua) Ulat grayak (S. litura)
Kutu daun persik(M. persicae) Tungau teh kuning (P. latus) Kutu kebul (B. tabaci) Wereng kapas (E. lybica)
Lalat pengorok daun (L. huidobrensis) Ulat buah tomat (Helicoverpa armigera)
Lalat buah (Bactrocera dorsalis)
Sumber: Setiawati et al. (2005)
Trips(Thysanoptera: Thripidae)
Morfologi Trips
6
membentuk pola segitiga (triangle) (Lewis 1973; Borror et al. 1996). Struktur mulut trips disebut probosis yang berbentuk seperti sebuah kerucut dan terletak di bagian belakang permukaan bawah kepala (Antonelli 2003). Trips memiliki alat mulut yang asimetris (mandibel kanan tereduksi) dengan tipe meraut-menghisap. Stilet pada alat mulut berfungsi untuk meraut jaringan tanaman dan untuk menu-suk serta menghisap cairan pada sel tanaman (Borror et al. 1996; Antonelli 2003). Menurut Lewis (1973), struktur yang khas pada bagian toraks adalah sayap. Jumlah sayap dua pasang dengan bentuk memanjang, berukuran sempit, dan mempunyai beberapa rangka sayap serta rambut-rambut yang berumbai. Subordo Terebrantia memiliki struktur sayap yang sejajar satu sama lain, sedangkan pada Subordo Tubulifera posisi sayap tumpang tindih, sehingga hanya satu pasang saja yang terlihat. Imago jantan maupun betina bersayap atau tidak bersayap. Beberapa spesies hanya imago betina saja yang memiliki sayap (Mound 2006).
Subordo Terebrantia memiliki abdomen dengan bagian ujung yang menge-rucut dan memiliki ovipositor pada ruas VIII dan IX, sedangkan pada Subordo Tubulifera, ujung abdomen berbentuk seperti tabung tanpa ovipositor. Organ yang berbentuk seperti tabung ini disebut genital opening organ, yang terletak antara ruas IX dan X abdomen (Lewis 1973; Borror et al. 1996). Imago jantan biasanya memiliki ukuran tubuh yang lebih kecil dan warna yang lebih pucat dibanding imago betina. Abdominal sternit jantan mempunyai satu atau lebih area glandular di daerah tengah pada beberapa spesies. Terdapat perbedaan terminal abdomen antara imago jantan dan betina. Betina mempunyai sebuah ovipositor yang terdiri atas dua pasang katup seperti gergaji, sedangkan pada imago jantan terlihat seperti aedeagus (Mound 2006).
Biologi Trips
Siklus hidup trips terdiri atas telur, dua instar nimfa yang aktif, prapupa, dan pupa (Mound dan Kibby 1998; Pourian et al. 2009). Trips dapat menghasilkan beberapa generasi pertahun dengan tipe perkembangan peralihan antara metamor-fosis bertahap (paurometabola) dan metamormetamor-fosis sempurna (holometabola) (Borror et al. 2005). Trips menyelesaikan siklus hidupnya sekitar 2-3 minggu (Ananthakrishnan 1993). Imago betina Subordo Terebrantia meletakkan telur se-cara tunggal di dalam jaringan tanaman dengan bantuan ovipositor, sedangkan imago betina Subordo Tubulifera meletakkan telur pada permukaan substrat de-ngan genital opening organ (Mound 2006). Telur berbentuk seperti ginjal berwar-na putih pucat. Jumlah telur yang dihasilkan 30-60 telur tergantung pada nutrisi, suhu, dan kelembaban (Ananthakrishnan 1993).
Nimfa instar-1 berwarna putih pucat atau transparan dengan mata berwarna merah, berukuran sekitar 0.5 mm. Nimfa instar-1 aktif bergerak dan memakan ja-ringan tanaman. Fase nimfa instar-1 berlangsung selama 2-3 hari. Nimfa instar-2 berwarna kuning tua keruh, berukuran sekitar 0.8 mm. Fase nimfa instar-2 ber-langsung selama 3-4 hari (Lewis 1973; Pourian et al. 2009).
ba-7 gian posterior dengan ukuran tubuh lebih kecil serta warna lebih pucat dibanding imago betina (Dibiyantoro 1998). Imago paling banyak ditemukan pada bagian dalam bunga dan daun. Lama hidup imago dapat mencapai 30 hari (Fekrat et al. 2009).
Trips berkembang biak secara seksual dan aseksual (parthenogenesis). Re-produksi secara partenogenesis terbagi menjadi tiga tipe yang berbeda, yaitu: arrhenotoky, thelytoky, dan deutherotoky. Arrhenotoky terjadi apabila telur imago betina yang tidak dibuahi menghasilkan keturunan yang semuanya jantan haploid. Thelytoky terjadi apabila telur imago betina yang tidak dibuahi menghasilkan keturunan yang semuanya betina diploid, sedangkan deutherotoky terjadi apabila telur imago betina yang tidak dibuahi menghasilkan keturunan jantan dan betina (Lewis 1973; Nault et al. 2006).
Tanaman Inang dan Gejala Serangan
Trips merupakan serangga hama yang bersifat polifag dengan kisaran inang yang luas. Trips merupakan hama penting pada tanaman sayuran dan hortikultura (Dibiyantoro 1998). Inang utama trips selain cabai adalah tembakau, kopi, ubi jalar, Vigna, Crotalaria, dan kacang-kacangan. Trips menyerang tanaman cabai sepanjang tahun. Trips menyerang tanaman sejak tanaman ada di persemaian. Ha-ma ini meraut daun, tunas dan buah serta mengisap cairan tanaHa-man dengan meng-gunakan alat mulutnya. Serangan hebat umumnya terjadi pada musim kemarau (Vos et al. 1991). Tanaman yang terserang menjadi kering dan menimbulkan warna keperakan, daun mengeriting, dan tunas terminal yang terserang menjadi kerdil (Kiers et al. 2000). Bunga dan daun yang terserang menimbulkan gejala berupa bintik-bintik putih atau bercak berwarna merah keperak-perakan dan daun mengeriting atau berkerut (Mound dan Kibby 1998).
Trips pada Tanaman Cabai
Beberapa spesies trips telah dilaporkan menyerang pertanaman cabai di ber-bagai negara (Tabel 2). Scirtothrips dorsalis, Thrips palmi, dan Thrips tabaci me-rupakan spesies yang sebagian besar ditemukan di Asia (Talekar 1991). Spesies yang pertama kali dilaporkan menyerang pertanaman cabai di Pulau Jawa adalah T. tabaci, namun berdasarkan hasil identifikasi dari survei Vierbergen ditemukan be-berapa spesies yang menyerang pertanaman cabai di Pulau Jawa, yaitu: T. parvispinus. T. hawaiiensis, T. florum, dan T. orientalis (Vos et al. 1991).
8
Tabel 2 Distribusi geografi spesies trips yang berasosiasi dengan tanaman cabai
No Spesies Lokasi
1 Frankliniella cephalica Amerika Serikat
2 Frankliniella intosa Taiwan
3 Frankliniella occidentalis Nederland
4 Gynaikothrips Karny India
5 Haplothrips chinensis Taiwan
6 Hercinothrips femoralis Amerika Serikat
7 S. dorsalis Burma, India, Sri Lanka, Taiwan,
Thailand
8 Taeniothrips simplex Amerika Serikat
9 T. hawaiiensis Taiwan
10 T. palmi Filipina, Jepang, Taiwan
11 T. tabaci Denmark, Libanon, Nederland
India, Indonesia, Jepang, Amerika Serikat
Sumber: Talekar (1991)
Statistik Demografi
Langkah awal dalam mempelajari perkembangan suatu populasi serangga adalah dengan mengetahui aspek-aspek demografinya. Demografi adalah analisis kuantitatif karakteristik suatu populasi, terutama hubungannya dengan pola per-tumbuhan populasi, hubungan ketahanan, dan pergerakan populasi. Aspek demo-grafi suatu populasi terdapat dalam neraca kehidupan (life table). Menurut Price (1997), neraca kehidupan adalah ringkasan pernyataan tentang kehidupan individu-individu dalam populasi/kelompok. Tarumingkeng (1992) menambahkan bahwa neraca kehidupan dapat digunakan untuk mengkalkulasikan berbagai statistik po-pulasi yang dapat memberikan informasi mengenai kelahiran (natalitas), kematian (mortalitas), dan peluang untuk berkembang biak, sehingga dapat digunakan se-bagai parameter perilaku perkembangan populasi.
9
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan Juni 2015-Maret 2016. Penelitian meli-puti pengamatan di lapangan yang dilakukan di empat kecamatan di Kabupaten Bogor dan pengamatan biologi serta statistik demografi T. parvispinus yang dila-kukan di Laboratorium WisH Batuhulung Bogor. Identifikasi diladila-kukan di Labo-ratorium Biosistematika Serangga, Departemen Proteksi Tanaman, Fakultas Per-tanian, Institut Pertanian Bogor, Jawa Barat.
Metode Penelitian
Pengamatan Trips pada Tanaman Cabai Besar dan Cabai Rawit
Kegiatan ini dilaksanakan pada bulan November 2015-Maret 2016. Peng-amatan trips pada cabai besar dan cabai rawit dilakukan di 14 petakan yang ter-sebar di empat kecamatan di Kabupaten Bogor. Setiap jenis cabai masing-masing terdiri atas tujuh petakan pengamatan (Lampiran 1 & Lampiran 2). Sebaran petakan pengamatan cabai besar yaitu: dua petakan di Kecamatan Cibungbulang, dua petakan di Kecamatan Tenjolaya, dan tiga petakan di Kecamatan Cisarua. Sebaran petakan pengamatan cabai rawit yaitu: dua petakan di Kecamatan Dramaga, tiga petakan di Kecamatan Tenjolaya, dan satu petakan di Kecamatan Cibungbulang. Pada setiap petakan, dipilih 10 tanaman sampel yang ditentukan secara acak.
Pengambilan sampel. Trips diamati pada bagian bunga dan daun untuk kedua jenis cabai. Pengamatan pada bunga dilakukan dengan cara menepuk-nepuk bunga diatas wadah plastik (diameter = 2.5 cm dan tinggi = 4.5 cm) yang telah diisi air (Gambar 1a). Setiap tanaman sampel diamati sebanyak 10 bunga. Pengamatan pada daun dilakukan dengan cara menepuk-nepuk ranting daun (10-15 helai daun) di atas nampan plastik (32 cm x 25 cm) yang diisi air (Gambar 1b). Setiap tanaman sampel diamati satu ranting daun. Trips kemudian dikumpulkan ke dalam wadah koleksi. Setiap wadah koleksi diberi label yang berisi informasi tanggal, lokasi, dan bagian tanaman inang yang diamati. Pengambilan sampel dilakukan hanya satu kali dari pukul 10.00 sampai 15.00 WIB. Trips yang telah dikumpulkan ke dalam wadah koleksi kemudian dibawa ke laboratorium untuk diidentifikasi dan dihitung ke-limpahannya.
10
Luas serangan. Luas serangan trips diamati pada tanaman yang sama de-ngan pengamatan kelimpahan trips. Besaran luas serade-ngan dihitung dede-ngan rumus:
L =
N x 100 % L = luas serangan trips
N = jumlah tanaman yang terserang N = jumlah tanaman contoh yang diamati
Kelimpahan trips. Sampel yang dikoleksi dari lapangan kemudian dihitung kelimpahannya berdasarkan jenis dan bagian tanaman cabai yang diamati. Imago dan nimfa trips yang terkumpul dipisahkan dalam penghitungan populasi. Proses penghitungan populasi dilakukan di bawah mikroskop stereo.
Identifikasi. Trips yang telah dikoleksi dari lapangan, kemudian identifikasi sampai tahap spesies. Sebanyak 31 dan 28 individu imago yang di-koleksi dari bagian bunga dan daun pada cabai besar serta 32 dan 33 individu imago yang dikoleksi dari bagian bunga dan daun pada cabai rawit diambil secara acak untuk diidentifikasi. Identifikasi trips dilakukan dengan cara membuat preparat mikroskop sementara.
Pembuatan preparat slide mengacu pada metode Mound dan Kibby (1998) dengan langkah kerja sebagai berikut: imago trips yang dikoleksi dimasukkan ke dalam cawan kaca berisi alkohol 70% untuk dipisahkan dari kotoran yang terbawa. Spesimen ditempatkan pada kaca penutup yang berdiameter 13 mm, dengan bagian ventral tubuh berada di atas, kemudian kedua sayap direntangkan serta posisi antena diluruskan dengan menggunakan jarum halus hingga posisinya tidak bertumpuk dan terlihat jelas. Setelah posisi spesimen tertata dengan baik, larutan Hoyer diteteskan pada kaca penutup, lalu segera ditutup dengan kaca obyek. Setelah kaca obyek diletakkan, posisi preparat slide segera dibalik, sehingga kaca penutup berada di atas kaca obyek. Preparat slide kemudian dikeringkan selama satu minggu pada suhu 35-45 ºC di kotak pengering, kemudian diberi cat kuku berwarna bening pada setiap pinggiran kaca penutup dan dikeringkan kembali selama satu hari. Identifikasi trips dilakukan berdasarkan kunci identifikasi dari Mound dan Kibby (1998), Sartiami dan Mound (2013), dan Subagyo (2014). Jumlah individu per spesies dari keseluruhan sampel yang diidentifikasi disajikan dalam bentuk tabel dan diagram lingkaran, dengan rumus:
Jumlah individu per spesies (%) = ∑ sp s s x
∑ s sp s s x 100
11 Pengamatan Biologi T. parvispinus
Penelitian ini berlangsung pada bulan Januari-Maret 2016. Kegiatan ini di-lakukan di Laboratorium WiSH Batuhulung Bogor. Kegiatan ini meliputi pe-ngamatan 50 individu nimfa instar-1 dengan umur kohort hingga individu terakhir mati dan identifikasi spesies yang diamati.
Persiapan tanaman inang. Tanaman cabai besar yang digunakan untuk perbanyakan dan pemeliharaan trips berumur 30-60 HST (hari setelah tanam). Ta-naman dipelihara dalam kurungan yang terbuat dari kain kassa organdi. Peme-liharaan tanaman cabai pada kurungan bertujuan untuk menghindari serangan dan infestasi telur dari serangga lain.
Pembuatan kurungan serangga. Kurungan serangga terbuat dari gelas plastik (diameter = 6 cm dan tinggi = 8 cm) (Gambar 2). Permukaan bagian atas di-buat lubang ventilasi dilapisi kain kassa organdi. Bagian bawah kurungan dilapisi tisu yang telah dibasahi.
Gambar 2 Kurungan pemeliharaan trips
Perbanyakan serangga uji.T. parvispinus yang digunakan sebagai popu-lasi awal dikumpulkan dari pertanaman cabai di Desa Cibatok II Kecamatan Ci-bungbulang Kabupaten Bogor. Sebanyak 30 individu imago betina dan jantan hasil koleksi dari lapangan dipelihara pada daun cabai umur 30 HST dalam kurungan gelas plastik. Setiap kurungan diinfestasikan satu individu imago betina dan jantan. Imago dikeluarkan dan dipindahkan pada daun dan kurungan baru setelah 24 jam. Daun yang telah terinfestasi telur, diamati hingga muncul nimfa instar-1. Perbanyakan terus dilakukan hingga jumlah serangga yang dibutuhkan cukup untuk pengujian.
Identifikasi serangga uji. Beberapa individu serangga uji yang digunakan sebagai populasi awal diidentifikasi untuk memastikan spesies. Proses identifikasi diawali dengan membuat preparat sementara. Pembuatan preparat slide mengacu pada Mound dan Kibby (1998).
12
Pengamatan lama hidup imago jantan dan betina dilakukan secara terpisah. Lama hidup imago jantan diamati dengan menginfestasikan imago pada daun cabai dalam kurungan serangga. Setiap satu helai daun dalam kurungan diinfestasikan satu individu imago jantan. Pengamatan dilakukan setiap hari hingga imago jantan terakhir mati. Pengamatan stadium praoviposisi, stadium oviposisi, siklus hidup, fekunditas, dan lama hidup imago betina dilakukan dengan menginfestasikan imago betina pada daun cabai dalam kurungan serangga. Setiap satu helai daun dalam kurungan diinfestasikan satu individu imago betina. Setiap hari, imago betina di-pindahkan pada daun cabai yang baru di dalam kurungan baru hingga imago betina terakhir mati. Daun cabai yang telah diinfestasikan imago betina pada hari se-belumnya, kemudian diamati setiap hari hingga muncul nimfa instar-1. Mound dan Masumoto (2005) melaporkan inkubasi telur dapat berlangsung selama 3-7 hari. Berdasarkan hal tersebut pengamatan dilakukan setiap hari selama tujuh hari. Pengamatan dilakukan dengan menggunakan mikroskop portable dinolite AM 2111 Basic dan mikroskop stereo.
Analisis Data
Data pengamatan luas serangan dan kelimpahan trips diperiksa melalui uji-t dengan bantuan program SPSS 16. Data hasil pengamatan trips dengan umur kohort selama satu generasi disusun dalam bentuk neraca kehidupan. Neraca kehidupan dengan umur kohort merupakan neraca kehidupan yang mengikuti perkembangan serangga dengan umur kohort dimulai dari kemunculan individu pertama sampai kematian individu terakhir yang bertahan hidup (Begon et al. 2006). Data-data yang dibutuhkan yaitu: x adalah kelas umur (hari), lx adalah peluang hidup setiap individu pada umur x, mx adalah fekunditas per individu pada umur x, dan lxmx adalah banyaknya keturunan yang dilahirkan pada kelas umur x. Penghitungan di-lanjutkan dengan menggunakan metode Jackknife. Metode Jackknife digunakan sebagai pendekatan umum untuk melakukan uji hipotesis dan menghitung selang kepercayaan. Adapun parameter yang diamati yaitu:
1. Laju reproduksi kotor (GRR) = Σ mx 2. Laju reproduksi bersih (Ro) = Σ lxmx
3. Laju pertambahan intrinsik (r) = ln (Ro) / T
4. Rataan masa generasi (T) = Σ xlxmx / Σ lxmx
13
HASIL DAN PEMBAHASAN
Kondisi Umum Petakan Penelitian
Petakan pengamatan yang diamati merupakan pertanaman cabai dengan sis-tem budi daya konvensional. Petakan pertanaman pada kedua jenis cabai terdiri atas pola tanam monokultur dan polikultur. Luas masing-masing petakan pengamatan sekitar 250-1000 m2 (Lampiran 1 & Lampiran 2). Tanaman cabai yang diamati merupakan tanaman cabai pada fase generatif. Tanaman di sekitar pertanaman cabai dicatat pada setiap petakan pengamatan (Tabel 3 & Tabel 4).
Tabel 3 Tanaman sekitar petakan pengamatan cabai besar
Petakan pengamatan Tanaman sekitar
1 Desa Cibatok I, Cibungbulang Jagung, pisang
2 Desa Cibatok II, Cibungbulang Kacang panjang, terung, singkong 3 Desa Cibitung Tengah, Tenjolaya Singkong, paria, ubi jalar
4 Desa Cibitung Tengah, Tenjolaya Terung, pisang, sawi
5 Desa Cibereum, Cisarua Pisang, bayam, singkong
6 Desa Cibereum, Cisarua Pisang, padi, kelapa, tomat
7 Desa Cibereum, Cisarua Mangga, kacang panjang, pisang
Tabel 4 Tanaman sekitar petakan pengamatan cabai rawit
Petakan Pengamatan Tanaman sekitar
1 Desa Dramaga, Dramaga Pepaya, sawi, bayam, pisang
2 Desa Dramaga, Dramaga Jambu biji, pisang
3 Desa Ciherang, Dramaga Pepaya, pisang, petai
4 Desa Cibitung Tengah, Tenjolaya Terung, pisang, sawi 5 Desa Cibitung Tengah, Tenjolaya Singkong, kelapa, pisang 6 Desa Cibitung Tengah, Tenjolaya Pepaya, singkong
7 Desa Cibatok II, Cibungbulang Belimbing, jati
Luas Serangan dan Kelimpahan Trips
Luas serangan imago, nimfa, dan total trips tidak berbeda nyata antar kedua
je-14
nis cabai yang diuji. Hal ini sesuai dengan laporan Vos et al. (1991) yang me-nyatakan bahwa trips merupakan hama utama pada pertanaman cabai (C.annuum L. dan C. frutescens L.) di Indonesia.
Populasi imago lebih tinggi dibandingkan nimfa pada bagian bunga dan daun untuk kedua jenis cabai (Tabel 5 & Tabel 6). Hal ini dipengaruhi lama fase nimfa yang lebih singkat daripada lama hidup imago (Table 8), sehingga menyebabkan populasi imago tumpang tindih. Faktor abiotik seperti suhu dan kelembaban pada saat pengamatan juga memberikan pengaruh. Berger et al. (2008) menyatakan bahwa, pada suhu yang tinggi imago akan mengalihkan energi yang dimilikinya untuk memproduksi telur yang lebih banyak, sementara pada suhu rendah imago akan menimbun energi yang dimilikinya untuk membentuk struktur tubuh.
Kelimpahan imago, nimfa dan total trips yang ditemukan pada bagian bunga dan daun untuk kedua jenis cabai relatif rendah (Tabel 5 & Tabel 6). Yulianti (2008) melaporkan bahwa populasi trips dapat mencapai 19.22 individu/tanaman dengan pengamatan yang dilakukan pada bagian daun muda, daun tua, dan bunga. Selain faktor biotik, populasi trips di pertanaman dapat dipengaruhi oleh kondisi abiotik terutama curah hujan, suhu, dan kelembaban. Pengambilan sampel pada penelitian ini dilakukan pada bulan November dengan kondisi curah hujan dan kelembaban yang tinggi (Lampiran 3). Lewis (1973) melaporkan bahwa, curah hujan dapat me-mengaruhi populasi trips, karena dapat membunuh nimfa dan menekan penyebaran trips. Hujan merupakan faktor lingkungan yang dapat berpengaruh langsung dan tidak langsung pada trips. Hujan yang lebat dapat menyebabkan trips jatuh dari tanaman. Selain itu, hujan dapat merangsang laju pertumbuhan daun baru yang mengurangi kepadatan trips per daun dan meningkatkan proporsi daun sehat (Kirk 1997).
Tabel 5 Luas serangan dan kelimpahan trips pada bagian bunga cabai besar dan rawit
Fase
Kelimpahan trips Luas serangan (%)
(individu/ranting daun)a
Tabel 6 Luas serangan dan kelimpahan trips pada bagian daun cabai besar dan rawit
Fase
Kelimpahan trips Luas serangan (%)
15 Vos et al. (1991) menyatakan bahwa, trips menjadi hama utama pada per-tanaman cabai terutama pada musim kemarau. Menurut Kakkar et al. (2010), suhu yang panas serta kelembaban yang tinggi menjadi faktor pendukung utama meledaknya populasi trips di suatu wilayah, sehingga pada musim hujan jumlah populasi trips sering menurun meskipun kelembaban cukup tinggi dibutuhkan untuk perkembangan trips secara sempurna. Lebih lanjut dijelaskan bahwa, pada suhu yang tinggi imago akan mengalihkan energi yang dimilikinya untuk memproduksi telur yang lebih banyak, sementara pada suhu rendah imago akan menimbun energi yang dimilikinya untuk membentuk struktur tubuh (Berger et al. 2008).
Spesies Trips pada Tanaman Cabai Besar dan Rawit
Berdasarkan hasil identifikasi, empat spesies trips ditemukan menyerang bagian bunga maupun daun pada pertanaman cabai besar dan cabai rawit. Keempat spesies tersebut adalah T. parvispinus, T. hawaiiensis, S. dorsalis, dan H. gowdeyi (Tabel 7). T. parvispinus, T. hawaiiensis, S. dorsalis termasuk dalam Subordo Terebrantia Famili Thripidae, sedangkan H. gowdeyi termasuk dalam Subordo Tubulifera familli Phlaeothripidae. Keempat spesies trips yang ditemukan me-rupakan serangga fitofag dengan kisaran tanaman inang yang luas (Chen dan Lo 1987; Ananthakrishnan 1993; Childers dan Nakahara 2006). Hasil ini sesuai dengan penelitian Yulianti (2008) dan Subagyo (2014) yang melaporkan bahwa, spesies T. parvispinus,T.hawaiiensis (Terebrantia: Thripidae), dan H. gowdeyi (Tubulifera: Phlaeothripidae) menyerang pertanaman cabai di Jawa Barat.
Thrips parvispinus merupakan spesies yang ditemukan paling dominan menyerang bagian bunga dan daun pada kedua jenis cabai (Gambar 3 & Gambar 4). Hasil ini sesuai dengan laporan Vos et al. (1991) yang menyatakan bahwa, T. parvispinus merupakan spesies yang dominan ditemukan menyerang pertanaman cabai (Capsicum spp.) di Indonesia. Yulianti (2008) juga melaporkan bahwa, spesies T. parvispinus ditemukan dominan menyerang pertanaman cabai di wilayah Jawa Barat sekitarnya, dengan rata-rata populasi 8.18-19.22 trips/tanaman. Sartiami dan Mound (2013) lebih lanjut menjelaskan bahwa, T. parvispinus adalah spesies yang paling banyak ditemukan berasosiasi dengan tanaman hortikultura di Pulau Jawa, dengan kerusakan yang cukup parah pada tanaman cabai dan kentang. Menurut Mound dan Collins (2000), T. parvispinus merupakan hama yang bersifat polifag, namun spesies ini lebih tertarik pada bunga yang berwarna putih dan wangi. Tabel 7 Spesies dan jenis kelamin trips pada tanaman cabai besar dan rawit
Jenis
T. parvispinus T. hawaiiensis S. dorsalis H. gowdeyi Jantan Betina Jantan Betina Jantan Betina Jantan Betina
Besar Bunga 31 5 14 1 1 0 1 0 9
Daun 28 3 15 0 2 0 5 0 3
Rawit Bunga 32 8 18 0 1 0 0 0 5
Daun 33 3 13 1 3 0 5 0 8
16
T. hawaiiensis, S. dorsalis, dan H. gowdeyi ditemukan dalam jumlah yang lebih rendah dari total individu imago yang diidentifikasi. Spesies-spesies tersebut diduga bukan merupakan hama utama pada pertanaman cabai karena bukan spesies yang dominan ditemukan pada pertanaman cabai. Spesies-spesies tersebut diduga hanya bersifat tinggal sementara pada pertanaman cabai. Inang utamanya adalah tanaman lain di sekitar petakan pengamatan. S. dorsalis dilaporkan memiliki ki-saran inang yang luas, yaitu: kacang-kacangan, karet, putri malu, jarak, mawar, mangga, teh, jeruk, pisang, kapas, terung, anggur, kiwi, mangga, melon, stroberi, tembakau, tomat, kentang, dan lada (Mound dan Palmer 2009). T. hawaiiensis dilaporkan menyerang berbagai tanaman sayuran, tembakau, mangga, jeruk, dan tembelekan(Chen dan Lo 1987; Childers dan Nakahara 2006; Reynaud et al. 2008). Menurut Nakahara dan Hilburn (1989), H. gowdeyi merupakan spesies yang umum ditemukan menyerang bagian bunga pada pertanaman budi daya. Lebih lanjut dijelaskan bahwa spesies ini juga ditemukan dominan pada tanaman penutup tanah atau gulma (Childers dan Nakahara 2006).
Gambar 3 Persentase jumlah individu per spesies dari total sampel pada bagian bunga
17 Nisbah Kelamin
Imago betina lebih dominan dibandingkan imago jantan dari total sampel yang diidentifikasi. Perbandingan nisbah kelamin antara jantan dan betina pada T. parvispinus dan T. hawaiiensis yaitu, 1:3.1 dan 1:3.5. Rasio ini dipengaruhi oleh beberapa faktor seperti, tipe reproduksi, nutrisi, suhu, dan kelembaban. Menurut Graham dan Knight (1967), kualitas pakan yang cukup baik, daya reproduksi betina yang tinggi, dan spesies partenogenesis cenderung menghasilkan betina yang lebih tinggi. Moritz (1997) menyatakan bahwa, suhu dan kelembaban dapat memberikan pengaruh terhadap frekuensi jumlah jantan dalam suatu populasi. Yadav dan Chang (2014) lebih lanjut menjelaskan bahwa, proporsi individu betina trips lebih tinggi pada kisaran suhu 22 ºC dan 31 ºC dibanding pada kisaran suhu 16, 19, 25, dan 28 ºC. Lama hidup imago betina yang lebih panjang dibanding imago jantan (Tabel 8) juga memberikan pengaruh, sehingga imago betina lebih dominan ditemukan di lapangan. Spesies S. dorsalis dan H. gowdeyi tidak dapat ditentukan nisbah kelaminnya, karena semua individu dalam spesies yang diidentifikasi hanya betina.
Karakter Morfologi
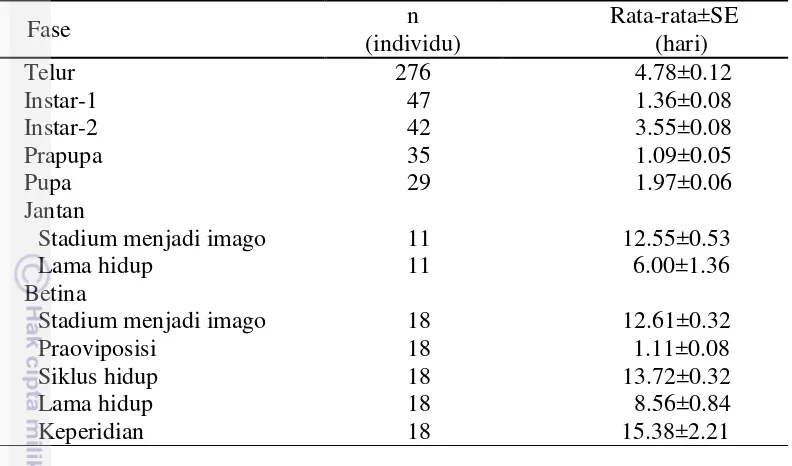
Imago betina T. parvispinus berwarna coklat dengan warna kepala dan toraks lebih pucat dibanding warna abdomen (bicoloured) (Gambar 5a). Terdapat dua pasang seta oseli pada bagian kepala, seta oseli III muncul pada garis segitiga oseli dengan ukuran yang lebih panjang dari seta oseli II (Gambar 5b). Antena terdiri atas VII ruas, ruas ke-III dan ke-IV dengan sense cone yang berbentuk garpu, dan ruas ke-VII berukuran sangat kecil (Gambar 5c). Venasi pertama dan kedua sayap memiliki deretan seta yang lengkap (Gambar 5d). Pola retikulasi pada metanotum medially (seperti kotak dengan ukuran yang serupa). Terdapat sepasang seta pada bagian tengah metanotum yang muncul di bawah garis atas metanotum (Gambar 5e). Tergit abdomen ruas ke-VIII tidak memiliki deretan microtrichia (Gambar 5f).
Gambar 5 Karakter morfologi T. parvispinus; (a) imago betina, (b) kepala, (c) antena, (d) sayap, (e) metanotum, dan (f) tergit abdomen
18
Gambar 6 Karakter morfologi T. hawaiiensis; (a) imago betina, (b) kepala, (c) antena, (d) sayap, (e) metanotum, dan (f) tergit abdomen
Gambar 7 Karakter morfologi S. dorsalis;(a) imago betina, (b) kepala, (c) antena, (d) sayap, (e) metanotum, dan (f) tergit abdomen
Imago T. hawaiiensis memiliki warna tubuh coklat atau bicoloured (Gambar 6a). Terdapat sepasang seta oseli pada bagian kepala, seta oseli III muncul di luar segitiga oseli (Gambar 6b). Antena terdiri atas VII ruas (Gambar 6c). Barisan seta pada venasi pertama sayap depan tidak lengkap (Gambar 6d). Pola retikulasi pada bagian atas metanotum berbentuk transversal, sedangkan di bagian tengah lo-ngitudinal paralel. Terdapat sepasang seta pada bagian tengah metanotum yang muncul dekat dengan garis atas metanotum (Gambar 6e). Tergit abdomen ruas ke-VIII memiliki deretan microtrichia yang lengkap, pendek, tetapi tidak teratur (Gambar 6f).
Imago S. dorsalis dicirikan dengan tubuh berwarna kuning, terdapat tanda coklat pada bagian tengah tergit abdomen ruas III-VII (Gambar 7a). Terdapat tiga pasang seta oseli pada bagian kepala, seta oseli III muncul di dalam segitiga oseli (Gambar 7b). Antena terdiri atas VIII ruas (Gambar 7c). Barisan seta pada venasi sayap depan tidak lengkap pada baris pertama maupun kedua (Gambar 7d). Me-tanotum dengan sepasang seta di bawah garis atas meMe-tanotum (Gambar 7e). Tergit abdomen ruas ke-VIII dengan deretan microtrichia yang lengkap, teratur, dan pan-jang (Gambar 7f).
0.5 mm
19
Gambar 8 Karakter morfologi H. gowdeyi;(a) imago betina, (b) antena, (c) sayap, dan (d) pronotum
Imago H. gowdeyi dicirikan dengan warna tubuh yang dominan berwarna coklat gelap. Ujung abdomen berbentuk tabung (Gambar 8a). Antena terdiri atas VIII segmen, segmen ke-III dan ke-IV berwarna kuning, pada dasar segmen ke-V dan ke-VI setengahnya berwarna kuning dan setengahnya berwarna coklat muda. Segmen ke-III dengan dua sense cone sedangkan segmen ke-IV dengan empat sense cone (Gambar 8b). Permukaan sayap bagian tengah terdapat delapan silia duplikat (Gambar 8c). Pronotum dengan lima pasang seta kapitat mayor (Gambar 8d).
Biologi T. parvispinus
T. parvispinus memiliki tipe metamorfosis peralihan antara paurometabola dan holometabola. Serangga ini tergolong dalam tipe peralihan karena mengalami lebih dari satu instar pradewasa yang memiliki kerangka sayap, perkembangan sayap terjadi pada bagian dalam tubuh, dan terdapat fase pupa sebelum menjadi imago (Borror et al. 2005). Tahap perkembangan T. parvispinus terdiri atas telur, nimfa, prapupa, pupa dan imago (Tabel 8 & Lampiran 4). Telur diletakkan secara tunggal di dalam jaringan epidermis daun. Telur berbentuk seperti ginjal, berwarna putih pucat, dan berwarna semakin keruh ketika akan menetas (Gambar 9a). Fase nimfa mengalami dua kali pergantian kulit. Nimfa instar-1 berwarna putih tran-sparan (Gambar 9b). Nimfa instar-2 berwarna kuning tua gelap yang semakin lama warnanya semakin kecoklatan (Gambar 9c). Pergantian fase nimfa ditandai dengan adanya eksuvia.
20
Gambar 9 Fase pradewasa T. parvispinus; (a) telur, (b) instar-1, dan (c) instar-2 Fase prapupa dan pupa ditandai dengan adanya kerangka sayap. Prapupa me-miliki kerangka sayap lebih pendek (sebatas toraks) dan berwarna putih transparan (Gambar 10a). Pupa memiliki kerangka sayap lebih panjang hingga abdomen. Kerangka sayap dan tungkai berwarna lebih gelap. Antena tertekuk ke belakang se-panjang kepala (Gambar 10b).
Gambar 10 Fase prapupa dan pupa T. parvispinus; (a) prapupa dan (b) pupa Imago betina dan jantan dapat dibedakan berdasarkan ukuran dan warna. Tubuh imago T. parvispinus betina berwarna coklat dengan warna kepala dan to-raks lebih pucat dibandingkan abdomen (Gambar 11a). Jantan mempunyai ukuran tubuh lebih kecil, segmen terminal abdomen lebih tumpul, dan keseluruhan tu-buhnya berwarna kuning (Gambar 11b).
Gambar 11 Fase dewasa T. parvispinus; (a) betina, dan (b) jantan
0.5 mm 0.5 mm
0.5 mm
0.5 mm 0.5 mm
21 Tabel 8 Karakter biologi T. parvispinus pada tanaman cabai
Fase n
Stadium menjadi imago 011 12.55±0.53
Lama hidup 011 6.00±1.36
Betina
Stadium menjadi imago 018 12.61±0.32
Praoviposisi 018 1.11±0.08
Siklus hidup 018 13.72±0.32
Lama hidup 018 8.56±0.84
Keperidian 018 15.38±2.21
Fase pradewasa T. parvispinus berlangsung singkat (Tabel 8). Stadium pra-dewasa yang singkat secara tidak langsung menyebabkan siklus hidup imago T. parvispinus menjadi lebih pendek. Menurut Amarasekare et al. (2008), perbedaan siklus hidup serangga pada beberapa tanaman inang dapat disebabkan oleh faktor nutrisi, senyawa alelokimiawi, dan struktur fisik daun tanaman inang. Spesies ta-naman yang berbeda menyediakan kualitas nutrisi dan unsur pokok kimia yang berbeda, hal ini dapat memengaruhi karakteristik suatu populasi terutama terhadap perkembangan, tingkat reproduksi, dan ketahanan serangga tersebut. Zhang et al. (2007) melaporkan bahwa, masa perkembangan pradewasa F. occidentalis ber-langsung selama 9.22, 10.19, 10.42, 12.15 dan 12.91 hari berturut-turut pada tanaman mentimun, kubis, kacang, cabai, dan tomat.
Speight et al. (2008)lebih lanjut menjelaskan bahwa, pertumbuhan dan per-kembangan populasi serangga ditentukan oleh faktor serangga itu sendiri (faktor internal) dan faktor luar (faktor eksternal). Salah satu faktor eksternal yaitu, suhu dan kelembaban yang sangat berkaitan terhadap iklim mikro perkembangan po-pulasi serangga. Yadav dan Chang (2014) melaporkan bahwa, pada kisaran suhu 16, 19, 22, 25, 28, dan 31 ºC masa perkembangan pradewasa T. palmi berlangsung selama 35.66, 26.83, 19.28, 14.13, 12.20, dan 9.57 hari.
Stadium praoviposisi merupakan selang waktu ketika imago betina pertama kali muncul hingga meletakkan telur. Stadium praoviposisi T. parvispinus ber-langsung singkat, yaitu selama 1.1 hari (Tabel 8). Masa praoviposisi yang singkat akan mempercepat proses peletakkan telur yang pertama. Hasil penelitian Zhang et al. (2007)melaporkan bahwa, stadium praoviposisi F. occidentalis berlangsung se-lama 1.67 hari pada tanaman cabai dengan suhu ±27ºC. Salmasi et al. (2003) melaporkan, pada suhu ±27 ºC stadium praoviposisi T. tabaci berlangsung selama 3.60 hari pada tanaman bawang.
22
nutrisi yang dimiliki tanaman inang berpengaruh terhadap biologi serangga, sehingga serangga kekurangan nutrisi yang dibutuhkan untuk perkembangannya (Speight et al. 2008). Lama hidup imago jantan F. occidentalis dilaporkan ber-langsung selama empat hari, sedangkan imago betina berber-langsung selama 8.23 hari pada tanaman cabai dengan kisaran suhu ±27 ºC pada saat pemeliharaan (Zhang et al. 2007). Salmasi et al. (2003) melaporkan bahwa, lama hidup imago betina T. tabaci berlangsung selama 16.15 hari pada tanaman bawang dengan kisaran suhu ±27 ºC pada saat pemelliharaan.
Nisbah kelamin merupakan jumlah relatif jantan dan betina dalam suatu po-pulasi. Semakin banyak betina yang dihasilkan, maka semakin cepat populasi serangga tersebut berkembang. Rasio jenis kelamin jantan dan betina T. parvispinus yaitu 1:1.63. Menurut Graham dan Knight (1967), kualitas pakan yang cukup banyak, daya reproduksi betina yang tinggi, serta spesies partenogenesis akan cen-derung menghasilkan betina yang lebih tinggi.
Sintasan dan Keperidian T. parvispinus
Peluang hidup T. parvispinus digambarkan dalam kurva sintasan (lx), se-dangkan natalitas digambarkan dalam kurva keperidian (mx).Sintasan atau peluang hidup T. parvispinus diperoleh dari pengamatan harian mulai dari fase nimfa instar-1 hingga menjadi dewasa. Kurva sintasan T. parvispinus menunjukkan bahwa, pe-luang hidup mulai menurun sejak individu berumur 2 hari setelah infestasi (Gambar 12 & Lampiran 5). Kurva laju kesintasan dari hasil pengamatan menunjukkan tingkat kematian yang lebih banyak terjadi pada fase pradewasa atau dapat disebut kurva kesintasan tipe III.
Menurut Price (1997), terdapat tiga jenis kurva bertahan hidup. Kurva tipe I, menggambarkan kematian organisme dalam jumlah sedikit ketika populasi ber-umur muda dan kematian dalam jumlah besar sewaktu populasi berber-umur lebih tua. Tipe II menunjukkan laju kematian konstan dan tipe III memperlihatkan kematian yang lebih besar terjadi pada populasi berumur muda.
23 Keperidian merupakan banyaknya keturunan yang mampu dihasilkan oleh setiap imago betina sepanjang generasi. Sifat biofisik dan biokimia tanaman menentukan tingkat pertumbuhan populasi. Kualitas dan kuantitas makanan akan menentukan taraf perkembangan serangga. Hal ini dikarenakan pertumbuhan, mor-talitas, dan keperidian serangga dipengaruhi oleh kandungan nutrisi makanan disamping faktor lingkungan (Speight et al. 2008). Zhang et al. (2007) melaporkan bahwa, keperidian F. occidentalis pada tanaman cabai sebanyak 7.67 telur per im-ago betina sepanjang generasi pada kisaran suhu ±27 ºC. Keperidian T. tabaci pada tanaman bawang dilaporkan sebanyak 31.63 telur per imago betina pada kisaran suhu ±27 ºC (Salmasi et al. 2003).
Kurva rataan keperidian (mx), menunjukkan bahwa peletakan telur dimulai dari hari ke-9 sampai hari ke-24 setelah infestasi.Imago betina trips meletakkan te-lur dalam jumlah sedikit pada awal fase imago dan jumlah terus bertambah seiring pertambahan umur imago dan kembali mengalamai fluktuasi hingga menjelang kematian imago. Rata-rata keperidian betina per hari mencapai titik maksimum pa-da hari ke-11 dengan rataan 4.0 telur (Gambar 12 & Lampiran 5).
Statistik Demografi T. parvispinus
Statistik demografi T. parvispinus meliputi laju reproduksi kotor (GRR), la-ju reproduksi bersih (Ro), lala-ju pertambahan instrinsik (r), rata-rata waktu satu ge-nerasi (T), dan waktu berlipat ganda (DT) (Tabel 9). Berdasarkan hasil peng-amatan, seluruh imago betina T. parvispinus sepanjang generasi mampu mengha-silkan keturunan sebanyak 25.60 individu per generasi. Nilai reproduksi bersih (Ro) T. parvispinus menunjukkan bahwa rata-rata banyaknya keturunan yang di-hasilkan oleh imago betina adalah 5.71 betina per induk per generasi atau dapat dikatakan bahwa populasi T. parvispinus dapat berlipat ganda sebanyak 5.71 kali dalam setiap generasinya. Tinggi rendahnya Ro akan memengaruhi populasi. Nilai Ro dipengaruhi oleh jumlah keturunan yang dihasilkan per imago betina dan pe-luang hidup imago betina.
Nilair menggambarkan laju pertambahan intrinsik pada lingkungan konstan dan keadaan sumber daya yang tidak terbatas. Laju pertambahan intrinsik di-tentukan oleh berbagai aspek yang berhubungan dengan siklus kehidupan or-ganisme tersebut, yaitu kematian, kelahiran, dan waktu perkembangan. Nilai r se-besar 0.15 menunjukkan bahwa, individu dapat bertambah sebanyak 0.15 kali dalam sumberdaya yang tidak terbatas atau percepatan pertambahannya sebesar 15% individu per hari. Tinggi rendahnya nilai r dapat dipengaruhi oleh keperidi-an, mortalitas pradewasa, dan masa dewasa.
Tabel 9 Karakteristik demografi T. parvispinus
Karakter Rata-rata ± SE
Laju reproduksi kotor (GRR) 25.60±0.14 individu/generasi Laju reproduksi bersih (Ro) 05.71±0.02 individu/induk/generasi Laju pertambahan intrinsik (r) 00.15±0.00 individu/induk/hari Rataan lama generasi (T) 11.49±0.01 hari
24
25
SIMPULAN DAN SARAN
Simpulan
1. Luas serangan dan kelimpahan imago, nimfa, dan total trips pada bagian bunga dan daun tidak berbeda nyata antar kedua jenis cabai.
2. Spesies trips yang ditemukan menyerang pertanaman cabai besar dan cabai rawit adalah T. parvispinus, T. hawaiiensis, S. dorsalis (Terebrantia: Thri-pidae)dan H. gowdeyi (Tubulifera: Phlaeothripidae).
3. Tipe metamorfosis T.parvispinus adalah tipe peralihan antara paurometabola dan holometabola. Tahap perkembangan T. parvispinus terdiri atas telur, nimfa, prapupa, pupa dan imago. Tipe perkembangan populasi T.parvispinus termasuk dalam kurva sintasan tipe III. Laju reproduksi kotor T. parvispinus (GRR) sebanyak 25.60 individu per generasi, laju reproduksi bersih (Ro) sebanyak 5.71 individu per induk per generasi, laju pertambahan intrinstik (r) sebesar 0.15 individu per induk per hari, rata-rata masa generasi (T) selama 11.49 hari, dan waktu berlipat ganda selama 4.57 hari.
Saran
26
DAFTAR PUSTAKA
Amarasekare KG, Mannion CM, Osborne LS. 2008. Life history of Paracoccus marginatus (Hemiptera: Pseudococcidae) on four host plant spesies under laboratory condition. J Environ Entomol. 37(3):630-635.
Ananthakrishnan TN. 1993. Bionomics of thrips. Annu Rev Entomol. 38:71-92. Antonelli LA. 2003. Thrips [internet]. Washington (US): Washington State
University (WSU) Puyallup; [diunduh pada 2015 Juni 20]. Tersedia pada: www.puyallup.wsu.edu/plantclinic/resources/pdf/pls36thrips.pdf.
Begon M, Townsend CR, Harper JL. 2006. Ecology: From Individuals to Eco-systems. Ed ke-2. Oxford (GB): Blackwell Publishing.
Berger D, Walters R, Gotthard K. 2008. What limits insect fecundity? body size and temperature dependent egg maturation and oviposition in a butterfly. Funct Ecol. 22:523-529.
[BPS] Badan Pusat Statistik. 2014. Produksi cabai besar, cabai rawit, dan bawang merah tahun 2014. Berita resmi satistik. No. 71/08/ Th. XVIII Agustus 2015 [internet]. [diunduh pada 2014 November 24]; Tersedia pada http://www. bps.go.id/tab_sub/view.php?kat=3&tabel=1&daftar=1&id_subyek=55%20 ¬ab=66.
Borror DJ, Triplehorn CA, Johnson NF. 1996. Pengenalan Pelajaran Serangga. Ed ke-6. Partosoedjono S, penerjemah. Yogyakarta(ID): Gajah Mada University Pr. Terjemahan dari: An Introduction to the Study of Insects.
Borror DJ, Triplehorn CA, Johnson NF. 2005. An Introduction to The Studies of In-sects. Ed ke-7. United States of America (US): Brooks/Cole.
Cahyono B. 2003. Cabai Rawit Teknik Budidaya dan Analisis Usaha Tani. Yogyakarta (ID). Kanisius.
Carey JR. 1993. Applied Demography for Biologist With Special Emphasis on In-sect. New York (US): Oxford University Pr.
Carey JR. 2001. Insect biodemography. Annu Rev Entomol. 46:79-110.
Chen JS, Lo PKC 1987. Diffrential preference of the flower dwelling thrips, Thrips hawaiiensis (Morgan) (Thysanoptera: Thripidae) to some gladiolus cultivars. Agric Res China 36 (3):371-326.
Childers CC, Nakahara S. 2006. Thysanoptera (thrips) within citrus orchards in Florida: Species distribution, relative, and seasonal abundance within trees, and species on vines and ground cover plants. J Insect Sci. 6:1-19.
Dibiyantoro ALH. 1994. Management of Thrips tabaci Lind with special reference on garlic (A. sativum L.) PhD. 3rd-Year Report. 1994. Univ.Newcastle (GB). 23-25.
Dibiyantoro ALH. 1998. Thrips pada Tanaman Sayuran. Bandung (ID): Balai Pe-nelitian Tanaman Sayuran.
Duriat AS. Sastrosiswojo S. 1994. Pengendalian Hama Penyakit Terpadu pada Ag-ribisnis Cabai. Jakarta (ID): Penebar Swadaya.
27 Funderburk J. 2001. Ecology of thrips. Di dalam: Marullo R, Mound L, eitor. Thrips and Tospoviruses. Proceedings of the 7th International Symposium on Thysanoptera; 2001 Juli 2-7; Reggio Calabria, Itali. Canberra (AU): CSIRO Entomology. hlm 121-128.
Graham SA, Knight FB. 1967. Principles of Forest Entomology. New York (US): McGrawHill Book Company.
Kakkar G, Seal DR, Jha VK. 2010. Common blossom trips, Frankliniella schultzei Trybom (Insecta: Thysanoptera: Thripidae). IFAS Extension. Florida (US): University of Florida.
Kiers E, Kogel WJ, Balkema BA, Mollema C. 2000. Flower visitation and ovipo-sition behaviour of Frankliniella occidentalis (Thysanoptera: Thripidae) on cucumber plants. J Applied Entomol. 124:27-32.
Kirk WDJ. 1997. Feeding. Di dalam: Lewis T, (ed.). Thrips and Crop Pests. Cambridge (GB): CABI. Pp. 119-174.
Kirk WDJ. 2001. The pest and vector from the west: Frankliniella ocidentalis. Di dalam: Rita M, Laurence M. (ed.). Thrips and Tospoviruses. Proceedings of the 7th International Symposium on Thysanoptera; 2001 Juli 2-7; Reggio Calabria, Itali. Canberra (AU): CSIRO Entomology. hlm 33-42.
Klose MJ, Sdoodee R, Teakle DS, Milne JR, Greber RS, Walter GH. 1996. Transmission of three strains of tobacco streak ilarvirus by different thrips species using virus infected pollen. J Phytopathol. 144:281-284.
Lewis T. 1997. Pest thrips in perspective. Di dalam: Lewis T, editor. Thrips and Crop Pest. Cambridge (GB): CABI. Pp.1-13.
Lewis T. 1973. Thrips: Their Biology, Ecology, and Economic Importance. London (GB): Academic Pr.
Moekasan TK, Prabaningrum L, Adiyoga W, Putter H. 2014. Panduan Praktis Budidaya Cabai Merah Berdasarkan Konsepsi PHT. Jakarta (ID): PT. Penebar Swadaya.
Mound LA. 2006. Taxonomy of the insect order Thysanoptera. Thrips. Taxonomy Workshop No.1; 2006 Juli 3-7; Malaysia. Kuala Lumpur (MY): Institute of Biological Science, University Malaya Kuala Lumpur.
Mound LA. 2008. Thysanoptera (Thrips) of the world-a checklist [internet]. Canberra (AU): CSIRO Entomology; [diunduh pada 2016 Juni 19]. Tersedia pada: http://www.ento.csiro.au/thysanoptera/worldthrips.html.
Mound LA, Collins DW. 2000. A Southeast Asian pest species newly recorded from Europe: Thrips parvispinus (Thysanoptera: Thripidae), its confused identity and potential quarantine significance. J Entomol. 97:197-200. Mound LA, Kibby G. 1998. Thysanoptera: An Identification Guide. Ed ke-2.
Canberra (AU): CSIRO Entomology.
Mound LA, Masumoto M. 2005. The genus thrips (Thysanoptera, Thripidae) in Australia, New Caledonia and New Zealand. Zootaxa 1020:1-64.
Mound La, Palmer JM. 2009. Identification, distribution and host plants of the pest species of Scirtothrips (Thysanoptera: Thripidae). Bull. Ent. Res. 71 (3):467-479.doi:10.1017/S0007485300008488.
Moritz G. 1997. Structure, growth and development. Di dalam: Lewis T, editor. Thrips and Crop Pest. Cambridge (GB): CABI . Pp. 15-63.
28
Nault BA, Shelton AM, Kaufmann GJL, Clark ME, Werren JL, La-Rosa CJC, Kennedy GC. 2006. Reproductive modes in onion thrips (Thysanoptera: Thripidae) populations from New York onion fields. Environ Entomol. 35(5): 1264-1271.
Prabaningrum L, Moekasan TK. 2007. Identifikasi status hama pada budidaya pa-prika (Capsicum annuum Var. Grossum) terhadap serangan Thrips parvispinus Karny (Thysanopthera: Thripidae) di Kabupaten Bandung, Jawa Barat. J Hort 17(2):161-167.
Price PW. 1997. Insect Ecology. Ed ke-3. New York (US): John Wiley & Sons. Pourian HR, Mirab M, Alizadeh M, Orosz S. 2009. Study on biology of onion
thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae) on cucumber (Var. Sulltan) in laboratory conditions. J Plant Protection Research. (49) 4:390-394.
Reynaud P, Balmes V, Pizzol J. 2008. Thrips hawaiiensis (Morgan, 1913) (Thysa-noptera: Thripidae), an Asian pest thrips now established in Europe. Bulletin OEPP/EPPO Bulletin. 38:155-160.
Rezende JAM, Galleti SR, Pozzer L, Resende RO, de Avila AC, Scagliusi SMM. 1997. Incidence, biological, and serological characteristicts of a tospovirus in experimental fields of zucchini in Sao Paulo state, Brazil. Fitopatol. 22:92-95.
Riley DG, Joseph SV, Srinivasan R, Diffie S. 2011. Thrips vectors of tospovirus. J Integrated Pest Management. 1(2):1-10.doi: 10.1603/IPM10020.
Rockwood LL. 2006. Introduction to Population Ecology. Malden (US): Blackwell Publishing.
Salmasi H, Hejazi M, Rahnemon AA. 2003. Life cycle of onion thrips, Thrips tabaci Lind. in insectarium. J Agric Sci 13:91-100.
Samadi B. 1997. Budidaya Cabai Merah Secara Komersial. Yogyakarta (ID): Ya-yasan Pustaka Nusantara.
Sartiami D. 2008. Kunci identifikasi ordo Thysanoptera pada tanaman pangan dan hortikultura. J ilmu Pert Indones. 13(2):103-110.
Sartiami D, Mound LA. 2013. Identification of the terebrantian thrips (Insecta, Thysanoptera) associated with cultivated plants in Java, Indonesia. ZooKeys 306:1-21.doi:10.3897/zookeys.306.5455.
Sastrosiswojo S. 1991. Thrips on vegetables in Indonesia. Di dalam: Talekar NS, editor. Thrips in Southeast Asia. Proc. Regional Consultation Workshop; 1991 March 13; Bangkok, Thailand. Taiwan (TW): AVRDC. hlm 12-17. Setiadi. 1994. Jenis dan Budi Daya Cabai Rawit. Jakarta (ID): Penebar Swadaya. Setiadi. 1996. Bertanam Cabai. Ed ke-7. Jakarta (ID): Penebar Swadaya.
Setiawati W, Udiarto BK, Muharam A. 2005. Pengenalan dan Pengendalian Hama-hama Penting pada Tanaman Cabai Merah. Bandung (ID): Balitsa, Balitbang Pertanian.
Speight MR, Hunter MD, Watt AD. 2008. Ecology of Insect: Concept and Application. Ed ke-2. Oxford (GB): John Wiley & Sons.
Subagyo VNO. 2014. Identifikasi thrips (Insecta: Thysanoptera) yang berasosiasi dengan tanaman hortikultura di Bogor, Cianjur, dan Lembang [tesis]. Bogor (ID): Institut Pertanian Bogor.
Ba-29 rat. Pros Sem Nas Masy Biodiv Indon. 1(4):874-877.doi: 10.13057/ psnmbi/m010438.
Sumarni N, Muharam A. 2005. Budidaya Tanaman Cabai Merah. Bandung (ID): Balai Penelitian Tanaman Sayuran.
Talekar NS. 1991. Di dalam: Talekar NS, editor. Talekar NS, editor. Thrips in Southeast Asia. Proc. Regional Consultation Workshop; 1991 March 13; Bangkok, Thailand. Taiwan (TW): AVRDC. hlm 61-65.
Tindall HD. 1983. Vegetables in the Tropics. Hongkong (HK): English Language Book Society (ELBS).
Tarumingkeng RC. 1992. Dinamika Pertumbuhan Populasi Serangga.Bogor (ID): IPB Pr.
Vos JGM, Sastrowiswojo S, Uhan TS, Setiawati W. 1991. Thrips on hot peppers in Java, Indonesia. Di dalam: Talekar NS, editor. Thrips in SoutheastAsia. Proc. Regional Consultation Workshop; 1991 March 13; Bangkok, Thailand. Taiwan (TW): AVRDC. Pp. 18-28.
Yadav R, Chang NT. 2014. Effects of temperature on the development and population growth of the melon thrips, Thrips palmi, on eggplant, Solanum melongena.J Insect Science. 14 (78):1-9.
Yulianti P. 2008. Spesies Trips (Ordo Thysanoptera) pada tanaman cabai dan ta-naman sekitarnya di Jawa Barat [skripsi]. Bogor (ID): Institut Pertanian Bo-gor.
30