SURAT PERYATAAN TESIS DAN SUMBER INFORMASI
Dengan ini, saya menyatakan bahwa tesis berjudul Interaksi Orangutan (Pongo pygmaeus wurmbii) Betina Dengan Individu Lain dan Hubungannya Dengan Profil Metabolit Glukokortikoid Di Stasiun Penelitian Orangutan Tuanan, Kalimantan Tengah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum pernah diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Semua data dan informasi yang digunakan berasal atau dikutib dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini sayamelimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2013
Neneng Mardianah
i
RINGKASAN
Neneng Mardianah. Interaksi Orangutan (Pongo pygmaeus wurmbii) Betina dengan Individu Lain dan Hubungannya dengan Profil Metabolit Glukokortikoid Di Stasiun Penelitian Orangutan Tuanan, Kalimantan Tengah. Dibimbing oleh Prof. Dr. drh Iman Supriatna, Dr. drh Muhammad Agil M. Sc, Agr dan Dr. Sri Suci Utami Atmoko.
Orangutan Kalimantan (Pongo pygmaeus wurmbii) merupakan salah satu primata endemik yang berstatus endangered menurut IUCN dan berstatus Apendiks I menurut CITES. Penurunan populasi disebabkan perusakan dan penyusutan habitat serta penangkapan illegal. Penelitian orangutan telah banyak dilakukan terutama di hutan primer, namun masih sedikit mengenai perilaku sosial pada orangutan yang berada di hutan sekunder. Stasiun Penelitian Orangutan Tuanan (SPOT) habitat orangutan yang termasuk dalam kategori hutan sekunder dan mengalami degradasi yang tinggi sehingga mudah sekali terbakar pada musim kemarau. Orangutan termasuk hewan frugivore dan sangat peka terhadap perubahan kondisi habitat. Perusakan habitat secara besar-besaran mempengaruhi perubahan perilaku sosial orangutan. Berkurannya daerah jelajah membuka peluang perjumpaan antar individu semakin besar, sedangkan orangutan memiliki sistem sosial semi soliter. Perilaku sosial pada orangutan hanya terlihat pada saat pengasuhan anak dan sosial reproduksi.
Pada satwa non-primata variasi kadar hormon stres sangat dipengaruhi oleh faktor lingkungan seperti iklim, kehadiran individu lain, status reproduksi dan status sosial. Pada saat melakukan hubungan sosial reproduksi akan terjadi peningkatan interaksi dengan individu lain. Orangutan betina sangat mempertimbangkan status dari individu jantan yang menjadi pasangannya. Perbedaan morfologi jantan yang bersifat bimaturisem membuat persaingan antar jantan dewasa berpipi dan berpipi untuk mendapatkan individu betina reproduktif semakin tinggi. Hal tersebut dapat membuat individu betina dalam keadaan terancam. Ancama yang terus menerus akan melebihi batas akan menyebabkan stres. Stres merupakan respon fisiologi dari dalam tubuh untuk menyesuaikan dengan perubahan lingkungan ataupun ancaman.
Metode pendeteksian kadar hormon stres dalam penelitian ini dengan cara non-invansif dengan menggunakan feses yang dikoleksi setiap pagi hari dengan menghomogenkan semua lapisan feses. Sedangkan metode pencatatan perilaku yang dilakukan dalam penelitian ini adalah Focal Animal Sampling secara
ii
Berdasarkan hasil penelitian diketahui bahwa perbedaan status reproduksi individu betina dan status sosial menpengaruhi respon individu betina terhadap
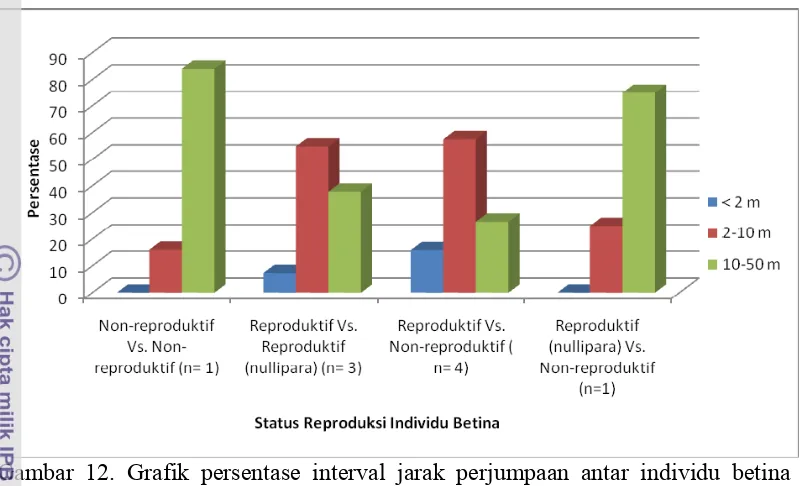
long call yang di dengar dan jarak perjumpaan dengan individu lain. Respon individu betina reproduktif nullipara dan non-reproduktif terhadap long call yang didengar dalam hubungan berpasangan memberikan respon abaikan Kondor (44,4%), Kerry (38,9%), Milo dan Mindy (5,7%). Sedangakan diluar hubungan berpasangan betina nullipara lebih memberikan respon positif terhadap long call Kondor (15,385) dan Milo (7,692%). Betina non reprodukti semua memberikan repon abaikan long call. Sebagian besar jarak perjumpaan yang terjadi antar individu betina non-reproduktif di 10-50m (84%), antar individu betina reproduktif di 2-10m (54,878%) dan antara individu reproduktif dengan non-reproduktif di 2-10m (41,176%). Individu betina non-reproduktif melakukan perjumpaan dengan individu jantan dewasa tidak berpipi umumnya dijarak 2-10m (69,293%) sedangkan individu betina non-reproduktif di 10-50m (33,155%). Perjumpaan antar individu betina reproduktif dengan jantan dewasa berpipi umumnya di 50m (38,357%) sementar individu betina non-reproduktif di 10-50m (48,951%). Selama hubungan berpasangan antara individu betina reproduktif dengan jantan dewasa tidak berpipi cenderung berjarak 2-10m (66,457%), sementara individu betina non-reproduktif pada 10-50m (58,155%). Kopulasi antara betina reproduktif dengan jantan dewasa berpipi dengan persentase kopulasi pasif (50%) dan aktif 50%. Kopulasi antara betina nullipara dengan jantan dewasa tidak berpipi kopulasi pasif 75%, 25% aktif dan 25% pemaksaan. Kategori kopulasi antara betina non-reproduktif dengan jantan dewasa tidak berpipi ataupun jantan dewasa berpipi semua dalam bentuk pasif
iii
mengalami peningkatan menjadi 347,50 ng/g (n=2). Mindy memiliki pola yang berbeda yaitu setelah terjadi interaksi jantan dewasa mengalami penurunan kadar hormon glukokortikoid, sebelum interaksi 470,53 ng/g (n=5) dan setelah interaksi mengalami penurunan 380,89 ng/g (n=3).
Berdasarkan hasil yang diperoleh bahwa dengan metode non-invansif melalui feses dapat diketahui profil horon glukokortikoid sebagai indikator stres. Didapatkan bahwa 11β-hydroxyetiocholanolone merupakan hormon glukokortikoid yang ditemukan dalam feses orangutan. Kedua penelitian tersebut dapat disimpulkan bahwa status reproduksi dan status sosial mempengaruhi interaksi dan interval jarak perjumpaan antar individu. Adanya korelasi antara interaksi dengan individu lain terhadap peningkatan kadar hormon glukokortikoid 11β-hydroxyetiocholanolone.
iv
SUMMARY
NENENG MARDIANAH. Interactions of female Orangutans (Pongo pygmaeus wurmbii) with other individuals and Its relationship with Glucocorticoid Metabolite Profiles In Orangutans Research Station in Tuanan, Central Kalimantan. Supervised by IMAN SUPRIATNA, MUHAMMAD AGIL and SRI SUCI UTAMI ATMOKO.
Bornean Orangutans (Pongo pygmaeus wurmbii) is one of the endemic primates the status of endangered according to the IUCN and was listed on Appendix I of CITES. Population declined caused by destruction, habitat decreased, and illegal arrests. The research of orangutans had been conducted mainly in primary forest, but still slightly research on of social behavior in orangutans located in secondary forest. Stasiun Penelitian Orangutan Tuanan (SPOT) is the orangutans habitat were included in the category of secondary forest and having a degraded was quite high so that easier to fire during the dry season. Orangutans is a frugivore animals and highly sensitive to the changes of habitat conditions. Habitat destruction massively affects the changes of social behavior of orangutans. Reduced range area increase in the occasion encounter between individuals increasingly higher, whereas the orangutans are a semi-solitary social system. Social behavior of orangutans is only visible when the parenting and social reproduction.
The level of stress hormones in non-primate animals highly influenced by environmental factors such as climate, the presence of other individuals, reproductive status and social status. At the time of social relationships, there will be increase in reproductive interactions with other individuals. The female of orangutans very considerate of the individual status of the male, who became her partner. The differences of male bimaturism morphological making a competition increasingly higher between flanged adult males and unflanged to get a female reproductive. It will effects a female to feels threatened. The threats continuously, will effects of excessive stress. Stress is a physiological response of the body to adjust the environmental changes or threats.
v
Based on observations it can be expressed that the difference of reproductive status and social status on the females individual affects the response female individuals towards the number of long call and distance encounter with another individual. The response of female reproductive nullipara and non-reproductive long call pairwise relations gives a respond to ignore Kondor (44.4%), Kerry (38.9%), Milo and Mindy (5.7%). Individual response of female reproductive nullipara and non-reproductive againts the number of long call in pairwise relations response to ignore Kondor (44.4%), Kerry (38.9%), Milo and Mindy (5.7%). Whereas beyond of the pairwise relations female nullipara responded positively to the long call Kondor (15.385) and Milo (7.692%). Of all the female non-reproductive gives a response to ignore the long call. Most of the encounter distance between female non-reproductive in a range 10-50m (84%), between the female reproductive at 2-10m (54.878%) and between female reproductive and non-reproductive at 2-10m (41.176%). Female reproductives doing an encounter, with unflanged adult males generally in the range 2-10m (69.293%), while female non-reproductive in a range 10-50m (33.155%). The encounter between female reproductive with flanged adult males generally at a distance of 10-50m with a percentage of 38.357% while the non-reproductive females with a distance of 10-50m at 48.951%. During the pairwise relations between individual female reproductive with unflanged adult male tends to within 2-10m with a percentage of 66.457%, while the females non-reproductive at a distance of 10-50m (58.155%). Copulation between female reproductive with flanged adult males by a passive copulation with a percentage (50%) and 50% for active. Copulation between female nullipara with unflanged adult male or flanged by a passive copulation 75%, 12,5% active and 12,5% for coercion. Category copulation between females non-reproductive to adult males or adult males flanged or unflanged all in the passive.
vi
was 291.87 ng / g (n=3), but after interaction increased to 634.75 ng / g (n=5). The encounter between June with unflanged adult males glucocorticoids caused by hormonal changes, the leveles of glucocorticoid hormones before the interaction was 267.70 ng / g (n=12) and after interaction increased to 347.50 ng / g (n=2). Mindy has a different pattern after the interaction with adult males, a levels of glucocorticoid hormones before interaction was 470.53 ng / g (n = 5) and after interaction decreased to 380.89 ng / g (n = 3).
Based on the results that have been obtainedthat the method of non-invansif through feces turns out that we can know the profile of glucocorticoid hormones as indicators of stress. It was found that 11β-hydroxyetiocholanolone a glucocorticoid hormone can be detected in the orangutans feces. Both of these studies concluded that the reproductive status and social status affects the interaction and interval of encounter between individuals. The correlation between the interaction with other individuals to increase the glucocorticoid hormone 11β -hydroxyetiocholanolone.
©
Hak Cipta Milik IPB, Tahun 2011
Hak Cipta Dilindungi Undang-Undang
Dilarang menguntip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Penguntipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
INTERAKSI ORANGUTAN (Pongo pygmaeus wurmbii) BETINA
DENGAN INDIVIDU LAIN DAN HUBUNGANNYA DENGAN
PROFIL METABOLIT GLUKOKORTIKOID DI STASIUN
PENELITIAN ORANGUTAN TUANAN, KALIMANTAN
TENGAH
NENENG MARDIANAH
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Strudi Biologi Repoduksi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah Subahanahuwata’ala atas segala limpahan karunia-Nya Shalawat dan keselamatan semoga senantiasa tercurah kepada insan pilihan dan rasul-Nya yang dengannya, kaum yang berada dalam kesesatan mendapat hidayah. Dialah nabi Muhammad SAW. Juga, semoga tercurah bagi keluarganya, sahabat dan saudara-saudaranya dari kalangan para rasul serta para malaikat yang selalu dekat dengan Allah SWT.
Alhamdulillah berkat rahmat Allah SWT dan kerja keras penulis, sehingga tesis dengan judul Interaksi Orangutan (Pongo pygmaeus wurmbii) Betina dengan Individu Lain dan Hubungannya dengan Profil Metabolit Glukokortikoid Di Stasiun Penelitian Orangutan Tuanan, Kalimantan Tengah telah berhasil diselesaikan. Karya tulis ini diajukan sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada program studi Biologi Reproduksi Departemen Klinik, Reproduksi dan Patologi Fakultas Kedokteran Hewan Institut Pertanian Bogor. Masih banyak kesalahan dan kekurangan dalam penulisan tesis ini. Oleh karena itu penulis sangat mengharapkan bimbingan, saran dan kritik membangun untuk dapat menyempurnakan tesis ini, sehingga dapat dijadikan bahan acuan oleh semua pihak.
Penulis mengucapkan terima kasih kepada Prof. Dr. drh Iman Supriatna, selaku Ketua Komisi Pembimbing dengan penuh ketulusan untuk senantiasa meluangkan waktu, memberi perhatian dan dengan sabar membimbing penulis untuk menyelesaikan tulisan ini, Kepada Dr. drh. Muhamad Agil, MSc. Agr. dan Dr. Sri Suci Utami Atmoko selaku anggota komisi pembimbing, yang dengan sabar membimbing penulis, memberikan dorongan, saran dan masukan yang membangun serta memberikan banyak kepercayaan kepada penulis untuk menyelesaikan tulisan ini.Ucapan terimakasih kepada Dr. Ir Dahrul Syah, M.Sc., Agr selaku dekan Pascasarjana IPB, Dr. drh Bambang Pontjo Priosoeryanto, MS. Ph.D. APVet., selaku ketua Departemen KRP Fakultas Kedokteran Hewan IPB, Prof. Dr. drh M. Agus Setiadi, selaku ketua Program studi BRP Fakultas Kedokteran Hewan IPB, serta terimakasih kepada seluruh dosen program studi BRP.
Penghargaan dan ucapan terima kasih disampaikan penulis kepada Prof Carel van Schaik, Tony Weingrill dan Maria van Noordwijk selaku menyandang dana beasiswa melanjutkan kegiatan pascasarjana ini hingga selesai. Drs. Imran SL Tobing M.Si. selaku Dekan Fakultas Biologi, Drs. Tatang Mitra Setia M.Si selaku komite beasiswa Fakultas Biologi bidang Primatologi yang telah memberikan kesempatan untuk bergabung dalam program kerjasama Universitas Nasional-Zurich of University dan kepercayaan untuk melanjutkan kegiatan pendidikan pascasarjana serta selalu memberi saran-saran yang mendukung. Selain itu, penulis juga ingin mengucapkan terimakasih kepada seluruh staf pengajar Fakultas Biologi Universitas Nasional, Jakarta atas semua bantuan dan perhatian yang diberikan selama ini.
diberikan kepada penulis untuk menyelesaikan penulisan ini. Abuk, Idun, Tono, Rahmat, Rahmadi dan Yandi atas segala kerjasama. Ayahanda Muhammad Sahud dan ibunda Nursiah atas do’a, pengorbanan dan kasih sayang yang tiada henti. Suami Abdul Azis dan ananda Rizky Adrian yang selalu mendukung, mendo’akan dan memberikan senyuman terindah dalam kekelahan menulis. Kepada kakak tercinta; Yopy Achmad, Luki Achmad, Darwanto, Anita, Nurasiah Astuti dan Sovi yang telah memberikan kasih sayang dan do’a.
Kepada teman seperjuangan; Fluer Scheele, Tobias Bollman, Davit Markus, Simon, Daniel dan Abigel Philips atas kebersamaan dan kerjasama. Teman diskusi Laura Chronika S.Pet, Oriza Savitriante S.Pt, Purwo Siswoyo S.Pt, drh. Nuryanto M.Si, drh. Leni Maylina, drh. Mawarsubangkit, drh. Dedi, drh Taufik M.Si dan Golib M.Si. Teman berbagi cerita; Ade Sunardi, Dwi Herlianingsih, Alm. Agnes Angkilinawati, Devi Margaret, Didik Nuraeli dan keluarga besar “Lutung” Forum Study Primata atas rasa persahabatan yang berarti. Tanpa mengecilkan arti, ucapan terima kasih juga disampaikan kepada seluruh kerabat dan handai taulan yang tidak tersebutkan
Akhir kata, penulis berharap semoga tesis ini dapat menjadi manfaat bagi bidang usaha pelestarian orangutan dan biologi reproduksi.
Bogor, Januari 2013
i Judul Tesis : Interaksi Orangutan (Pongo pygmaeus wurmbii) Betina dengan
Individu Lain dan Hubungannya Profil Metabolit Glukokortikoid di Stasiun Penelitian Orangutan Tuanan, Kalimantan Tengah
Nama : Neneng Mardianah NIM : B 352100041
Disetujui oleh Komisi Pembimbing
Ketua
Prof.Dr.drh. Iman Supriatna
Dr. drh. Muhammad Agil, M.Sc.Agr
Anggota Anggota
Dr. Sri Suci Utami Atmoko
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana IPB Biologi Reproduksi
Prof. Dr. drh. M. Agus Setiadi Dr. Ir. Dahrul Syah, M.Sc.Agr
viii
DAFTAR ISI
DAFTAR ISI viii
DAFTAR GAMBAR xi
DAFTAR TABEL xii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
Perumusanan Masalah Penelitan 3
Hipotesis 4
DAFTAR PUSTAKA 4
TINJAUAN PUSTAKA 6
Orangutan (Pongo pygmaeus wurmbii) 6
Taksonomi 6
Morfologi 6
Distribusi dan status konservasi 7 Biologi dan siklus menstruasi 8 Interaksi sosial orangutan 9
Konsep stres 13
Biosintesis hormon steroid 14 Biosintesis glukokortikoid 15
Sekresi hormon glukokortikoid 16 Metabolisme dan ekskresi hormon glukokortikoid 17
DAFTAR PUSTAKA 19
INTERAKSI ORANGUTAN (Pongo pygmaeus wurmbii) BETINA
BERKERABAT DENGAN INDIVIDU LAIN 24
ABSTRAK 24
PENDAHULUAN 24
BAHAN DAN METODE 25
ix
Peralatan 26
Hewan penelitian 26
Cara kerja 27
Analisa Data 29
Hasil dan Pembahasan 29
Respon Individu Betina Terhadap Long Call 29 Interval Jarak Perjumpaan Antar Individu Betina Berkerabat 32
Interval Jarak Perjumpaan Individu Betina dengan Individu Jantan
Dewas Berpipi dan Tidak Berpipi 34 Interval Jarak Antar Individu Betina dengan Individu Jantan Dewasa
Tidak Berpipi Pada Saat Berpasanga (Consort) 36 Inisiatif Individu Betina dalam Perilaku Seksual 37
Perilaku Seksual Kopulasi 38
Simpulan dan Saran 41
DAFTAR PUSTAKA 42
PROFIL HORMON GLUKOKORTIKOID PADA ORANGUTAN
(Pongo pygmaeus wurmbii) BETINA yang BERINTERAKSI
dengan INDIVIDU LAIN MENGGUNAKAN METODE
NON-INVANSIF 44
ABSTRAK 44
PENDAHULUAN 44
METODELOGI PENELITIAN 46
Waktu dan Lokasi Penelitian 46 Hewan Penelitian dan Status Reproduksi 46
Alat dan Bahan 46
Koleksi Feses 46
Analisa Hormon 47
Pengeringan dan Penghalusan Feses 47
Ekstraksi Feses 48
Pengujian Validitas Assay Hormon Glukokortikoid 48 Pengukuran Konsentrasi Metabolit Hormon 49
Analisa Data 49
x
Kadar hormon 11β- hydroxyetiocholanolone antar individu betina berdasarkan status reproduksi padasaat perjumpaan
dan tidak ada perjumpaan 50
Simpulan dan Saran 54
xi
DAFTAR GAMBAR
1. Morfologi orangutan (Pongo pygmaeus wurmbii) jantan dan betina 7 2. Persebaran oranguatan di Kalimantan 8 3. Siklus menstruasi pada primata 9 4. Mekanisme kerja hormon steroid pada target 14 5. Biosintesis perubahan kolesterol menjadi hormon steroid di korteks
adrenal 16
6. Sekresi hormon adrenokortikoid 17 7. Metabolisme glukokortikoid di dalam tubuh 18 8. Letak geografis camp Tuanan 26 9. Grafik persentase respon individu betina terhadap long call di dalam
hubungan kebersamaan dengan individu jantan 30 10. Grafik persentase respon individu betina terhadap long call di luar
hubungan kebersamaan dengan individu jantan 32 11. Silsilah keturunan orangutan betina berkerabat di areal penelitian
orangutan Tuanan 33
12. Grafik pesentase interval jarak perjumpaan antar individu betina
berkerabat berdasarkan perbedaan status reproduksi 34 13. Grafik persentase interval jarak perjumpaan individu betina reproduktif
dan non-reproduktif dengan individu jantan dewasa tidak berpipi dan
berpipi 35
14. Grafik persentase interval jarak antar individu betina dengan jantan
dewasa tidak berpipi pada saat hubungan berpasangan (Consort) 37
15. Koleksi feses 47
16. Kadar hormon 11β- hydroxyetiocholanolone orangutan betina 51 17. Kadar hormon 11β- hydroxyetiocholanolone pada individu
xii
DAFTAR TABEL
1. Individu orangutan betina pada saat pengamatan Oktober 2009-Juli 2010 27 2. Inisiatif individu betina dan jantan dalam perilaku seksual 38 3. Durasi waktu (menit) interaksi seksual antar individu betina dengan
1
PENDAHULUAN
Latar Belakang
Orangutan merupakan salah satu anggota suku Pongidae dan satu-satunya kera besar yang hidup di Asia, namun saat ini orangutan hanya dapat ditemukan di dua Pulau yaitu Sumatera dan Borneo (Groves 2001; van Schaik 2006). Persebaran orangutan di Sumatera hanya ditemukan di dua provinsi (Nanggroe Aceh Darussalam dan Sumatera Utara), sedangkan di Kalimantan orangutan hampir terdapat di semua provinsi kecuali Kalimantan Selatan, sementara di wilayah Malaysia beberapa populasi berada di bagian Sabah dan Serawak (Meijaard et al. 2001; Soehartono et al. 2007). Orangutan banyak ditemukan di dataran rendah, namun pada beberapa tahun terakhir populasi orangutan liar di alam mengalami penurunan. Faktor yang menyebabkan penurunan tersebut salah satunya adalah kehilangan dan kerusakan habitat, yang sangat mempengaruhi perilaku (Meijaard et al. 2001).
Status konservasi orangutan kalimantan, menurut International Union for Conservation of Nature and Nature Resources (IUCN) masuk ke dalam kategori terancam punah (endangered) sedangkan orangutan sumatera masuk dalam kategori kritis (critical endangered) hal ini disebabkan jumlah populasi dari orangutan sumatera yang terus mengalami penurunan. Menurut Convention on Internatonal Trade in Endangered Species of Wild Fauna and Flora (CITES) orangutan masuk dalam kategori Apendiks I yaitu merupakan jenis satwa yang dilindungi penuh dan dilarang untuk ditangkap, dibunuh, dipelihara, dan diperdagangkan baik dalam keadaan hidup maupun mati (Soehartono et al. 2007; Wich et al. 2008).
Penelitian-penelitian mengenai orangutan telah dan masih dilakukan di beberapa habitat orangutan liar umumnya memiliki tipe habitat hutan primer, di antaranya Taman Nasional Tanjung Puting (Kalimantan Tengah), Taman Nasional Gunung Palung (Kalimantan Barat), Taman Nasional Kutai (Kalimantan Timur), dan Ketambe serta Suaq Balimbing di Taman Nasional Gunung Leuser (Nangroe Aceh Darussalam). Namun masih sedikitnya penelitian yang dilakukan di tipe hutan sekunder atau terdegradasi oleh karena itu perlu untuk dilakukan penelitian lebih lanjut, sehingga hasil yang diperoleh dapat diperbandingkan untuk melihat dampak perubahan lingkungan terhadap perilaku orangutan.
2
Orangutan termasuk dalam hewan frugivore, mereka sangat peka terhadap perubahan kondisi habitat dikarenakan seluruh aktivitas bersumber dari hutan (Supriatna & Wahyono 2000). Perusakan habitat yang dilakukan secara besar-besaran berdampak negatif pada aktivitas orangutan seperti berkurannya area jelajah, terbatasnya sumber makanan yang mengakibatkan peluang perjumpaan antar individu semakin besar (Supriatna & Wahyono 2000; van Schaik 2006; Singleton et al. 2009). Tekanan akibat perubahan habitat dan perjumpaan dengan individu lain jantan maupun betina serta terdapatnya perbedaan status individu betina di suatu lokasi yang sama, secara terus menerus hingga melebihi batas toleransi dapat menyebabkan peningkatan kadar hormon stres (Smith & French 1997; Astuti 2006).
Penelitian mengenai orangutan telah banyak dilakukan terutama mengenai perilaku harian maupun sosial, namun masih sedikit informasi mengenai pengaruh perilaku sosial yang juga memperhatikan respon fisiologis. Stres merupakan respon fisiologi yang berasal dari dalam tubuh untuk menyesuaikan diri terhadap perubahan lingkungan maupun ancaman dari individu lain dengan adanya rangsangan yang berulang. Tanggap cekaman yang timbul dapat berupa kegelisahan, penurunan daya fertilitas, bahkan dapat menimbulkan kematian. Indikator tanggap cekaman dapat diketahui dengan melakukan pegukuran kadar hormon di dalam tubuh melalui darah, feses, urine, air susu maupun saliva (Thanos et al. 2008).
Hormon yang ada di dalam tubuh beredar melalui pembuluh darah, untuk hormon yang tidak terikat pada protein akan diekskresikan melalui urine dan feses. Hasil tersebut dapat dimanfaatkan untuk melakukan pengukuran profil hormon pada satwa liar. Beberapa penelitian telah berhasil mendeteksi kondisi fisiologi melalui pengukuran metabolit hormon dalam urine dan feses. Diketahui bahwa pada kondisi tertekan ataupun terancam menyebabkan terjadinya peningkatan kadar hormon stres, hal ini dikarena adanya kejadian hormonal di dalam tubuh. Pada keadaan normal hormon stres disekresikan secara episodik sehingga membentuk pola harian, sekresi hormon stres yang berlebih dan berlangsung secara terus menerus dapat menghilangkan pola harian hormon (Brook & Marshall 1996; Möstl & Palme 2002).
Tujuan Penelitian
1. Mempelajari interaksi antara individu betina dengan status reproduksi non-reproduktif, reproduktif dan nullipara terhadap individu lain.
2. Mempelajari profil hormon stres pada individu betina dengan status reproduksi non-reproduktif, reproduktif dan nullipara yang berinteraksi dengan individu lain.
3. Mempelajari korelasi perilaku sosial dengan profil hormon stres.
Manfaat Penelitian
3 Perumusan Masalah Penelitian
Berdasarkan hasil penelitian sebelumnya menyatakan bahwa tingkat reproduksi pada orangutan sangat rendah, hal ini disebabkan oleh rendahnya kemampuan individu betina untuk dapat melahirkan anak selama masa hidup. Menurut Wich et al. (2009). Orangutan betina hanya dapat melahirkan 3-5 anak selama masa hidupnya dan disetiap kelahirannya individu betina hanya mampu melahirkan satu anak dengan interval kelahiran sangat panjang yaitu 6-8 tahun. Rendahnya sistem reproduksi orangutan di dukung dengan sistem sosial dari orangutan yang bersifat semi soliter menyebabkan penurunan jumlah populasi orangutan di alam.
Pada saat melakukan hubungan sosial reproduksi individu betina sangat memperhatikan perbedaan status dari individu jantan yang menjadi pasangannya. Orangutan jantan memiliki tingkat perbedaan seksual dilihat dari perbedaan bentuk morfologi. Orangutan jantan memiliki sifat bimaturism yaitu adanya perbedaan morfologi antara individu jantan dominan dan tidak dominan yang diketahui setelah memasuki masa matang kelamin. Individu jantan dominan dapat dilihat dari munculnya sifat kelamin sekunder seperti berkembangnya bantalan pipi (cheek pad), kantong suara dan rambut panjang disekitar punggung serta lengan. Namun, sifat kelamin sekunder yang muncul pada individu jantan dominan tidak ditemukan pada individu jantan tidak dominan sehingga memiliki morfologi yang sama seperti individu betina.
Persaingan antar individu jantan mengalami peningkatan pada saat proses pemilihan pasangan, namun dalam hal ini individu betina menjadi pengambil keputusan (decision maker) dimana individu betina lebih memilih berpasangan dengan individu jantan dengan status yang lebih tinggi atau jantan dewasa berpipi (flanged). Tujuan individu betina berpasangan dengan jantan dewasa berpipi adalah untuk mendapatkan keturunan dengan genetika terbaik yang berasal dari individu jantan, mendapatkan perlindungan bagi dirinya serta keturunannya dari individu jantan lain. Hal ini menyebabkan tingginya tingkat persaingan antara individu jantan untuk mendapatkan individu betina yang diharapkan, sehingga menyebabkan tekanan pada individu betina.
4
Cekaman yang terus menerus melebihi batas toleransi dari suatu individu akan menyebabkan stres, misalnya cekaman yang disebabkan karena kehadiran individu lain disuatu tempat yang sama menyebabkan peningkatan kadar hormon stres baik pada orangutan jantan maupun betina (Smith & French 1997; Marty 2009). Stres merupakan respon fisiologi dari dalam tubuh individu untuk menyesuaikan diri terhadap perubahan maupun ancaman yang terjadi (Smith & French 1997). Pada primata non-manusia variasi kadar hormon stres tidak hanya dipengaruhi oleh variasi iklim, kehadiran individu lain, namun umur ataupun status sosial dan status reproduktif merupakan hal penting yang harus diperhatikan.
Metode pendeteksian aktivitas hormon biasanya dilakukan dengan metode invansif yaitu pengambilan sampel darah secara langsung dan berulang-ulang untuk analisa hormon steroid, namun hal ini tidak memungkinkan dilakukan pada satwa liar karena mengakibatkan hewan tersebut stres dan mengubah konsentrasi hormon stres yang diuji. Oleh karena itu pendekatan dengan metode non-invansif melalui feses merupakan metode yang lebih memungkinkan digunakan untuk satwa liar karena hewan tidak perlu dilakukan anastesi dan hewan tidak merasa terancam terhadap uji yang ingin dilakukan, sehingga dalam penggujian kadar metabolit hormon stres mendapatkan hasil yang lebih akurat.
Hipotesis
Harapan dari penelitian ini untuk menjawab hipotesis sebagai berikut:
1. Terdapat perbedaan interaksi antara individu betina dengan status reproduksi non-reproduktif, reproduktif dan nullipara terhadap kehadiran individu lain. 2. Adanya peningkatan dan fluktuasi kadar hormon stres pada individu betina
dengan status reproduksi non-reproduktif, reproduktif dan nullipara setelah terjadi interaksi dengan individu lain.
DAFTAR PUSTAKA
Astuti P. 2006. Kajian Metabolit Testosteron dan Kortisol di dalam Feses dan Urine dalam Hubungannya dengan Kualitas Spermatozoa Owa Jawa (Hylobates moloch) di Penangkaran. (Disertasi). Bogor: Pascasarjana Fakultas Kedokteran Hewan, Institut Pertanian Bogor.
Azwar, Gondanisam, Mistar HK, Ambriansyah. 2004. Survey Keanekaragaman Hayati (biodiversity) pada Hutan Rawa Gambut di area Mawas, Propinsi Kalimantan Tengah. Borneo Orangutan Survival Foundation. Jakarta.
Brook CGD, Marshall NJ. 1996. Essential Endocrinology, 3th Edition. Blackwell Science.
5 Marty P. 2009. Behavioural endocrinology of male Bornean orangutans (Pongo pygmaeus wurmbii). (Thesis). Anthropological Institute and Museum University of Zürich.
Meijaard E, Rijksen, HR, Kartikasari SN. 2001. Di Ambang Kepunahan, Kondisi Orangutan Liar di Awal Abad ke-21. Gibbon Foundation Indonesia. Jakarta. Möstl E, Palme R. 2002. Hormones as Indicators of Stress. Domestic Animal
Endocrinology 23:67–74.
Singleton I, Knott CD, Morrogh-Bernard C, Wich SA, van Schaik CP. 2009. Ranging Behavior of Orangutan Female and Social Organization, eds Orangutans: Behavioral Ecology and Conservation. Oxford University Press. US. 10: 205-215.
Smith TE, French JA. 1997. Psychosocial Stress and Urinary Cortisol Excretion in Marmoset Monkeys (Callithrix kuhli), Physiol Behav 62: 225-232.
Soehartono T, Soesilo HD, Andayani N, Atmoko SSU, Sihite J, Saleh C, Sutrisno A. 2007. Strategi dan Rencana Aksi Konservasi Orangutan Indonesia. Direktorat Jenderal Perlindungan Hutan dan Konservasi Alam. Departemen Kehutanan, Jakarta.
Supriatna J, Wahyono EH. 2000. Panduan Lapangan Primata Indonesia. Yayasan Obor Indonesia, Jakarta.
Thanos PKA, Cavigelli SA, Michaelides M, Olvet. M, Patel U, Diep MN, Volkow ND. 2008. Non-Invasive Method for Detecting the Metabolic Stress Response in Rodents: Characterization and Disruption of the Circadian Corticosterone Rhythm. University Park, PA, US. 58: 219-228.
van Schaik CP. 2006. Diantara Orangutan. Kera Merah Dan Kebangkitannya Kebudayaan Manusia. Yayasan Penyelamatan Orangutan Bormeo (BOS), Jakarta.
Wich SA, Meijaard E, Marshall AJ, Husson S, Ancrenaz M, Lacy RC, van Schaik CP, Sugardjito J, Simorangkir T, Traylor-Holzer K, Galdikas BMF, Doughty M, Supriatna J, Dennis R, Gumal M, Singleton I. 2008. The status of the orangutan: an overview of this current distribution. Oxford University Press. US.
6
TINJAUAN PUSTAKA
Orangutan (Pongo pygmaeus wurmbii)
Taksonomi
Menurut Groves (2001) orangutan dapat dibedakan menjadi dua jenis yaitu orangutan sumatera dan orangutan kalimantan, perbedaan kedua jenis orangutan tersebut dilihat dari perbedaan morfologi dan genetika. Menurut taksonomi orangutan termasuk dalam kingdom Animalia, phylum Chordata, kelas Mammalia, ordo Primata, family Pongidae, Genus Pongo, termasuk jenis Pongo pygmaeus. Menurut Warren et al. (2001); Arora et al. (2010) orangutan kalimantan dibagi kembali menjadi tiga anak jenis berdasarkan perbedaan genetika. Tiga anak jenis orangutan Kalimantan juga menempati daerah persebaran yang berbeda yaitu Pongo pygmaeus pygmaeus (Kalimantan Barat dan Serawak), Pongo pygmaeus wurmbii (Kalimantan Tengah dan Kalimantan Barat) dan Pongo pygmaeus morio (Kalimantan Timur dan Sabah).
Morfologi
Orangutan merupakan jenis kera besar yang hampir seluruhnya ditutupi oleh rambut kecuali bagian wajah, telapak tangan dan telapak kaki. Menurut Napier & Napier (1967); MacKinnon (1974); van Schaik et al. 2009 orangutan mempunyai memiliki warna rambut jingga sampai merah kecokelatan sesuai dengan pertambahan umur. Rambut pada orangutan kalimantan berwarna kecokelatan dan akan berkembang menjadi cokelat gelap sesuai pertambahan umur. Umumnya bentuk rambut pada orangutan keras, kaku dan lebih rapuh dengan ujung rambut pecah, untuk membedakan individu yang telah dewasa dapat dilihat rambut pada bagian tangan dan punggung yang lebih panjang dan berwarna gelap (Gambar 1).
Bentuk muka orangutan kalimantan lebih bulat, selain itu orangutan jantan dewasa pada bagian pipi dapat mengalami pelebaran karena adanya penebalan otot yang disebut dengan bantalan pipi (cheek pad). Pada orangutan jantan dan betina terdapat kantong suara pada bagian leher yang berguna untuk mengeluarkan seruan panjang (long call), namun kantong suara pada orangutan betina ini belum diketahui fungsinya (Galdikas 1985; Setia et al. 2009). Menurut Galdikas (1985) orangutan betina mengeluarkan seruan yang hampir sama dengan seruan panjang yang biasa dikeluarkan jantan dewasa, tetapi tanpa jeritan yang bergelombang. Seruan ini pernah dilakukan oleh betina dewasa pada waktu berselisih dengan anak yang sedang dalam proses penyapihan dan seruan dapat terdengar dari jarak sejauh 400 m. Ukuran tubuh jantan dewasa berbantalan pipi (flanged) dua kali lebih besar dari pada betina yaitu dengan berat tubuh jantan berkisar antara 50-90 kg, sedangkan berat tubuh orangutan betina berkisar 30-50 kg.
7
akan melahirkan menunjukkan pembengkakan bagian perineal yang sedikit demi sedikitnya pada saat beberapa bulan sebelum kelahiran (Galdikas 1985).
a. b.
c.
Gambar 1 : Morfologi orangutan (Pongo pygmaeus wurmbii) a) Flanged male, b)betina dewasa dan anak, c) betina dewasa bunting (pembengkakan perineal, perut dan puting susu) (Mardianah 2009).
Distribusi dan Status Konservasi
Orangutan banyak ditemukan di areal hutan hujan tropis yang merupakan habiat utama orangutan yaitu mulai dari dataran rendah, rawa gambut hingga hutan pegunungan. Kepadatan yang terbesar ditemukan pada tipe habitat hutan dataran rendah sekitar areal sungai dan hutan rawa gambut. Di Kalimantan orangutan dapat ditemukan pada ketinggian 500 m dpl, sedangkan orangutan sumatera dilaporkan dapat mencapai hutan pegunungan dengan ketinggian 1.000-1.500 m dpl (Soehartono 2007; Arora et al. 2010). Faktor ketinggian menggambarkan ketersediaan pakan. Keberadaan buah-buahan berdaging semakin menurun tajam pada tempat yang lebih tinggi, selain itu suhu yang dingin dan curah hujan yang tinggi merupakan suatu tekanan bagi orangutan untuk bertahan hidup.
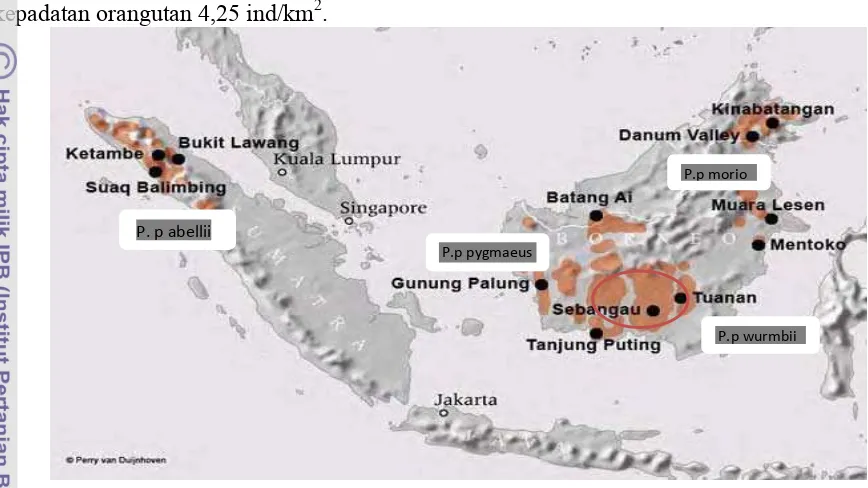
8 Orangutan di Indonesia berdasarkan surat keputusan Menteri Kehutanan No. 522/kpts-II/1997 merupakan jenis satwa yang dilindungi penuh dan dilarang untuk ditangkap, dibunuh, dipelihara, dan diperdagangkan baik dalam keadaan hidup maupun mati. Orangutan juga dilindungi hukum internasional, berdasarkan IUCN, orangutan kalimantan termasuk satwa yang terancam punah (endangered) (Soehartono et al. 2007). Menurut van Schaik (2005) jumlah populasi orangutan kalimantan di areal SPOT dengan luas area sebesar 2.730 km2 memiliki rata-rata kepadatan orangutan 4,25 ind/km2.
Gambar 2 : Persebaran orangutan di Kalimanatan dan Sumatera, serta Stasiun Penelitian Orangutan Tuanan(van Schaik et al. 2005)
Biologi dan Siklus Menstruasi
Orangutan memiliki siklus bereproduksi, dimulai dari masa pubertas atau dewasa kelamin, siklus menstruasi dan masa subur. Menurut hasil penelitian sebelumnya menyatakan bahwa tingkat reproduksi pada orangutan sangat rendah. Menurut Wich et al. (2009); Knott et al. (2009) orangutan betina liar yang berada di Kalimantan memasuki masa pubertas atau mampu memiliki keturunan pertama kali pada umur 14-16 tahun dengan jarak kelahiran inter birth interval (IBI) 6-8 tahun, sedangkan orangutan jantan pada umur 10 tahun mulai terlihat sifat-sifat reproduksi sekunder dan mulai melakukan perilaku reproduksi (Delgado & van Schaik 2000; Knott et al. 2009). Orangutan memiliki tingkat reproduksi yang rendah dengan lama kebuntingan 8-9 bulan dan setiap kelahiran orangutan hanya melahirkan satu anak. Selama masa hidupnya orangutan betina hanya mampu melahirkan 3-5 anak dan anak mulai tidak menyusu pada umur 5-6 tahun, kemudian anak akan hidup mandiri atau lepas dari pengawasan induk setelah berumur 7–15 tahun untuk anak jantan dan umur 7-12 tahun untuk anak betina (Wich et al. 2009; Knott et al. 2009). Orangutan yang memasuki masa pubertas ditandai dengan melakukan proses pemilihan pasangan.
Orangutan yang berada di dalam pemeliharaan atau captive (kebun binatang ataupun rehabilitasi) memasuki masa pubertas dan mampu mempunyai keturunan pertamakali jauh lebih muda dibandingkan orangutan liar, disebabkan
P.p pygmaeus
P.p wurmbii P.p morio
9
faktor habitat dan pemenuhan nutrisi yang tercukupi (van Schaik 2006). Orangutan jantan yang berada di captive akan memasuki masa pubertas pada umur kurang lebih 8-9 tahun, sedangkan orangutan betina akan memasuki masa pubertas pada umur 6 tahun dengan jarak kelahiran inter birth interval (IBI) 5-6 tahun (Delgado & van Schaik 2000; Wich et al. 2009; Knott et al. 2009).
Sama seperti jenis kera lainya orangutan juga mengalami siklus mestruasi (Gamabr 3). Menurut Maple (1980) dan Dixson (1998) siklus menstruasi pada orangutan kurang lebih 26-37 hari dengan lama waktu menstruasi 3-4 hari sama dengan jenis kera besar lainnya, tetapi lebih atau kurang tergantung dari mekanisme hormonal yang mempengaruhi siklus menstruasi (Dunbar 1988; Dixson 1998). Pada primata regenerasi penghancuran endometrium dengan pendarahan kurang lebih dua minggu setelah ovulasi (Singer 2000; Nelson 2005).
Menstruasi terjadi setelah proses ovulasi dimana terbentuk korpus luteum menyebabkan peningkatan hormon progresteron sehingga terjadi penebalan pada dinding endometrium, apabila tidak terjadi kebuntingan atau fertilisasi maka korpus luteum akan mengalami regresi sekitar 4 hari sebelum menstruasi dan diikuti oleh proses pendarahan. Pada akhir mestruasi semua lapisan endometrium terlepas kecuali lapisan dalam (Nelson 2005).
Pada orangutan berakhirnya siklus menstruasi akan diikuti dengan masa subur/fertil adalah masa pematangan sel telur sampai dengan pelepasan sel telur menuju tuba falopii (ovulasi). Perubahan perilaku orangutan betina yang sedang subur ditandai dengan meningkat ketanggapan (reseptivitas) atau kesediaan seks (proreseptivitas) terhadap kehadiran individu jantan (Galdikas 1985; Atmoko et al. 2009). Lama masa fertil/subur pada orangutan antara 2-3 hari satu hari setelah proses ovulasi. Ovulasi diperkirakan terjadi pada pertengahan siklus menstruasi, yaitu antara hari ke 11-13 (Disxon 1998; Nelson 2005).
Gambar 3 : Siklus menstruasi pada primata (Nelson 2005).
Interaksi Sosial Orangutan
10 satuan individu orangutan di lokasi yang sama < 50 m (Dunbar 1988; van Schaik 2006). Interaksi sosial dimulai saat suatu individu melakukan perjumpaan antara dua atau lebih individu orangutan yang salah satu atau kedua-duanya saling melihat, mengacuhkan, saling menyerang dan bermain. Beberapa aktifitas sosial yang teramati diantaranya adalah menelisik atau mengutui, perilaku berpasangan (consortship), kopulasi, dan mengasuh anak. Interaksi sosial dapat terjadi hampir pada setiap individu orangutan kecuali pada sesama jantan dewasa sangat jarang teramati (Galdikas 1985; van Schaik 2006). Berikut adalah aktifitas sosial dan interaksi sosial yang terjadi pada orangutan meliputi:
1. Aktifitas sosial
a. Pemilihan pasangan
Perilaku reproduksi pada orangutan dimulai dari proses pemilihan pasangan, saat satu individu memasuki dewasa kelamin ditunjukan dengan perubahan perilaku. Orangutan betina remaja yang telah memasuki dewasa kelamin akan menunjukan ketertarikannya pada individu jantan yaitu memamerkan organ genital atau sampai mendorong dirinya (van Schaik 2006). Menurut Galdikas (1985); van Schaik (2006) orangutan jantan dewasa yang memasuki dewasa kelamin juga menunjukan perubahan perilaku, yaitu mulai mendekati individu betina dan mengikuti individu betina pada jarak tertentu (consortship).
Faktor penting yang harus diperhatikan dalam proses pemilihan pasangan pada orangutan adalah status sosial dan umur (Dixson 1998; Atmoko et al. 2009; Knott et al. 2009). Pada saat proses pemilihan pasangan individu betina lebih memilih berpasangan dan kopulasi dengan individu jantan dewasa berpipi. Manfaat yang didapatkan oleh individu betina tersebut diantaranya adalah manfaat genetik, dimana anak akan mendapatkan gen-gen terbaik yang berasal dari individu jantan (van Schaik & van Hooff 1996; van Schaik 2006; Atmoko et al. 2009b). Strategi yang dilakukan oleh individu jantan dewasa berpipi untuk mendapatkan individu betina salah satunya dengan cara mengeluarkan long call
yang berfungsi untuk menarik perhatian individu betina yang berada di sekitarnya, sedangkan jantan dewasa tidak berpipi cenderung berjalan jauh memperluas daerah jelajah untuk bertemu dengan betina yang siap kawin (Delgado & van Schaik 2000; Atmoko et al. 2009a; Setia et al. 2009). Betina reproduktif biasanya akan memberikan respon positif setelah mendegar long call. Hal ini terlihat dari perubahan perilaku betina tersebut, yaitu mendekati long call namun pada betina yang sedang membawa anak cenderung memberikan respon menghindari long call (Galdikas 1985; Atmoko et al. 2009a).
Menurut Galdikas (1985) dan Atmoko et al. (2009b) sikap penolakan dalam hubungan berpasangan yang dilakukan orangutan betina merupakan usaha betina untuk mengurangi usaha kopulasi jantan. Hal ini karena akan mempengaruhi aktifitas betina seperti makan, berjelajah dan menyusui bayi. Pada saat terjadi hubungan seksual antara jantan dewasa dengan betina dewasa yang mempunyai anak, sering terlihat perlawanan yang dilakukan anak terhadap jantan yang ingin melakukan hubungan seksual dengan induknya (Galdikas 1985; Atmoko et al. 2009b).
b. Hubungan berpasangan (consortship)
11
kebersamaan antara individu jantan dan betina yang berlangsung beberapa hari hingga beberapa minggu, selama proses ini terjadi akan diselingi dengan beberapa kali kopulasi (Galdikas 1985; Atmoko et.al 2000). Hubungan berpasangan tergantung dari masa estrus individu betina, hubungan ini sangat penting untuk menentukan keberhasilan kopulasi karena akan terjadi peningkatan frekuensi kopulasi (Atmoko et al. 2009b; Setia et al. 2009). Orangutan jantan dewasa selalu kembali ke betina pasangannya pada waktu betina siap kawin kembali untuk memastikan kebuntingan dan keturunanya (Schurmann 1982; Galdikas 1985; Atmoko et al. 2009b). Hubungan berpasangan yang terjadi antara jantan dan betina tidak hanya terjadi pada satu individu yang sama tetapi dilakukan dengan beberapa individu yang berbeda.
c. Kopulasi
Perilaku kopulasi terjadi peningkatan setelah berakhirnya siklus menstruasi dan diikuti dengan masa subur/fertile. Orangutan betina yang sedang memasuki masa estrus akan menunjukan ketanggapan atau kesediaan seks terhadap jantan. Ketanggapan seks (reseptivitas) ialah kesediaan betina untuk melakukan kopulasi sedangkan kesediaan seks (proseptivitas) ialah semua perilaku yang dilakukan betina untuk memulai interaksi seks (Galdikas 1985).
Perilaku orangutan betina yang sedang birahi sama dengan jenis monyet atau kera lainnya yaitu berinteraksi dengan individu jantan untuk memamerkan atau memperlihatkan organ genitalnya di depan individu jantan yang diharapkan (Rijksen 1978; Galdikas 1985; Fox 2002; van Schaik 2006). Selain itu melekatkan organ genitalnya ke organ genital jantan dan membuat gerakan menusuk-nusuk (intromisi). Berdasarkan hasil penelitian Fox (2002) bentuk kopulasi dibedakan dalam dua golongan, yaitu: suka sama suka dan pemaksaan. Sementara itu Shurmann dan van Hoof (1986); Fox (2002) menambahkan dengan memasukan bentuk kopulasi setengah kerjasama.
c.1. Suka sama suka (betina aktif)
Kopulasi suka sama suka atau aktif adalah kopulasi yang ditandai dengan adanya inisiatif kedua individu mengadakan kopulasi dan lebih sering terjadi dalam hubungan berpasangan (consortship). Menurut Fox (2002), kopulasi suka sama suka atau aktif adalah kopulasi yang dimulai dari inisiatif individu betina untuk mengawali terjadinya proses kopulasi sedangkan individu jantan bersifat melayani individu betina. Bentuk kopulasi ini lebih sering terjadi di dalam proses hubungan berpasangan (consorthsip).
c.2. Pemaksaan (pemerkosaan)
12 c.3. Kopulasi setengah kerjasama (betina pasif)
Menurut Fox (2002) kopulasi setengah kerjasama ini ditandai dengan pasifnya individu betina terhadap usaha individu jantan, namun tidak ada perlawanan yang ditunjukan individu betina terdapat usaha individu jantan. adanya inisiatif jantan, sedangkan betina tidak mengiginkan terjadinya proses kopulasi namun tidak ada perlawanan yang dilakukan oleh betina atas tindakan yang dilakukan jantan. Menurut Fox (2002), inisiatif yang dilakukan dalam memulai kopulasi setengah kerjasama biasanya jantan lebih mengarahkan posisi kopulasi betina. Kopulasi setengah kerjasama ini banyak terjadi antara individu jantan dewasa tidak berpipi dengan betina dewasa atau betina remaja (Fox, 2002).
2. Hubungan antar individu lain
Bentuk hubungan antar individu orangutan yang sesama jenis kelamin terlihat dalam bentuk agresi, agonistik dan pengelompokan sosial, sedangkan hubungan sosial antar individu orangutan yang berbeda jenis kelamin bertujuan melakukan hubungan sosial reproduksi. Agresi adalah bentuk pamer kemarahan dengan cara menyerang atau pengejaran. Agonistik merupakan bentuk sikap tindak ketidak senangan akan kehadiran individu lain. Kedua peristiwa tersebut terjadi karena perebutan daerah kekuasaan dan perebutan individu betina yang tanggap dan sedia seks (estrus) (van Schaik & van Hooff 1996; Delgado & van Schaik 2000; Atmoko et al. 2009a). Hubungan antar individu kelamin sejenis dapat terjadi pada:
a. Hubungan betina dengan betina
Bentuk interaksi sosial yang terlihat pada sesama individu betina diantaranya adalah agresi, agonistik dan pengelompokan sosial. Tingkat persaingan antara individu betina dewasa dapat dilihat dari kedekatan kekerabatan suatu individu, perbedaan umur individu betina dan banyaknya jantan dewasa yang bersedia kawin dengan betina tersebut. Berdasarkan kekerabatanya maka dibedakan menjadi dua kelompok yaitu:
a.1. Hubungan betina yang tidak berkerabat
Interaksi antar individu betina dengan kekerabatan yang jauh yaitu interaksi sosial agresi dan agonistik. Kedua peristiwa tersebut terjadi disebabkan adanya usaha perebutan daerah kekuasaan dan sumber pakan yang ada (van Schaik & van Hooff 1996; Delgado & van Schaik 2000; Singleton et al. 2009). Individu betina remaja lebih cenderung menghindar diri dari kehadiran betina dewasa, baik yang sedang membawa anak maupun betina dewasa yang tidak bawa anak. Perbedaan status pada individu betina di satu tempat yang sama menimbulkan rasa takut untuk melakukan perjumpaan (Galdikas 1985; van Schaik & van Hooff 1996; Setia et al. 2009). Secara fisiologis interaksi sosial yang terjadi antar sesama jenis kelamin dapat menyebabkan peningkatan kadar hormon stres suatu individu (Smith & French 1997; Nelson 2005).
a.2. Hubungan betina yang berkerabat
13
b. Hubungan antar individu betina dengan individu jantan
Hubungan antar individu betina dengan individu jantan bersifat sosial reproduksi dimana tujuannya adalah mencapai sukses bereproduksi. Individu betina merupakan penentu pengambil keputusan dalam usaha sukses reproduksi. Perbedaan status sosial pada individu jantan merupakan pertimbangan individu betina dalam pengambil keputusan. Individu betina lebih memilih berpasangan dengan individu jantan dominan untuk mendapatkan keuntungan dari individu jantan pasangannya diantaranya adalah askes mendapat makanan dan perlindungan dari individu lain. Hubungan antar individu betina dengan individu jantan dimulai dari proses pemilihan pasangan, hubungan berpasangan dan berakhir pada proses kopulasi.
c. Hubungan sosial ibu dan anak
Hubungan sosial ini merupakan hubungan sosial yang konsisten setelah pengelompokan berpasangan (Rodman & Mitani, 1987; Galdikas, 1984). Proses pembelajar dan bermain bersama terjadi di dalam hubungan tersebut (Singleton et al. 2009). Menurut Atmoko (1991); Fox (2002) hubungan kopulasi yang terjadi pada individu betina yang membawa anak, sering telihat perlawanan yang dilakukan oleh individu anak terhadap usaha individu jantan yang ingin melakukan hubungan kopulasi dengan induknya. Perlawanan ini ternyata lebih efektif, karena jantan dewasa merasa terganggu
Konsep Stres
Menurut Smith dan French (1997); Nelson (2005) stres merupakan keadaan biologis, emosional dan tingkah laku yang tidak spesifik, tetapi dapat mengganggu ataupun mengancam keselamatan. Gejala stres terlihat sebagai gangguan psikologis, emosi maupun kegelisahan (Smith & French 1997). Berdasarkan perbedaan waktu stres dapat dibagi menjadi tiga yaitu stres jangka pendek ditandai dengan terjadi peningkatan denyut jantung, ketegangan otot dan perpendek pernafasan; stres jangka panjang dengan gejala berkurangnya nafsu makan, gangguan pencernaan, kulit serta penurunan libido; stres internal dengan gejala yang muncul ketakutan, ketidak mampuan berkonsentrasi. Penyebab stres (stresor) berasal dari situasi lingkungan, rangsangan atau kejadian yang mampu menganggu dan mengancam keselamatan makhluk hidup (Guyton & Hall 1996).
14
Biosintesis Hormon Steroid
Berdasarkan organ targetnya hormon steroid terbagi menjadi dua sub kelompok yaitu hormon seksual (progestrin dan estrogen) dan homon adrenal. Pada kolesterol struktur kerangka C-27 berasal dari Acetyl-CoA yang telah mengalami serangkaian proses yaitu diawali dengan pembentukan asetat menjadi mevalonat dengan bantuan enzim HMG-CoA reduktase yang kemudian diubah menjadi squalane dan dilanjutkan dengan lanosterol. Selanjutnya, lanosterol diubah menjadi kolesterol sebagai produk dengan mengambil tiga gugus karbon. Kolesterol banyak terdapat di membran sel yang merupakan salah satu komponen terpenting bagi kelangsungan hidup sel, selain itu kolesterol juga sebagai prekursor hormon steroid (Litwack &Schmidt 2002; Steimer 2003).
Biosintesis hormon steroid dimulai dari pengubahan kolesterol menjadi pregnenolon. Pengaturan biosintesis hormon steroid disebabkan oleh peningkatan cAMP intraseluler atau Ca2+ melalui inositol trifosfat. Rangsangan pada cAMP dapat bersifat akut atau kronis (beberapa jam sampai hari). Rangsangan dimulai sejak pengiriman kolesterol ke dalam inner mitokondria dengan perantara
steroidogenic acute regulatory (StAR), sedangkan rangsangan kronis terjadi pada saat pengubahan kolesterol menjadi pregnanolon (Gambar 4). Pada keadaan tersebut proses konversi berlangsung di dalam mitokondria dengan bantuan enzim
side chain cleavage (SCC), NADPH, oksigen dan sitokrom P450 sesuai kebutuhan. Fungsi Ca2+ pada proses ini adalah sebagai peningkat enzim SCC (Litwack &Schmidt 2002).
Gambar 4. Mekanisme kerja hormon steroid pada target (Steimer 2003)
Reseptor hormon steroid terletak di dalam sitoplasma sel, berbeda pada hormon protein yang berada di dinding sel. Tahap pertama hormon masuk ke dalam sel secara difusi dan segera berikatan dengan reseptor protein spesifik di dalam sitoplasma. Reseptor hormon steroid secara inaktif dipelihara dalam suatu
15
dalam nukleus terjadi ikatan kompleks antara hormon dan reseptor yang mempengaruhi koaktivator dan faktor transkripsi secara menyeluruh untuk menghasilkan kompleks transkripsional aktif yang kemudian akan meningkatkan ekspresi target gen dan menimbulkan efek hormon steroid (West 1991; Kuiper et al. 1998).
Biositesis Glukokortikoid
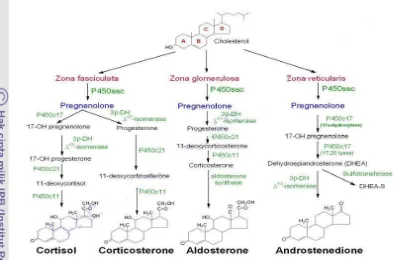
Hormon glukokortikoid merupakan hormon steroid yang disintesis dari kolesterol dan diproduksi oleh kelenjar adrenalis bagian korteks. Hormon disekresikan oleh organ endokrin yang diatur oleh hipotamus dan syaraf pusat. Korteks adrenal mensintesis molekul steroid yang dibagi menjadi tiga kelompok hormon yaitu glukokortikoid, mineralkortikoid dan androgen dengan zona atau lapisan penghasil yang berbeda-beda. Glukokortikoid berfungsi mengatur metabolisme karbohidrat, mineralokortikoid berfungsi mengatur kadar cairan (natrium dan kalium) di dalam tubuh dan androgen yang fungsinya hampir sama dengan hormon steroid yang dihasilkan oleh gonad jantan. Korteks adrenal terdiri dari 3 bagian utama yaitu zona glomerulosa, zona fasciculata, dan zona reticularis (Guyton 1996; Nelson 2005).
Menurut Martin dan Crump (2003); King (2009) pada setiap zona di korteks adrenal memiliki kesamaan dimana jalur utama yang dilewati untuk proses sintesis yaitu kolesterol diubah menjadi pregnenolon yang dibantu oleh enzim sitokrom P450 (CYP). Produk akhir yang dihasilkan oleh hormon steroid tergantung pada enzim yang berada di dalam sel masing-masing zona (Gambar 5). Kolesterol yang akan diubah menjadi pregnenolon di dalam korteks adrenal harus diangkut ke dalam mitokondria di mana CYP11A1. Proses transportasi dibantu oleh protein regulator akut steroidogenik (StAR).
Menurut King (2009) konversi prenenolone untuk progesteron memerlukan dua kegiatan enzim HSD3B2: dehidrogenase 3β-hidroksisteroid dan
Δ4,5-isomerase. Zona glomerulosa membutuhkan enzim P450c17 yang mengubah pregnenolon dan progesteron untuk C17 analog dihidroksilasi. Dengan demikian, jalur glukokortikoid (deoxycortisol dan kortisol) dan androgen (dehydroepiandosterone (DHEA) dan androstenedione) di simpan di dalam sel. Zona glomerulosa mengandung enzim yang bertanggung jawab untuk mengkonversi corticosterone untuk aldosteron. Pregnenolone yang dihasilkan di zona fasciculate akan diubah menjadi dua produk yang dibantu oleh dua enzim yang berbeda yaitu 17-OH pregnenolone dibantu enzim P450c17 dan progresteron dibantu enzim 3β-DH∆45-isomer. Progresteron kemudian mengalami biositesa menjadi 11-deoxycorticosterone dibantu dengan enzim P450c21 yang kemudian dikonversikan menjadi corticosterone (Martin & Crump 2003).
16 Enzim 3β-DH∆45-isomer membantu proses konversi Dehydroepiandrosterone menjadi Adrostenedione.
Gambar 5 : Biosintesis perubahan kolesterol menjadi hormon steroid di korteks adrenal (Martin & Crump 2003; King 2009).
Sekresi Hormon Glukokortikoid
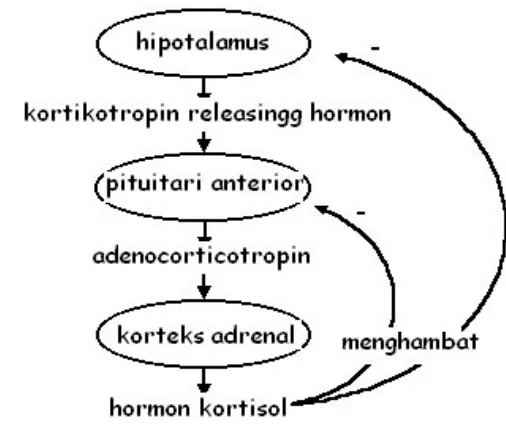
Hormon adalah senyawa yang yang secara normal dikeluarkan oleh kelenjar endokrin atau jaringan tubuh dan dilepaskan ke pembuluh darah menuju jaringan sasaran, berinteraksi secara selektif dengan reseptor khas dan menunjukkan efek biologis. Smith dan French (1997); Nelson (2005) menyatakan bahwa respon stres dipengaruhi oleh sistem endokrin di dalam hipotalamus dan kelenjar hipofisis. Tubuh mempunyai kemampuan fisiologis untuk beradaptasi terhadap kondisi stres. Respon neuroendokrin terhadap stres terjadi pada saat implus syaraf simpatik dari hipotalamus menstimulir kelenjar endokrin segera melepaskan katekolamin (epinepbrin/adrenalin dan noradrenalin) ke dalam aliran darah. Sekresi adreno corticotropic hormone (ACTH) diatur secara umpan balik oleh steroid yang beredar (Gambar 6). Hormon stres bebas berada di dalam darah dan memiliki umpan balik negatif terhadap pelepasan corticotrophin releasing hormone (CRH) dari hipotalamus terhadap kortikotrof hipofisis (Gambar 6).
17
Gambar 6: Sekresi hormon glukokortikoid (Nelson 2005).
Menurut Nelson (2005) menyatakan bahwa hormon steroid pada korteks adrenal dan medulla berperan dalam suatu respon fisiologi terhadap keadaan darurat. Indikator stres dapat dilihat dari peningkatan kadar hormon stres yang terjadi pada pagi hari dan mencapai titik terendah pada malam hari. Beredarnya hormon stres dalam darah kurang lebih 10% dan sisahnya akan berikatan di dalam trancortin maupun albumin yang kemudian akan dihancurkan dalam beberapa jam setelah berada di dalam organ target. Steroid adrenal akan didegradasi di dalam hati dan dikonjugasikan dalam bentuk glukoronida dan sulfat, 25% dari jumlah tersebut akan dieksresikan ke dalam empedu sampai dikeluarkan memalalui feses ataupun urine. Hormon yang telah terkonjugasi mempunyai sifat yang tidak aktif (inaktivasi).
Metabolisme dan Ekskresi Hormon Glukokortikoid
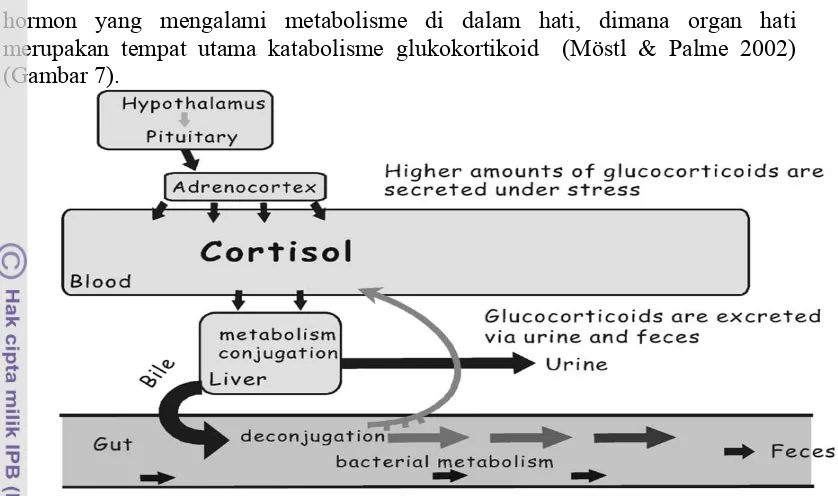
Hormon yang telah di sekresikan akan beredar ke seluruh tubuh melalui aliran darah karena hormon tidak memiliki saluran khusus dan secara lemah berikatan dengan protein. Pada waktu tertentu hormon akan dipecah menjadi senyawa tidak aktif dan selanjutnya dieksresikan dari dalam tubuh dalam bersama feses dan urin (Möstl & Palme 2002). Hormon yang tidak terikat di dalam jaringan juga akan diubah di dalam hati sebelum dieksresikan melalui usus dan ginjal (Guyton & Hall 1996).
18 hormon yang mengalami metabolisme di dalam hati, dimana organ hati merupakan tempat utama katabolisme glukokortikoid (Möstl & Palme 2002) (Gambar 7).
Gambar 7. Metabolisme glukokortikoid di dalam tubuh (Möstl & Palme 2002). Pemilihan sampel secara non-invansif ataupun invansif dalam deteksi hormon biasanya dikaitan dengan tujuan penelitian karena masing-masing sampel mempunyai kelebihan dan kekurangan. Pada satwa liar pengkoleksi darah dengan intensitas yang demikian tinggi akan menimbulkan traumatik, sehingga profil hormon yang terdeteksi akan berubah dari sifat alaminya akibat stres yang sangat tinggi. Pada proses koleksi metode invansif (sampel darah) seringkali hewan harus di tangkap, dibius atau dika ndangkan pa da ukura n ya ng se mpit . Resiko kematian menjadi semakin tinggi, terutama dengan adanya pembiusan selain itu dapat menyebabkan stres yang tinggi pada hewan tersebut. Pada satwa liar untuk mendapatkan hasil profil hormon yang akurat sebaiknya pengkoleksian sampel dilakukan dengan cara non-invansif, dengan menjadikan satwa liar tidak stres yang akan menganggu pola pelepasan hormon (Schwarzenberger 2007; Thanos et al. 2008) .
Keuntungan menggunakan metode non-invansif yaitu tidak perlu dilakukan anastesi sehingga menggurangi kejadian stres pada saat penangkapan maupun anastesi dan hewan tidak merasa tertekan ataupun terancam sehingga hasil yang didapatkan menjadi lebih akurat (Heistermann et al. 2001; Hua et al. 2004; Schwarzenberger 2007). Kerugian menggunakan metode non-invansif (feses) adalah sampel feses harus diambil secara keseluruhan karena kandungan metabolit hormon pada bagian luar dan dalam berbeda.
19
Kadar glukokortikoid yang berada di urine maupun feses setiap spesies berbeda-beda dan waktu yang dibutuhkan setiap hormon yang dieksresikan melalui urine ataupun feses membutuhkan waktu yang berbeda pula pada setiap spesies (Möstl & Palme 2002; Schwarzenberger 2007; Thanos et al. 2008). Salah satu hormon glukokortikoid yang telah diuji yaitu kadar hormon kortisol yang ditemukan di dalam feses pada masing-masing spesies memerlukan waktu yang berbeda disebabkan perbedaan metabolisme masing-masing jenis, misalnya pada
Macaca fasicularis kadar kortisol dapat terdeteksi pada 22 jam, Chimpanzee 26 jam (Hua et al. 2004; Heistermann et al. 2001).
Menurut Marty (2009) kadar hormon stres pada orangutan liar dapat dideteksi 48 jam kemudian setelah terjadinya interaksi sosial, namun jika dideteksi lebih dari 50 jam ke atas kadar hormon stres yang terdeksi di dalam feses akan menurun. Hal ini juga sesuai dengan hasil penelitian Weingrill et al. (2008) yang menyatakan bahwa kadar hormon stres pada orangutan di captive dapat dideteksi melalui feses setelah 48 jam. Metabolit hormon glukokortikoid yang dideteksi sebagai indikator stres pada orangutan adalah 11β- hydroxyetiocholanolone.
DAFTAR PUSTAKA
Ackerman U. 1996. Human Physiology. Mosby, St Louis.
Atmoko SSU, Mitra ST, Goossens B, James SS, Knott CD, Bernard M, van Schaik CP, van Noordwijk MA. 2009 a. Orangutan Matting Behavior and Strategi, eds. Orangutans: Geographic Variation in Behavior Ecology and Conservation. Oxford University Press. US.
Atmoko SSU, Singleton I, van Noordwijk MA, van Schaik CP, Mitra ST. 2009 b. Male-male Relationships in Orangutan. eds. Orangutans: Geographic Variation in Behavior Ecology and Conservation. Oxford University Press. US.
Atmoko SSU. 2000. Bimaturism in Orangutan Males: Reproductive and Ecological Strategies. (Thesis). Faculty Biology Utrecht University. The Nederlands.
Atmoko SSU. Perilaku Seksual Orangutan (Pongo pygmaeus abelii, Lesson 1827). 1991. Betina Dewasa di Pusat Penelitian Ketambe Aceh Tenggara. (Skripsi). Sarjana Biologi Fakultas Biologi Universitas Nasional. Jakarta. Arora N, Natera A, van Schaik CP, Erik P. Willemsa, van Noordwijka MA,
20 Brook CGD, and Marshall NJ. 1996. Essential Endocrinology, 3th Edition.
Blackwell Science. John Street, London.
Delgado RAJR , van Schaik, CP. 2000. The Behavioral Ecology and Conservation The Orangutan (Pongo pymaeus): A Tale of Two Island. Evolutionary Antrophology 20: 201-218.
Denhard M. 2004. Sample Collection, Handling, Storage and Preparation. Institute for Zoo and Wildlife Research, Berlin. Germany.
Dixson AF. 1998. Primate Sexsuality Comparative Studies of the Prosimians, Monkeys, Apes. and Humans Beings. Oxford Universty Press. US.
Dunbar RIM. 1988. Primate Social System: Study Behavior Adaptation. Dept. Zoology Universitas Liverpool, Croom Helm.Itd.
Fox EA. 2002. Female Tactics to Reduce Sexsual Harassment in The Sumatera Orangutan (Pongo pymaeus abelii). Behavior Ecologi Socio Biologi. Springer 52: 93-101.
Fowler ME. 1999. Zoo and Wild Animal Medicine 4th ed. Philadelphia: W.B. Saunders Company. Vol. 34-35.
Galdikas BMF. 1985. Adult Male Sociality and Reproductive Tactic Among Orangutan at Tanjung Puting. American Journal of Primatology. 45; 9-24. Ganong WF. 1983. Fisiologi Kedokteran Edisi 10. EGC. Jakarta.
Groves CP. 2001. Primate Taxsonomi Smithsonian Institution Press. Washington and London. US.
Guyton AC, Hall JE. 1996. Textbook of Medical Physiology, 9th ed. Saunders Company, Philadelphia, Pennsylvania. US.
Hau J, Abelson KS, Carlsson HE, Royo F. 2004. Stress-Assassed by Quantification of Non-invansif Stress Markers. University of Copenhagen & State Hospital and Uppsala University.
Heistermann M, Uhrigshardt J, Husung A, Kaumanns W, Hodges JK. 2001. Measurement of fecal steroid Metabilites in the lion-tail Macaque (Macaca silenus): a Non-invansif Tool for Assessing Female Ovarian Function. Primate report. American Journal of Primatology. 59:27-42.
Knott CD, Thompson ME, Wich SA. 2009. The Ecology of Female Reproduction in Wild Orangutan. eds. Orangutans: Geographic Variation in Behavior Ecology and Conservation. Oxford University Press. US.17: 171-188.
21
Kuiper GJM, Carlquist M, Gustaffson JA. 1998. Estrogen Is a male and female Hormones. Science and Medicine.10: 36-45.
Litwack G, Schmidt TJ. 2002. Biochemistery of Hormones II: Steroid Hormones in: G. Litwack and TJ. Schmidt. Textbook of Biochemistery and Clinical Correlations 5th ed. John Wiley and Sons. Pp 959-988.
MacKinnon JR. 1974. The Behaviour and Ecology of Wild Orangutan (Pongo pygmaeus). Animal Behaviour. 22: 3-74.
Maple TL. 1980. Orangutan Behavior. Primate Behavior and Development Series van Nostrand Reinhold Company.US.
Martin PA, Crump MH. 2003. The Adrenal Gland. In: Pineda MH, Dooley MP, editor. Veterinary Endocrinology and Reproduction. 5th Ed. US: Lowa State Press. Pp 165-200.
Marty P. 2009. Behavioural endocrinology of male Bornean orangutans (Pongo pygmaeus wurmbii). (Thesis). Anthropological Institute and Museum University of Zürich.
Meijaard E, Rijksen, HR, Kartikasari SN. 2001. Di Ambang Kepunahan, Kondisi Orangutan Liar di Awal Abad ke-21. Gibbon Foundation Indonesia. Jakarta. Möstl E, Palme R. 2002. Hormones as Indicators of Stress. Domestic Animal
Endocrinology 23:67–74.
Napier JR and Napier PHA. 1967. Hand Book of Living Primates. Academic Press. US.
Nelson R. 2005 An Introduction to Behavioral Endocrinology. Third Edition. The Ohio State Univesity. Sinauer Associates, Inc Publishers Sunderland, Massachusetts.
Rijksen HD. 1978. A Field Study on Sumatra Orangutans (Pongo pygmaeus abelii, Lesson 1827) Ecology Behaivior and Conservation. (Disseration) Argricultur University Wageningen.
Rodman CG, Mitani JC. 1987. Orangutans Sexual Dimorpisme in A Solitary Spesies in Primate Societies. The University of Chicago Press.
Schwarzenberger F, Mostl E, Palme R, Bamberg E. 1996. Faecal Steroid Analysis for non invansive Monitoring of Reproductive Status in Farm, Wild and Zoo Animals. Animal Reproduction Scince. 42: 515-526.
22 Singleton I, Knott CD, Morrogh-Bernard C, Wich SA, van Schaik CP. 2009.
Ranging Behavior of Orangutan Female and Social Organization, eds Orangutans: Behavioral Ecology and Conservation. Oxford University Press. US.
Smith TE, French JA. 1997. Psychosocial Stress and Urinary Cortisol Excretion in Marmoset Monkeys (Callithrix kuhli), Physiology Behavior 62(2): 225-232. Schurmann CL, Van Hooff JARAM. 1986. Reproduktive Strategies of the
Orangutan. New data and a reconsideration of existing sosiosexsual model. International Journal of Primatology. 7: 265-287.
Schurmann CL. 1982. Courtship and Mating Behavior of Wild Orangutan in Sumatera dalam Primate Behavior and Sociobiology, A. B. Chiarelli dan R. S. Corruccini, Springer Verlag, Berlin Heidelberg.
Soehartono T, Soesilo HD., Andayani N, Atmoko SSU, Sihite J, Saleh C, Sutrisno A. 2007. Strategi dan Rencana Aksi Konservasi Orangutan Indonesia. Direktorat Jenderal Perlindungan Hutan dan Konservasi Alam, Departemen Kehutanan. Jakarta.
Steimer. 2003. Steroid Hormone Metabolism. (Internet).
http://www.gfmer.ch/books/reproductive health/steroid hormone metabolism (6 September 2012)
Supriatna J. Wahyono EH. 2000. Panduan Lapangan Primata Indonesia. Yayasan Obor Indonesia. Jakarta.
Thanos PKA, Cavigelli SA, Michaelides M, Olvet. M, Patel U, Diep MN, Volkow ND. 2008. Non-Invasive Method for Detecting the Metabolic Stress Response in Rodents: Characterization and Disruption of the Circadian Corticosterone Rhythm. University Park, PA, US. 58: 219-228.
van Schaik CP, van Hooff JARAM. 1996. Toward an Understanding of The Orangutan’s Social System in McGrew, WC, Marchout, LF, and Nishida T, eds. Grear Ape Societies. University Press, Canbreidge, pp 3-15.
van Schaik CP. 2006. Diantara Orangutan. Kera Merah Dan Kebangkitannya Kebudayaan Manusia. Yayasan Penyelamatan Orangutan Bormeo (BOS), Jakarta.
23
Wich SA, Meijaard E, Marshall AJ, Husson S, Ancrenaz M, Lacy RC, van Schaik CP, Sugardjito J, Simorangkir T, Traylor-Holzer K, Galdikas BMF, Doughty M, Supriatna J, Dennis R, Gumal M, Singleton I. 2008. The Status of The Orangutan: an Overview of This Current Distribution. Oxford University Press. US.
Wich SA, Vries de Han, Ancrenaz M, Perkins L, Shumaker RW, Suzuki A, and
van Schaik CP. 2009. Orangutan Life History Variation. eds. Orangutans:
Geographic Variation in Behavior Ecology and Conservation. Oxford University Press. US.
Weingrill T, Willems EP, Zimmermann, Steinmetz H, Heistermann M. 2011. Spesies-Specific Patterns In fecal Glukokortikoid and Androgen Levels Zoo
Living Orangutan (Pongo spp.). American Journal of primatology. 172:
446-457.