_____
intesis dari asam mudian dapat diu
mudian mengala
loil‐glukosa. Gal kosa akan mengh
ng dapat terhidro
onizer et al. 2006
mbar 2.1. Produk
al. 2006
___________Senyawa F
NTESIS
SENY
enyawa fenolik
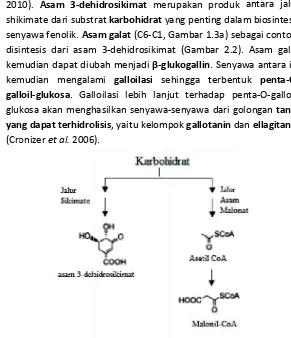
wali melalui jalur s hidrosikimat mer rat karbohidrat y am galat (C6‐C1,
m 3‐dehidrosikima
bah menjadi β‐gl
ami galloilasi se loilasi lebih lanj hasilkan senyawa
Fenolik pada Sayura
YAWA
FENO
at (Gambar 2.2).
lukogallin. Senyaw
ehingga terbentu
jut terhadap pe
‐senyawa dari go pok gallotanin da
dan asam malonat
an Indigenous
OLIK
r terjadi di ar 2.1) (Wink antara jalur m biosintesis bagai contoh, . Asam galat wa antara ini uk penta‐O‐
nta‐O‐galloil‐ olongan tanin
an ellagitanin
t (Cronizer et
©SEAFAST
Center
Senyawa Fenolik pada Sayuran Indigenous ____________________________________
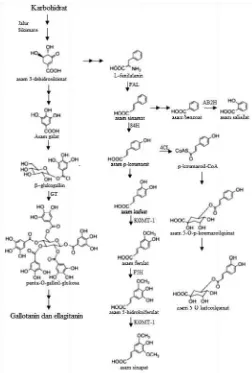
Di tanaman, asam 3‐dehidrosikimat selain diubah menjadi
asam galat juga digunakan untuk menyintesis L‐fenilalanin dan
mulai memasuki jalur fenilpropanoid. Melalui bantuan enzim
fenilalanin amonia liase, L‐fenilalanin dikonversi menjadi asam sinamat (C6‐C3). Pada kondisi tanaman mengalami penyerangan
oleh jamur, bakteri, atau virus, tanaman akan memproduksi asam
salisilat (C6‐C1) sebagai senjata pertahanan. Untuk menyintesis
asam salisilat, asam sinamat dikonversi terlebih dahulu menjadi
asam benzoat. Enzim asam benzoat 2‐hidroksilase kemudian
mengatalisi perubahan asam benzoat menjadi asam salisilat
(Gambar 2.2) (Cronizer et al. 2006).
Pada kondisi normal, asam sinamat diubah menjadi asam p‐
koumarat (C6‐C3, Gambar 1.5a) atau p‐koumaroil‐CoA dengan
bantuan enzim sinamat 4‐hidroksilase atau p‐koumarat:CoA
ligase. Asam p‐koumarat kemudian dikonversi menjadi asam kafeat (C6‐C3). Pada awalnya, diketahui bahwa asam kafeat
merupakan prekursor langsung untuk sintesis asam 5‐O‐
kafeoilquinat yang banyak terdapat di buah dan sayuran. Namun
hasil penelitian di bidang biologi molekular terbaru menyebutkan bahwa rute utama dalam sintesis senyawa tersebut adalah melalui p‐koumaroil‐CoA (Gambar 2.2) (Cronizer et al. 2006).
Asam kafeat yang didapat dari hasil konversi asam p‐
koumarat kemudian diubah menjadi asam ferulat (C6‐C3) dengan
bantuan enzim asam kafeat/5‐hidroksiferulat O‐metiltransferase.
Asam ferulat yang terbentuk dapat diubah menjadi asam sinapat
(C6‐C3) melalui produk antara 5‐hidroksiferulat (Gambar 2.2).
Kedua asam tersebut, ferulat dan sinapat, merupakan prekursor
untuk sintesis lignin (Cronizer et al. 2006).
©SEAFAST
Center
____________________________
FAL, fenilalanin am sinamat 4‐hidroksila KOMT‐1, asam caffe hidroksilase; ACoAC.
Gambar 2.2
___________Senyawa F
monia‐liase; AB2H, a se; GT, galloiltransf eat/5‐hydroksiferulat
2. Biosintesis feno
Fenolik pada Sayura
asam benzoat 2‐hid erase; 4CL, p‐kouma O‐metiltransferase;
olik (Cronizer et al
an Indigenous
droksilase; S4H,
arat:CoA ligase; F5H, ferulat 5‐
l. 2006).
©SEAFAST
Center
Seny
SS ka fl h 4‐ re
Gam
Flav
yawa Fenolik pada S
S, stilben sintase; alkon isomerase; avanon 3‐hidroksi idroksilase; IOMT,
‐reduktase; LDOX, eduktase; ANR, anto
mbar 2.3. Biosinte 2006). vonoid dan Stilb
Sayuran Indigenou
KS, kalkon sintas IFS, isoflavon sinta ilase; FLS, flavono isoflavon‐Ometiltr leukosianidin deok osianidin reductase esis flavonoid d
en
us _____________________
se; KR, kalkon red ase; FNS, flavon s ol sintase; F3’H, ansferase; DFR, dih ksigenase; LAR, leu e.
an stilben (Cron
________________
duktase; CHI,
sintase; F3H, flavonol 3’‐ hidroflavonol
kosianidin 4‐
nizer et al.
©SEAFAST
Center
_____________________________________Senyawa Fenolik pada Sayuran Indigenous
Flavonoid dan stilben memiliki jalur yang sedikit berbeda
dari senyawa fenolik lainnya. Jika senyawa fenolik lain hanya
diawali dari jalur shikimate, maka pada biosintesis flavonoid dan stilben melibatkan juga produk dari jalur asam malonat (Gambar
2.1). Flavonoid (Gambar 1.7) memiliki dua cincin benzen yang
dihubungkan oleh tiga buah atom karbon (C6‐C3‐C6). Cincin
benzen B dan jembatan C3 pada flavonoid berasal dari p‐koumaril‐
CoA yang merupakan produk turunan dari asam 3‐dehidrosikimat
dari jalur shikimate (Gambar 2.1). Produk dari jalur malonat, yaitu
malonil‐CoA, digunakan sebagai cincin A pada flavonoid. Seperti
halnya flavonoid, stilben juga merupakan produk hasil kondensasi p‐koumaril‐CoA dan malonil‐CoA (Cronizer et al. 2006).
Malonil‐CoA dari jalur asam malonat didapat dari asetil‐CoA.
Perubahan asetil‐CoA menjadi malonil‐CoA dibantu oleh enzim
asetil‐CoA karboksilase. Sebenarnya asetil‐CoA dapat terbentuk di
berbagai bagaian sel tumbuhan, seperti mitokondria, plastid,
peroksisom, dan sitosol. Namun demikian, asetil‐CoA untuk
sintesis malonil‐CoA pada biosintesis flavonoid dan stilben
terbentuk di sitosol. Di sitosol, enzim ATP‐sitrat liase mengonversi sitrat, ATP, dan Co‐A menjadi asetil‐CoA, oksaloasetat, ADP, dan inorganik fosfat (Davies dan Schwinn 2006).
A. Flavonoid
Gambar 2.3 memperlihatkan tahapan‐tahapan biosintesis
flavonoid. Kalkon sintase merupakan enzim yang mengatalis reaksi
antara 1 molekul p‐koumaril‐CoA dengan 3 molekul malonil‐CoA
membentuk narigenin‐kalkon. Pada beberapa tanaman, meskipun
sangat jarang, malonil‐CoA yang digunakan untuk menyintesis
kalkon ini dapat digantikan oleh kafeoil‐CoA atau feruloil‐CoA
©SEAFAST
Center
Senyawa Fenolik pada Sayuran Indigenous ____________________________________
(Davies dan Schwinn 2006). Jika enzim kalkon sintase berinteraksi
dengan enzim kalkon reduktase, maka produk yang dihasilkan
adalah 2’‐isoliquiritigenin. Perbedaan antara kedua senyawa
tersebut (narigenin‐kalkon dan 2’‐isoliquiritigenin) yaitu tidak
terdapatnya gugus 2’‐hidroksi pada 2’‐isoliquiritigenin.
Narigenin‐kalkon dikonversi menjadi narigenin oleh enzim
kalkon isomerase (CHI) tipe 1. Terdapat dua jenis enzim kalkon
isomerase (CHI) pada tanaman, yaitu CHI tipe 1 dan CHI tipe 2. CHI
tipe 1 adalah enzim yang terdapat di semua jenis tanaman, baik itu kacang‐kacangan maupun bukan kacang‐kacangan. Enzim ini hanya
dapat mengonversi narigenin‐kalkon menjadi narigenin (flavanon,
C15). Sebaliknya, enzim CHI tipe 2 yang hanya terdapat di tanaman kacang‐kacangan tidak hanya dapat mengonversi narigenin‐kalkon
menjadi narigenin, namun juga dapat mengonversi 2’‐
isoliquiritigenin menjadi liquiritigenin (flavon, C15) (Gambar 2.3)
(Cronizer et al. 2006).
Flavanon naringenin dan flavon liquiritigenin merupakan
bahan baku untuk memproduksi falvonoid dari golongan isoflavon.
Narigenin dan liquiritigenin berturut‐turut dikonversi menjadi
isoflavon genistein dan daidzein oleh enzim flavon sintase. Metilasi pada atom karbon nomor 7 dan atom oksigen yang terikat
di karbon nomor 4’ (cara penomoran dapat dilihat pada Gambar
1.7c) pada daidzein menghasilkan senyawa isofalvon lainnya,
secara berturut‐turut yaitu isoformonetin dan formononetin.
Reaksi metilasi tersebut dikatalisis oleh enzim isoflavon‐
Ometiltransferase (Gambar 2.3) (Cronizer et al. 2006).
Reaksi oksidasi terhadap naringenin dapat menghasilkan
senyawa flavonoid dari golongan flavon. Reaksi ini dibantu dengan
enzim flavon sintase dan membutuhkan hadirnya NADPH serta
©SEAFAST
Center
_____________________________________Senyawa Fenolik pada Sayuran Indigenous
oksigen. Senyawa flavon yang dihasilkan yaitu apigenin (Gambar
2.3) (Cronizer et al. 2006).
Selain menjadi substrat untuk senyawa dari golongan
isoflavon dan flavon, naringenin juga digunakan sebagai subtrat
untuk membetuk senyawa dari golongan dihidroflavonol/
flavanonol. Enzim flavanon 3‐hydroksilase membantu terjadinya reaksi hidroksilasi naringenin pada atom karbon nomor 3
membentuk senyawa dihidrokaemferol (flavanonol). Hidroksilasi
lebih lanjut terhadap dihidrokaemferol pada atom C nomor 3’ yang dikatalisis oleh enzim flavonol 3’‐hidroksilase menghasilkan
senyawa dihidroquersetin (flavanonol). Selain dihidroksilasi,
dihidrokaemferol juga dapat dikonversi menjadi kaemferol yang
merupakan senyawa dari golongan flavonol melalui pembentukan
ikatan rangkap dengan katalis enzim flavonol sintase (Gambar 2.3)
(Cronizer et al. 2006).
Senyawa flavonoid dari golongan leukosianidin dibentuk dari
dihidroquersetin dengan katalis enzim dihidroflavonol 4‐
reduktase. Golongan leukosianidin ini merupakan senyawa penting
pada tanaman dalam memproduksi tanin terkondensasi. Enzim
leukosianidin 4‐reduktase secara langsung mengatalisis
perubahan leukosianidin menjadi (+)‐katekin (golongan flavan‐
3ol). Isomer (+)‐katekin, yaitu (‐)‐epikatekin, merupakan senyawa
dari golongan flavan‐3ol yang banyak terdapat di tanaman,
terutama teh. Berbeda dengan isomernya, (‐)‐epikatekin tidak
secara langsung terbentuk dari leukosianidin. Pertama, dengan
katalis enzim leukosianidin deoksigenase leukosianidin dikonversi
terlebih dahulu menjadi sianidin (golongan antosianidin). Sianidin
inilah yang kemudian diubah menjadi (‐)‐epikatekin oleh enzim
antosianidin reduktase. Tanin terkondensasi didapat dari hasil
polimerasi leukosianidin dan flavan‐3ol (Cronizer et al. 2006).
©SEAFAST
Center
Senyawa Fenolik pada Sayuran Indigenous ____________________________________
Enzim‐enzim yang terlibat dalam biosintesis flavonoid pada
umumnya merupakan enzim yang berlokasi di sitosol tanaman.
Produk akhir dari biosintesis flavonoid tersebut kemudian diangkut
menuju subselular atau extraselular. Flavonoid yang terlibat
sebagai pigmen dalam tanaman kebanyakan diangkut menuju
vakuola (Davies dan Schwinn 2006).
B. Stilben
Seperti halnya flavonoid, stilben juga disintesis dengan
menggunakan satu molekul p‐komaroil‐CoA dan tiga molekul
malonil‐CoA (Gambar 2.3). Jika dalam sintetis flavonoid enzim yang berperan dalam mengondensasi keempat molekul tersebut adalah enzim kalkon sintase, maka pada biosintesis stilben, enzim yang digunakan adalah stilben sintase. Enzim stilben sintase dan kalkon
sintase memiliki struktur kimia yang sangat mirip sehingga
dipercaya kedua enzim tersebut merupakan enzim dari golongan
yang sama, yaitu enzim poliketida. Perbedaan pada kedua enzim
tersebut adalah enzim kalkon sintase selalu terdapat di jaringan
tanaman, sebaliknya enzim stilben sintase baru terbentuk jika
terinduksi oleh tekanan lingkungan. Tekanan lingkungan yang
dimaksud seperti radiasi sinar UV, trauma, dan infeksi (Cronizer et
al. 2006).
©SEAFAST
Center