PETA PAUTAN GENETIK MARKA RAPD DAN ANALISIS QTL
KELAPA SAWIT MENGGUNAKAN POPULASI SILANG BALIK
GENERASI PERTAMA MENUJU PERBAIKAN KUALITAS MINYAK
Oleh
EDY IRWANSYAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
EDY IRWANSYAH. Peta Pautan Genetik Marka RAPD dan Analisis QTL Kelapa Sawit Men99unakan Popu\asi Stan9 BaKkGene~asi :\-ma Menuju Pehaikan k a ! Nnyak Dibawah bimbingan SUDARSONO, HAJRIAL ASWIDINNOOR, M ACHMAD
CHOZIN, dan D W ASMONO.
lndikator utama rninyak kelapa sawit bermutu baik adalah tingginya kandungan asam oleat. Adanya peningkatan jumlah penduduk dunia, peningkatan perhatian konsumen terhadap nutrisi esensial produk pangan (khususnya minyak goreng), dan lebih 90% produk kelapa sawit digunakan untuk pangan; merupakan faktor pendorong perlu dilakukannya perbaikan kualitas minyak kelapa sawit.
Penelitian ini bertujuan: (1) mendapatkan data karakteristik komponen hasil dan kualitas minyak kelapa sawit populasi BC, (eks Brazil); khususnya untuk menyeleksi pohon sebagai tetua donor BC,, (2) mendapatkan peta pautan tetua donor dan pemulih; untuk memetakan marka molekuler yang terpaut dan sebagai prasyarat analisis QTL, (3) rnendapatkan peta QTL komponen kualitas minyak; guna mengetahui lokasi dan pengaruh lokus yang mengendalikan karakter kualitas minyak.
Upaya perbaikan kualitas minyak kelapa sawit dapat dilakukan melalui program jangka panjang melalui silang balik (teknik pemulian konvensional) yang berkelanjutan dan program jangka pendek melalui seleksi dengan bantuan marka molekuler. Kombinasi teknik pemuliaan konvensional dengan bantuan marka molekuler merupakan kunci penting dalam upaya percepatan mendapatkan kelapa sawit yang memiliki mutu rninyak tinggi sekaligus produksi tinggi.
Dari keragaan populasi BC,, berdasarkan analisis lintas dan regresi bertatar, peubah berat tandan dan persentase minyak per tandan memiliki pengaruh langsung paling besar dalam menentukan besarnya hasil minyak per tandan (komponen hasil). Sedangkan peubah miristat (C-14), palmitat (C-16), stearat (C-18), linoleat (C-18:2), dan linolenat (C-18:3) memiliki pengaruh paling besar dalam menentukan besamya kandungan asam oleat (komponen kualitas minyak). Guna pengembangan program pemuliaan untuk menghasilkan kelapa sawit yang memiliki kandungan oleat tinggi, terpilih pohon 93-107-33 T sebagai tetua donor untuk menghasilkan populasi BC,.
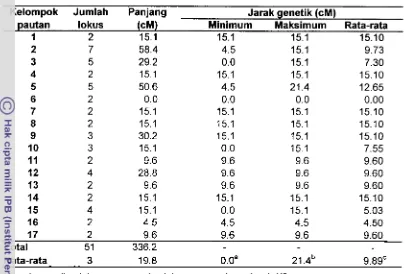
Dan 165 primer acak 10-mer yang ditapis pada kedua tetua BC,, diperoleh 108 yang polimorfis dan menghasilkan 600 lokus: 314 polimorfis (tetua donor= 195, tetua pemulih= 119) dan 286 monomorfis. Dengan uji chi kuadrat (P>0.05), lokus yang bersegregasi 1:1, untuk tetua donor=141, tetua pemulih=53. Peta pautan dikonstruksi menggunakan perangkat lunak MAPMAKEREXP 3.0b: peta tetua donor (LOD 3.0, 0=0.25)=7 kelompok pautan (KP), terdiri dari 51 lokus (336.2 cM), peta tetua pemulih=4 KP (LOD 1.69,0=0.25), terdiri dari 14 l o b s (282.9 cM).
ABSTRACT
EDY IRWANSYAH. Genetic Linkage Maps sf W D Markers and QTL Analysis of Oil Palm Using Population of First Generation of Backcross Toward Oil Quality Improve- ment. Under s u p e ~ s o r : SUDARSONO, HAJRIAL ASWIDINNOOR, M ACHMAD CHOZIN, and DWI ASMONO.
The main indicator that oil palm highly quality is highly oleic acid content. There are increasing world people number, increasing of consumer attention to essential nu- trition of food product (specific oil food), and more 90% product of oil palm used for food; a s factor which determining for improvement palm oil quality.
Objectives this research: (1) to get the characteristic data of yield and quality components of oil palm from the BC, (ex Brazil) population; specific for selected donor parent BC,, (2) to get the linkage map of donor and recurrent parent; for mapping linked molecular marker and as requirement of QTL analysis, (3) to get the QTL map- ping of oil quality component for knowing location and effect locus which controlling oil quality components.
Effort of improvement oil palm quality can conducted via long term program via backcrossing (conventional breeding technique) that continuing and short term program via marker-assisted selection. Combination of the both technique, conven- tional breeding with marker-assisted selection a s important key for acceleration effort to get palm oil that have oil quality and production highly.
Based on path analysis and stepwise regression, performance of BC, popula- tion on bunch weight and percentage of oil per bunch variable that have bigger direct effect for determining yield of oil per bunch content (yield component). Otherwise, miristic (C- 14), palmitic (C- 16), stearic (C-18), linoleic (C-18:2), and linolenic (C- 18:3) variable that have bigger effect for determining oleic acid content (oil quality compo- nent). For breeding program development to yielding oil palm that have oleic content highly, selected tree 93-107-33 T a s donor parent to generate BC, population.
From 165 random primers 10-mer was screened to both parent BC,, yielded 108 polymorphic (total 600 locus): 314 polymorphic (donor parent= 195, recurrent parent= 119) and 286 monornorphic. With chi square test (P>0.05), segregating locus that have 1 : 1, for donor parent= 141, recurrent parent=53. Linkage map were con- structed using MAPMAKERIEXP 3.0b software: donor parent map (LOD 3.0,8=0.25)= 7 linkage groups (LG), consist of 51 loci (336.2 cM), recurrent parent map=4 KP (LOD 1.69,8=0.25), consist of 14 loci (282.9 cM).
SURAT PERNYATAAN
Saya menyatakan dengan sebenar-benarnya, bahwa segala pernyataan dalam
disertasi yang berjudul:
PETA PAUTAN GENETIK MARKA RAPD DAN ANALISIS QTL KELAPA SAWIT MENGGUNAKAN POPULASI SILANG BALIK GENERASI PERTAMA MENUJU PERBAIKAN KUALITAS MINYAK
adalah gagasan atau hasil penelitian disertasi saya sendiri dengan bimbingan
komisi pembimbing, kecuali yang dengan jelas ditunjukkan rujukannya. Disertasi
ini belum pernah diajukan untuk memperoleh gelar pada program sejenis di
perguruan tinggi lain.
Semua data dan informasi yang digunakan telah dinyatakan dengan jelas dan
dapat diperiksa kebenarannya.
EDY I ~ ~ A N S Y A H
PETA PAUTAN GENETIK MARKA RAPD DAN ANALISIS QTL
KELAPA SAWIT MENGGUNAKAN POPULASI SILANG BALIK
GENERASI PERTAMA MENUJU PERBAIKAN KUALITAS MINYAK
Oleh
EDY IRWANSYAH
Disertasi
sebagai salah satu syarat untuk memperoleh
Gelar Doktor
pada Sekolah Pascasarjana lnstitut Pertanian Bogor
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Peta Pautan Genetik Marka RAPD dan Analisis
QTL
Kelapa Sawit Menggunakan Populasi Silang Balik Generasi Pertama Menuju Perbaikan Kualitas Minyak
Nama : Edy Irwansyah
Nomor Pokok : 995043
Program Studi : Agronomi/Biologi Molekuler Tanaman
Menyetujui:
1. Komisi Pembimbing
Dr Ir Sudarsono, MSc
Ketua
\
Dr Ir Hairial Aswidinnoor, MSc Prof Dr Ir M hchmad Chozin, MAnr
m g o t a +'"egota
Dr Ir Dwi Asmono, MS Anggota
EDY
IRWANSYAH
adalah putra pertama dari ayah Abdul Rivai dan ibu RismiahPohan, dilahirkan pada tanggal 7 September 1969 di Marbau Kabupaten
Labuhan Batu, Sumatera Utara. Pada tanggal 3 Agustus 2003 menikah dengan
Feriyanti.
Pendidikan dasar dan menengah diselesaikan di Kabupaten Labuhan
Batu, Sumatera Utara; yaitu Sekolah Dasar Negeri di Marbau (1983), Sekolah
Menengah Tngkat Pertama Negeri di Marbau (1986), dan Sekolah Menengah
Tngkat Atas Negeri 2 di Rantau Prapat (1989).
Gelar sarjana pertanian (S,) diperoleh dari Fakultas Pertanian Uurusan
Agronomi) Universitas Islam Sumatera Utara di Medan (1993), gelar magister
sains (S,) dari Program Pascasarjana (sekarang Sekolah Pascasarjana), Pro-
gram Studi Agronomi/Biologi Molekuler Tanaman, Institut Pertanian Bogor di
Bogor (1999). Pada tahun yang sama melanjutkan studi pada jenjang doktor
(S,) pada Program Studi Agronomi/Biologi Molekuler Tanaman, juga di Sekolah
Pascasarjana Institut Pertanian Bogor.
Sejak Nopember 1995 sampai sekarang, penulis menjadi staf pengajar
tetap Yayasan Universitas Islam Sumatera Utara pada Fakultas Pertanian di
PRAKATA
Dengan segala puji dan rasa syukur, penulis sampaikan kepada Allah
SW, atas keridoan, rahmat, dan karunia-Nya disertasi ini dapat diselesaikan.
Penelitian yang telah dilaksanakan dalam rentang waktu 17 bulan (Januari 2002
sampai Mei 2003), berjudul: "Peta Pautan Genetik Marka RAPD dan Analisis
QTL Kelapa Sawit Menggunakan Populasi Silang Balik Generasi Pertama Menuju
Perbaikan Kualitas Minyak.
Disertasi ini terdiri dari 7 bab. Bab-V merupakan pengembangan dari
naskah artikel yang telah dipublikasikan ke jurnal ilmiah dan publikasi poster.
Artikel yang telah dipublikasikan (J Penel Pertanian (22) 1 : 1-7) berjudul: "Analisis
Awal Pemetaan Genetik Kelapa Sawit pada Populasi Silang Balik Generasi
Pertama Hasil Persilangan Elaeis oleifera (Eks Brazil) dengan E. guineensis".
Poster yang telah dipublikasikan (International Seminar on Biotechnology for
Sustainable Agriculture: State of The Art of Research And Product Commer-
cialization. Bogor, Indonesia, 7-8 October 2003), berjudul: "Oil Palm Genetic
Linkage Maps Based on RAPD Markers Using Pseudo-Testcross Strategy". Bab-
VII merupakan pembahasan dan kesimpulam umum, juga merupakan opini
dalam upaya perbaikan mutu minyak kelapa sawit.
Bantuan dan dukungan dari berbagai pihak berupa dana, tenaga, pikiran,
dan doa serta lainnya yang bersifat material maupun immaterial merupakan
karunia Allah S W sebagai wujud dari rahrnat dan hidayat-Nya. Untuk itu, dalam
kesempatan ini penulis menyampaikan terima kasih dan penghargaan yang
setinggi-tingginya kepada:
1. Dr
Ir
Sudarsono, MSc yang bertindak selaku ketua komisi pembimbing; DrIr
Hajrial Aswidinnoor, MSc; Prof Dr Ir M Achmad Chozin, MAgr; dan Dr IrDwi Asmono, MS masing-masing bertindak selaku anggota komisi
hingga selesainya disertasi ini. Banyak pengalaman dan ilmu berharga yang
telah penulis peroleh sejak dari perkuliahan sampai selesainya disertasi ini
ditulis. Bimbingan, motivasi, saran, dan doa yang mereka berikan sangat
membantu penajaman analisis, sintesis, dan sistematika berpikir dalam
penyelesaian disertasi ini.
2. Dr Z Poeloengan (Direktur Pusat Penelitian Kelapa Sawit, PPKS), atas izinnya
dapat melakukan penelitian di Laboratorium Biologi Molekuler Balai
Penelitian Marihat Pematang Siantar dan Laboratorium Oleo Pangan milik
PPKS.
3. Ir Ridwan A Lubis (Mantan Kepala Balai Penelitian Marihat, 2003), Drs Gale
Ginting (GM Unit Produksi Marihat); atas izinnya melakukan penelitian di
Laboratorium Biologi Molekuler dan kebun percobaan, juga fasilitas mess
untuk tempat tinggal selama melakukan penelitian.
4. Dr A Razak Purba; Dr Iman Yani Harahap; H Subarjo; Ir Dira P
Komalaningtyas; Ir Amir Purba, MS; Ir Edy Suprianto;
Ir
Yurna Yenni; IrNajib Fauzi; Dr Sudharto PS; Dr Agus Susanto; Dr Sjafrul Latif, Dr Condro
Utomo dan seluruh Staf Balai Penelitian Marihat lainnya, atas fasilitas-
fasilitasnya, juga motivasi, saran, dan doa yang diberikan.
5. Ir Yusran Pangaribuan, MSi; atas izinnya memakai beberapa fasilitas yang
ada di Laboratorium Kimia dan Ir Indra Eko Setiyo, MSi; atas izinnya
memakai fasilitas perpustakaan dan internet, di Balai Penelitian Marihat.
6. Dr li Haryati, atas izimya untuk melakukan analisis kandungan asarn lemak
dan beta-karoten. Pak Warnoto dan Ibu Sabarida, SSi; atas bantuannya
dalam melakukan analisis asam lemak dan beta-karoten.
7. Pak Sutrisno, Pak Juki, Pak Pangihutan Panggabean; atas bantuannya di
kebun percobaan. Pak Egenar Panggabean dan Pak Sainun Manurung;
atas bantuannya dalam analisis tandan kelapa sawit. Pak Tamsil Pohan atas
Ibu Krista Manalu dan Pak Yusmanto; atas bantuannya di Laboratorium
Biologi Molekuler. Pak Ngalimin dan Ibu Tumini atas pelayanan dan perhatian
yang baik selama tinggal di Mess Balai Penelitian Marihat. Rudianto,
Sukamto, Rahmat Lubis, Mislan serta semua karyawan tetap dan tidak tetap
di PPKS atas segala bantuannya baik langsung maupun tidak langsung.
8. Seluruh staf pengajar dan pegawai yang ada di lingkup Sekolah Pascasa jana
IPB, atas segala layanan dan bantuan yang telah diberikan.
9. Bapak Prof Usman Nasution (Rektor UISU), Bapak Prof Djohar Arifin Husin
(Pjs. Rektor UISU, 1999), Bapak Ir Zainal Abidin, MS (Mantan Dekan Fakultas
Pertanian UISU, 2000), Ibu Ir Markhaini, MS (Dekan Fakultas Pertanian UISU,
2000-2004); atas izin, dukungan, bantuan dana, juga motivasi dan doa
yang diberikan dalam menyelesaikan studi program doktor di IPB.
10. Seluruh staf pengajar dan pegawai Fakultas Pertanian UISU, atas segala
layanan dan bantuan yang telah diberikan.
1 1. Ayahanda, Ibunda, adik-adik tercinta, bapak dan ibu mertua, khususnya
istri tercinta Feriyanti, SSi; atas segala dukungan, semangat, doa, dan
bantuan materi maupun moril sehingga disertasi ini dapat diselesaikan.
12. Dr Memen Surachman; Dr Catur Herison; Dr Rustikawati; Ir Dwi Hapsoro,
MSc; Ir Yusnita, MSc; Ir Eliyanti, MSi; Ir Zuyasna, MSc; Ir Ahmad Riduan,
MSi; Ir Soaloon Sinaga, MSi; Ir Ilyas Marzuki, MSi; Ir Sri Wening, MSi; dan
sahabat-sahabat lainnya, atas diskusi, saran, dan bantuan lainnya.
13. Bapak Dr Ir Dwi Asmono, MS; selain sebagai anggota komisi pembimbing,
beliau juga telah memfasilitasi dana penelitian yang cukup besar yang beliau
sisihkan sebagian dari dana proyek Riset Unggulan Terpadu (RUT), APBN,
dan INCO-DEV (EEC) pada Pusat Penelitian Kelapa Sawit.
Terimakasih, semoga disertasi ini bermanfaat.
Bogor, 10 Mei 2004
DAFTAR
IS1
Halaman
DAFTAR TABEL
...
xiv...
DAFTAR GAMBAR xv
DAFTAR LAMPIRAN
...
xvi...
DAFTAR SING KATAN xvii
BAB-I: PENDAHULUAN
...
1...
Latar Belakang 1
Tujuan
...
4...
Kegunaan 4
BAB-11: TINJAUAN PUSTAKA
...
6...
Kelapa Sawit 6
1
.
Asal...
2.
Botani...
...
.
3 Syarat Tumbuh
...
.
4 Pemuliaan
...
.
5 Produksi dan Kualitas Minyak
...
Marka Molekuler
...
.
1 Potensi
2
.
RAPD...
3
.
Aplikasi RAPD pada Tanaman Kelapa Sawit...
Peta Pautan Genetik dan
Quantitative
TraitLoci
(QTL)...
MAS-BC pada Tanaman Kelapa Sawit
...
BAB-111: KERAGAAN KOMPONEN HASIL
DAN
KUALlTASMlNYAK
KELAPASAWT DAM POPULASI SILANG BALlK GENERASI PERTAMA HASIL
PERSILANGAN HIBRIDA
Elaeis oleifera
(EKS BRAZIL)X
E.
guineensis
DENGAN E.
guineensis
...
Abstrak
...
...
Abstract
PENDAHULUAN
...
...
B MDAN METODE
...
Bahan Tanaman...
Pengamatan...
Komponen Hasil...
Komponen Kualitas Minyak
...
Analisis Data
...
HASIL DAN PEMBAHASAN
Keragaan Komponen Hasil dan Kualitas Minyak
...
...
Keragaan Komponen Hasil
Halaman
...
KESIMPUIAN DAN SARAN
BAB-IV: OPTlMASl PROSEDUR EKSTRAKSI DNA DARI DAUN DAN
AIYALISIS RAPD KELAPA SAWT
...
...
Abstrak...
Abstract...
PENDAHULUAN...
BAtlAN DAN METODE
Ekstraksi DNA dari Daun Kelapa Sawit
...
...
Bahan Tanaman...
Optimasi Prosedur...
Pengamatan...
Analisis RAPD Kelapa Sawit
Sampel DNA
...
...
Optimasi Prosedur
...
Pengamatan
...
HASIL DAN PEMBAHASAN
...
Ekstraksi DNA...
Analisis RAPD...
KESIMPUIANBAB-V: KONSTRUKSI PETA PAUTAN GENETIK KELAPA SAWIT MENGGUNAKAN POPULASI SIIANG BALIK GENERASI PERTAMA
HASlL PERSIIANGAN HIBRIDA Elaeis oleifera (EKS BRAZIL) X E
.
...
guineensis DENGAN E
.
guineensis...
Abstrak
...
Abstract
PENDAHULUAN
...
...
BAHAN DAN METODE
...
Bahan Tanaman
...
Ekstraksi
DNA
...
Arnplifikasi DNA
...
Penapisan dan Pemilihan
Ptirner
Analisis Segregasi Marka RAPD dan Pernilihan Lokus
...
Konstruksi Peta Pautan
...
...
HASIL DAN PEMBAHASAN
KESIMPUIAN DAN SARAN
...
BAB-VI: ANALISIS QTL KOMPONEN HASIL DAN K W A S
MINYAK
KELAPA SAWIT DARI POPULASI SILANG BALIK GENERASI
...
PERTAMA (EKS BRAZIL)
Halaman
...
BAHAN DAN METODE
...
Bahan Tanaman
Data Kuantitatif Komponen Hasil dan Kualitas Minyak
...
...
Ekstraksi DNA dan Analisis RAPD
...
Penapisan dan Pemilihan Primer
Analisis Segregasi Marka RAPD dan Pemilihan Lokus
...
Konstruksi Peta Pautan
...
...
Identifikasi QTL
HASIL DAN PEMBAHASAN
...
QTL Komponen Hasil...
...
QTL Komponen Kualitas Minyak
KESIMPULW DAN SARAN
...
BAB-VII: UPAYA MENUJU PERBAIKAN KUALITAS MINYAK K E W A SAWIT
...
Abstrak
Abstract
...
...
PENDAHULUAN
...
SIIANG BALIK GENERASI KEDUA
MARKER ASSISTED BREEDING
...
Konversi ke Marka Molekuler Spesifik
...
Pemetaan QTL Berbasiskan Peta Pautan Genetik Berkerapatan...
Tinggi
...
REKAYASA GENETIKA
...
KESIMPULW
...
D m A R PUSTAKA
DAFTAR TABEL
Nomor J u d u l Halaman
[image:14.570.85.493.90.717.2]1 Kandungan asam lemak dan beta-karoten E. oleifera, E. guineensis,
...
dan hibridanya
2 Keragaan sepuluh peubah komponen hasil dari 23 progeni BC, dan
...
kedua tetuanya
3 Keragaan tujuh peubah komponen kualitas minyak dari 23 progeni
...
BC, dan kedua tetuanya
4 Matriks koefisien korelasi antara sepuluh peubah komponen hasil
...
kelapa sawit dari 23 progeni BC,
5 Matriks pengaruh langsung (koefisien lintas) dan pengaruh tidak
langsung dari sembilan peubah komponen hasil terhadap hasil minyak per tandan
...
6 Matriks koefisien korelasi antara tujuh peubah komponen kualitasminyak kelapa sawit dari 23 progeni BC,
...
7 Matriks pengaruh langsung (koefisien lintasan) dan pengaruh tidak
langsung dari enam peubah komponen hasil terhadap persentase
...
asam oleat
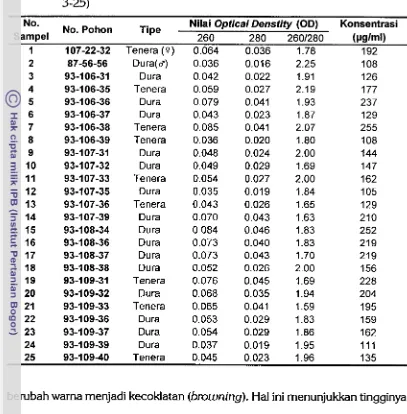
8 Nilai optical density (OD) 260,280,2601280 dan konsentrasi DNA
dua tetua populasi BC, (nomor 1 dan 2) serta 23 progeninya (nomor
3-25)
...
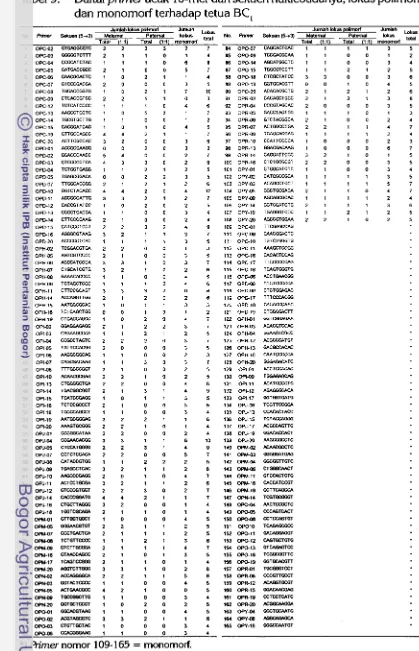
9 Daftar primer acak 10-mer dan sekuen nukleotidanya, lokus polimorf
...
dan monomorf terhadap tetua BC,
10 Karakteristik peta pautan tetua donor yang dikonstruksi dengan nilai
...
minimum LOD 3.0 dan fraksi rekombinasi maksimum 0.25
11 Karakteristik peta pautan tetua pemulih yang dikonstruksi dengan
...
nilai minimum LOD 1.69 dan fraksi rekombinasi maksimum 0.25 12 Karakteristik QTL komponen hasil dari peubah berat per buah,
persentase minyak per mesokarp, dan hasil minyak per tandan;
...
dideteksi dari 23 progeni BC,
13 Karakteristik QTL komponen kualitas minyak dari peubah persentase asam lemak dari: miristat, palmitat, stearat, oleat dan linolenat;
...
dideteksi dari 23 progeni BC,
14 Daftar sernbilan belas spesies tanaman, lokudnukleotida ID, dan
...
protein ID yang memiliki produk gen FAD2
Tabel Lampiran
1 Ringkasan analisis faktor tunggal (tipe TeneraIDura) terhadap sepuluh
...
peubah komponen hasil
2 Ringkasan analisis faktor tunggal (tipe Tenera/Dura) terhadap tujuh
...
peubah komponen kualitas minyak
3 Daftar lokus polimorfis dan hasil skoring terhadap dua tetua BC,
DAFTAR GAMBAR
Nomor J u d u i Halaman
...
1 Kerangka dan bagan alur penelitian
2 Keragaan E. oleifera. (A) E. oleifera (eks Brazil), tahun tanam 1940.
...
(B) E. oleifera (eks Suriname), tahun tanam 1952
...
3 Contoh kartu analisis tandan (individuil)
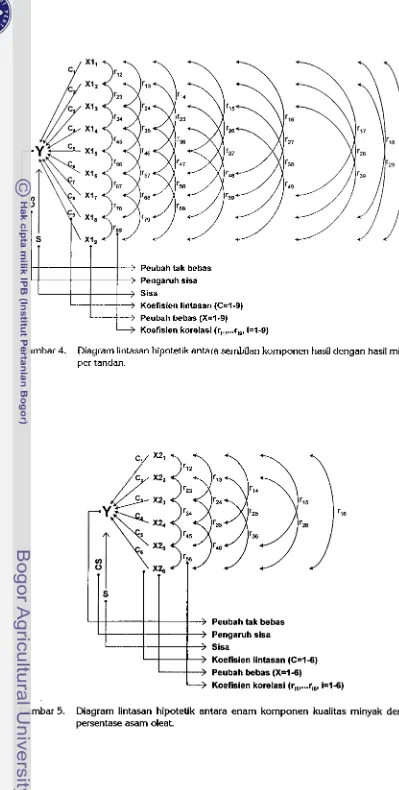
4 Diagram lintasan hipotetik antara sembilan komponen hasil dengan
...
persentase hasil minyak
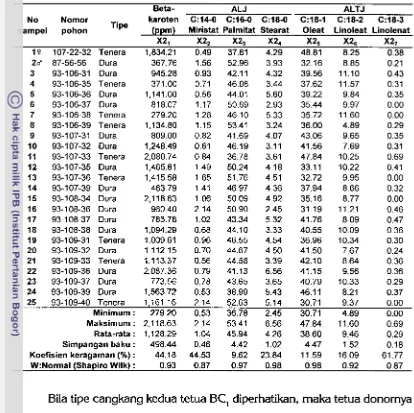
5 Diagram lintasan hipotetik antara enam komponen kualitas minyak
...
dengan persentase asam oleat
6 Diagram lintasan hipotetik dari komponen hasil antara berat tandan
(XI ,) dan persentase minyak per tandan (XI,) terhadap hasil minyak
...
per tandan (Y)
7 Diagram lintasan hipotetik dari komponen kualitas minyak antara
miristat
(a,),
palmitat (X2,), stearat (X2,), linoleat @2J, dan linolenat...
(X2,) terhadap oleat (Y)
8 Profil fragmen DNA dari dua tetua populasi BC, eks Brazil (lane 1
...
dan 2) serta 23 progeninya (lane 3-25)
9 Profil fragmen DNA hasil amplifikasi dengan primer OPJ-0 1 terhadap
dua tetua populasi BC, eks Brazil (lane 1 dan 2) serta 23 progeninya
...
(lane 3-25)
10 Peta pautan genetik tetua donor, dibentuk dengan nilai LOD mini-
...
mum 3.0 dan fraksi rekombinasi maksimum 0.25
1 1 Peta pautan genetik tetua pemulih, dibentuk dengan nilai LOD mini-
...
mum 1.69 dan fraksi rekombinasi maksimum 0.25
12 Peta pautan genetik dan plot LOD yang menunjukkan eksistensi QTL berasosiasi dengan peubah: (A) berat per buah pada KP-2, (B)
persentase minyak per mesokarp pada
KP-
10, (C) persentase minyakper mesokarp pada KP-12, dan @) hasil rninyak per tandan pada KP-12
13 Peta pautan genetik dan plot LOD yang menunjukkan eksistensi
QTL berasosiasi dengan peubah persentase asam lemak dari: (A)
miristat pada KP-2, (B) miristat pada KP-5, (C) palmitat pada KP-2,
(D) stearat pada KP-2,
(E)
oleat pada KP-2,(F)
oleat pada KP-5, (G)...
oleat pada KP-12, dan (H) linolenat pada KP-2
14 Profil fragmen DNA sampel hasil amplifikasi dengan primer OPC-
14. Lajur: M = marka 1
kb
DNA ladder, 1 = tetua donor BC, (mater-...
nal), 2 = tetua pemulih BC, (paternal), 3-25 = progeni BC,
15 Prakiraan disain primer untuk mengisolasi gen FAD2 pada kelapa sawit. (A) Sekuen asam amino conserve (parsial) dari 19 spesies
tanaman yang gen FAD2-nya telah diidentifikasi. (B) Disain primer
...
berdasarkan susunan asam amino (parsial) yang conserve
16 Hubungan phylogenetic sekuen asam amino yang menyandikan
...
DAFTAR
LAMPIRAN
Nomor J u d u l Halaman
...
1 Silsilah populasi silang balik generasi pertama
(BJ
4 1 S) 982 Prosedur Baku Pembuatan Larutan Kimia dan Ekstraksi DNA Daun
Kelapa Sawit
...
993 Prosedur Baku Analisis RAPD Kelapa Sawit
...
1064 Visualisasi fragmen DNA (23 progeni BC. dan kedua tetuanya) hasil
DAFTAR SINGKATAN AFLP A W ALTJ BC bp BSA CAPS CYY CIRAD CPO CTAB D dATP dCTP dGTP DNA dNTP dTTP FAD FIPS
FK
FP GCA IOPRI IRHO Jacq. KIAA KP LOD MAB MAS mdpl MRS: amplified fragment length polymorphism
: asam lemak jenuh
: asam lemak tak jenuh
: backcross
: base pair
: bulk segregant analysis
: cleaved amplified polymorphic sequence
: composite interval mapping
: Centre de cooperation internationale en recherche agronomique
pour le d6veloppement
: crude palm oil
: cetyl-trimethyl-ammoniumbromide
: Dura
: 2'-deoxyadenosine 5'-triphosphate
: 2'-deoxycytidine 5'-triphosphate
: 2'-deoxyguanosine 5'-triphosphate
: deoxyribonucleic acid
: 2'-deoxy any base 5'-triphosphate
: 2'-deoxythymidine 5'4riphosphate
: fatty acid desaturase
: Familiy and Individual Palm Selection
: faktor konversi
: faktor pengenceran
: general combining ability
: Indonesian Oil Palm Research Institute
: I'Institut de recherche pour les huiles et okagineux
: Jacquin (Nicolaus Josef von Jacquin)
: kloroform:isoamilalkohol
: kelompok pautan
: log of the odd
: marker-assisted backcrossing
: marker-assisted selection
: meter dari permukaan laut
: Marihat Research Station
OD P PCR PKO PPKS PTGS PVE PWP QTL RAPD RFLP RP RRS SCA SCAR SSR TAE Taq TE tn TPVE
UV
v
: optical density
: Pisifera
: polymerase chain reaction
: palm kernel oil
: pusat penelitian kelapa sawit
: post transcriptional gene silencing
: phenotypic variance explained
: polyvinilpolypyrrolidone
: quantitative trait loci
: random amplified polymorphic D M
: restriction fragment length polymorphisms
: recurrent parent
: reciprocal recurrent selection
: specific combining ability
: sequence-characterized amplified region
: simple sequence repeats
: [Trisl-[Acetic Acid Glacial]- [EDTA]
: Thermus aquaticus
: [Tris]
-
[EDTA]: tidak nyata
: total phenotypic variance explained
: ultra violet
BAB-I
PENDAHULUAN
Latar Belakang
Sampai saat ini, kelapa sawit (Elaeis guineensis Jacq.) merupakan
penghasil minyak nabati terbesar kedua (25%) di dunia setelah kedelai (29%)
(Casson 2003; Mayes et al. 2000). Walaupun demikian, dari segi produksi, kelapa
sawit memiliki produktivitas yang lebih tinggi dibandingkan dengan tanaman
penghasil minyak nabati lainnya (Pamin 1998), sepuluh kali lebih tinggi
dibandingkan dengan tanaman kedelai (Yuan dan Weng 1996).
lndonesia dikenal sebagai produsen minyak kelapa sawit terbesar kedua
setelah Malaysia. Pada 1996, produksi lndonesia sebesar 4.5 juta ton, sedangkan
Malaysia sebesar 8.4 juta ton. Kemudian, pada 2000 produksi Indonesia sebesar
6.9 juta ton dan Malaysia sebesar 10.8 juta ton.Walaupun produksi Malaysia
lebih besar dibandingkan Indonesia, tetapi pertumbuhan produksi lndonesia
lebih besar. Pamin (1998) memperkirakan, pada 2010 lndonesia telah menjadi
produsen minyak kelapa sawit terbesar pertama di dunia.
Seiring dengan meningkatnya jurnlah penduduk dunia, meningkat pula
kebutuhan penduduk terhadap minyak kelapa sawit dan derivatnya. Hal ini
disebabkan sebagian besar minyak kelapa sawit digunakan untuk pangan (90x1,
selebihnya non-pangan (Jalani 1998). Menyadari adanya trend pada tingkat
konsumen dalarn memperhatikan mutu minyak dan nutrisi esensial, maka dalam
usaha pemuliaan kelapa sawit, selain produksi, mutu minyak juga perlu
diperbaiki. Mutu minyak dinilai dari tinggi-rendahnya kandungan asam lemak
tak jenuh (ALTJ) dan komponen minor minyak kelapa sawit.
Upaya pemuliaan tanaman untuk merakit kelapa sawit yang memiliki
kualitas minyak tinggi tanpa mengorbankan produksi
crude palm
oil(CPO)
ke arah ini dimulai sejak 1978, yaitu dengan mempersilangkan E. oleifera X E.
guineensis, juga silang balik (backcross, BC) hibridanya dengan E. guineensis
(Lubis et al. 1994; Hutomo et al. 1995). E. oleifera, merupakan plasma nutfah
yang baik untuk perbaikan mutu minyak, ditunjukkan dengan lebih tingginya
komposisi kandungan ALTJ (70-83%) dibandingkan dengan E. guineensis (40-
60%) (Syukur dan Lubis 1985). Gen dari spesies liar dapat diintrogresikan ke
tanaman budidaya melalui persilangan kerabat jauh (interspecific hybn'diza-
tion), dilanjutkan dengan metoda silang balik (Chen dan Adelberg 2000). Dalam
ha1 ini, gen penentu ALTJ dan komponen minor minyak yang ada pada E.
oleifera (donor parent, tetua donor) dapat diintrogresikan ke dalam spesies
komersial, E. guineensis (recurrentparent, tetua pemulih) melalui metode silang
balik. Menurut Komalaningtyas et al. (1998) bahwa kemajuan seleksi melalui
silang balik konvensional kelapa sawit sangat lambat. Hal ini disebabkan berbagai
kendala, seperti siklus pemuliaan kelapa sawit yang lama (10-13 tahun) dan
adanya faktor sterilitas akibat persilangan dua spesies yang berkerabat jauh.
Ada dua strategi potensial untuk meningkatkan efisiensi perbaikan genetik
kelapa sawit, yaitu melalui marker assisted breeding (MAB) dan rekayasa
genetika (Mayes et al. 1993). MAE3 ini dapat dilakukan dengan menggabungkan
teknologi marka molekuler ke dalam program seleksi silang balik, lazimnya
disebut marker-assisted backrossing (MAS-BC). Melalui MAS-BC, hambatan
pautan (linkage drag) dapat dikurangi, mempercepat transfer blok genom tar-
get dari plasma nutfah spesies kerabat liar ke dalam background plasma nutfah
yang diinginkan (Forbes 2000), dan mampu mengurangi jurnlah generasi seleksi
(Hospital et al. 1992).
Metode MAS-BC memerlukan ketersediaan peta pautan genetik (genetic
linkage map), informasi tentang lokasi dan pengaruh gen yang berasosiasi
dengan sifat kuantitatif tertentu (quantitatiue trait loci,
QTL)
sebagai faktorKonstruksi peta pautan genetik berbasis marka molekuler dapat dilakukan
dengan strategi pseudo-testcross dua arah (Grattapaglia dan Sederoff 1994).
Dengan strategi ini akan didapatkan dua set peta, yaitu peta maternal dan pa-
ternal dari populasi pemetaan.
Masih sedikit laporan tentang pemetaan pautan genetik dan QTL yang
berasosiasi dengan komponen hasil, bahkan belum ada laporan untuk
komponen kualitas minyak kelapa sawit. Peta pautan genetik kelapa sawit
menggunakan marka restriction fragment length polymorphism (RFLP)
dilaporkan oleh Mayes et al. (1997) dan Rance et al. (2001), menggunakan
marka amplified fi-agment length polymorphism (AFLP) dan simple sequnce
repeats (SSR, atau microsatellite) oleh Rival et al. (200 1). Marka random am-
plified polymorphic DNA (RAPD) dan marka molekuler lainnya belum banyak
dilaporkan untuk pembuatan peta pautan genetik kelapa sawit. QTL kelapa
sawit yang berasosiasi dengan beberapa komponen hasil kelapa sawit dilaporkan
oleh Rance et al. (2001).
Meskipun hingga saat ini pemetaan QTL yang berasosiasi dengan kualitas
minyak kelapa sawit belum pernah dilaporkan, tetapi pemetaan QTL yang
berasosiasi dengan kualitas minyak pada tanaman selain kelapa sawit telah
banyak dilaporkan, antara lain pemetaan QTL yang berasosiasi dengan
konsentrasi asam lemak pada jagung (Goldman et al. 1994; Alrefai et al. 1995),
QTL
yang berasosiasi dengan asam oleat tinggi (Schierholt et al. 2000; Sharmaet al. 2002) atau asam linoleat pada Brassica
napus
(Somers et al. 1998), danQTL pengendali asam linolenat pada rapeseed (Jourdren et al. 1996a, 1996b).
Sedangkan model pemetaan QTL dengan populasi silang balik dan manfaat
penggunaan marker assisted selection (MAS) telah dikaji oleh beberapa grup
peneliti (Loriew et al. 1995; Ahmad 2000; Moreau et al. 2000; Reyes-Valdes
Teknik yang merupakan kombinasi seleksi berbasis marka molekuler
dengan teknik pemuliaan konvensional merupakan alat bantu strategis yang
dapat mempersingkat waktu seleksi. Dengan perkataan lain, ketersediaan dan
perkembangan marka molekuler yang semakin pesat dapat mempercepat
pencapaian tujuan program pemuliaan untuk menyediakan sumberdaya genetik
yang memiliki karakter unggul dalam waktu yang singkat. Peta pautan genetik
dianggap sebagai salah alat bantu untuk mencapai tujuan itu. Peta pautan
genetik dipersiapkan untuk analisis QTL kelapa sawit menuju perbaikan kualitas
minyak tanpa mengorbankan produksi CPO.
Tujuan
1. Mendapatkan data karakteristik komponen hasil dan kualitas minyak kelapa
sawit dari populasi BC,.
2. Mendapatkan peta pautan genetik kelapa sawit dari populasi pemetaan BC,.
3. Mengetahui lokus-lokus DNA yang berasosiasi dengan karakter komponen
hasil dan kualitas minyak.
4. Mengetahui marka-marka DNA spesifik untuk identifikasi karakter
komponen hasil dan asam lemak kelapa sawit.
Kegunaan
1 . Dasar pengembangan MAB pada kelapa sawit, sehingga pada akhirnya
diperoleh bahan tanaman yang memiliki kualitas minyak dan produksi CPO
tinggi dalam waktu relatif cepat dan dengan sumberdaya genetik yang tepat.
2. Marka QTL yang berasosiasi dengan karakter unggul kelapa sawit yang
konsisten memberikan efek genetik terbesar digunakan sebagai dasar isolasi
Marker-assisted backcrossing
I
Backcrossing
1
I
Kelapa sawitE. oleifera E. guineensis
Marker-assisted selection
I
E. guineensis
CPO
>
7.9 tonALTJ > 40%
t'
m
Marka molekuler
I
-
nailtan genetik
Verifikasi
L A
Alignment asam amino tanaman lair-
Sekuen QTL (+)
gen interest
~ s o s i a s i ~
-
dengan karakter'L
A
kinteresfr
u
L . Tanaman transgenik
I Rekayasa Genetik;
L
d
BAB-I1
TINJAUAN PUSTAKA
Kelapa Sawit
1. Asal
Tanaman kelapa sawit (E. guineensis Jacq.) berasal dari Afrika Barat,
ada beberapa bukti kuat yang mendukungnya. Di Abydos (3000 SM) ditemukan
lemak dalam kendi yang terkubur di makam diduga berasal dari kelapa sawit;
fosil polen mirip dengan polen kelapa sawit yang dipelihara dan berada di Afrika
Barat dari Miocene dan lapisan muda pada delta Niger, dan bukti lingustik yang
menyebutkan ditemukannya spesies pohon mirip kelapa sawit. E. oleifera atau
E. melanococca merupakan spesies kelapa sawit yang banyak tumbuh di
Amerika. Ada pendapat lain bahwa kelapa sawit berasal dari Amerika yang
kemudian dibawa ke Afrika. Ada dua alasan kelapa sawit berasal dari Amerika,
yaitu (1) palma tersebut tumbuh di area pantai Brazil, dan (2) seluruh genera
berasal dari Amerika. Ketika Columbus menemukan Amerika, diyakini bahwa
kelapa sawit sudah tumbuh di Amerika. Akan tetapi, tidak ada catatan otentik
tentang ha1 itu (Hartley 1988).
Tanaman kelapa sawit diintroduksi ke Indonesia pada 1848. Sebanyak
empat bibit kelapa sawit ditanam di Kebun Raya Bogor. Dari keempat bibit
tersebut, dua bibit diintroduksi dari Bourbon atau Mauritius pada Februari 1848,
dua bibit yang lain diintroduksi dari Amsterdam pada Maret 1848 (Pamin 1998).
2.
BotaniKelapa sawit yang mempunyai bahasa latin Elaeis guineensis Jacq.,
berasal dari bahasa Yunani, yaitu elaion yang berarti minyak, guineensis
menunjukkan bahwa tanaman kelapa sawit berasal dari Pantai Guinea,
kelapa sawit, nama lengkapnya Nicolaus Josef von Jacquin. Tanaman ini
tergolong pada kelompok monokotil.
Kelapa sawit akarnya terdiri dari primer, sekunder, tersier dan kuarter;
merupakan akar serabut yang sebagian besar berada dekat permukaan tanah
dengan kedalaman 15-30 cm. Batangnya tegak, tidak bercabang, berdiameter
40-75 cm, tinggi batang dalam pembudidayaan sekitar 15- 18 m. Daunnya
majemuk dengan pelepah daun tersusun melingkari batang berbentuk spiral.
Panjang pelepah daun mencapai 9 m, panjang helaian daun mencapai 1.2 m
yang jumlahnya 100-160 pasang. Untuk perkebunan kelapa sawit, jumlah
pelepah yang dipertahankan sekitar 30-50 pelepah (Hartley 1988).
Tpe pembungaan kelapa sawit adalah monoecious, berarti bunga jantan
dan betina terdapat pada satu tanaman, tetapi pada tandan yang berbeda. Rasio
bunga jantan terhadap betina dapat dipengaruhi keadaan iklim. Pada tanaman
yang mengalami masa kekeringan, bunga jantan mendominasi. Sementara itu
pada musim hujan bunga betina yang mendominasinya. Kadangkala dijumpai
bunga hermaprodit pada tanaman muda yang berumur sekitar 2-4 tahun.
Namun, bunga hermaprodit tersebut akan menyusut atau menghilang dengan
sendirinya sejalan dengan bertambahnya umur tanaman. Bunga tumbuh pada
setiap ketiak pelepah daun, satu tandan bunga berupa bunga jantan atau bunga
betina dengan masa siap polinasi yang berbeda, sehingga te jadi penyerbukan
silang. Buah, tersusun dalam satu tandan yang merupakan buah batu, terdiri
dari kulit buah, daging buah, cangkang, dan inti. Minyak, sebagian besar (20-
27%) terdapat pada bagian perikarp, yaitu pada kulit buah dan daging buah,
sementara itu pada bagian inti hanya mengandung minyak sekitar 4-6% (Hartley
1988).
Berdasarkan ketebalan cangkangnya, kelapa sawit dapat dibedakan atas
(a) Dura; ketebalan cangkang 2-8 mm, kandungan mesokarp tergolong rendah
sampai menengah (35-55x1, di bagian luar tidak terdapat lingkaran sabut.
(b) Tenera; ketebalan cangkang 0.5-4 mm, kandungan mesokarp tergolong
menengah sampai tinggi (60-96%), terdapat lingkaran sabut pada bagian
luarnya.
(c) Pisifera; ketebalan cangkang sangat tipis, bahkan tidak ada (Hartley 1988).
Ada perbedaan karakter morfologi yang cukup jelas antara E. oleifera
dan E. guineensis (Supriyanto et al. 2000). Perbedaan morfologi yang paling
nyata terletak pada keragaan batang dan daun tanaman. Pada E. oleifera,
pertumbuhan batang sangat lambat, dan umumnya laju pertumbuhannya hanya
setengah kali dari pertumbuhan batang E. guineensis. Bentuk daun E. oleifera
dicirikan dengan susunan anak daun yang berada dalam satu arah. Anak daun
pada E. oleifera cenderung lebih lebar dibandingkan dengan E. guineensis.
Sedangkan bentuk buahnya, E. oleifera umumnya lebih kecil dibandingkan
dengan
E.
guineensis, dan terdapat kecenderungan selalu membentuk buahpartenokarpi. Buah E. oleifera umumnya berwarna jingga terang ketika masak.
Pada E. oleifera belum pernah ditemukan buah jenis Pisifera. Gambar 2
memperlihatkan keragaan E. oleifera asal Brazil dan Suriname.
3. Syarat Tumbuh
Kelapa sawit tumbuh baik pada iklim tropis zone katulistiwa dengan tipe
iklim Af dan Am (menurut klasifikasi Koppen), dengan curah hujan minimal 9
bulan sebanyak 2000-3000 mm/tahun yang menyebar sepanjang tahun. Lahan
pertanaman mulai dari dataran rendah sampai pada ketinggian tidak lebih dari
600 mdpl dan sangat baik bila tanah tersebut bertopografi datar. Syarat tanah
untuk mendukung pertumbuhan tanaman adalah tanah yang subur bersolum
dalam, berdrainase baik, pH 5.5-7.0, yaitu tanah-tanah aluvial yang bertekstur
Gambar 2. Ke~agaan E. oleifera. (A) E. ow,- I
,-.-.
-.
-,, tahun tanam 1940. (B) E. old@(eks Suriname), ta hun tanam 1952.
4. Pemuliaan
Program
pemuliaan kelapa
sawitdl
Indonesia dimulai pada
1 9 1 0 ~ ,menggunakn
materialtanaman secara
terbatas dari
ernpat bilikdapa
&witindukvarietas
Durayang
ditanam
pertama
M
i
di
Kebun
RaysBogor pada
1848(Barnin 1998).
Bermula
pada
1970, MadhatR e s d
Station(MRS, sekarang
Balaiknelitian
Mari hat) mengintroduksjplasma nutfah baru
,
be
kerjasarnadeng
an
rlnstitut
de
& b epour
les
h u l k etoi&ginewc
(IRHO),
krancis.
Introduksitersebut
meliputi Dura IMmu dariLame; TenerWiera
Ivory Coast;Tenerd
Pisifera Yangambi dari Djonggo
deEala,
Isangui,dm
Yawenda;Tmera/Pisifera
Nifor
dariCalabar,
Angola, d mNigeria;
Tenerdhifera Yacobeue;
TeneraPisiFera
Dami
dari New Guinea;dan
polen E.oleifera
dari
SanAlberto
(Colombia).Populasi
program pemuliaan dari
1848- 195 1 d mintroduksi
bam, klahmenjadi
stitute (IOPRI; Pusat Penelitian Kelapa Sawit, PPKS) sampai sekarang (Pamin
1998).
Salah satu tujuan penting pemuliaan kelapa sawit di Indonesia adalah
membuat kultivar yang mempunyai hasil minyak tinggi. Berbagai observasi
menunjukkan bahwa vigor hibrida berasosiasi dengan hasil yang dapat dicapai
pada progeni F, dari persilangan antara dua grup heterosis yang independen,
Dura (D) dan Pisifera (P). Diantara metode seleksi yang ada untuk perbaikan
hasil minyak, metode reciprocal recurrent selection (RRS) yang dipilih sebagai
prosedur favorit, sebab metode ini memiliki kemampuan untuk meningkatkan
hasil minyak dari hibrida DxP dan mempertahankan keragaman genetik dari
masing-masing grup heterosis. PPKS telah menggunakan metode ini untuk
perbaikan hasil minyak sejak 1974 (Asmono et al. 1998).
Metode RRS adalah suatu skema yang sangat menarik, baik untuk pro-
gram pemuliaan maupun produksi benih dan klon kelapa sawit. Pertama,
pemilihan tetua untuk memproduksi hibrida komersial didasarkan atas pengujian
progeni, sehingga hanya hibrida-hibrida yang telah diuji disalurkan kepada
konsumen. Kedua, skema seleksi memungkinkan untuk mengeksploitasi
sesegera mungkin persilangan-persilangan terbaik dan perbaikannya dapat
dilakukan melalui selfing tetua terpilih sehingga daya gabung khusus (specific
combining ability, SCA) dapat dieksploitasi secara optimal. Ketiga, hibrida
komersial dapat direproduksi menggunakan berbagai tipe persilangan, Dura
diseleksi Dura dan TeneraPisifera diseleksi Tenera. Setelah berakhirnya siklus
seleksi, dimungkinkan untuk memproduksi benih dengan cara reproduksi secara
pasti persilangan-persilangan terbaik dari hasil pengujian dan menyilangkan
dua tetua yang memiliki daya gabung umum (general combining ability, GCA)
yang baik, meskipun persilangan tersebut belum diuji. Dengan menggunakan
tanaman unggul dari hasil pengujian, progeni dapat diperbanyak secara kultur
tanaman pada bagian titik tumbuh untuk kultur jaringan). Pemilihan tanaman
unggul dilakukan dengan mengeksploitasi keragaman di dalam famili di antara
famili-famili yang diuji pada pengujian progeni. Klon-klon yang dihasilkan dari
ortet yang dipilih dari pengujian progeni perlu diuji terlebih dahulu pada pengujian
klonal sebelum dilepas secara komersial (Purba et al. 1997).
Program seleksi dengan tujuan utama untuk memperbaiki CPO, strategi
seleksi yang diterapkan berbasis pada famili dan individu, lazim disebut Family
and Individual Palm Selection (FIPS) (Asmono et al. 1999).
Strategi pemuliaan tanaman untuk mentransfer karakter penting dari
satu spesies ke spesies lainnya dapat dilakukan melalui program silang balik.
Asmono (1999) menjelaskan, silang balik merupakan prosedur umum yang
digunakan untuk mentransfer karakter-karakter spesifik dari satu spesies ke
spesies lainnya. Saat ini dikenal dua spesies utama kelapa sawit: E. guineensis
dan E. oleifera. Kelapa sawit komersial yang dikenal saat ini, E. guineensis,
memiliki berbagai keunggulan, utamanya kandungan CPO yang tinggi. Namun
demikian, beberapa komponen penting, seperti kandungan ALTJ, pada E. gui-
neensis umumnya sangat rendah (40-60%). Di sisi lain, E. oleifera dikenal
sebagai spesies kelapa sawit yang memiliki kandungan CPO sangat rendah.
Namun demikian, persentase ALTJ-nya sangat tinggi (70-83%), dan mempunyai
pertumbuhan meninggi yang lambat. PPKS, dengan menggunakan metode
silang balik, saat ini berupaya untuk mentransfer karakter unggul
E.
oleifera,utamanya ALTJ tinggi, ke
E.
guineensis. Harapan dari upaya tersebut adalahdihasilkannya kelapa sawit unggul yang mempunyai kandungan CPO sekaligus
ALTJ yang tinggi.
5. Produksi dan Kualitas Minyak
Meningkatnya perhatian konsumen terhadap kesehatan (khususnya
minyak nabati), membuat para peneliti kelapa sawit memberikan perhatian yang
yang mengambil keunggulan dari plasma nutfah E. oleifera perlu dilanjutkan
agar terjadi integrasi ale1 yang bermutu terbaik dari
E.
oleifera ke dalam populasiE. guineensis (Pamin 1998).
Kelompok spesies E. guineensis, Dxi? mampu menghasilkan CPO sangat
tinggi, 7.0-7.9 ton/ha/tahun (Asmono et al. 1998). Tetapi, komponen kualitas
minyak yang dihasilkan
E.
guineensis tergolong rendah, kandungan ALTJ-nyaberkisar 40-60%. Sebaliknya, E. oleifera, walaupun produksinya tergolong
rendah, kurang dari 1 ton/ha/tahun, tetapi kandungan komponen kualitas
minyaknya tergolong sangat tinggi, kandungan ALTJ-nya berkisar 70-83%
(Syukur dan Lubis 1985). Tabel 1 memperlihatkan kandungan asam lemak dan
beta-karoten E. oleifera, E. guineensis, dan hibridanya.
Tabel 1. Kandungan asam lemak dan beta-karoten E. oleifera, E. guineen-
sis, dan hibridanya
E. oleifera orijin E. oleifera X
Karakteristik E. guineensis
Suriname E. guineensis
1. Asam lemak jenuh (%)
a. Asam miristat 0.1 0.8 1.5
b. Asam palmitat 24.9 36.8 41.8
c. Asam stearat 2.4 3.2 4.7
2. Asam lemak tak jenuh (%)
a. Asam oleat 66.0 45.3 37.4
b. Asam linolenat 5.0 13.0 13.1
3. Iodine value (%) 70.0 62.6 54.9
4. Beta-karoten (ppm) 4300-4600 1200-2400 500
Sumber: Choo dan Yap (1997), Syukur dan Lubis (1985).
Marka Molekuler
1. Potensi
Plasma nutfah merupakan sumber genetik yang perlu mendapat
perhatian, tidak hanya mengumpulkan dan memelihara, tetapi juga
mengkarakterisasi keragaman genetik, mengevaluasi sifat-sifat yang dikehendaki
dan memanfaatkannya untuk pemuliaan tanaman. Keragarnan genetik secara
Dengan kemajuan teknologi, keragaman genetik dapat terjadi melalui variasi
somaklonal, fusi protoplas maupun transfer gen. Informasi tentang keragaman
genetik plasma nutfah perlu diketahui karena sangat penting untuk membedakan
genotipe individu di dalam maupun antar spesies secara tepat yang sangat
diperlukan dalam pengembangan program pemuliaan tanaman (Bennett 1993).
Teknik pemuliaan secara molekuler (molecular breeding) merupakan
ilmu pengetahuan yang melibatkan pemakaian marka DNA untuk memfasilitasi
proses pemuliaan tersebut. Berdasarkan sejarahnya, para pemulia tanaman
mengandalkan sifat fenotip tanaman dalam rangka pemuliaan tanaman untuk
sifat unggul. Para pemulia dapat menggunakan teknik pemuliaan molekuler
sebagai alat untuk melengkapi teknik pemuliaan klasik (Forbes 2000).
Sekarang, teknologi marka moderen mengandalkan amplifikasi sejumlah
kecil DNA melalui polymerase chain reaction (PCR). Fragmen yang
teramplifikasi dipisahkan melalui gel elektroforesis dan divisualisasikan melalui
pewarnaan dengan dye spesifik untuk DNA. Dengan marka molekuler, proses
seleksi dapat dipercepat. Selain itu, lokus DNA yang bertanggung jawab terhadap
sifat kuantitatif tertentu dapat dipetakan (Jung 1999).
Potensi penggunaan marka sebagai alat untuk melakukan karakterisasi
genetik dalam program pemuliaan telah dikenal sejak puluhan tahun yang lalu.
Marka bisa dikategorikan sebagai marka morfologi, sitologi, dan yang terbaru
adalah marka molekuler (Moritz dan Hillis 1996; Sessions 1996).
Saat ini, kemajuan dalam bidang biologi berkembang sangat cepat dan
pesat. Biologi molekuler merupakan salah satu cabang ilmu yang mempelajari
organisme pada tingkat DNA. Teknik ini sangat membantu pemulia tanaman
dalam melakukan studi genetik dengan ketepatan yang lebih akurat. Untuk
mendapatkan informasi genetik dapat dilakukan dengan menggunakan marka
(Kongkiatngam et al. 1995; Powell et al. 1996; Karp dan Edwards 1997; Plieske
dan Struss 200 1).
Marka molekuler dapat memberi gambaran yang cukup tinggi tentang
perbedaan genetik individu, baik pada tingkat spesies maupun dengan kerabat
jauhnya. Menurut Tanksley (1983), penanda molekuler dapat mendeteksi variasi
genetik pada tingkat jaringan atau seluler, dan polimorfismenya tidak dipengaruhi
oleh lingkungan.
Marka molekuler yang pertama dan paling sederhana dikenal dengan
marka protein yang lazim disebut isozim. Polimorfisme protein dideteksi dengan
cara elektroforesis, dan perbedaan yang terdeteksi antar ale1 bergantung pada
pergantian asam-asam amino yang bermuatan. Untuk pencirian dan analisis
gen yang jumlahnya beragam, aplikasi marka isozim mempunyai keterbatasan
karena jumlah lokus yang bisa digunakan terbatas (Murphy et al. 1996).
Perkembangan dan penggunaan penanda RFLP relatif lebih belakangan
dibandingkan dengan isozim, meskipun demikian prinsip interpretasi analisis
genetik dari isozim dan RFLP sama. Setelah isozim, RFLP mendapat perhatian
yang lebih besar dari pakar genetika molekuler dan pemulia tanaman. Hal ini
karena RFLP dapat mengungkapkan perbedaan-perbedaan yang lebih banyak
antara individu-individu tanaman dibandingkan dengan isozim. Penanda RFLP
mendasarkan pada perbedaan dalam ukuran fragmen DNA nukleus
(kromosom), organel, atau total DNA yang dihasilkan dari pernotongan dengan
enzim restriksi. Individu-individu tanaman yang mempunyai perbedaan sekuen
DNA akan mempunyai perbedaan distribusi dari situs restriksi untuk suatu enzim
restriksi. Fragmen DNA hasil restriksi dipisahkan menurut ukuran berat molekul
pada gel elektroforesis, kemudian dipindahkan ke membran nilon dan
hibrida DNA yang terbentuk pada membran nilon yang dihibridisasi dengan
suatu probe berbeda antar individu yang diuji (McCouch dan Tanksley 1991).
2. RAPD
Marka RAPD yang dikembangkan oleh Williams et al. (1990) dihasilkan
melalui amplifikasi DNA secara in vitro dengan menggunakan teknik PCR. PCR
yang ditemukan Mullis (1980) berkembang sangat cepat sebagai teknik dalam
biologi molekuler. RAPD-PCR adalah metode sintesis DNA atau RNA secara in
vitro atau teknik memperbanyak molekul DNA atau RNA dengan menggunakan
sekuen acak primer oligonukleotida (10 basa primer) dan DNA polymerase
tahan panas yang dapat mempertahankan aktivitasnya sampai suhu 94°C.
Prosedur dalam teknik PCR terdiri dari proses: (1) denaturasi (denaturation)
DNA template menjadi DNA utas tunggal dengan suhu sekitar 94"C, (2)
penempelan (annealing) primer ke ujung utas DNA dengan suhu sekitar 55"C,
dan (3) pemanjangan (extension) DNA atau amplifikasi DNA yang dimulai dari
primer di ujung 5' ke ujung 3' dengan suhu sekitar 72°C. Ketiga langkah tersebut
merupakan prosedur satu siklus yang dapat diulang sebanyak 20-35 kali.
PCR membutuhkan DNA polymerase, dNTPs, DNA template, primer,
dan buffer reaksi. PCR dapat memperbanyak molekul DNA secara dramatik
yaitu sebagai deret geometri. DNA polymerase yang digunakan pertama kali
adalah fragmen Klenow. Kelemahan fragmen ini adalah tidak tahan suhu tinggi
sehingga setiap siklus harus diganti. Dengan ditemukannya Taq polymerase,
kekurangan ini dapat diatasi. Taqpolymerase, diperoleh dari 7hermus aquaticus,
suatu bakteri yang tahan suhu tinggi. DNA template, diperoleh dari hasil isolasi
DNA yang akan diamplifikasi. Rimer, merupakan beberapa nukleotida yang
berfungsi sebagai inisiasi proses sintesis DNA. Dalam PCR dibutuhkan 2 primer,
yaitu left primer (fonoawd) dan right primer (reverse), sedangkan apabila
Semenjak diperkenalkan oleh Williams et al. (1990), teknik RAPD menjadi
salah satu cara yang banyak digunakan untuk berbagai penelitian di bidang
biologi molekuler dan pemuliaan tanaman.
Kesederhanaan teknik RAPD dibandingkan dengan RFLE diantaranya:
(1) DNA tidak perlu dipotong dengan enzim restriksi, (2) sampel DNA yang
diperlukan relatif sedikit, (3) tidak memerlukan pemindahan DNA ke membran
nilon, (4) tidak memerlukan hibridisasi DNA, dan (5) tidak memerlukan prosedur
labelling. Teknik RAPD didasarkan pada amplifikasi DNA secara in vitro dengan
PCR, yaitu dengan mengatur variasi temperatur pada mesin PCR. Siklus termal
akibat adanya variasi temperatur tersebut adalah denaturasi, penempelan primer,
dan perpanjangan pita DNA. Selama pengulangan siklus tersebut dibantu oleh
enzim Taq DNA polymerase (Nair 1993).
Kemudahan teknik RAPD dibandingkan dengan RFLP adalah: (1)
pengetahuan latar belakang genom tanaman tidak diperlukan; (2) hasil RAPD
dapat diperoleh secara cepat jika dibandingkan dengan analisis RFLP yang
memerlukan banyak tahapan, dan (3) beberapa jenisprimer arbitrari dapat dibeli
dan digunakan untuk analisis genom semua jenis organisme (Williams et al.
1990). Sedangkan keterbatasannya adalah sangat sensitif terhadap kondisi reaksi
dan profil temperatur (Vos et al. 1995). Selain itu, marka RAPD bersifat dominan,
yaitu dalam populasi yang bersegregasi, individu yang homosigot dengan
individu yang heterosigot tidak dapat dibedakan dengan menggunakan penanda
RAPD, karena baik individu yang homo- atau yang heterosigot akan sama-
sama memberikan hasil pita DNA untuk suatu penanda RAPD tertentu ( R o ~ i n g
et al. 1995).
Marka RAPD dihasilkan melalui proses amplifikasi DNA seperti halnya
dalam melakukan PCR. Perbedaannya terletak pada penggunaan primer
oligonukleotida yang sekuennya dibuat secara random. Diantara jutaan
sekuen DNA-nya 100% sama atau hampir sama dengan
primer
oligonukleotidarandom
serta ada beberapa arah yang memiliki orientasi berlawanan yang hanyaberjarak beberapa ribu pasang basa antara satu dengan yang lain. Akibatnya,
jika genom tersebut dipakai sebagai
template
untuk reaksi PCR, berbagai ukuranpotongan DNA dapat teramplifikasi. Berbagai ukuran potongan DNA hasil
amplifikasi ini akan dapat dengan mudah dipisahkan berdasarkan ukurannya
dengan menggunakan teknik elektroforesis dan hasilnya dapat dilihat sebagai
pita-pita DNA dengan berbagai ukuran. Sekuen DNA dimana
primer
oligonukleotida berkomplementasi dapat berbeda-beda antara individu satu
dengan individu yang lain. Hal ini mengakibatkan kegagalan pembentukan
komplementasi antara DNA
template
danprimer
oligonukleotidanya. Kegagalanini menyebabkan hilangnya potongan DNA ukuran tertentu dari hasil amplifikasi
PCR yang dilakukan. Pita RAPD merupakan lokus DNA yang bersifat dominan
dan polimorfismenya diskor berdasarkan ada atau tidaknya pita (Rafalski
et al.
1994).
Marka RAPD sangat sesuai untuk pemetaan genetik, pemuliaan tanaman
dan hewan, dan sidik-jari DNA, dengan kegunaan khusus untuk kajian genetika
populasi. Penanda RAPD juga dapat memberikan efisiensi pengujian untuk
polimorfisme, yang dapat mempercepat identifikasi dan isolasi fragmen DNA
kromosom yang spesifik (Williams
et al.
1990).3. Aplikasi
RAPD
pada Tanaman Kelapa SawitPada awalnya, marka molekuler telah digunakan secara luas dalam kajian
keragaman genetik kelapa sawit. Beberapa contoh kajian pada kelapa sawit
yang menggunakan RAPD, diantaranya Shah
et al.
(1994) menggunakan markaRAPD untuk determinasi variasi genetik kelapa sawit di Afrika, Rajanaidu
et
al.(2000) yang telah melakukan penapisan populasi kelapa sawit menggunakan
marka RAPD dan R W Moretzsohn
et
al. (2000) telah melakukan pembentukanToruan-Mathius
et a!.
(1997) dengan RAPD melakukan penapisan 75primer
terhadap sampel individu Dura, Pisifera, dan Tenera mendapatkan 10
primer
(OPC-7,OPD-2,OPD-3,OPD-6,OPD-15,OPD-16,OPD-20,OPH-3,OPH-9,
dan
OPH-
12) yang diduga mampu membedakan Dura, Pisifera, dan Tenera.Peta Pautan Genetik dan Quantitatiue Trait Loci (QTL)
Penetapan marka genetik terhadap posisi relatif pada kromosom
berdasarkan frekuensi pindah silang, disebut sebagai peta genetik (Karp 1996),
lazimnya disebut peta pautan genetik. Menurut Grattapaglia dan Sederoff (1 994),
pada dasarnya konstruksi peta pautan genetik melibatkan aplikasi teknik biologi
molekuler pada konsep pewarisan sifat. Terdapat dua ha1 yang dibutuhkan dalam
pembentukan peta pautan genetik, yaitu reproduksi seksual dan generasi progeni
serta marka yang mengikuti hukum Mendel dalam jumlah yang banyak. Langkah
pertama pembentukan peta pautan genetik adalah seleksi individu yang
disilangkan untuk mendapatkan peluang besar mendeteksi polimorfisme
genetik. Biasanya, silsilah yang digunakan dalam konstruksi peta pautan genetik
melibatkan persilangan antara dua galur
inbred
yang diperoleh melalui generasisilang dalam. Progeni F, memiliki genotipe monomorfik. Silang dalam atau
backcross
dilakukan untuk membentuk generasi F, atau BC,, yangmenyebabkan genotipe marka molekulernya bersegregasi. Pada F, yang
dibentuk dengan menyilangdalamkan generasi F, , marka akan bersegregasi
1 (AA):2(Aa): 1 (aa) untuk marka kodominan, dan 3(A-): 1 (aa) untuk marka
dominan. Pada silang balik terhadap galur homozigot resesif, marka dominan
dan kodominan akan bersegregasi 1 (AA): 1 (Aa).
Faktor genetik yang bertanggung jawab terhadap variasi fenotipik yang
QTL hanya menunjukkan suatu region pada genom, dan dapat diperbandingkan
terhadap satu atau lebih gen fungsional (Falconer dan Mackay 1996).
QTL merupakan segmen pada kromosom yang dijumpai pada genom
yang mengandung satu atau beberapa gen yang pada akhirnya mengarahkan
ekspresi beberapa sifat kuantitatif. Dewasa ini, para peneliti di bidang biotekno-
logi tanaman dapat menggunakan QTL untuk menemukan gen yang diinginkan
dan gen baru, yaitu berbagai sifat agronomi seperti hasil, status tanaman dan
komponennya, sifat kualitas, resistensi, dan stres lingkungan (Forbes 2000).
Untuk tujuan analisis QTL diperlukan informasi berupa: (1) peta pautan
genetik berkerapatan tinggi, dan (2) data kuantitatif yang diukur dari parameter
yang diinginkan. Menurut Liu (1998), peta pautan genetik adalah model abstrak
berupa urutan linier kelompok gen dan marka. Gen dapat didefinisikan sebagai
faktor mendelian atau faktor DNA. Marka dapat berupa marka sitologi, suatu
varian yang berdasarkan perubahan gen atau protein yang telah diketahui atau
fragmen DNA yang tidak diketahui fungsinya. Gen dan marka tersebut hams
memiliki pewarisan sifat sederhana yang dapat ditelusuri dari generasi ke
generasi. Gen dengan fungsi yang telah diketahui dapat dipertirnbangkan sebagai
marka jika memiliki variasi yang dapat dideteksi. Peta pautan genetik didasarkan
pada rekombinasi kromosom homolog selama meiosis, jadi peta ini merupakan
peta meiosis. Dalam meiosis, jika dua atau lebih marka berlokasi dekat dalarn
suatu kromosom, alel-ale1 marka tersebut biasanya diwariskan bersama. Oleh
karena itulah peta tersebut diberi istilah peta pautan genetik.
MAS-BC pada Tanaman Kelapa Sawit
Program pemuliaan untuk meningkatkan kualitas minyak kelapa sawit
tanpa mengorbankan produktivitas CPO, dapat dilakukan dengan metode silang
tetua donor, E. oleifera, sebagai sumber gen yang menentukan kadar ALTJ
tinggi, terhadap tetua pemulih; E. guineensis, sebagai sumber gen yang
menentukan produksi CPO tinggi, (2) melakukan silang balik turunannya
terhadap tetua pemulih, dan (3) mengidentifikasi individu hasil silang balik
sehingga diperoleh individu BC, terpilih. Menurut Ribaut dan Hoisington (1998),
kegiatan silang balik dan identifikasi individu turunannya tersebut hams dilakukan
secara berulang hingga 10- 15 kali.
Walaupun secara teoritis metode ini mungkin untuk dilakukan, tetapi
upaya ini terbentur oleh waktu pelaksanaan seleksi (pada tanaman kelapa sawit,
10-13 tahun per siklus), volume pekerjaan yang sangat besar karena haws
melakukan silang balik 10-15 kali, dan adanya sterilitas pada sebagian hasil
persilangan. Dengan demikian, diperlukan metode atau strategi lain untuk
mengatasi kendala tersebut. Ribaut dan Hoisington (1998), mengemukakan
bahwa dengan bantuan marka molekuler (metode MAS), hanya diperlukan satu
atau dua kali daur silang balik.
Menurut Openshaw et al. (1994), pada BC, dan generasi silang balik
selanjutnya, individu terpilih telah mernbawa transfer gen dari hasil silang baliknya
terhadap tetua pemulih. Proporsi harapan genom tetua donor berkurang menjadi
setengah setiap generasi silang balik. Rata-rata persentase harapan genom tetua
pemulih
(%W
recwrentparent) pada setiap generasi silang balik dihitung denganrumus:
%RP
= 100[1-(0.5)"+']dimana
n
adalah jumlah silang balik. Dengan demikian, metode MAS-BC, yangmerupakan kombinasi metode MAS dengan program silang balik untuk
mendapatkan introgresi gen baru dari tetua donor ke tetua pemulih, diharapkan
dapat membantu program pemuliaan kelapa sawit untuk mendapatkan tanaman
BAB-I11
KERAGAAN KOMPONEN HASlL DAN KUALITAS MINYAK KELAPA SAWlT DARl POPULASI SILANG BALlK GENERASI
PERTAMA HASlL PERSILANGAN HI BRlDA Elaeis oleifera
(EKS BRAZIL)
X E. guineensis
DENGAN E. guineensisAbstrak
Populasi silang balik generasi pertama (BC,) hasil persilangan hibrida [Elaeis oleifera (eks Brazil) X E. guineensis; donorparent, maternal] dengan E. guineensis (recurrent parent, paternal) telah ditanam sejak 1993 di Afdeling Ill Kebun Bah Jambi - Pematang Siantar (Simalungun). Evaluasi telah dilakukan untuk melihat keragaan komponen hasil dan kualitas minyak populasi tersebut. Berdasarkan analisis lintas dan regresi bertatar, peubah berat tandan dan persentase minyak per tandan memiliki pengaruh langsung paling besar dalam menentukan besarnya hasil minyak per tandan pada komponen hasil. Sedangkan peubah miristat (C-14), palmitat (C-16), stearat (C-18), linoleat (C- 18:2), dan linolenat (C-18:3) memiliki pengaruh paling besar dalam menentukan besarnya kandungan oleat pada komponen kualitas minyak. Pada populasi ini hambatan pautannya masih besar, yaitu tingginya kandungan oleat belum disertai dengan tingginya hasil minyak per tandan. Guna pengembangan program pemuliaan untuk menghasilkan kelapa sawit yang memiliki kualitas minyak tinggi (oleat), terpilih pohon 93-107-33 T
sebagai tetua donor untuk menghasilkan populasi BC,.
Kata kunci: BC,, komponen hasil, komponen kualitas minyak, asam oleat.
PERFORMANCE YIELD AND QUALITY OIL PALM COMPONENTS OF BACKCROSS OF FIRST GENERATION FROM CROSSING
HYBRID Elaeis oleifera (EKS BRAZIL)
X
E. guineensisWITH E. guineensis
Abstract
Population of first generation of backcross (BC,) from crossing hybrid [Elaeis oleifera
(ex Brazil) X E. guineensis; donorparent, maternal] with E. guineensis (recurrent par- ent, paternal) was planting since 1993 at Afdeling Ill Bah Jambi Plantation - Pematang Siantar (Simalungun). The evaluations were doing to seek performances of yield and oil quality of the population. Based on path analysis and stepwise regression, variables of weight of bunch and percentage of oil per bunch have direct effect very big for determining yield oil per bunch at the yield components. Meanwhile, variables miristic (C-14), palmitic (C- 16), stearic (C-18), linoleic (C-18:2), and linolenic (C-18:3) has di- rect effect very big for determining content of oleic at the oil quality components. In this population, linkage drag event still big, i.e. high level of oleic content not yet followed high level of yield of oil per bunch. Breeding program toward provide high oil quality, tree 93-107-33 T were selected a s donor parent to generate BC, population.
PENDAHULUAN
Sebagai penghasil minyak nabati terbesar kedua setelah kedelai (Mayes
et al. 2000), kelapa sawit (Elaeis guineensis Jacq.), merupakan komoditas
ekspor yang memberi kontribusi penting terhadap perekonomian nasional,
utamanya sebagai penghasil devisa negara. Volume ekspor akan meningkat
seiring dengan peningkatan jumlah penduduk dunia, karena sebagian besar
minyak kelapa sawit digunakan untuk pangan (go%), selebihnya (10%) non-
pangan (Jalani 1998).
Sejalan dengan program pengembangan bahan tanaman kelapa sawit,
kegiatan pemuliaan tidak hanya difokuskan pada peningkatan produktivitas
tanaman, melainkan juga pada aspek kualitas minyak yang dihasilkan. Kualitas
minyak kelapa sawit ditentukan oleh besarnya kandungan asam lemak tak jenuh
(ALTJ) dan komponen minornya (diantaranya: beta-carotene, tocopherol, dan
tocotrienol). Salah satu upaya yang dilakukan dalam rangka peningkatan kualitas
minyak adalah dengan memanfaatkan keunggulan spesies E. oleifera melalui
program silang balik (Suprianto et al. 2000). Sementara itu, Asmono et al.
(1999) mengemukakan adanya kepedulian akan kesehatan mendorong para
pemulia kelapa sawit menaruh perhatian yang lebih serius terhadap perbaikan
kualitas minyak.
Ada dua spesies kelapa sawit, yaitu
E.
guineensis dan E. oleifera.E.
guineensis dikenal sebagai kelapa sawit komersial, memiliki berbagai
keunggulan, utamanya kandungan minyak mentah (crude palm oil, CPO) yang
tinggi. Narnun dernikian, beberapa komponen penting, seperti kandungan ALTJ,
umumnya sangat rendah (40-60%). Sedangkan E. oleifera dikenal sebagai
spesies kelapa sawit yang memiliki kandungan CPO sangat rendah, tetapi
memiliki persentase ALTJ sangat tinggi (70-83%) dan pertumbuhan meninggi
metode silang balik, saat ini bempaya untuk mentransfer karakter unggul E.
oleifera, utamanya ALTJ tinggi, ke E. guineensis. Tujuan utamanya adalah untuk
menghasilkan kelapa sawit unggul yang mempunyai kandungan CPO sekaligus
ALTJ yang tinggi (Asmono et
al.
1999). E. oleifera, selain memiliki kadar ALTJyang tinggi, juga memiliki kandungan beta-karoten yang tinggi. Kandungan
beta-karoten pada E. oleifera mencapai 4300-4600 ppm, sedangkan hibridanya
terhadap E. guineensis mencapai 1200-2400 ppm (Choo dan Yap 1997; Syukur
dan Lubis 1985 ).
Secara khusus penelitian ini bertujuan menseleksi progeni BC, (eks Brazil)
yang memiliki kualitas minyak paling baik, utamanya kandungan oleat (C: 18- 1)
untuk dijadikan tetua donor BC,. Secara umum bertujuan untuk mengetahui
keragaan komponen hasil dan komponen kualitas minyak populasi BC,.
BAHAN DAN METODE
Bahan Tanaman
Penelitian menggunakan 23 progeni BC, dari persilangan hibrida [E. olei-
fera (eks Brazil) X E. guineensis; donorparent, maternal] dengan E. guineensis
(recurrent parent, paternal), silsilah disajikan pada Lampiran 1. Progeni tersebut
mempakan existingpopulation yang ditanam sejak 1993 di Afdeling 111 Kebun
Bah Jambi (Pematang Siantar
-
Simalungun) rnilik Balai Penelitian Marihat-
PPKS. Kedua tetua populasi BC, itu juga digunakan dalam penelitian ini.
Pengamatan
Untuk melihat keragaan komponen hasil dan kualitas minyak, telah
dilakukan pengurnpulan data selama 17 bulan. Pengukuran atau analisis semua
peubah dilakukan sesuai dengan prosedur baku yang dilakukan di Laboratorium
Analisis Tandan Balai Penelitian Marihat
-
PPKS (tidak dipublikasikan). HasilPuslit Kelapa Sawit
Bagian Laboratorium
Analisis
Tbndan
(KARTU INDIVIDUIL)
Kebun : ... Program : No. Analisis :
[image:42.831.133.695.85.465.2]Tgl.Panen : [ I