NIDA WAFIQAH NABILA M. SOLIN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
KERAGAMAN GENETIK DAN IDENTIFIKASI PENANDA SPESIFIK
KARAKTER PRODUKSI PADA PROGENI DAN TETUA
iii
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Keragaman Genetik dan Identifikasi Penanda Spesifik Karakter Produksi pada Progeni dan Tetua Kelapa Sawit dengan Marka SSR adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
NIDA WAFIQAH NABILA M. SOLIN. Keragaman Genetik dan Identifikasi Penanda Spesifik Karakter Produksi pada Progeni dan Tetua Kelapa Sawit dengan Marka SSR. Dibimbing oleh SOBIR dan NURITA TORUAN-MATHIUS.
Studi tesis ini mengamati hubungan antara marka mikrosatelit dengan karakter produksi kelapa sawit (Elaeis guineensis Jacq.) dalam upaya untuk mendapatkan penanda spesifik yang dapat bermanfaat dalam pemilihan tetua.
Percobaan pertama dilakukan dengan mengamati keragaman genetik 80 progeni kelapa sawit yang telah dikelompokkan berdasarkan karakter produksi dan tinggi rendah karakter produksinya. Masing-masing 20 individu diamati untuk karakter minyak per tandan (OB), jumlah tandan (BN), produksi minyak (OY) dan tandan buah segar (FFB). Hasil penelitian menunjukkan keragaman genetik yang cukup tinggi dari keempat karakter, yang ditunjukkan oleh rata-rata persentase polimorfik dan PIC yang cukup tinggi, yaitu 52,78% dan 0.51. Konstruksi dendogram berdasarkan UPGMA dapat membedakan karakter satu dengan yang lain dengan persentase kemiripan genetik 56-100% atau keragaman genetik ~44%. Pemisahan karakter ke dalam empat kelompok ini juga dikonfirmasi menggunakan PCoA (Principle Coordinate Analysis). Selain itu juga didapatkan marka yang spesifik untuk karakter yang diamati, yaitu satu lokus spesifik terhadap karakter OB, empat lokus spesifik terhadap karakter BN, satu lokus spesifik terhadap karakter OY dan dua lokus spesifik terhadap karakter FFB.
Percobaan kedua dilakukan untuk mengidentifikasi marka yang spesifik terhadap keempat karakter. Pendekatan korelasi dan regresi stepwise digunakan dalam percobaan ini. Hasil penelitian menunjukkan bahwa terdapat 11 marka spesifik yang diperoleh menggunakan pendekatan korelasi, serta 16 marka spesifik diperoleh menggunakan pendekatan regresi stepwise. Dari hasil tesebut diketahui bahwa satu marka berkorelasi dan beregresi positif terhadap OB, dua marka berkorelasi dan beregresi positif terhadap BN, satu marka beregresi positif terhadap OY, namun hanya satu marka yang berkorelasi positif terhadap OY, serta tidak ada maka yang berkorelasi maupun beregresi positif terhadap FFB.
Percobaan ketiga dilakukan untuk mengetahui keragaman genetik tetua Dura yang digunakan sebagai induk betina pada percobaan 1 dan 2, dan peranannya dalam keragaman keempat karakter yang diuji. Dua populasi Dura hasil selfing dengan 30 sampel per populasi digunakan untuk analisis. Hasil analisis menunjukkan pemisahan tetua ke dalam dua kelompok berdasarkan selfingnya. Grup A, dengan keragaman genetik ~20%, merupakan hasil selfing D1. Sedangkan Grup B, dengan keragaman genetik ~28% merupakan hasil selfing D2. Kedua bahan genetik ini memiliki keragaman genetik yang rendah di dalam kelompok, tetapi keragaman genetik lebih tinggi antar kelompoknya (~32%), sebagaimana dikonfirmasi dengan menggunakan PCoA dan Structure. Hasil analisis uji t menunjukkan bahwa pasangan D1 dan D2 dengan keempat karakter, memiliki pengaruh yang signifikan terhadap karakter OB, OY dan FFB, sementara pada karakter BN, pengaruh Dura tidak signifikan.
SUMMARY
NIDA WAFIQAH NABILA M. SOLIN. Genetic Diversity and Identification of Specific Production Character Markers of Oil Palm Progeny and Parent with SSR Marker. Supervised by SOBIR dan NURITA TORUAN-MATHIUS.
This thesis studies the relationship between microsatellite markers with the character of the production of oil palm (Elaeis guineensis Jacq.) in an attempt to get specific markers that can be useful in the parental selection.
The first experiment was carried out by observing the genetic diversity of 80 progeny palm oil that has been classified by the character of high and and low production. Each of these 20 individuals were observed for the characters of oil to bunch (OB), bunch number (BN), oil yield (OY) and fresh fruit bunches (FFB). The results showed a fairly high genetic diversity of the four characters, which is shown by the quite high average percentage of polymorphic and PIC, ie 52.78% and 0.51. Construction of dendogram based on UPGMA can distinguish the characters from one to another by the percentage of genetic similarity 56-100% or ~44% of genetic diversity. The separation of the characters into the four groups was also confirmed using PCoA (Principle Coordinate Analysis). It also found the specific markers for the characters observed, ie one marker specific to the character of OB, four markers specific to the character of BN, one marker specific to the character of OY and two marker specific to the character of FFB.
The second experiment was conducted to identify specific markers of the four characters. Correlation and stepwise regression approach used in this experiment. The results showed that there are 11 specific markers were obtained using the correlation approach, as well as 16 specific markers obtained using stepwise regression approach. From the results it is known that one marker positively correlated and regressed to the OB, two markers positively correlated and regressed to the BN, one marker positively regressed to OY, but only one marker that positively correlated to OY, and no one is positively correlated or regressed to FFB.
The third experiment was conducted to determine the genetic diversity of Dura parents which is used as female parent in experiments 1 and 2, and its role in the diversity of the four characters tested. Two populations of Dura selfing with 30 samples per population used for analysis. The analysis showed parental separation into two groups based on the selfing. Group A, with ~20% genetic diversity, is the result of D1 selfing. While Group B, with ~28% genetic diversity is the result of D2 selfing. Both genetic material has a low genetic diversity within groups, but higher genetic diversity between group (~ 68%), as well as confirmed by PCoA and Structure. The results of t-test analysis showed that the pair D1 and D2 with a fourth character, has a significant effect on the character of the OB, OY and FFB, while the character of BN, Dura influence is not significant.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
NIDA WAFIQAH NABILA M. SOLIN
KERAGAMAN GENETIK DAN IDENTIFIKASI PENANDA SPESIFIK
KARAKTER PRODUKSI PADA PROGENI DAN TETUA
Judul Tesis : Keragaman Genetik dan Identifikasi Penanda Spesifik Karakter Produksi pada Progeni dan Tetua Kelapa Sawit dengan Marka SSR
Nama : Nida Wafiqah Nabila M. Solin NIM : A253110191
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Sobir, MSi Ketua
Diketahui oleh
Dr Nurita Toruan-Mathius, MS Anggota
Ketua Program Studi Pemuliaan dan
Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu EK, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
: Nida Wafiqah Nabila M. Solin
セim@ : A253110191
Disetujui oleh
Komisi Pembimbing
Prof Dr Ir Sobir, MSi Dr Nur' a Toruan-Mathius MS
Ketua Anggota
Diketahui oleh
Ketua Program Studi Pemuliaan dan
Bioteknologi Tanaman
Dr Ir Yudi anti Wahyu EK, MS
Tanggal Ujian: 13 Maret 2014
Sekolah Pascasarjana
GMM]ZZZヲ セiZZbセュ@ Syah, MScAgr
Tanggal Lulus:
o
3
APR
2014
--PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT. atas segala karunia-Nya sehingga tesis ini berhasil diselesaikan. Adapun judul tesis ini yaitu “Keragaman Genetik dan Identifikasi Penanda Spesifik Karakter Produksi pada Progeni dan Tetua Kelapa Sawit dengan Marka SSR”.
Terima kasih penulis ucapkan kepada Bapak Prof Dr Ir Sobir, MSi dan Ibu Dr Nurita Toruan-Mathius, MS selaku komisi pembimbing, atas bimbingan, arahan, kritik dan saran, serta dukungan moril mulai dari pembuatan proposal, pelaksanaan penelitian, sampai penyelesaian tesis ini.
Penulis juga mengucapkan terima kasih kepada Pimpinan Laboratorium Bioteknologi, Plant Production and Biotechnology Division, PT. SMART Tbk atas izin yang diberikan untuk menggunakan fasilitas penelitian, sampai penelitian diselesaikan dengan baik. Terima kasih juga penulis sampaikan kepada Direktur Lembaga Pengelola Dana Pendidikan yang telah memberikan beasiswa tesis kepada penulis.
Terima kasih penulis ucapkan kepada Bapak Ir Ismail Maskromo, MSi, Bapak Roberdi SP, MSi, Bapak Azis Natawijaya, SP, MSi yang banyak membantu penulis dalam menyelesaikan tesis ini. Ucapan terima kasih juga penulis sampaikan kepada Bapak Prof Sudarsono, MSc, atas keluangan waktunya untuk berdiskusi dengan penulis. Terima kasih juga penulis ucapkan kepada Yogo Adhi Nugroho, SP, MSi, Wulan Artutiningsih, SP, MSi, Intana Pamela dan Diana Mekar Jayanti, yang banyak membantu pelaksanaan penelitian mulai dari pengambilan sampel sampai visualisasi gel acrylamide, serta rekan-rekan PBT 2011 yang banyak membantu baik selama perkuliahan sampai penulisan tesis ini.
Dengan segenap rasa hormat dan bangga serta terima kasih penulis sampaikan kepada Ayahanda tercinta Bapak Dr Mutsyuhito Solin, MPd dan Ibunda tercinta Ibu Dr Sri Minda Murni, MS, serta abang Syafiq Anshori M. Solin, SS dan adik-adik, Thareq Muhammad M. Solin dan Aqilah Nadira Safia M. Solin, yang telah memberikan dukungan moril dan materiil, kasih sayang, bimbingan dan doa kepada penulis. Penulis juga mengucapkan terima kasih kepada Allen Wijaya, SP atas kesabaran, serta dukungan dan doanya untuk penulis. Tak lupa penulis mengucapkan terima kasih kepada Nenek Dra. Masniari Poeloengan, MSi dan Kakek Dr. Amri Jahi, MS atas segala bantuan dan motivasinya selama penulis berada di Bogor.
Akhirnya, dengan segala kerendahan hati penulis berharap agar hasil penelitian ini dapat bermanfaat dan dapat digunakan untuk kepentingan penelitian serta kemajuan ilmu pengetahuan.
Bogor, Maret 2014
RIWAYAT HIDUP
DAFTAR ISI
DAFTAR ISI vii
DAFTAR TABEL ix
DAFTAR GAMBAR ix
DAFTAR LAMPIRAN x
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 3
2 TINJAUAN PUSTAKA 4
Kelapa Sawit 4
Pemuliaan Kelapa Sawit 5
Mikrosatelit 5
Penerapan Metode Mikrosatelit pada Kelapa Sawit 6
Populasi Half-Sib 7
Bulk Segregant Analysis (BSA) 7
Analisis Statistik untuk Data Molekuler 8
3 KERAGAMAN GENETIK KARAKTER PRODUKSI POPULASI DxP PADA FAMILI PATERNAL HALF-SIB KELAPA SAWIT BERDASARKAN MARKA DNA SSR 9
ABSTRAK 9
ABSTRACT 9
PENDAHULUAN 10
BAHAN DAN METODE 11
Tempat dan Waktu Penelitian 11
Bahan Tanaman dan Isolasi DNA 11
Pemilihan Populasi 11
Amplifikasi PCR 12
Analisis Data 12
HASIL DAN PEMBAHASAN 14
Uji Signifikansi Individu di dalam Bulk 14
Analisis Mikrosatelit dan Polimorfisme 14
Polymorphism Information Content (PIC) 15
Jumlah alel (Na), jumlah alel efektif (Ne), Indeks Informasi (I), heretozigositas teramati (Ho), Heterozigositas harapan (He) dan indeks Fiksasi (F) 16
Keragaman Genetik Antar dan Intra Populasi 17
KESIMPULAN 20
4 IDENTIFIKASI MARKA MOLEKULER SPESIFIK UNTUK KARAKTER PRODUKSI POPULASI DxP PADA FAMILI PATERNAL HALF-SIB KELAPA SAWIT 21
ABSTRACT 21
PENDAHULUAN 22
BAHAN DAN METODE 23
Tempat dan Waktu Penelitian 23
Bahan Tanaman dan Isolasi DNA 23
Pemilihan Populasi 23
Amplifikasi PCR 25
Analisis Data 25
HASIL DAN PEMBAHASAN 25
KESIMPULAN 29
5 KERAGAMAN GENETIK POPULASI TETUA SAUDARA KANDUNG (SIBS)
KELAPA SAWIT DURA DELI BERDASARKAN MARKA DNA SSR 30
ABSTRAK 30
ABSTRACT 31
PENDAHULUAN 32
BAHAN DAN METODE 33
Tempat dan Waktu Penelitian 33
Bahan Tanaman dan Isolasi DNA 33
Amplifikasi PCR 35
Analisis Data 35
HASIL DAN PEMBAHASAN 35
Analisis Mikrosatelit dan Polimorfisme 35
Jumlah Alel Efektif (Ne), Heterozigositas Harapan (He), Heterozigositas Teramati (Ho) dan Polymorphism Information Content (PIC) 36
Keragaman Genetik Antar dan Intra Populasi 38
Uji Signifikansi Tetua dan Individu dengan Keempat Karakter yang Dianalisis 40
KESIMPULAN 41
6 PEMBAHASAN UMUM 42
7 KESIMPULAN UMUM 45
DAFTAR TABEL
1. Daftar primer beserta urutan basa nukleotida dan suhu annealing yang digunakan 13 2. Rekapitulasi uji t dari keempat karakter yang dianalisis 14 3. Jumlah alel dan Polymorphism Information Content (PIC) pada mikrosatelit yang
dievaluasi 15
4. Rata-rata jumlah alel (Na), jumlah alel efektif (Ne), Indeks Informasi (I), heretozigositas teramati (Ho), Heterozigositas harapan (He) dan indeks Fiksasi (F)
pada setiap karakter 16
5. Daftar primer beserta urutan basa nukleotida dan suhu annealing yang digunakan 24 6. Matriks Korelasi Antar Marka dan Karakter minyak per tandan (OB) 25 7. Matriks Korelasi Antar Marka dan Karakter jumlah tandan (BN) 26 8. Matriks Korelasi Antar Marka dan Karakter Produksi Minyak (OY) 27 9. Matriks Korelasi Antar Marka dan Karakter Tandan Buah Segar (FFB) 27 10.Rekapitulasi Jumlah Marka yang Berkorelasi Nyata terhadap Masing-Masing
Karakter produksi 28
11.Rekapitulasi Jumlah Marka yang Beregresi Nyata terhadap Masing-Masing
Karakter produksi 29
12.Daftar primer beserta urutan basa nukleotida dan suhu annealing yang digunakan 34 13.Jumlah alel dan polimorfisme dari mikrosatelit yang dievaluasi 36 14.Jumlah alel efektif (Ne), heterozigositas harapan (He), heterozigositas teramati (Ho)
dan Polymorphism Information Content (PIC) pada 20 lokus SSR yang polimorfik 37 15.Rekapitulasi uji t tetua D1 dan D2 dengan keempat karakter yang dianalisis 41
DAFTAR GAMBAR
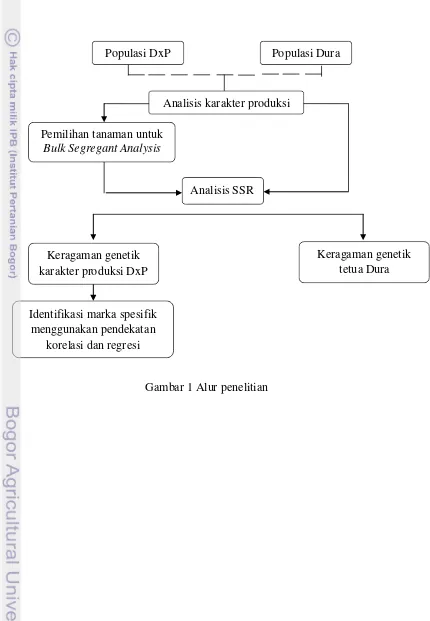
1. Alur penelitian 3
2. Pola amplifikasi dua marka mikrosatelit dari bahan genetik kelapa sawit dengan 4 alel (a) lokus monomorfik; (b) lokus polimorfik. 10 lokus paling kiri merupakan individu di dalam bulk dengan produksi rendah, dan 10 lokus paling kanan merupakan individu di dalam bulk dengan produksi tinggi 15
3. Principal Coordinate Analysis (PCoA) dari empat karakter produksi kelapa sawit 17
4. Principal Coordinate Analysis (PCoA) pada masing-masing karakter yang diuji
menggunakan 27 primer mikrosatelit. 18
5. Dendogram kemiripan genetik individu dalam bulk untuk karakter produksi jumlah tandan buah segar (bunch number/BN), berat tandan buah segar (fresh fruit bunches/FFB), jumlah minyak per tandan (oil to bunch/OB), dan produksi minyak
(oil yield/OY) sawit berdasarkan marka SSR 19
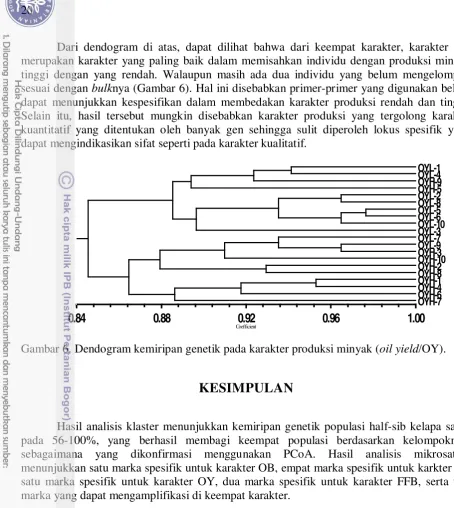
6. Dendogram kemiripan genetik pada karakter produksi minyak (oil yield/OY). 20
7. Principal Coordinate Analysis pada 60 sampel kelapa sawit yang digunakan 38
8. Kesamaan genetik populasi tetua kelapa sawit dengan hasil klastering terbaik
(K=2) dari 60 pohon tetua. 39
9. Dendogram kesamaan genetik 60 sampel populasi tetua saudara kandung (sibs)
DAFTAR LAMPIRAN
1. Uji t pada keempat karakter yang diuji 52
1
PENDAHULUAN
Latar Belakang
Kelapa sawit (Elaeis guineensis Jacq.) merupakan komoditi perkebunan yang mempunyai peran penting dalam berbagai aspek kehidupan di Indonesia, khususnya aspek perekonomian dalam negeri. Indonesia memiliki beberapa keunggulan sebagai salah satu negara yang sangat potensial untuk menanamkan investasi di bidang perkebunan kelapa sawit dan industri hilirnya. Hal tersebut ditunjang dengan meningkatnya permintaan minyak sawit dunia untuk kebutuhan pangan (edible oil), industri (oleochemical), dan sumber energi alternatif berbasis biodiesel (Wahyono 2006). Secara konsisten pendapatan nasional dari kelapa sawit terus menunjukkan peningkatan dan berpeluang menjadi komoditi yang paling menguntungkan dibandingkan dengan komoditi lainnya (Sugema 2007).
Luas areal tanaman kelapa sawit di Indonesia, hanya sekitar 7 363 847 pada tahun 2008 kemudian berkembang sangat cepat hingga mencapai 9 074 621 pada tahun 2012. Hal ini seiring dengan meningkatnya produksi CPO, yaitu 17 539 788 pada tahun 2008 hingga mencapai 23 521 071 ton pada tahun 2012 (Direktorat Jenderal Perkebunan 2012). Indonesia sampai saat ini merupakan negara pengahasil Crude Palm Oil (CPO) terbesar di dunia.
Pengembangan industri kelapa sawit memerlukan beberapa upaya untuk mencapai tujuan peningkatan produktivitas nasional, salah satunya adalah pemanfaatan benih unggul bermutu yang didukung dengan ketersedian sumber daya genetik (plasma nutfah) yang mempunyai tingkat keragaman genetik yang tinggi (Direktorat Jenderal Perkebunan 2010). Benih unggul bermutu dapat dihasilkan dari program pemuliaan yang efisien dan terarah. Peningkatan produksi melalui penyediaan bibit unggul berdaya hasil tinggi merupakan salah satu upaya strategis, yang terus dilakukan untuk memenuhi kebutuhan minyak sawit yang semakin meningkat dari tahun ke tahun (Asmono et al. 1999).
Lamadji et al. (1999) menyatakan bahwa terdapat beberapa hambatan dalam pemuliaan secara konvensional, yaitu memerlukan waktu yang cukup lama, gen yang menjadi target seleksi untuk diekspresikan pada sifat-sifat morfologi sulit ditentukan, karena penampilan fenotipe tanaman bukan hanya ditentukan oleh komposisi genetik tetapi juga oleh lingkungan, rendahnya frekuensi individu yang diinginkan dalam suatu populasi sehingga menyulitkan kegiatan seleksi untuk mendapatkan hasil yang valid secara statistik, dan pautan gen antara sifat yang diinginkan dengan yang tidak diinginkan sulit dipisahkan saat persilangan.
karakter sesuai dengan peta pautan dan genom bertujuan untuk menyeleksi tanaman sesuai karakter yang diinginkan (Kasim dan Azrai 2004). Kemampuan penggunaan pita DNA tanaman pada setiap tahap pengembangan tanaman, membuat marka molekuler sebagai alat yang cepat dan akurat untuk mengevaluasi kebenaran dan kemurnian suatu kultivar (Lim and Rao 2005). Marka molekuler dapat mendeteksi variasi genetik dan sifat polimorfismenya tidak dipengaruhi oleh lingkungan (Tanksley 1983).
Marka SSR untuk kelapa sawit pertama kali dikembangkan oleh CIRAD Perancis. Billotte et al. (2001) berdasarkan hasil analisis data multivariate melaporkan kemampuan marka SSR yang sangat efisien untuk menunjukkan struktur keragaman genetik genus Elaeis sesuai dengan daerah asalnya. Berdasarkan tingkat variabilitas aleliknya yang tinggi, marka SSR dapat menjadi perangkat yang sangat bermanfaat untuk kajian genetik genus Elaeis, antara lain untuk identifikasi plasma nutfah dan pemetaan genetik intra atau interspesifik. Saghai-Maroof et al. (1994) mengemukakan beberapa alasan pemakaian SSR untuk analisis molekular yaitu: (1) melimpah, (2) terdistribusi dengan seragam, (3) sangat polimorfis, (4) kodominan, (5) dihasilkan dengan cepat melalui PCR, (6) relatif sederhana untuk ditafsirkan, dan (7) mudah diakses oleh laboratorium lain melalui publikasi sekuen primer. Bahkan Powell et al. (1996) membuktikan bahwa dari empat marka molekuler yang diuji (RFLP, RAPD, AFLP dan SSR), marka SSR memiliki kandungan informasi (kemampuan membedakan genotipe) yang paling tinggi untuk mengevaluasi plasma nutfah kedelai dibandingkan dengan marka molekuler yang lain.
Integrasi marka DNA ke dalam program marker-assisted selection (MAS) diketahui mampu meningkatkan efektivitas seleksi. Ketersediaan peta lokus sifat kuantitatif (quantitative trait loci/QTL) yang berasosiasi dengan komponen produktivitas minyak, merupakan prasyarat penerapan MAS. Lokus DNA yang berasosiasi dengan komponen produktivitas minyak dan mempunyai pengaruh genetik yang besar akan bermanfaat untuk meningkatkan efektifitas seleksi (Singh et al. 2009).
Teknik yang merupakan kombinasi seleksi berbasis marka molekuler dengan teknik pemuliaan konvensional merupakan alat bantu strategis yang dapat mempersingkat waktu seleksi, sehingga dapat mempercepat pencapaian tujuan pemuliaan tanaman, yaitu untuk menyediakan sumberdaya genetik yang memiliki karakter unggul dalam waktu yang singkat.
Tujuan Penelitian
Manfaat Penelitian
Manfaat penelitian ini adalah untuk memperoleh marka spesifik karakter produksi kelapa sawit, sehingga dapat dijadikan sebagai alat untuk seleksi berbasis marka (Marker Assisted Selection/MAS), yang dapat mempersingkat proses pemuliaan kelapa sawit.
Gambar 1Alur penelitian Analisis karakter produksi
Analisis SSR
Keragaman genetik karakter produksi DxP
Pemilihan tanaman untuk Bulk Segregant Analysis
Identifikasi marka spesifik menggunakan pendekatan
korelasi dan regresi
Keragaman genetik tetua Dura
2
TINJAUAN PUSTAKA
Kelapa Sawit
Kelapa sawit (Elaeis guineensis Jacq.) termasuk dalam subfamily Cocoidae dari family Arecaceae atau Palmae. Populasi alami E. guineensis sebagian besar ditemukan di Afrika Barat, sedang spesies E. oleifera di Amerika Tengah dan Selatan. Famili Palmae memuat lebih dari 225 genus dan 2600 spesies. Dari jumlah tersebut hanya 35 genus dan 75 spesies termasuk E. guineensis yang diketahui jumlah kromosomnya (Madon et al 1996).
Kelapa sawit (Elaeis guineensis Jacq.), berasal dari bahasa Yunani, yaitu elaion yang berarti minyak dan guineensis yang menunjukkan bahwa tanaman kelapa sawit berasal dari pantai Guinea Afrika Barat, sedangkan Jacq., adalah singkatan dari nama belakang Nicolaus Josef von Jacquin, orang yang memberi nama kelapa sawit secara botani (Hartley 1988). Tanaman ini memiliki genom diploid dengan 16 pasang kromosom homolog (2n = 32).
Tanaman kelapa sawit diintroduksi ke Indonesia pada tahun 1848. Sebanyak empat bibit kelapa sawit ditanam di Kebun Raya Bogor. Dari keempat bibit tersebut, dua bibit diintroduksi dari Bourbon atau Mauritius pada Februari 1848, dua bibit yang lain diintroduksi dari Amsterdam pada Maret 1848 (Pamin 1998).
Tipe pembungaan kelapa sawit adalah monoecious, yaitu bunga jantan dan betina ada di satu tanaman, tetapi pada tandan yang berbeda. Rasio bunga jantan terhadap betina dapat dipengaruhi keadaan iklim. Pada musim kemarau biasanya bunga jantan yang mendominasi sedangkan pada musim penghujan bunga betina yang mendominasi. Kadangkala dijumpai bunga hermaprodit pada tanaman kelapa sawit muda yang berumur sekitar 2-4 tahun, bunga ini akan menyusut atau hilang sejalan dengan bertambahnya umur tanaman. Pada setiap ketiak pelepah daun kelapa sawit tumbuh hanya satu tandan bunga, dapat berupa bunga jantan atau bunga betina. Periode antesis bunga jantan dan reseptif bunga betina tidak bersamaan sehingga memungkinkan terjadinya penyerbukan silang antar pohon kelapa sawit. Buah kelapa sawit merupakan buah batu yang terdiri atas kulit buah, daging buah, cangkang dan inti yang tersusun dalam satu tandan (Hartley 1988).
Berdasarkan ketebalan cangkangnya, kelapa sawit dapat dibedakan menjadi kelapa sawit tipe Dura, Pisifera, dan Tenera dengan ciri-ciri sebagai berikut:
a) Dura: persentase mesokarp terhadap buah bervariasi antara 35-55%, meskipun ada yang mencapai 65%; ketebalan cangkang 2-8 mm; tidak mempunyai lingkar serabut di sekeliling inti; inti relatif besar dan rendemen minyak relatif rendah (17-18%). Dura sangat baik digunakan sebagai induk betina dalam produksi benih komersial (Hartley 1988).
c) Tenera: merupakan hasil persilangan Dura dengan Pisifera; banyak ditanam secara komersil di perkebunan dan mempunyai karakteristik gabungan dari kedua induk dura dan pisifera. Ketebalan cangkang 0.4-4 mm; di sekelilingnya ada lingkar serabut dan perbandingan mesokarp terhadap buahnya cukup tinggi mencapai (60-96%). Tenera menghasilkan tandan relatif lebih banyak dibandingkan dengan Dura, walaupun ukuran tandannya lebih kecil dari Dura. Rendemen minyak mencapai 22-24% (Soehardjo et al. 1996). Tenera merupakan tanaman kelapa sawit komersial yang ditanam untuk menghasilkan minyak sawit.
Pemuliaan Kelapa Sawit
Program pemuliaan kelapa sawit di Indonesia dimulai pada tahun 1910-an, menggunakan material tanaman secara terbatas dari empat bibit kelapa sawit induk varietas Dura yang ditanam pertama kali di Kebun Raya Bogor pada tahun 1848 (Pamin 1998). Berbagai penelitian pemuliaan terus dilakukan untuk menghasilkan tanaman kelapa sawit yang ideal sesuai harapan pengusaha perkebunan terkait produktivitas maupun konsumen minyak terkait kualitas minyak sawit tersebut.
Menurut Asmono et al. (2005), pemuliaan kelapa sawit di Indonesia umumnya ditujukan untuk menghasilkan bahan tanaman kelapa sawit unggul yang memiliki produktivitas minyak tinggi dan karakteristik sekunder (auxiliary traits) tertentu dan spesifik misalnya kualitas minyak, fenologi, ketahanan terhadap cekaman biotik atau cekaman abiotik. Kualitas minyak yang dimaksud adalah perbaikan kandungan CPO, kandungan asam laurat pada minyak inti sawit (PKO), perbaikan asam lemak tak jenuh ALTJ, β karoten, tokoferol serta tokotrienol.
Mikrosatelit
Tanaman menyimpan informasi genetiknya di dalam genom inti dan organel, yaitu kloroplas dan mitokondria.Beberapa mekanisme seperti delesi, inversi, translokasi, dan transposisi yang dapat terjadi secara alami maupun diinduksi, dapat menyebabkan terjadinya penggantian atau perubahan basa nukleotida pada sekuen DNA. Mekanisme perubahan tersebut tidak selalu mengubah fenotipe tanaman sehingga penggunaan marka morfologi menjadi terbatas pemanfaatannya, sedangkan marka DNA yang langsung berinteraksi dengan sistem genetik lebih mencerminkan keadaan genom sesungguhnya. Pemanfaatan marka DNA memberikan alternatif analisis keragaman genetik (DNA fingerprinting) yang lebih baik terutama untuk karakterisasi populasi tanaman karena mampu menyediakan polimorfisme pita DNA dalam jumlah yang banyak, konsisten, dan tidak dipengaruhi lingkungan (Putri 2010).
besar dan panjang DNA-nya dapat mencapai 100 juta bp. DNA minisatelit, rata-rata sekuen berulangnya sekitar 15 bp dan dijumpai pada klaster yang mengandung 1000-3000 pengulangan.
DNA mikrosatelit terdiri atas sekuen-sekuen pendek dengan ukuran 2-5 bp dan berada pada klaster yang rata-rata pengulangannya maksimum 100 kali (Karp 1996). Pola ulangan DNA mikrosatelit terdiri atas pola di-, tri- atau tetranukleotida berulang. Pola ini ditemukan pada semua organisme, baik prokariot atau eukariot. Ulangan dinukleotida yang paling sering ditemukan pada tanaman adalah AA/TT dan AT/TA, sedangkan pada hewan GT/AC (Hoelzel 1998).
Mikrosatelit, yang juga dikenal sebagai Simple Sequece Repeats (SSRs). SSR memiliki pengulangan tandem yang pendek dari urutan DNA (2-6 bp) dan sangat polimorfik karena variasi dalam jumlah unit berulang. SSR diwariskan dalam pola yang kodominan dan sangat terwariskan. Di samping itu, SSR mudah diskoring dan dapat diproduksi dengan cepat menggunakan teknologi PCR (Bindu et al. 2004). Billotte et al (2005) berkolaborasi dengan MPOB telah mengembangkan 390 mikrosatelit pada kelapa sawit.
Penerapan Metode Mikrosatelit pada Kelapa Sawit
Bilotte et al (2001) melaporkan hasil pengembangan marka SSR kelapa sawit yang dilakukan secara bertahap, mulai dari penapisan pustaka SSR yang diperkaya dengan unit pengulangan (GA)n, (GT)n, dan (CCG)n, sampai kepada karakterisasi akhir 21 lokus SSR. Juga telah dilaporkan tentang sekuen primer, perkiraan kisaran ukuran alel, dan heterosigositas yang diharapkan pada E. guineensis dan E. oleifera. Isolasi dan karakterisasi mikrosatelit pada E. guineensis telah dilakukan oleh Cheruku et al (2009).
Populasi Half-Sib
Baro et al. (2000) menyajikan hasil untuk analisis sensitivitas pada deteksi QTL menggunakan pool DNA yang dipilih dari populasi half-sib. Simulasi menunjukkan bahwa deteksi konklusif dapat dicapai dengan famili setidaknya 500 half sib jika tetua jantan yang dipilih pada kriteria bahwa sebagian besar marka alel hilang atau xed.
Bovi et al. (2003) melakukan penelitian half-sib pada kelapa sawit King dan menyimpulkan bahwa model sistem kawin acak harus digunakan ketika memperkirakan parameter genetik. Setidaknya 50% dari selfing terjadi pada saat penyerbukan dalam tetua betina. Multi-efek indeks, menggunakan semua efek acak dari model linear menyediakan pilihan dengan akurasi tertinggi utuk setiap sifat, apapun tingkat heritabilitas idividu, dan harus digunakan untuk program pemuliaan tanaman kelapa sawit King. Di bawah model allogamous, ukuran efektif dari populasi yang diteliti adalah setara dengan 93 individu yang tidak berhubungan. Ukuran ini dikurangi menjadi 32 di bawah model campuran, bahkan begitu populasi ini memiliki variabilitas yang cukup untuk memungkinkan kemajuan genetik melalui seleksi.
Bulk Segregant Analysis (BSA)
Teknik bulked segregant analysis dapat digunakan untuk menentukan gen (gene tagging). Dalam metode BSA, DNA di-pool (digabung) dari individu-individu yang memiliki kesamaan fenotipe. Sebagai contoh, untuk mencari marka molekuler yang terpaut dengan karakter ketahanan terhadap penyakit dalam suatu populasi bersegregasi, dibuat dua bulk sample DNA; salah satu bulk terdiri atas DNA tanaman-tanaman yang tahan dan bulk kedua adalah DNA pool dari tanaman-tanaman yang peka. Perbedaan (polymorphism) suatu marka antara kedua bulk diduga terpaut dengan karakter ketahanan penyakit tersebut. BSA semula digunakan untuk karakter kualitatif, tetapi kesederhanaan metode dan biaya analisis yang relatif murah telah mendorong penggunaannya untuk karakter-karakter kuantitatif (Mackay and Caligari 2000).
Michelmore (1991) menggunakan metode BSA menggunakan marka SSR untuk menemukan marka yang berkaitan dengan gen ketahanan terhadap cekaman suhu tinggi. Marka SSR kemudian diskrining pada tetua dan kedua sample DNA bulk, dimana beberapa kombinasi primer mengungkapkan pita yang polimorfik, tidak hanya di kalangan genotipe tetua, tetapi juga antar sepasang DNA bulk. Berdasarkan evaluasi dari DNA bulk, tanaman F2 dianalisis dengan primer ko-segregasi untuk mengkonfirmasi marka yang terpaut dengan sifat ketahanan terhadap suhu tinggi. Hasil yang diperoleh Barakat et al. (2012) pada tanaman gandum menunjukkan bahwa marka SSR yang dikombinasikan dengan metode BSA dapat digunakan untuk mengidentifikasi marka molekuler yang terkait dengan sifat ketahanan tersebut. BSA dapat mengukur variasi yang ada di dalam pool segregants yang telah diurutkan sesuai dengan fenotip dan menggunakan korelasi antara pengukuran dan pool fenotip untuk menetapkan lokasi peta yang mungkin (Brauer et al 2006).
untuk mendeteksi gen dengan pengaruh yang besar bisa sama seperti yang diperoleh dengan genotiping individu yang selektif (Semagn et al. 2010). Marka yang terpaut juga dapat teridentifikasi dengan membandingkan pool homozigot (AA atau aa) dengan DNA tetua. Marka kodominan akan menghasilkan kedua pita tetua di pool homozigot jika tidak terpaut dengan gen target, namun akan menghasilkan hanya satu pita tetua jika terpaut. Dengan demikian, marka akan memunculkan pita yang polimorfik dalam pool homozigot jika tidak terpaut dengan gen target, tetapi monomorfik jika terpaut. Oleh karena itu, membangun pool homozigot adalah kunci untuk BSA (Wu and Huang 2006).
Analisis Statistik untuk Data Molekuler
Metode statistik dan multivariat secara luas digunakan untuk analisis pola genotipe DNA dalam studi keragaman tanaman. Analisis koordinat utama (Principal coordinate analysis/PCoA) merupakan metode multivariat yang umum digunakan untuk studi keragaman genetik. PCoA juga digunakan untuk menunjukkan penyebaran posisi relatif dari materi yang diuji dalam dua atau tiga dimensi sehingga jarak geometrikal sejumlah materi yang diuji dapat merefleksikan jarak genetik di antara materi tersebut dengan sedikit distorsi. Pengelompokan materi dalam plot sebaran akan menunjukkan set berdasarkan kemiripan genetik secara individu (Mohammadi and Prasanna 2003).
3
KERAGAMAN GENETIK KARAKTER PRODUKSI
POPULASI DxP PADA FAMILI PATERNAL HALF-SIB
KELAPA SAWIT BERDASARKAN MARKA DNA SSR
ABSTRAK
Pemuliaan kelapa sawit untuk memperoleh tanaman dengan produksi yang tinggi memerlukan waktu yang sangat panjang. Marka mikrosatelit dapat digunakan untuk menjelaskan keragaman genetik individu dan untuk memilih individu dengan sifat yang diinginkan. Tujuan penelitian ini adalah (1) untuk mengestimasi keragaman genetik karakter produksi populasi DxP dalam famili paternal half-sib kelapa sawit berdasarkan marka SSR dan (2) mencari kandidat marka spesifik untuk karakter minyak per tandan (oil to bunch/OB), jumlah tandan (bunch number/BN), produksi minyak (oil yield/OY) dan tandan buah segar (fresh fruit bunches/FFB). Rata-rata persentase lokus polimorfik adalah 52,78% dengan rata-rata nilai PIC 0,51. Dari hasil analisis mikrosatelit diperoleh satu lokus spesifik terhadap karakter OB, empat lokus spesifik terhadap karakter BN, satu lokus spesifik terhadap karakter OY dan dua lokus spesifik terhadap karakter FFB. Analisis klaster menunjukkan kemiripan genetik pada 56-100%, yang memisahkan keempat karakter berdasarkan bulknya, sebagaimana yang dikonfirmasi menggunakan Principal Coordinate Analysis (PCoA).
Kata kunci: kelapa sawit, keragaman genetik, pengelompokan populasi, lokus spesifik.
ABSTRACT
Oil palm breeding to obtain plants with high production requires a very long time. Microsatellite markers can be used to describe an individual's genetic diversity and to select individuals with the trait of interest. The purpose of this study was (1) to estimate the genetic diversity of DxP population based on yield component in the oil palm paternal half-sib families based on SSR markers and (2) search for a specific candidate markers for the character of oil to bunch (OB), bunch number (BN), oil yield (OY) and fresh fruit bunches (FFB). The average percentage of polymorphic loci was 52.78% with an average PIC value is 0.51. From the results obtained by the analysis of microsatellite, one marker specific to the character of OB, four markers specific to the character of BN, one marker specific to the character of OY and two markers specific to the character of FFB. Cluster analysis showed 56-100% of genetic similarity, which separates the four characters based on the bulk, as well as confirmed by Principal Coordinate Analysis (PCoA).
10
PENDAHULUAN
Kelapa sawit (Elaeis guineensis Jacq.) merupakan komoditi perkebunan yang mempunyai peran penting di Indonesia, khususnya dalam aspek perekonomian dalam negeri. Indonesia saat ini merupakan negara pengahasil Crude Palm Oil (CPO) terbesar di dunia (Direktorat Jenderal Perkebunan 2010). Selain itu Indonesia memiliki keunggulan sebagai salah satu negara yang sangat potensial untuk menanamkan investasi di bidang perkebunan dan industri hilir kelapa sawit.
Hartley (1988) menyatakan bahwa pemuliaan kelapa sawit bertujuan meningkatkan kuantitas minyak sawit dan minyak inti. Lubis (1992) dan Pamin (1999) menyebutkan tujuan lain dari pemuliaan kelapa sawit adalah untuk mendapatkan varietas dengan produksi minyak yang tinggi. Peningkatan peran kelapa sawit tidak terlepas dari kontribusi pemuliaan tanaman dalam mendukung penyediaan bahan tanaman unggul. Usaha merakit bahan tanaman kelapa sawit unggul sangat ditentukan oleh ketersediaan bahan dasar plasma nutfah dan variabilitas genetiknya. Informasi yang berkaitan dengan berbagai karakter morfologi dan genetik diperlukan dalam mendukung upaya pemberdayaan potensi plasma nutfah pada program seleksi. Informasi genetik sangat bermanfaat untuk memberikan kelengkapan informasi tanaman dan mampu mencerminkan potensi setiap individu (Asmono 1998).
Bahan tanaman unggul kelapa sawit yang banyak digunakan merupakan hibrida persilangan Dura x Pisifera, atau lazim disebut DxP. Untuk menghasilkan bahan tanaman yang unggul diperlukan material genetik yang berkualitas dan beragam, serta strategi pemuliaan yang efektif dan berkelanjutan. Pemuliaan kelapa sawit dikarakterisasi menggunakan skema reciprocal recurrent selection yang menggunakan dua tipe populasi awal, Dura dan Pisifera, untuk persilangan dan pengujian progeni, darimana tetua terbaik akan dipilih, dan benih tipe Tenera diproduksi (Corley and Tinker 2003).
Kelapa sawit secara alami merupakan tanaman tahunan dengan siklus generatif yang panjang, oleh karena itu pemuliaan konvensional dapat memakan waktu beberapa tahun. Hal tersebut sangat menghambat kemajuan dan efisiensi dalam pemilihan individu. Berbagai teknik biologi molekuler telah tersedia saat ini untuk mendeteksi variabilitas genetik dan untuk membangun hubungan kemiripan genetik antara individu-individu dan dapat digunakan dalam berbagai aplikasi dalam program pemuliaan.
Tujuan penelitian ini adalah (1) untuk mengestimasi keragaman genetik karakter produksi populasi DxP dalam famili paternal half-sib kelapa sawit berdasarkan marka SSR dan (2) mencari kandidat marka spesifik untuk karakter minyak per tandan (oil to bunch/OB), jumlah tandan (bunch number/BN), produksi minyak (oil yield/OY) dan tandan buah segar (fresh fruit bunches/FFB).
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di laboratorium Genomic and Traanscriptomic, Plant Production and Biotechnology Division, PT SMART Tbk., Sentul. Pengambilan sampel tanaman dilaksanakan di kebun percobaan Indra Sakti Estate, Pekanbaru. Waktu pelaksanaan penelitian dimulai dari bulan Agustus 2012 hingga November 2013.
Bahan Tanaman dan Isolasi DNA
Tanaman yang digunakan adalah sampel daun muda kelapa sawit yang terdiri dari 160 tanaman progeni half-sib (Dura x Pisifera) milik PT SMART Tbk yang ditanam di Pekanbaru. Tetua betina merupakan dua tanaman saudara kandung (sibs) hasil selfing dari Dura Deli, dan tetua jantan merupakan Pisifera AVROS.
DNA genom total diperoleh dengan menggerus sampel menggunakan Nitrogen cair, kemudian sampel diisolasi menggunakan GenElute Plant Genomic DNA Miniprep Kit (Sigma-Aldrich 2008) mengikuti instruksi pabrik. Kualitas DNA diketahui melalui elektroforesis menggunakan gel agarose 1%, dan kuantitas DNA diperkirakan menggunakan NanoDrop HND-1000 spektrofotometer (NanoDrop Technologies Inc.), dan terakhir kosentrasi DNA diencerkan hingga 20 ng μL-1.
Pemilihan Populasi
Populasi tanaman kelapa sawit tersebut diamati karakter-karakter yang terkait dengan produksi, dianalisis untuk mengetahui tingkat keragaman populasi. Sebanyak empat karakter yang diamati, yaitu jumlah tandan buah segar (bunch number/BN), berat tandan buah segar (fresh fruit bunches/FFB), jumlah minyak per tandan (oil to bunch/OB), dan produksi minyak (oil yield/OY). Pengamatan dilakukan selama 10 tahun. Analisis keragaman dan pengklasifikasian berdasarkan nilai terendah dan tertinggi karakter produksi tersebut, menjadi dasar penentuan tanaman yang digunakan dalam seleksi primer SSR menggunakan metode Bulk Segregant Analysis (BSA).
Amplifikasi PCR
Metode standar isolasi DNA dan evaluasi parameter PCR untuk amplifikasi mikrosatelit adalah tahap pertama penelitian yang melibatkan analisis keempat karakter yang digunakan. Seleksi primer dilakukan dengan menyeleksi lokus yang dapat membedakan bulk yang tinggi dan rendah. Primer-primer yang polimorfik pada setiap bulk di masing-masing lokus kemudian diuji untuk setiap individu di dalam bulk.
Sebanyak 80 sampel progeni terseleksi dievaluasi menggunakan Polymerase Chain Reaction (PCR) dengan 50 marka mikrosatelit sesuai dengan kondisi amplifikasi yang dilaporkan oleh Billotte et al. (2001). Polyacrylamide Gel Electrophoresis (PAGE) 6%w/v digunakan untuk visualisasi produk hasil amplifikasi PCR dengan metode pewarnaan mengikuti Benbouza et al. (2006). Pita DNA yang didapat dari lokus SSR diskor secara manual sesuai metode Allou et al. (2008).
Analisis Data
Analisis berdasarkan hasil skoring pita DNA pada gel acrylamid. Pita diskor secara manual sebagai data biner dengan ada (1) atau tidak (0) pita. Kemudian data di eksport berdasarkan permintaan perangkat lunak yang digunakan.
Tabel 1. Daftar primer beserta urutan basa nukleotida dan suhu annealing yang digunakan Kode
Primer Locus Forward marker (5' - 3') Reverse Marker (5' - 3')
Suhu annealing
(Tm) ˚C
P2 mEgCIR3847 CGTTAGTGTCGCTTATTATG AAATGAGGAAGCGTAAC 51
P3 mEgCIR3574 AGAGACCCTATTTGCTTGAT GACAAAGAGCTTGTCACAC 51
P4 mEgCIR0878 CAAAGCAACAAAGCTAGTTAGTA CAAGCAACCTCCATTTAGAT 51
P5 mEgCIR2518 GATCCCATGGTAAAGACT AAGCCTCAAAAGAAGACC 51
P6 mEgCIR2569 TAGCCGCACTCCCACGAAGC CCAGAATCATCAGACTCGGACAG 51
P8 mEgCIR2427 GAAGGGGCATTGGATTT TACCTATTACAGCGAGAGTG 51
P9 mEgCIR0783 GAATGTGGCTGTAAATGCTGAGTG AAGCCGCATGGACAACTCTAGTAA 51
P11 mEgCIR0059 TGCAGGGGATGCTTTTATT CCCTTAATTCCTGCCTTATT 51
P12 mEgCIR3869 CCAATGCAGGGGACATT GAAGCCAGTGGAAAGATAGT 51
P15 mEgCIR3519 CCACTGCTTCAAATTTACTAG GCGTCCAAAACATAAATCAC 51
P16 mEgCIR3639 ACGTTTTGGCAACTCTC ACTCCCCTCTTTGACAT 51
P18 mEgCIR0134 AGTTTGGGGCTTACCTG CTTCCACGCACCCTCTC 51
P19 mEgCIR0046 AGCCTTAGTATTTTGTTGAT CCTCTGATTTGTCCTTTTGG 51
P21 mEgCIR3890 GTGCAGATGCAGATTATATG CCTTTAGAATTGCCGTATC 51
P25 mEgCIR3389 GTCCATGTGCATAAGAGAG CTCTTGGCATTTCAGATAC 51
P26 mEgCIR3543 GTTCCCTGACCATCTTTGAG GTCGGCGATTGATTAGATTC 51
P28 mEgCIR0779 AATGCAGACCAAGCTAATCATATAC GTTCAGGTGATGGTGACTCAGATAG 51
P29 mEgCIR2144 ACAAGGCTCTTCAAGAGAT CCACTGCCAACACTAGTAC 51
P31 mEgCIR3555 CATCAGAGCCTTCAAACTAC AGCCTGAATTGCCTCTC 51
P32 mEgCIR3569 AAGGCTTGGAGTTGAGGTAT CACCATTGCATCATTATTCC 51
P34 mEgCIR3809 CCTTGCATTCCACTATT AGTTCTCAAGCCTCACA 51
P35 mEgCIR3275 GAAGCCTGAGACCGCATAGA TTCGGTGATGAAGATTGAAG 51
P36 mEgCIR2590 GTAGTTAAGGGACTTGTAGTC AAGTCTCTTGTGCTGATACA 51
P37 mEgCIR3260 AGGGCAAGTCATGTTTC TATAAGGGCGAGGTATT 51
P38 mEgCIR0801 TGGTTGGCAGGTATTATTAG TTAGAGGCTGTGATGAGTTG 51
P39 mEgCIR0521 GTGACTTTGGGCTGAAT ACAGCATCTCCAACTCTATC 51
HASIL DAN PEMBAHASAN
Uji Signifikansi Individu di dalam Bulk
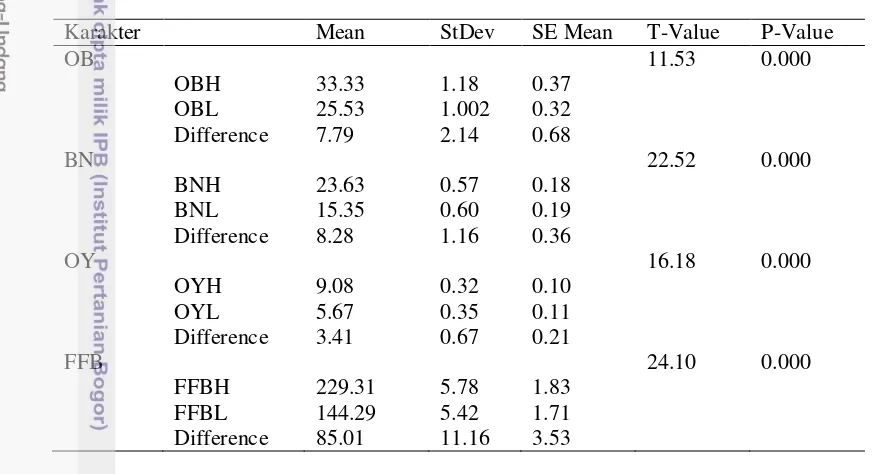
[image:31.595.68.508.238.475.2]Dari hasil analisis uji t (Tabel 2) dapat dilihat bahwa keempat pasang karakter yang dianalisis menunjukkan hasil yang signifikan, yang berarti bahwa ada perbedaan yang nyata antara bulk tinggi dan rendah pada keempat karakter yang dianalisis. Hal ini mengindikasikan bahwa metode bulk segregant analysis yang digunakan pada penelitian ini sudah benar. Hasil analisis uji t pada keempat karakter dapat dilihat pada Lampiran 1.
Tabel 2. Rekapitulasi uji t dari keempat karakter yang dianalisis
Karakter Mean StDev SE Mean T-Value P-Value OB
OBH 33.33 1.18 0.37 11.53 0.000
OBL 25.53 1.002 0.32
Difference 7.79 2.14 0.68 BN
BNH 23.63 0.57 0.18 22.52 0.000
BNL 15.35 0.60 0.19
Difference 8.28 1.16 0.36 OY
OYH 9.08 0.32 0.10 16.18 0.000
OYL 5.67 0.35 0.11
Difference 3.41 0.67 0.21 FFB
FFBH 229.31 5.78 1.83 24.10 0.000 FFBL 144.29 5.42 1.71
Difference 85.01 11.16 3.53
Analisis Mikrosatelit dan Polimorfisme
Hasil yang diperoleh menunjukkan bahwa dari 50 primer yang dievaluasi, terdapat 27 primer polimorfik yang digunakan untuk analisis selanjutnya. Sebanyak 15 primer polimorfik terdapat pada karakter OB, 18 primer pada karakter BN, 11 primer pada karakter OY, dan 13 primer pada karakter FFB (Gambar 2).
Dari keempat karakter yang diamati, persentase lokus polimorfik terbesar sampai terkecil berturut-turut adalah OB 55,56%, BN 66.67%, OY 40.74% dan FFB 48,15%, dengan rata-rata 52,78%. McGregor et al. (2000) menyatakan bahwa polimorfisme merupakan gambaran amplifikasi yang diperoleh dari perbedaan fragmen DNA yang diobservasi dan diskor sebagai ada atau tidaknya perbedaan sekuen sehingga menunjukkan ada tidaknya variasi.
(a) OY – pada lokus P39
[image:32.595.51.497.148.763.2](b) BN – pada lokus P21
Gambar 2. Pola amplifikasi dua marka mikrosatelit dari bahan genetik kelapa sawit dengan 4 alel (a) lokus monomorfik; (b) lokus polimorfik. 10 lokus paling kiri merupakan individu di dalam bulk dengan produksi rendah, dan 10 lokus paling kanan merupakan individu di dalam bulk dengan produksi tinggi
Polymorphism Information Content (PIC)
Dari hasil analisis menggunakan 27 primer pada empat karakter, diperoleh nilai Polymorphism Information Content antara 0,29-0,68 (Tabel 3).
Tabel 3. Jumlah alel dan Polymorphism Information Content (PIC) pada mikrosatelit yang dievaluasi
Kode
Primer Locus
Jumlah alel teramplifikasi PIC
OB BN OY FFB
P2 mEgCIR3847 3 0 0 4 0.67
P3 mEgCIR3574 0 4 0 0 0.67
P4 mEgCIR0878 2 0 0 3 0.51
P5 mEgCIR2518 0 2 3 0 0.53
P6 mEgCIR2569 3 0 3 0 0.58
P8 mEgCIR2427 0 2 3 0 0.50
P9 mEgCIR0783 3 3 0 3 0.56
P11 mEgCIR0059 2 2 2 0 0.37
P12 mEgCIR3869 0 3 0 0 0.49
P15 mEgCIR3519 3 2 0 2 0.53
P16 mEgCIR3639 0 0 3 0 0.43
P18 mEgCIR0134 3 0 0 3 0.55
P19 mEgCIR0046 3 3 0 0 0.51
P21 mEgCIR3890 3 4 3 0 0.66
P25 mEgCIR3389 0 0 0 3 0.48
P26 mEgCIR3543 3 0 0 0 0.51
P28 mEgCIR0779 0 4 0 0 0.68
P29 mEgCIR2144 2 3 3 3 0.54
P31 mEgCIR3555 0 0 2 3 0.43
P32 mEgCIR3569 0 0 0 3 0.56
P34 mEgCIR3809 0 2 0 0 0.36
P35 mEgCIR3275 3 4 0 0 0.57
P36 mEgCIR2590 2 2 3 2 0.38
P37 mEgCIR3260 2 3 2 3 0.44
P38 mEgCIR0801 0 3 2 3 0.52
P39 mEgCIR0521 3 3 0 3 0.52
Rata-rata nilai PIC yang diperoleh sebesar 0,51. Nilai ini mengindikasikan bahwa marka SSR tersebut cukup informatif digunakan untuk melihat keragaman antar empat karakter yang diuji. Walaupun begitu, nilai ini lebih kecil daripada yang didapatkan Thongthawee et al. (2010) pada populasi tetua kelapa sawit, yaitu 0,58-0,82, dan Ajambang et al. (2012) yang memperoleh rata-rata nilai PIC sebesar 0,60. Nilai PIC yang lebih kecil ini diakibatkan oleh populasi yang digunakan dalam penelitian ini memiliki keragaman genetik yang cukup dekat, sehingga variasi dan frekuensi alel yang ditemukan tidak sebanyak pada populasi yang keragaman genetiknya jauh. Hal ini sesuai dengan pernyataan DeVicente and Fulton (2003) yang menyatakan bahwa nilai polimorfisme ditentukan oleh frekuensi kemunculan alelnya. Nilai PIC diperlukan untuk memilih marka yang dapat membedakan antar individu yang digunakan. Kuantifikasi PIC didasarkan pada jumlah alel yang dapat dihasilkan oleh suatu marka dan frekuensi dari tiap alel dalam set genotipe yang diuji.
Botstein et al. (1980) mengklasifikasikan nilai PIC menjadi tiga kelas, yaitu PIC>0.5 adalah sangat informatif, 0.25>PIC>0.5 termasuk sedang dan PIC<0.25 memiliki nilai informatif yang rendah. Menurut Anderson et al. (1993), semakin besar nilai PIC dalam suatu marka, maka semakin baik marka tersebut digunakan sebagai marka molekuler. Terdapat 18 dari 28 marka yang memiliki nilai PIC > 0.5, yang berarti marka-marka tersebut dapat diseleksi untuk studi keragaman kelapa sawit.
Jumlah alel (Na), jumlah alel efektif (Ne), Indeks Informasi (I), heretozigositas teramati (Ho), Heterozigositas harapan (He) dan indeks Fiksasi (F)
Ne adalah sebuah ukuran dari jumlah alel efektif yang diperoleh dari masing-masing karakter. Nilai ini adalah nilai resiprok atau nilai kebalikan dari homozigositas. Semakin tinggi nilai Ne, maka semakin banyak individu yang heterozigot. Jumlah alel per lokus tertinggi, terdapat pada karakter BN yaitu 2,833 dan terendah pada karakter OB yaitu 2,667. Sedangkan jumlah alel efektif tertinggi terdapat pada karakter FFB yaitu 2,493 dan terendah pada karakter OB yaitu 2,307 (Tabel 4). Jumlah ini lebih rendah daripada yang didapatkan Zulhermana et al. (2010) (3,67), Agustina et al. (2010), (3,85), Ajambang et al. (2010), (4,71), Billotte et al. (2001), (5.3), tetapi lebih tinggi dari yang diperoleh Maizura et al. (2006), (1,9) dan Allou et al. (2008), (1,75).
Tabel 4. Rata-rata jumlah alel (Na), jumlah alel efektif (Ne), Indeks Informasi (I),
heretozigositas teramati (Ho), Heterozigositas harapan (He) dan indeks Fiksasi (F) pada setiap karakter
Pop Na Ne I Ho He F
OB Mean 2,67 2,31 0,88 0,82 0,55 -0,49 SE 0,13 0,11 0,05 0,05 0,03 0,06 BN Mean 2,83 2,37 0,89 0,80 0,55 -0,47
SE 0,19 0,16 0,06 0,05 0,03 0,08 OY Mean 2,72 2,35 0,88 0,84 0,54 -0,50
SE 0,14 0,17 0,07 0,08 0,04 0,06 FFB Mean 2,92 2,50 0,95 0,88 0,57 -0,49
SE 0,14 0,17 0,06 0,07 0,03 0,07
polen dari Pisifera terpilih. Upaya-upaya perbaikan genetik kelapa sawit ini menyebabkan keragaman genetik menyempit, sehingga alel yang diperoleh juga semakin sedikit. Bakoume et al. (2007) menemukan bahwa alel pada populasi liar kelapa sawit menunjukkan penurunan yang disebabkan oleh seleksi bertahun-tahun pada bahan genetik. Arias et al. (2012) menggambarkan hasil ini sebagai kecenderungan umum kehilangan keragaman genetik yang disebabkan oleh perbaikan genetik pada kelapa sawit.
Secara umum, keempat karakter menunjukkan nilai rata-rata heterozigositas teramati (Ho) lebih tinggi daripada nilai rata-rata heterozigositas harapan (He). Ini berarti bahwa setiap karakter memiliki heterozigositas yang tinggi.
Keragaman Genetik Antar dan Intra Populasi
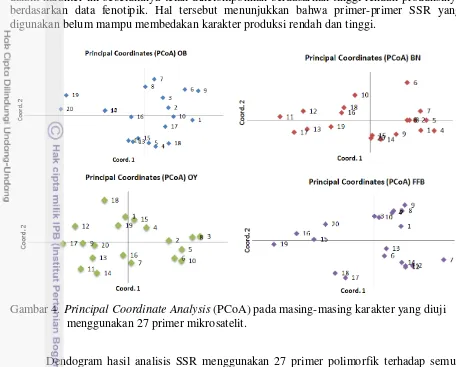
Hasil PCoA empat karakter menunjukkan informasi dari empat kelompok berdasarkan kesamaan genetiknya. Karakter-karakter ini memiliki keragaman genetik yang tinggi di dalam kelompoknya, tetapi keragaman genetik rendah antar karakter. Karakter OB membentuk kelompok sendiri, demikian juga pada BN, OY dan FFB. Persentase keragaman dijelaskan oleh tiga sumbu pertama, yaitu 31.95%, 25.23% dan 17.66% (Gambar 3).
Dari hasil PCoA dapat dilihat bahwa tiga sumbu pertama pada individu di dalam bulk menjelaskan variasi kumulatif sebesar 74,85% dari data. Hal ini menunjukkan bahwa hanya terdapat 26,15% variasi yang tidak dapat dijelaskan dari data tersebut. Peakall and Smouse (2012) menyatakan bahwa PCoA adalah suatu proses dimana sumbu utama variasi terletak pada set data multidimensi. Setiap sumbu berturut-turut secara proporsional menjelaskan lebih sedikit dari variasi total, sehingga ketika ada kelompok yang berbeda, dua atau tiga sumbu pertama akan mengungkapkan sebagian besar pemisahannya.
PCoA (Principal Coordinate Analysis) adalah skala atau metode ordinasi yang dimulai dengan matriks kesamaan atau ketidaksamaan antara satu set individu dan bertujuan untuk menghasilkan plot grafis dengan dimensi rendah dari data, sehingga jarak geometris antara individu-individu dalam plot mencerminkan jarak genetik antara mereka dengan distorsi minimal. Agregasi individu dalam plot seperti itu akan mengungkapkan set genetik individu yang serupa (Melchinger 1993).
Gambar 3. Principal Coordinate Analysis (PCoA) dari empat karakter produksi kelapa sawit
Dari hasil PCoA pada masing-masing karakter (Gambar 4), dapat dilihat bahwa individu nomor 1-10 belum sepenuhnya terpisah dari individu nomor 11-20. Individu di
12 6534 7 8 910 11 12 1415161713192018 21 22 23 24 2526 27 2829 30 31 32363435 33
37 38 3940 4142 43 44 454647 4850515552495354
56 5758 5960 61 6263 64 65 66 67 6871747572737769707876
79 80 Co o rd . 2 Coord. 1
Principal Coordinates (PCoA)
OB
BN
OY
dalam karakter ini sebelumnya telah dikelompokkan berdasarkan tinggi-rendah produksinya berdasarkan data fenotipik. Hal tersebut menunjukkan bahwa primer-primer SSR yang digunakan belum mampu membedakan karakter produksi rendah dan tinggi.
Gambar 4. Principal Coordinate Analysis (PCoA) pada masing-masing karakter yang diuji menggunakan 27 primer mikrosatelit.
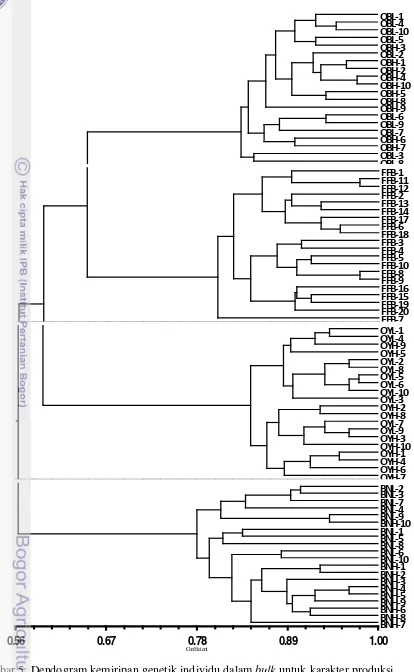
Dendogram hasil analisis SSR menggunakan 27 primer polimorfik terhadap semua individu dalam bulk untuk karakter produksi sawit menunjukkan pengelompokan sesuai empat karakter produksi yaitu OB, BN, OY dan FFB dengan tingkat kemiripan masing-masing sebesar 56 – 100% (Gambar 5). Hal ini menunjukkan bahwa primer-primer yang digunakan mampu menunjukkan perbedaan lokus antar karakter produksi yang diuji.
Kemiripan genetik yang tinggi (~56%) ini merefleksikan jarak genetik yang sempit antara tetua yang digunakan pada persilangan. Nilai ini dapat dijelaskan oleh beberapa faktor di antaranya: 1) dua tetua Dura Deli hasil selfing yang disilangkan dengan Pisifera AVROS yang sama, 2) pertukaran global sumber tanaman hasil program pemuliaan oleh perusahaan komersil selama beberapa tahun (Richardson 1995), dan 3) populasi asli yang terbatas, yang dijelaskan oleh beberapa peneliti (Bakoume 2007; Cochard 2009).
Gambar 5. Dendogram kemiripan genetik individu dalam bulk untuk karakter produksi jumlah tandan buah segar (bunch number/BN), berat tandan buah segar (fresh fruit bunches/FFB), jumlah minyak per tandan (oil to bunch/OB), dan produksi minyak (oil yield/OY) sawit berdasarkan marka SSR
Coefficient
0.56 0.67 0.77 0.87 0.98
OBL-10 OBL-5 OBH-3 OBL-2 OBH-1 OBH-2 OBH-4 OBH-10 OBH-5 OBH-8 OBH-9 OBL-6 OBL-9 OBL-7 OBH-6 OBH-7 OBL-3 OBL-8 Coefficient
0.56 0.67 0.78 0.89 1.00
FFB-1 FFB-11 FFB-12 FFB-2 FFB-13 FFB-14 FFB-17 FFB-6 FFB-18 FFB-3 FFB-4 FFB-5 FFB-10 FFB-8 FFB-9 FFB-16 FFB-15 FFB-19 FFB-20 FFB-7 Coefficient
0.56 0.67 0.78 0.89 1.00
OYL-1 OYL-4 OYH-9 OYH-5 OYL-2 OYL-8 OYL-5 OYL-6 OYL-10 OYL-3 OYH-2 OYH-8 OYL-7 OYL-9 OYH-3 OYH-10 OYH-1 OYH-4 OYH-6 OYH-7 Coefficient
0.56 0.67 0.78 0.89 1.00
Dari dendogram di atas, dapat dilihat bahwa dari keempat karakter, karakter OY merupakan karakter yang paling baik dalam memisahkan individu dengan produksi minyak tinggi dengan yang rendah. Walaupun masih ada dua individu yang belum mengelompok sesuai dengan bulknya (Gambar 6). Hal ini disebabkan primer-primer yang digunakan belum dapat menunjukkan kespesifikan dalam membedakan karakter produksi rendah dan tinggi. Selain itu, hasil tersebut mungkin disebabkan karakter produksi yang tergolong karakter kuantitatif yang ditentukan oleh banyak gen sehingga sulit diperoleh lokus spesifik yang dapat mengindikasikan sifat seperti pada karakter kualitatif.
Gambar 6. Dendogram kemiripan genetik pada karakter produksi minyak (oil yield/OY).
KESIMPULAN
Hasil analisis klaster menunjukkan kemiripan genetik populasi half-sib kelapa sawit pada 56-100%, yang berhasil membagi keempat populasi berdasarkan kelompoknya, sebagaimana yang dikonfirmasi menggunakan PCoA. Hasil analisis mikrosatelit menunjukkan satu marka spesifik untuk karakter OB, empat marka spesifik untuk karkter BN, satu marka spesifik untuk karakter OY, dua marka spesifik untuk karakter FFB, serta tiga marka yang dapat mengamplifikasi di keempat karakter.
Coefficient
0.84 0.88 0.92 0.96 1.00
4
IDENTIFIKASI MARKA MOLEKULER SPESIFIK UNTUK
KARAKTER PRODUKSI POPULASI DxP PADA FAMILI
PATERNAL HALF-SIB KELAPA SAWIT
ABSTRAK
Karakter produksi dan komponennya merupakan karakter utama potensi hasil. Informasi marka spesifik akan memperbaiki kecepatan dan ketepatan seleksi. Tujuan penelitian ini adalah untuk mengidentifikasi marka spesifik karakter produksi menggunakan pendekatan korelasi dan regresi. Hasil analisis menunjukkan bahwa terdapat 11 marka spesifik yang diperoleh menggunakan pendekatan korelasi, serta 16 marka spesifik yang diperoleh menggunakan pendekatan regresi stepwise. Satu marka berkorelasi dan beregresi positif terhadap OB, dua marka berkorelasi dan beregresi positif terhadap BN, dua marka beregresi positif terhadap OY, namun hanya satu marka yang berkorelasi positif terhadap OY, serta tidak ada maka yang berkorelasi maupun beregresi positif terhadap FFB. Marka-marka yang terpilih secara bersamaan berdasarkan analisis korelasi dan regresi dapat dipertimbangkan sebagai kandidat marka untuk menyeleksi karakter dengan produksi minyak tinggi.
Kata kunci: kelapa sawit, karakter produksi, marka spesifik, korelasi, regresi
ABSTRACT
Character of production and its component is the main character of the potential yield. Information about specific markers will improve the velocity and accuracy of the selection. The objective of this study was to identify specific markers of character production using correlation and regression approach. The results showed that there are 11 specific markers obtained from the correlation approach and 16 specific markers obtained from the stepswise regression approach. One marker positively correlated and regressed to the OB, two markers positively correlated and regressed to the BN, two markers positively regressed to the OY, but only one marker positively correlated to the OY, and there are no marker were positively correlated nor regressed to the FFB. Markers simultaneously selected based on correlation and regression analysis can be considered as candidate markers for selecting a character with high oil production.
22
PENDAHULUAN
Peningkatan potensi hasil merupakan tujuan utama dalam program pemuliaan kelapa sawit. Karakter produksi dan komponennya merupakan karakter utama potensi hasil. Pengembangan metode seleksi pada kelapa sawit diarahkan untuk mengidentifikasi genotipe harapan dengan waktu yang lebih cepat dan pada tahap awal pertumbuhan tanaman. Perolehan genotipe harapan pada awal pertumbuhan tanaman akan mereduksi kompleksitas dan ukuran populasi yang akan dievaluasi pada skala kebun.
Sifat agronomi yang bernilai ekonomi tinggi, seperti produksi, dikendalikan oleh banyak gen, sehingga disebut karakter kuantitatif. Karakter kuantitatif diterjemahkan sebagai karakter dengan nilai yang memiliki distribusi kontiniu yang diperoleh dari hasil pengukuran dan perhitungan. Karakter kuantitatif dikontrol oleh banyak gen dan masing-masing gen bersifat aditif terhadap karakter tersebut. Ekspresi gen-gen yang mengendalikan karakter kuantitatif sangat dipengaruhi oleh lingkungan. Pengaruh lingkungan yang besar terhadap keragaman karakter kuantitatif merupakan salah satu kesulitan yang dihadapi pemulia dalam melakukan seleksi (Falconer 1985).
Pengembangan marka molekuler menyediakan alat untuk menilai jarak genetik pada tingkat DNA pada spesies tanaman (Melchinger and Gumber 1998). Selain itu, data molekuler juga telah sering dimasukkan dalam beberapa metode untuk memprediksi penampilan hibrida. Data molekuler telah terintegrasi ke dalam model untuk memprediksi hasil dalam berbagai cara. Antara lain untuk memperhitungkan jarak genetik tetua dan analisis korelasi dengan hasil progeni (Ajmone et al. 1998)
Selama 20 tahun terakhir, upaya utama dalam pemuliaan telah berubah dari sistem seleksi tradisional berbasis silsilah fenotipik menjadi genetika molekuler dengan penekanan pada identifikasi lokus sifat kuantitatif dan seleksi dibantu marka (marker assisted selection/MAS). MAS yang menggunakan marka DNA untuk memilih genotipe optimal, merupakan alat yang sangat baik untuk memilih sifat-sifat yang menguntungkan, yang sulit untuk diukur, yang menunjukkan heritabilitas rendah dan/atau diekspresikan pada saat akhir perkembangan tanaman (Davies et al. 2006; Wilde et al. 2007), serta untuk menilai potensi genetik. Pengembangan dan implementasi dari skema MAS menggunakan marka yang diidentifikasi akan memperbaiki kecepatan dan ketepatan seleksi (Henning et al. 2011).
Namun demikian dalam interpretasi data, data molekuler sendiri tidak dapat diinterpretasi untuk aplikasi di lapangan karena data genotipik itu sendiri tidak dapat diekspresikan. Marka molekuler hanya akan terekspresi melalui data fenotipik. Oleh sebab itu diperlukan informasi fenotipik untuk mengetahui apakah data molekuler yang diperoleh dapat memberikan informasi yang lebih akurat dan lebih efektif sebagai alat bantu program pengembangan tanaman (Smith and Smith 1992).
Korelasi merupakan hasil akhir dari pengaruh gen yang bersegregasi atau faktor lingkungan yang mengendalikan karakter-karakter yang berkorelasi. Faktor genetik yang menyebabkan terjadinya korelasi (positif/negatif) antara lain adanya pleitropi, yaitu satu atau beberapa gen yang mengendalikan ekspresi lebih dari satu karakter (Ruchjaningsih et al. 2000) atau pengaruh gen komplementer lainnya (Singh 1980).
model (Lande and Thompson 1990). Signifikansi ambang batas yang dapat memaksimalkan respon seleksi tidak dapat ditentukan secara analitis (Hospital et al. 1997), namun demikian, ketika hanya efek marka signifikan yang diperkirakan, hanya sebagian dari varians genetik yang akan ditangkap dan efek dipertahankan dalam model dapat sangat berlebihan (Beavis 1998; Hayes 2007), terutama ketika banyak efek yang diuji.
Tujuan dari penelitian ini adalah untuk mengidentifikasi marka spesifik karakter produksi menggunakan pendekatan korelasi dan regresi.
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di laboratorium Genomic and Traanscriptomic, Plant Production and Biotechnology Division, PT SMART Tbk., Sentul. Pengambilan sampel tanaman dilaksanakan di kebun percobaan Indra Sakti Estate, Pekanbaru. Waktu pelaksanaan penelitian dimulai dari bulan Agustus 2012 hingga November 2013.
Bahan Tanaman dan Isolasi DNA
Tanaman yang digunakan adalah sampel daun muda kelapa sawit yang terdiri dari 160 tanaman progeni half-sib (Dura x Pisifera) milik PT SMART Tbk yang ditanam di Pekanbaru. Tetua betina merupakan dua tanaman saudara kandung (sibs) hasil selfing dari Dura Deli, dan tetua jantan merupakan Pisifera AVROS.
DNA genom total diperoleh dengan menggerus sampel menggunakan Nitrogen cair, kemudian sampel diisolasi menggunakan GenElute Plant Genomic DNA Miniprep Kit (Sigma-Aldrich 2008) mengikuti instruksi pabrik. Kualitas DNA diketahui melalui elektroforesis menggunakan gel agarose 1%, dan kuantitas DNA diperkirakan menggunakan NanoDrop HND-1000 spektrofotometer (NanoDrop Technologies Inc.), dan terakhir kosentrasi DNA diencerkan hingga 20 ng μL-1.
Pemilihan Populasi
Populasi tanaman kelapa sawit tersebut diamati karakter-karakter yang terkait dengan produksi. Sebanyak empat karakter yang diamati, yaitu jumlah tandan buah segar (bunch number/BN), berat tandan buah segar (fresh fruit bunches/FFB), jumlah minyak per tandan (oil to bunch/OB), dan produksi minyak (oil yield/OY). Pengamatan dilakukan selama 10 tahun. Hasil analisis keragaman dan pengklasifikasian berdasarkan nilai terendah dan tertinggi karakter produksi tersebut, menjadi dasar penentuan tanaman yang digunakan dalam seleksi primer SSR menggunakan metode Bulk Segregant Analysis (BSA).
Tabel 5. Daftar primer beserta urutan basa nukleotida dan suhu annealing yang digunakan Kode
Primer Locus Forward marker (5' - 3') Reverse Marker (5' - 3') Suhu annealing (Tm) ˚C
P2 mEgCIR3847 CGTTAGTGTCGCTTATTATG AAATGAGGAAGCGTAAC 51
P3 mEgCIR3574 AGAGACCCTATTTGCTTGAT GACAAAGAGCTTGTCACAC 51
P4 mEgCIR0878 CAAAGCAACAAAGCTAGTTAGTA CAAGCAACCTCCATTTAGAT 51
P5 mEgCIR2518 GATCCCATGGTAAAGACT AAGCCTCAAAAGAAGACC 51
P6 mEgCIR2569 TAGCCGCACTCCCACGAAGC CCAGAATCATCAGACTCGGACAG 51
P8 mEgCIR2427 GAAGGGGCATTGGATTT TACCTATTACAGCGAGAGTG 51
P9 mEgCIR0783 GAATGTGGCTGTAAATGCTGAGTG AAGCCGCATGGACAACTCTAGTAA 51
P11 mEgCIR0059 TGCAGGGGATGCTTTTATT CCCTTAATTCCTGCCTTATT 51
P12 mEgCIR3869 CCAATGCAGGGGACATT GAAGCCAGTGGAAAGATAGT 51
P15 mEgCIR3519 CCACTGCTTCAAATTTACTAG GCGTCCAAAACATAAATCAC 51
P16 mEgCIR3639 ACGTTTTGGCAACTCTC ACTCCCCTCTTTGACAT 51
P18 mEgCIR0134 AGTTTGGGGCTTACCTG CTTCCACGCACCCTCTC 51
P19 mEgCIR0046 AGCCTTAGTATTTTGTTGAT CCTCTGATTTGTCCTTTTGG 51
P21 mEgCIR3890 GTGCAGATGCAGATTATATG CCTTTAGAATTGCCGTATC 51
P25 mEgCIR3389 GTCCATGTGCATAAGAGAG CTCTTGGCATTTCAGATAC 51
P26 mEgCIR3543 GTTCCCTGACCATCTTTGAG GTCGGCGATTGATTAGATTC 51
P28 mEgCIR0779 AATGCAGACCAAGCTAATCATATAC GTTCAGGTGATGGTGACTCAGATAG 51
P29 mEgCIR2144 ACAAGGCTCTTCAAGAGAT CCACTGCCAACACTAGTAC 51
P31 mEgCIR3555 CATCAGAGCCTTCAAACTAC AGCCTGAATTGCCTCTC 51
P32 mEgCIR3569 AAGGCTTGGAGTTGAGGTAT CACCATTGCATCATTATTCC 51
P34 mEgCIR3809 CCTTGCATTCCACTATT AGTTCTCAAGCCTCACA 51
P35 mEgCIR3275 GAAGCCTGAGACCGCATAGA TTCGGTGATGAAGATTGAAG 51
P36 mEgCIR2590 GTAGTTAAGGGACTTGTAGTC AAGTCTCTTGTGCTGATACA 51
P37 mEgCIR3260 AGGGCAAGTCATGTTTC TATAAGGGCGAGGTATT 51
P38 mEgCIR0801 TGGTTGGCAGGTATTATTAG TTAGAGGCTGTGATGAGTTG 51
P39 mEgCIR0521 GTGACTTTGGGCTGAAT ACAGCATCTCCAACTCTATC 51
P46 mEgCIR0326 GCTAACCACAGGCAAAAACA AAGCCGCACTAACATACACATC 51
Amplifikasi PCR
Sebanyak 80 sampel progeni terseleksi dievaluasi menggunakan Polymerase Chain Reaction (PCR) dengan 50 marka mikrosatelit sesuai dengan kondisi amplifikasi yang dilaporkan oleh Billotte et al. (2001; 2005). Polyacrylamide Gel Electrophoresis (PAGE) 6%w/v digunakan untuk visualisasi produk hasil amplifikasi PCR dengan metode pewarnaan mengikuti Benbouza et al. (2006). Pita DNA yang didapat dari lokus SSR diskor secara manual sesuai metode Allou et al. (2008).
Analisis Data
Analisis dilakukan berdasarkan hasil skoring pita DNA pada gel acrylamid. Pita diskor secara manual sebagai data biner dengan ada (1) atau tidak (0) pita. Kemudian data dieksport berdasarkan permintaan perangkat lunak yang digunakan. Identifikasi marka yang berasosiasi dengan karakter yang diuji, dilakukan menggunakan pendekatan deskriptif berupa analisis korelasi dan regresi yang dijalankan menggunakan Minitab ver. 16 (Minitab, State College, PA).
HASIL DAN PEMBAHASAN
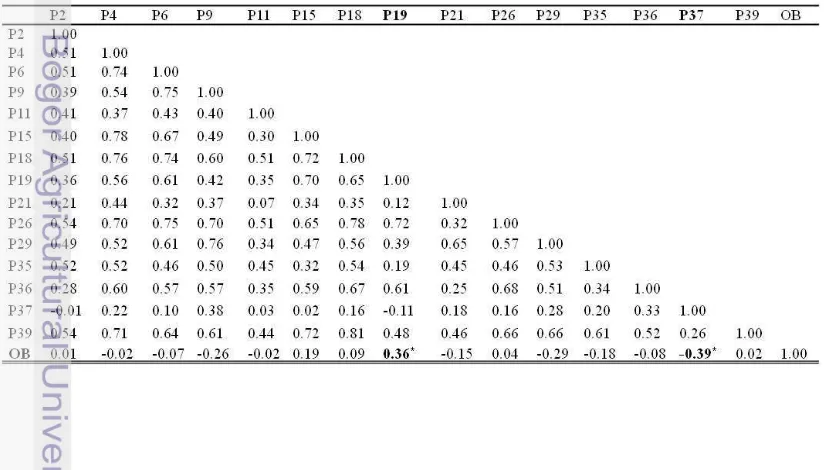
Hasil analisis korelasi antara marka SSR dengan karakter minyak per tandan (oil to bunch/OB) menunjukkan bahwa terdapat dua marka yang memiliki korelasi nyata yaitu P19 dan P37 (Tabel 6). Berdasarkan hasil regresi stepwise, terdapat tiga marka yang berkorelasi terhadap OB yaitu P19, P37 dan P29. Model regresi karakter minyak per tandan yaitu OB = 29.9 + 2.91 P19 - 1.17 P37 - 3.68 P29. Marka-marka yang
[image:42.595.90.502.564.799.2]berkorelasi nyata kemungkinan besar merupakan lokus-lokus yang berasosiasi dengan karakter OB sehingga kedua marka ini dapat diuji dan dieksplorasi lanjut pada populasi yang lebih sesuai dengan ukuran populasi minimum yang cukup dalam upaya perbaikan genetik karakter ini. Hasil analisis regresi stepwise untuk keempat karakter dapat dilihat pada Lampiran 2.
Hasil analisis korelasi menunjukkan bahwa terdapat empat marka SSR yang berkorelasi nyata terhadap jumlah tandan, yaitu P3, P29, P37, dan P46 (Tabel 7), sedangkan berdasarkan hasil analisis regresi stepwise terdapat lima marka SSR yang berkontribusi tinggi terhadap karakter jumlah tandan, yaitu P3, P29, P46, P37 dan P12. BN = 18.3 - 1.44 P3 - 1.80 P29 + 1.56 P46 + 3.81 P37 - 2.34 P12 merupakan model regresi karakter jumlah tandan. Perbedaan arah regresinya menunjukkan perbedaan alel yang spesifik untuk karakter ini. Marka-marka yang berkorelasi nyata dapat digunakan sebagai kandidat marka untuk menyeleksi genotipe-genotipe kelapa sawit yang memiliki jumlah tandan banyak.
Tabel 7. Matriks Korelasi Antar Marka dan Karakter jumlah tandan (BN)
Tabel 8. Matriks Korelasi Antar Marka dan Karakter Produksi Minyak (OY)
Terdapat tiga marka SSR yang berkorelasi nyata dengan karakter tandan buah segar (FFB), yaitu P36, P37 dan P39 (Tabel 9). Ketiga marka tersebut juga memiliki nilai koefisien regresi tertinggi terhadap karakter tandan buah segar. Model regresi karakter tandan buah segar yaitu FFB = 236 - 14.7 P36 - 9.28 P37 - 7.19 P39. Informasi marka-marka yang berasosiasi dengan masing-masing karakter produksi diringkas pada Tabel 10. Pada tabel tersebut terlihat bahwa terdapat total 11 marka SSR yang berasosiasi pada 4 karakter produksi. Marka-marka tersebut dapat diuji ulang dengan ukuran populasi yang lebih besar untuk memverifikasi apakah marka-marka tersebut betul-betul berasosiasi dengan masing-masing karakter produksi.
Tabel 9. Matriks Korelasi Antar Marka dan Karakter Tandan Buah Segar (FFB)
produksi yang diharapkan, sebaliknya marka yang berkorelasi negatif dengan individu sebaiknya dihindari, karena dikhawatirkan akan menurunkan produksi.
Tabel 10. Rekapitulasi Jumlah Marka yang Berkorelasi Nyata terhadap Masing-Masing Karakter produksi
Karakter Marka r B P
OB P19 0.36 2.91 0.002
P37 -0.39 -1.17 0.135
BN P3 -0.51 -1.44 0.001
P29 -0.44 -1.80 0.015
P37 0.31 3.81 0.001
P46 0.31 1.57 0.270
OY P36 0.42 1.54 0.000
P38 -0.48 -1.05 0.019
FFB P36 -037 -14.72 0.184
P37 -0.39 -9.28 0.203
P39 -0.31 -7.19 0.431
Dari data tersebut juga dapat dilihat bahwa terdapat marka yang berkorelasi positif terhadap suatu karakter tetapi berkorelasi negatif dengan karakter lainnya. P37 berkorelasi positif terhadap BN, tetapi berkorelasi negatif terhadap OB dan FFB. Perbedaan ini menunjukkan bahwa kehadiran marka ini pada suatu individu, akan meningkatkan jumlah tandan yang dihasilkan, tetapi menurunkan produksi minyak per tandan dan tandan buah segar yang dihasilkan. Selain itu, P36 berkorelasi positif terhadap OY, tetapi berkorelasi negatif terhadap FFB. Ini berarti bahwa kehadiran marka ini pada suatu individu, akan meningkatkan produksi minyak, tetapi menurunkan tandan buah segar yang dihasilkan. Ini sesuai dengan pernyataan Falconer (1989) yang menyatakan bahwa korelasi negatif memberi indikasi bahwa peningkatan suatu sifat akan menurunkan sifat yag lain, sedangkan korelasi positif terjadi bila peningkatan suatu sifat akan meningkatkan sifat lain yang dikorelasikan.