Kanthi Arum Widayati
GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
I hereby declare that dissertation entitled Categorization in Macaca fascicularis is original result of my own research supervised under advisory committee and has never been submitted in any form at any institution before. All information from other authors cited here are mentioned in the text and listed in the reference at the end part of the dissertation.
Bogor, February 1st, 2012
Kanthi Arum Widayati
KANTHI ARUM WIDAYATI. Categorization in Macaca fascicularis. Supervised by BAMBANG SURYOBROTO, AKICHIKA MIKAMI, ACHMAD FARAJALLAH.
Categorization is an ability to group individuals or events into different classes mediated by conceptualized mental images. There are several levels of categorization and within a taxonomy the levels are nested. At the most concrete level of categorization, all or most members of the category shared common physical attributes that differ from other categories. The higher the level of category, the fewer common attributes between members of the group. In addition to humans, the ability to categorize has also been proposed in animals. One example of categorization in animal is species discrimination. Using matching-to-sample task, present experiment tested ability of the long-tailed macaque (Macaca fascicularis) in discriminating dichotomous-stimuli of different animals. The species has been shown to be able to see photos as representations of real object so I used facial photos of humans and animals for the stimuli. First, I tested their ability to classify humans and macaques into separate group. Second, I tested their ability to discriminate their conspecific from other macaques. And the last, I tested whether the subjects were able to discriminate non-human animals from humans. In all of these experiments I found that the subjects showed high performance in categorizing objects, even when I discarded details of visual informations, such as color and local shapes. The ability to identify objects with reduced representation of physical properties means the subjects were able to generalize attributes of members of the group. This would indicate that the subjects created a higher level abstraction. On the other hand, in discriminating intrageneric macaque species I found that they were able to extract uniqueness of each species. More over, I also found that the subjects were able to put photos of non-human animals that shared very few similarities in physical percepts into one group. I suggested that the subjects could create a more abstract concept based on non-percepts relations as a basis to put the objects into one category. Thus, I concluded that M. fascicularis were able to perform multiple levels of categorizations.
KANTHI ARUM WIDAYATI. Kemampuan kategorisasi Macaca fascicularis. Dibimbing oleh BAMBANG SURYOBROTO, AKICHIKA MIKAMI, ACHMAD FARAJALLAH.
Kategorisasi adalah kemampuan seseorang untuk mengelompokkan individu-individu atau kejadian-kejadian ke dalam kelompok yang berbeda. Kemampuan kategorisasi didukung oleh konsep yang dibentuk di otak. Ada beberapa tingkat kategorisasi. Pada tingkat yang paling dasar, semua anggota kelompok memiliki banyak persamaan ciri fisik dibandingkan dengan kelompok lainnya. Semakin tinggi tingkat kategorisasi, persamaan ciri fisik di antara anggota kelompok semakin sedikit. Selain manusia, hewan diduga juga memiliki kemampuan kategorisasi. Salah satu contoh kategorisasi adalah diskriminasi spesies hewan yang berbeda. Penelitian ini bertujuan untuk mengetahui kemampuan diskriminasi spesies pada Macaca fascicularis. Monyet ini memiliki kemampuan untuk melihat foto sebagai representasi dari benda sebenarnya. Penelitian ini menggunakan stimulus berupa foto-foto wajah dari manusia dan hewan dengan metode matching-to-sample task. Saya melakukan tiga eksperimen utama. Pertama, saya ingin menguji apakah M. fascicularis dapat mengelompokkan manusia dan makaka ke dalam dua kelompok terpisah. Kedua, saya ingin mengetahui apakah monyet mampu membedakan antara individu-individu spesiesnya dengan individu-individu-individu-individu dari spesies lain. Terakhir, saya ingin menguji apakah monyet mampu membedakan antara manusia dan hewan lain non manusia. Hasil penelitian menunjukkan bahwa M. fascicularis mampu memisahkan dan mengelompokan objek-objek ke dalam kategori yang diujikan walaupun saya menyingkirkan informasi visual dari stimulus seperti warna dan bentuk. Kemampuan monyet untuk mengidentifikasi objek berdasarkan sedikitnya informasi fisik mengindikasikan adanya kemampuan dalam membentuk konsep yang lebih abstrak. Selain itu, pada eksperimen ke dua, monyet saya berhasil menemukan ciri-ciri unik dari masing-masing spesies makaka. Saya juga menemukan bahwa monyet berhasil mengelompokkan foto-foto hewan yang berbeda secara fisik ke dalam satu kelompok. Saya menduga bahwa monyet dapat membuat konsep yang lebih abstrak berdasarkan hubungan non-perseptual sebagai dasar untuk mengelompokkan objek ke dalam satu kategori. Saya menyimpulkan bahwa dalam mengkategori hewan, M. fascicularis menggunakan beberapa tingkatan abstraksi.
KANTHI ARUM WIDAYATI. Categorization in Macaca fascicularis. Supervised by BAMBANG SURYOBROTO, AKICHIKA MIKAMI, ACHMAD FARAJALLAH.
Categorization is an ability to group individuals or events into different classes mediated by conceptualized mental image. There are several levels of categorization and within a taxonomy the levels are nested. At the most concrete level of categorization, all or most members of the category shared common physical attributes that differ from other category. The higher the level of category, the fewer common attributes between members of the group.
In addition to humans, the ability to categorize has also been proposed in animals. In addition to humans, the ability to categorize has also been proposed for animals. Being able to identify, visually or otherwise, a new object as a member of a category is an advantage for animals. It should help them to distinguish between food or non-food, or to discriminate between species of animals. This species discrimination is important to prevent hybridization among species.
Using matching-to-sample task, present experiments tested the ability of the long-tailed macaque (Macaca fascicularis) in discriminating dichotomous-stimuli of different animals. The species has been shown to be able to see photos as representations of real objects so we used facial photos of humans and animals for the stimuli. Using operant conditioning method, I trained monkeys to associate matching to sample stimuli against a distractor stimulus. First, I showed monkeys a sample stimulus as a reference to be matched. To ensure that the subjects paid attention to the sample stimulus, they had to touch it and for this they received reward that they find beneath the stimulus. Next, I presented a matching stimulus and a distractor stimulus side-by-side. The subjects must choose one of them. When the subjects chose the matching stimulus, they received a piece of food as a reward; the response was counted as a correct one. When they chose the distractor, they did not get any rewards. Subject's motivation for reward warranted the choice of the matching stimulus. The location of matching and distractor stimuli on the tray was arranged pseudorandomly. I blocked every 20 trials into one session and measured their correct rate. If the subject chose the matching stimuli higher than 90% in a session, I interpreted they were able to associate matching to sample stimuli. Logically, this may be inferred as the subject had developed dichotomic concepts of matching against distractor. When they showed this competence, they went to test phase. I expect that monkeys were able to transfer their concept learned in training phase into new stimuli by showing the same performance in both baseline and test trials. In the test phases, I introduced new matching and distractor stimuli and see their response into the stimuli. I did three major experiment. First, I tested their ability to classify human and macaques into separate group. Second, I tested their ability to discriminate their conspecific from other macaques. And the last, I tested whether the subjects able to discriminate non-human animals from human.
generalize attributes of members of the group. This would indicate that the subjects have ability to create a higher level abstraction. On the other hand, in discriminating intragenic macaque species, I found that they were able to extract uniqueness of each species. More over, I also found that the subjects able to put photos of non-primates animals that shared very few similarities in physical properties into one group. Monkeys may able to create a logical concept such as A and non-A. I suggest that the subjects could create abstract concepts free from the physical properties as a basis to put objects into one category. Thus, I conclude that M. fascicularis are able to perform multiple levels of categorizations.
All Rights Reserved
It is prohibited to cite all or a part of this dissertation without referring to and mentioning the source. Citation is permitted for the purposes of education, research, scientific paper, report, or critism writing only; and it does not defame the name and honor of Bogor Agricultural University.
KANTHI ARUM WIDAYATI
Dissertation
submitted in partial fulfillment of the requirements for a Doctoral Degree in Animal Bioscience Major of Graduate School of Bogor Agricultural University
GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
1. Dr. Entang Iskandar (Researcher, Primate Research Center, Bogor Agricultural University)
2. Dr. Yamato Tsuji (Assistant Professor, Primate Research Institute, Kyoto University, Japan)
Examiners in the Open examination:
1. Prof. Ir. Wasmen Manalu, Ph.D. (Professor of Faculty of Veterinery Medicine, Bogor Agricultural University)
Title : Categorization in Macaca fascicularis Name : Kanthi Arum Widayati
Student ID : G362070051 Major : Animal Biosciences
Endorsed by,
Supervisory Committee
Dr. Bambang Suryobroto Chair
Prof. Dr. Akichika Mikami, MD Dr. Achmad Farajallah
Member Member
Acknowledged by,
Chair of Major of Animal Biosciences Dean of Graduate School,
Dr. Bambang Suryobroto Dr. Ir. Dahrul Syah, M.Sc.Agr.
The title of this dissertation is Categorization in Macaca fascicularis. The experiment took place in the Laboratory of Zoology Departement of Biology Faculty of Mathematics and Natural Sciences Bogor Agricultural University, Bogor and Primate Research Institute, Kyoto University, Japan.
I want to send my gratitude to Dr. Bambang Suryobroto, Dr. Achmad Farajallah and Prof. Dr. Akichika Mikami, MD as author’s advisors; to Mr. Adi Surahman, Mrs. Ani and Mr. Mamat for taking care of the monkeys. To all individuals with their unique personality in Zoo Corner (Dr. Tetri Widiyani, Sarah Nila, S.Si, Puji Rianti, M.Si, Islamul Hadi, M.Si, Eneng Nunuz R, S.Si, Elda Irma Kawulur, M.Si, Andi Darmawan, M.Si) for great discussion and their supports for the experiments.
I also want to send my appreciation to Dr. Yamato Tsuji from Primate Research Institute, Kyoto University, Dr. Harry Susianto from Faculty of Psychology, University of Indonesia, Dr. Entang Iskandar from Primate Research Center, Bogor Agricultural University and Prof. Wasmen Manalu, PhD from Faculty of Veterinary Medicine, Bogor Agricultural University for improving the dissertation.
I also want to send my gratitude to Dra. Taruni Sri Prawasti.M.Si, Dr. Dedy Duryadi Solihin, Tri Heru Widarto, M.Sc, Dr. Dyah Perwitasari, Dr. Rika Raffiudin, Dr. Tri Atmowidi, Msi for their support during the study.
I deeply send my gratitude to Yayasan SDM IPTEK HABIBIE CENTER for the scholarship for academic year 2008/2009 and 2009/2010. I also want to send my appreciation to program SANDWICH LIKE from Directorate General of Higher Education which gave me financial support when I work in Primate Research Institute, Kyoto University, Japan.
I also want to appreciate and gives love to Ucok, Sukhoi and Kerok for their great performances.
Bogor, February 1st 2012
Author was born in Bogor on 29th of September 1982 as the first child of
four from the parent, Hidayat Banjaransari and Sri Wilujeng.
Page
LIST OF FIGURES... xiv
APPENDIX …... xiv
INTRODUCTION …... 1
LITERATURE STUDY... 3
Categorization in Human ... 3
Categorization in Animal... 3
Operant Conditioning Behavior... 5
METHODS... 6
Subjects... 6
Apparatus... 6
Stimuli and Procedure... 7
a. Human vs Macaques... 10
b. Macaca fascicularis vs Other Macaques... 11
c. Human vs Non-Human... 12
Data Analysis... 13
RESULTS... 14
Human vs Macaques... 14
Macaca fascicularis vs Other Macaques... 18
Human vs Non-human... 20
DISCUSSION... 22
CONCLUSION... 27
Page 1 Schematic diagram of operant conditioning match-to-sample task of
the experiment in baseline (a) and test trials (b)... 8
2 Example of stimuli used for Human vs Macaques experiment... 9 3 Example of stimuli used for Human vs Non-Human experiment …... 13 4 Monkeys performances in categorizing human and monkeys in color
stimuli...
9 Monkeys performances in categorizing conspecific and other monkeys using colored stimuli...
19
10 Monkeys performances in categorizing conspecific and other monkeys using black and white stimuli...
19
11 Monkeys performances in categorizing human and non-human using colored and black and white stimuli...
21
APPENDIX
Page 1 Monkeys performance in Human vs Macaques experiments... 32 2 Monkeys performance in M. fascicularis vs other Macaques
experiments... 34
3 Monkeys performances in Human vs Non-Human experiments... 35 4 Test of the similarity of the monkey performances between baseline
INTRODUCTION
We live in a world full of ever-changing objects. It is impossible for us to
memorize each and every new individuals we encountered. Our brain need to have
an ability to deal with infinite informations that is continuously coming from the
surrounding environment. One way to overcome the memory storage constraint is
to represent factual objects as conceptualized mental images. A concept concludes
a lot of informations that had been collected at the time we perceive the object and
abstracts them to assist the process of grouping individuals or events into different
categories (Rosch et al. 1976). These categories reduce the number of bits of
information to manageable classes. There are several levels of categorization and
within a taxonomy the levels are nested (Rosch et al. 1976). At the most concrete
level of categorization, all or most members of the category shared common
physical attributes that differ from other category. The higher the level of
category, the fewer common attributes between members of the group.
In addition to humans, the ability to categorize has also been proposed in
the animals. Being able to identify, visually or otherwise, a new object as a
member of a category is an advantage for the animals. It should help them to
distinguish between food or non-food, and to discriminate between species of
animal. This species discrimination is important to prevent hybridization among
species (Yoshikubo 1985, Fujita 1987, Fujita and Watanabe 1995, Fujita et al.
1997).
Present experiment aims to test the ability of species discrimination by
primates. I did three major experiments. First, I tested whether the monkeys were
able to categorize monkey individuals as a class against human individuals as
another class. Second, I tested whether they were able to discriminate their
conspecific from other macaque species. Third, I tested their ability to categorize
non primate animal as non-human class against human individuals as human
class. Previous studies showed that monkeys were able to see photos as
representations of real objects (Kyes et al. 1982). I therefore used photos of
ability. The stimuli consisted of information on physical properties of the object.
The physical properties represented in photos may be modified in many ways to
alter the amount of information in the stimuli. These modified photos provide a
way to deduce levels of categorization of the monkey. Some experiments found
that the monkeys look to their conspecific longer than to non-conspecific so they
used duration of visual fixation to indicate this discriminative capability (Demaria
and Thierry 1988, Fujita 1993, Fujita and Watanabe 1995, Fujita et al. 1997).
However, although counting the fixation time is easy to be described
quantitatively, it is uncertain as to what the reaction time measures. For instance,
the longer the time could represent two facts; the monkeys likes the picture in the
stimuli, or it can be the opposite. In fact, in agonistic bouts, the amonkeys tend to
look longer to their opponent (de Waal et al. 1976). Thus, the reaction time may
better be interpreted as a measure for the attention of the monkeys and may not
carry information about species discrimination. Moreover, it is not clear whether
the monkeys are truly able to distinguish between species or their familiarity with
their own species due to experience in their life history made them see conspecific
longer. Instead of counting fixation time of each stimulus, present experiment test
their categorization ability in discriminating dichotomous-stimuli of two different
LITERATURE STUDY
Categorization in Humans
Categorization is the ability to put an object to a group based on some
internal representations of the group (Rosch et al. 1976). The ability to categorize
objects and events and extending this behavior to new instances is fundamental to
many human activities. I sort the objects and events around us into categories,
while still being able to recognize some or all of the individuals. In general,
categorization could be divided into two levels (Behl-Chadha 1996). If the
detected physical properties of the individuals within a category are mostly
similar, we call it as basic level of abstraction (Rosch et al. 1976). For example;
we could put Asians, Africans and Caucasians into one group; that is a human
group. If the connection between members of one category is not only based on
perceptual similarity but more on relations between concepts, we call it as abstract
level of categorization (Mervis and Rosch 1975). For example, we can put rice,
bread and fruit into the food or edible group and put a chair, a plane and a soap
into the non-food group. Although it is basically divided into two levels, the
relation between each class often overlap and sometimes it is very difficult to
differentiate between classes. These levels of categorization are basic to
understand language, number and social relationship with other humans. This
ability obviously needs memory, learning and reasoning.
In humans, categorizing behavior was predicted to start at infantile age.
Quinn (1993, 2002) found that the 3-4 month old infants attended to natural
objects as though they belong to groups of basic level. The experiments used the
preferential looking procedure and measure the looking time of the pictures. The
infants tended to look at the picture of a new categories for longer period of time.
Their ability was regarded as in the basic-level of categorization.
Categorization in Animal
The field of research on the ability of animals to categorize objects was
that the pigeon were able to sort photographs on the basis of whether the image
contained human being or not. It means the pigeon were able to make a class of
human and a class of non-human, which is the definition of categorization. Since
then, several studies have demonstrated categorization in other animals, most of
them used primates as models (ex: Tanaka 2001, Jitsumori and Matsuzawa 1991,
Santos 2000). Some of the researches aimed to know whether categorization
processes in the animal is based on similar processess compare to human (ex:
Farbe-Thorpe 2003, Freedman et al. 2002,2003, Hampson et al. 2004). In other
case, some studies wanted to find out the levels of categorization that could be
performed by the animals (ex: Tanaka 2000, Jitsumori and Matsuzawa 1991,
Santos 2000, Inoue et al. 2008, Vonk & McDonald 2002; Vonk & McDonald
2004). In both cases, the studies used photos of biologically significant objects,
such food vs non food, animals vs non-animals, and the last is own species vs
non-own species or species discrimination (Tanaka 2001, Jitsumori and
Matsuzawa 1991, Santos 2000, Inoue et al. 2008, Fujita et al 1995, 1997, Vonk &
McDonald 2002; Vonk & McDonald 2004).
Species discrimination is important to prevent hybridization among species
(Yoshikubo 1985, Fujita 1995, Fujita et al. 1997). Some studies were conducted to
find out which part of the body are really important to identify and categorize the
species. Those studies found that face provide information about species, sex, age
and emotion of an individual (Pascalis and Bachevalier 1988, Tomonaga 2007).
Fujita et al. (1995) demonstrated that Sulawesi macaques performed basic level of
categorization in identifying their conspecific. The stimuli used in that experiment
were photos of monkey presented sequentially. By counting the fixation time of
each stimulus, they found that the Sulawesi macaques see their conspecific longer
than non-conspecific. Thus, the photos carry specific information which could be
categorized by conspecific or non-conspecific (Fujita et al. 1995, Pascalis and
Bachevalier 1988, Tomonaga 2007). However, although the fixation time is easy
to be described quantitatively, it is difficult to draw conclusion from it. For
example, the longer reaction time could mean that monkey likes the picture in the
look longer to his/her opponents (de Waal et al. 1976).
Looking at previous work in primates, there are some data on
categorization in great apes, M. mulatta, M. fuscata and Sulawesi macaques but
not really in long-tailed macaques (M. fascicularis). A few studies showed that M.
fascicularis were able to see photos as a representations of the real objects (Dasser
1987, Kyes et al. 1982). More over, some studies also found that those monkeys
were able to identify individual within species (Dasser 1987), read facial
expression from the drawing and could discriminate between drawings of some
bodies of old world primate species (Dittrich 1994). However, there were no
direct experiment to infer the level of categorization in this species.
Operant Conditioning Behavior
In this experiment, I used the operant conditioning methods which refers to
process in which the frequency of occurrence of the particular type of behavior
was modified by the consequences of the behavior (Reynolds 1975). In operant
conditioning, the behavior (called operant) came from animal motivation to
response to a given stimulus. The stimulus used in operant conditioning is
discriminative. In this particular experiment, monkeys were trained to choose a
specific stimulus (the plus stimulus, S+) from another alternative to get a reward.
If they choose the other (the minus stimulus, S-), they would not get any rewards.
The behavior to choose a specific card is an operant. The reward is the
consequences of the behavior and it should increase animal motivation to choose
the plus stimulus. The specific task in this operant conditioning method is
matching to sample (Miller et al. 1996) with photographs of human, macaques
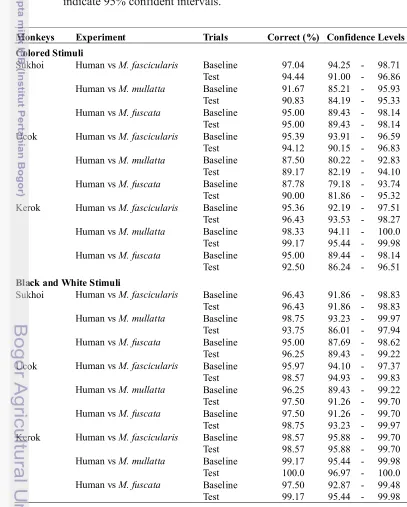
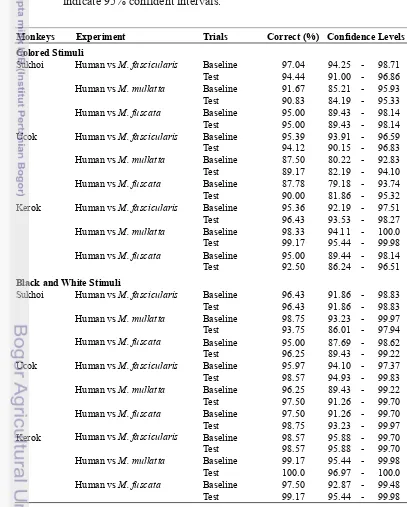
METHODS
Subjects
The subjects were two adult M. fascicularis males, named Ucok and
Sukhoi and one female named Kerok. Ucok were born in Pangandaran National
Park, with present age at least 19 years old. Kerok and Sukhoi were born in
Department of Biology, Bogor Agricultural University with ages are 14 and 9
years old, respectively. Both male monkeys were maintained at the Department of
Biology of Bogor Agricultural University. The female monkey was maintained at
Primate Research Institute, Kyoto University. They were reared in individual
cages and tested in the same cage. Experiments were conducted according to the
Guide for the Care and Use of Laboratory Animals by the National Institute of
Health, U.S.A. (1985), and the Guide for the Care and Use of Laboratory Primates
by the Primate Research Institute, Kyoto University (1986, 2002). The training
phases were held about two years for Ucok and one year for Sukhoi and two
months for Kerok depended on the ability of learning of the monkeys. The test
phase was conducted for few months for all monkeys.
Apparatus
A modified version of the Wisconsin General Test Apparatus (WGTA) was
used for the presentation of stimuli to the macaques. The apparatus used consisted
of a horizontal tray containing three shallow food-wells mounted on a portable
shelf. A sample stimulus and a matching stimulus were placed over the food-wells
so that they always associated with a food reward beneath. When setting up the
food reward between trials, an opaque screen was placed between the cage and the
experimenter to prevent animals from seeing the process. The eye of the
experimenter was covered with sunglasses, so the monkey could not use eye
direction of the experimenter as cue to find the matching stimulus. To control the
animal’s motivation level, animals were deprived of food after 5 pm until the next
morning before the day of a training session or a test session. Foods were
Stimuli and Procedure
I assessed the ability of M. fascicularis in categorizing objects by
conducting three experiments. First, Human vs Macaques experiment, aimed to
test the ability of M. fascicularis in categorizing monkeys and humans into two
different conceptual classes. Second, M. fascicularis vs Other Macaques
experiment, to test the ability of M. fascicularis in categorizing conspecific
differently from M. mulatta or M. fuscata. And the last one, Human vs Non
Human experiment, to test the ability of M. fascicularis in categorizing various
kinds of animal differently from human.
Fujita et al. (1995) demonstrated the ability of species discrimination of
Sulawesi macaques using photos as stimuli. Other studies of categorization also
used photos (Demaria and Thierry 1988, Fujita 1993, Fujita and Watanabe 1995,
Fujita et al. 1997) but those former studies did not control feature of the photos so
it is difficult to delineate which component of the photo provides informations to
do categorization. Each species of the animal possess distinctive non-facial,
physical features; for instance, posture and proportion of legs which may or may
not be present at the photo frames. For that reason, I used facial photos and
controlled the background color by changing it into green. Because of limitation
of visual fixation time to infer species discrimination as noted in Introduction,
present experiments tested their categorization ability based on dichotomic
discrimination of one species against other. To do this, I employed
match-to-sample task that often were used in working memory experiments (Miller et al.
1996, Rao et al. 1997) and later to study categorization experiments (Freedman et
al. 2002; Hampson et al. 2004; Inoue et al. 2008; Tanaka 2001). Using operant
conditioning method, at first, the monkeys were trained to respond only to
particular stimulus, not based on previous experience in their life. Every trial
consisted of two presentations. The first presentation shows sample stimulus, and
the second matching and distractor stimuli side by side. The sample and matching
stimuli always belonged to a same category that dichotomically differ to the
distractor (Figure 1). I assumed that each stimulus was independent from each
be a distractor in other trial depended to the sample stimulus.
I trained the monkeys to associate sample and matching stimuli against
distractor stimulus. First, I showed the monkeys a sample stimulus as reference to
be matched (Figure 1a). To ensure that the subjects paid attention to the sample
stimulus, they had to touch the sample stimulus and for this they received reward
that they may find beneath the stimulus. Next, I presented side-by-side matching
and distractor stimuli which one had to be chosen by the subjects. When the
subjects chose the matching stimulus, they received food as a reward; the
response was counted as a correct one. When they chose the distractor, they did
not get any rewards. Subject's motivation for reward warranted the choice of the
matching stimulus. I repeated the trials consecutively with inter-trial intervals for
about 30 second. The location of matching and distractor stimuli on the tray was
arranged pseudorandomly. I blocked every 20 trials into one session and measured
session, it means they were able to associate matching to sample stimuli. Then I
introduced a set of new photographs of the same human and the same monkey. I
repeated this training until the same criterion was reached (90% correct rate in 3
continuous sessions). After the monkey cleared the criterion of third set, I
introduced the photographs of a different human and a different monkey. This
procedure was repeated for six set of new humans and monkeys. After the monkey
cleared all of these training phases, I started the test session. At this stage,
logically, this may be inferred as the subjects had developed dichotomic concepts
of matching against distractor. When they showed this competence, they went to
test phase (see below for details) where the matching photos were new ones. If the
subjects were able to transfer the learned concept to novel stimuli, it proved that
a. Human vs Macaques
In this experiment, I assessed categorization ability of M. fascicularis to
discriminate monkeys from humans using facial features. Both primates possess
distinctive physical features, notably different shape of face and the presence of
hair in the monkey's face which is absent from human. These distinctions would
allow us to safely infer the discriminative ability on the different conceptual class
of human and monkey. For monkey category, I used photos of both sexes of three
species of macaques, those were M. fascicularis, M. mulatta and M. fuscata. For
human category, I used female and male photos. Some of the female heads were
covered by scarf. Nevertheless, all pictures showed the whole face.
For training phase, sample and matching stimuli were the same and
monkey's stimuli were always M. fascicularis (see Figure 2b and 2d). For
practical reason, I used 90% correct rate as threshold to infer subject's competence
in discriminating human from monkey categories. Furthermore, to make sure they
have achieved this ability, they have to show it in three successive sessions.
To test if a subject could apply the learned concept of human and monkeys
to new individuals, I changed the pictures of matching stimulus with different
individuals that belonged to the same category as the sample stimuli (eg. Figure
2c , 2e, 2g); for example, if the sample stimulus is monkey b then the matching
stimulus is monkey d. The subject should compare the novel matching stimulus to
the available distractor. If the subject associate the sample and the
different-picture of matching stimuli, I may infer that they put those two different-pictures into one
class that dichotomically differs from the class of distractor. This would evidenced
they transfer the human and monkey concepts to novel stimuli. This would prove
their categorization ability. For practical reason, the test sessions consisted of
baseline and test trials in certain proportion. In the baseline trials, the stimuli is the
same as in training trials so this would provide a reference to compare the test
trials.
The stimulus photos give subjects various informations about color and
shape of each species. Thus, I expected that subjects used informations from
of categorization. To test whether the subjects has levels of abstraction, I extended
the experiment to see whether the subjects would still have the ability to identify
objects if the informations on the physical properties of the stimuli were reduced.
For this reason, I deviced extended experiments that stripped off certain visual
information from the stimuli. First, I took color off the photos and presented it in
black and white. In the next step, I tried to test whether size of the stimuli could
affect the performance of the subjects in categorizing humans differently from
monkeys. I thought that altering the size of the photos would also perspectively
change the shape of the figures. I reduced the size of the stimuli to one quarter of
the original and showed it to monkeys. In this case, I tested them using pairs of
photos of colored human and M. fascicularis. I continued the test by giving the
subjects the blurred, black and white, original size photos of human and
macaques. These photo manipulations aimed to test whether the subjects could
categorize human differently from monkeys though the stimuli lack informations
about color and shape. In my last attempt to ascertain whether the subjects could
categorize human differently from macaques, I reduced the size of the black and
white, blurred photos of human and monkeys to one quarter of the original size.
The complete and modified physical properties of the photo stimulate the varied
amount of information perceived on the objects. Thus, by comparing subject's
responds to the stimuli, I may interpret their level of categorization. In total, I
used 82 photos as the stimuli for the test.
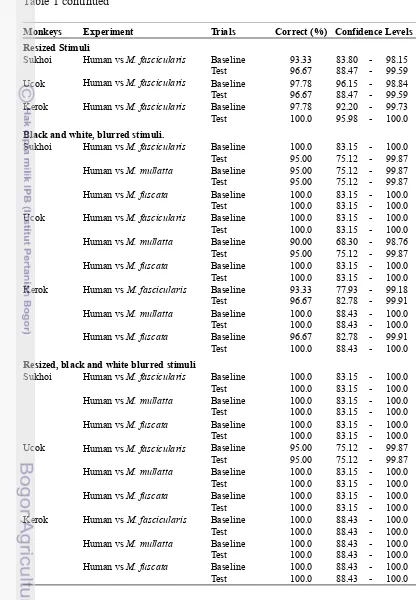
b. Macaca fascicularis vs Other Macaques
In this experiment, I assessed categorization ability of M. fascicularis to
discriminate their own species from other species of macaques. In this
experiment, I used several facial photos of M. fascicularis, M. mulatta and M.
fuscata. I did this in two conditions, that is in color and black and white.
After the first experiment of Human v Macaques, I assured that monkeys
have mastered to do maching-to-sample task. In this intra-generic experiment I
used two paradigms, that is, M. fascicularis vs M. mulatta and M. fascicularis vs
whether subjects were able to find the difference between each species and use it
to group them into separate class. In test phase, I changed the pictures of matching
stimulus with different individuals that belonged to the same species. Same as the
first experiment, the test session consisted of certain proportion of baseline and
test trials. In baseline trials of both paradigms, the photos of matching stimuli
were the same to sample stimuli. In total, we used 40 photos as the stimuli for the
test.
c. Human vs Non-Human
In this experiment, I assessed categorization ability of M. fascicularis to
discriminate various kinds of animals from human using facial features. In this
experiment, I used several photos of human (see the first experiment) and
non-primate animals, such as mammals, reptiles and amphibians (Figure 3). I
wondered whether subjects were able to ignore the physical difference of various
animals and creating a new class, that is non-human group. As in the second
experiment, I did this in color and black and white. In test trials, I changed the
pictures of matching stimulus with different individuals that belonged to the same
category as the sample stimuli; for example, if the sample stimulus was monkey
2b then the matching stimulus was animal 3a. The subjects should compare the
novel matching stimulus to the available distractor. If the subjects could associate
the sample and the different-picture of matching stimuli, I may infer that they put
those two pictures into one class that dichotomically differs from the class of
distractor. This would mean they are able to categorize non-human animals
differently from human. The baseline stimuli were the same stimuli used in
Human vs Macaques experiment (see Figure 2). In total, I used 68 photos as the
Data Analysis
In this experiment, monkeys have to respond to particular stimuli. I
defined the response of monkeys as correct when they choose matching stimulus,
and false when they choose distractor stimulus. This binomial responses was
dependent on stimuli presented in baseline and test trials; I therefore treat baseline
and test trials as independent variables and the effect of individuals in the analysis
as random effect. Thus, I used Generalized Linear Mixed Model (GLMM,
Venables and Ripley 2002) using R software version 2.10.1 (R Development Core
Team 2010) to analyze the discrete, binomial data.
In test phase, I also conducted several baseline trials using stimuli that
used in training phase and test trials using new stimuli. I expect that monkeys
were able to transfer their concept learned in training phase into new stimuli by
showing the same performance in both baseline and test trials. For that reason,
RESULTS
Human vs Macaques
As mentioned before, this experiment aimed to test the ability of monkeys
in categorizing monkeys and humans into two different conceptual classes. The
two classes were presented dichotomously as matching and distractor stimuli. In
baseline trials, the photos of matching stimuli were the same to sample stimuli. As
expected, in this trials all subjects could associate matching to sample stimuli.
They showed high performance of choosing the correct stimuli at the proportion
around 90% of the trials. Because the matching stimuli were always of a different
category to the distractor stimuli, it might mean they developed different and
mutually exclusive concepts of human and monkeys. These results provided a
reference to test whether subjects could transfer the newly developed mental
concepts to identify new individuals. I did this by changing the matching stimulus
with pictures of different humans and different kinds of monkeys. In this test, I
found that all subjects associate sample stimuli to new individuals of matching
stimuli (Figure 4, Appendix 1, Appendix 4). This same result to baseline
(glmmPQL, p=0.15; Venables and Ripley 2002) would indicate that the subjects
were able to correctly identify new individuals as members of its own class or of
human class. This would mean that monkeys categorized humans differently from
monkeys.
Since the stimuli used in the above experiment contained information on
physical properties (that is, shape and color) of the objects, I suggested that the
subjects used those properties to create a concept (see Discussion for detailed). I
was wondering whether the subjects would still have the ability to identify objects
if the informations of the physical properties were reduced. I expected that the
subjects will be able to gain informations from the stimuli with a reduced physical
properties and combine them with previous concepts they learned before and this
would lead them to create higher level of abstraction. For this reason, I deviced
First, I took color off from the photos and presented it in black and white. I found
that the subject's performance in both baseline and test phases were the same
(glmmPQL, p=0.19) (Figure 5, Appendix 1). This result indicates that even
without color, the subjects categorized humans differently from monkeys.
In the next step, I tested whether size of the stimuli could affect the
performance of the subjects in categorizing humans differently from monkeys. I
thought that altering the size of the photos would also perspectively change the
shape of the figures. I reduced the size of the stimuli to one quarter of the original
and showed it to monkeys. In this case, I tested them using pairs of photos of
colored human and M. fascicularis. I found that the subject's performance in both
baseline and test phase were the same (glmmPQL, p=0.85)(Figure 6, Appendix 1,
Appendix 4). It means the subjects did not affected by the reduction in size of the
I continued the test by giving the subjects the blurred and black and white original
size photos of human and macaques. These photo manipulations tested whether
the subjects could categorize human differently from monkeys though the stimuli
lack informations about color and shape. I found that the subjects were able to
categorize human separately from monkeys. Their performance in both baseline
and test phases were the same (glmmPQL, p=0.53) (Figure 7, Appendix 1,
Appendix 4).
In my last attempt to find out whether the subjects could categorize human
differently from macaques, I reduced the size of the black and white, blurred
photos of human and monkeys to one quarter of the original size. I found that
even in a condition lacking important physical informations, such as color, shape
and size, their performance in both baseline and test phases were the same
(glmmPQL, p=0.57) (Figure 8, Appendix 1, Appendix 4). It means that the
subjects could categorize human differently from monkeys. Overall, by seeing the
high performance of the subjects when tested by the manipulated stimuli, I
concluded that they were able to develop higher level of abstraction based on
available concepts they have learned before.
Macaca fascicularis vs other macaques
This experiment aimed to test the ability of monkeys in categorizing M.
fascicularis differently from other macaque species. In this experiment I used two
paradigms, that is, M. fascicularis vs M.mulatta and M. fascicularis vs M. fuscata.
In baseline trials of both paradigms, the photos of matching stimuli were the same
to sample stimuli. Thus, all subjects could associate matching to sample stimuli at
around 90% of trials. Similar with human vs macaques experiment, the matching
stimuli were always of a different category to the distractor stimuli, so this result
might mean they developed different and mutually exclusive concepts between
M. fascicularis and the other macaques; that is M. mulatta and M. fuscata. This
result provided a reference to test whether the subjects could use the developed
concepts of M. mulatta to identify new individuals of the species; the same also
different individual of monkeys. I found that all subjects associate sample
stimulus to new individuals of matching stimuli (Figure 9, Appendix 2, Appendix
4). This same result to baseline (glmmPQL, p=1) indicate that the subjects were
able to correctly identify new individuals as members of their respective class.
This would mean that the subjects categorized M. fascicularis differently from the
two other macaque species.
As in the first experiment, I ascertained this categorization ability by
taking color off the photos used in the stimuli and presented it in black and white
color. I found that all subjects associate sample stimulus to black and white
version of the matching stimuli (Figure 10, Appendix 2, Appendix 4). This same
result to baseline (glmmPQL, p=1) would indicate that even without color, the
subjects could also categorize M. fascicularis differently from other macaques.
Human vs Non-Human
This experiment aimed to test ability of monkeys in categorizing human
differently from non-primate animals. In this experiment, I used several photos of
human and non-primate animals, such as mammals, birds and reptiles and
amphibians. I tested the subjects using both color and black and white photos. For
baseline trials, I used same photos of human and M. fascicularis that used in
baseline trials of Human vs Macaques experiment. I found that the subjects did
high performance in test trials, similar to that in baseline trials (glmmPQL, p=0.65
for colored stimuli and p=0.70 for black and white stimuli) (Figure 11, Appendix
3, Appendix 4). It means that they were able to maintain concept of human and
DISCUSSION
Fujita and his coworkers (1993, 1995, 1997) found that monkeys see their
conspecific longer compared to other species, and this were thought to lead to
species discrimination. His works using various kinds of pictures of macaques
give us an insight into how monkey learn to categorize via species discrimination.
However, the methods that he used could be doubted; for example, there were
several interpretation for how monkeys see conspecific in longer duration. First,
the longer reaction time might mean that monkey's attention varied in response to
different pictures regardless of their species membership. Second, they were
mostly wild born so they have already familiar with their own species since
opportunities to learn facial properties of members of its own species exist in the
life history of the monkeys. Thus, monkeys may prefer to look longer at a picture
that is similar to their group mate and, by doing so, exclude pictures of another
species. Although it can be considered as species discrimination, it's still not clear
if monkeys can discriminate various species excluding their own. Third, since
most experiments (Demaria and Thierry 1988, Fujita 1993, Fujita and Watanabe
1995, Fujita et al. 1997) did not control features, such as backgrounds, of the
stimulus, it is hard to conclude which part of the stimulus attracts the monkeys
attention.
The method of matching to sample task I used might be more reliable than
counting perceptual duration for several reasons. The task of associating matching
and sample solves the first and second problems of Fujita (see experiments on M.
fascicularis vs non M. fascicularis below). More over, by controlling the
background of stimuli, and the use of a specific body part (that is, face), I tried to
reduce information noise which may distract my deduction. Thus, since my
stimuli represent real objects (Dasser 1987, Kyes et al. 1982), this experiment
showed that monkey could discriminate species. Therefore, my method should
make a strong conclusion about the ability of categorization in M. fascicularis.
The first experiment to show the ability of categorization in M.
introduced pictures of human and monkey to the subjects. There were consistent
similarities and differences of physical properties between human and monkey
pictures. It is not unreasonable to conclude that the subjects used these similarities
and differences to create a conceptualized mental image of human that differs to
that of monkey. There are some physical properties that provide information to
create concepts. First is shape. The global and local shapes of human's and
monkey's faces are very different. While human faces are oval, monkey's faces are
rounded with hair on it. Another possibility is that the subjects detected the
presence of the eye. This mechanism, called “eye direction detector”, is important
to understand facial emotion (Farroni et al. 2002, Baron-Cohen et al. 1999).
Given the eyes, its angles with nose and lips of both species are different. The
second is color. In this experiment, I used colored photographs. As seen in
stimulus photos (Figure 2), compared to uniformly presented background color,
the global color of the monkeys was different to that of human. Indeed, this had
been shown by Santos (2001) that colors are used as information to categorize
object. Those informations may help the subjects to recognize and discriminate
between categorically human and monkey.
The resulting concepts of human and monkey learned from training phase
were used by M. fascicularis individuals as basis to categorize pictures in the test
phase. In every trial of test phase I changed the baseline pictures with new ones.
This would prove the ability of the subjects to transfer its concepts to respond to
novel stimuli. For the monkey pictures, I used three different species of macaques.
I found that proceeding from baseline to test trials performance of the subjects
remained the same. These indicate that the subjects categorized monkey as
monkey and human as human. It is interesting to note that although two of the
subjects (Ucok and Sukhoi) had never saw other species (M. fuscata and M.
mulatta) for their entire life, they categorize photos of those species as monkey
instead of human which in captivity they see everyday. It might be concluded that
they had concepts of human and monkey based on different color and shape of the
stimuli. I may interpret this fact as the monkeys recognized natural discontinuity
As this kind of categorization ability was based on perceptual similarity of
physical attributes of the photos discussed above I may also describe this as the
concrete level of abstraction (Vonk and MacDonald 2002).
In nature, sometimes there are conditions (such as at long distance or less
light conditions) where monkeys could not see detailed information of object to
do categorization. Thus, they have to be able to categorize objects based on broad,
generalized concept that is developed on concept with complete and detailed
physical percepts. According to Rosch et al. (1976), the world is structured
because real-world properties do not occur independently of each other. Because
of this correlation, few properties of an object would suffice to predict the whole
properties. After seeing the results of the above test (which were positive), I
wondered whether the subjects would still have the ability to identify objects if
the informations on the physical properties of the stimuli were reduced. This
would simulate natural conditions and further proof the ability to transfer the
learned concept in my subjects. I then manipulated the stimuli by taking color off
the matching and distractor stimuli. The result showed that the ability of all of the
subjects to categorize between human and monkey was not hampered. They could
successfully transferred their mental image to the black and white photos. This
fact showed that the subjects did not depend on color and use other informations
to discriminate the stimuli. Some studies showed that monkey may use global
shape (Dittrich 1994) and local features (Demaria and Thierry 1988) of object as
informations in categorizing object. Jitsumori and Matsuzawa (1991) showed that
M. mulatta and M. cyclopsis wereable to classify human shilouettes as human and
assume that monkeys used global shape to recognize human. Rosch et al. (1976)
explained that levels of abstraction are nested. At the basic level, objects within
category shared most similarity in concrete properties. When it goes to the more
abstract levels, it share less properties among each other. Thus, we may follow
that the use of global shape is one way monkey used to generalize the physical
properties of an object. To test whether the subjects might be able to generalize
their physical percepts, I blurred or resized the stimuli to reduce and/or to change
tests, the M. fascicularis subjects showed high performance of discrimination.
This ability to classify blurred or resized photos of human as human and blurred
or resized photos of monkey as monkey demonstrated that the subjects were
looking for features of blurred or resized matching stimulus that have common
properties with those of sample stimulus. It seemed that the subjects subtracted
the local shape differences to get a generalized concept of object. Compared to
basic level of abstraction proofed before, this generalized level of abstraction
showed that the subjects performs at least two levels of categorization by learning
from their experience in training and test phases.
For second experiment, I tested the ability of M. fascicularis in
discriminating M. fuscata or M. mulatta against their conspecific. Those macaques
belong to fascicularis group (Fooden 1969) so they shared many similar physical
properties of face, such as the existence of facial hair and other local shapes. Thus,
the monkeys have to extract the differences from total facial properties to get the
uniqueness of each species. Result of the experiment showed that monkeys could
discriminate their conspecific from other species. They may used color as
information, since the hair color of M. fascicularis in the stimuli were lighter than
that of M. mulatta and darker from M. fuscata. More over, the facial color of M.
fascicularis is relatively brownish while M. mulatta paler and M. fuscata reddish.
In Human vs Macaques experiments, when color information was discarded, the
monkeys used (whether global or local) shape informations to categorize the two
different classes. Thus, I took the color off the stimuli and found that the subjects
again showed high performance of discriminations. This adds to my conclusion
that the subjects were able to detect the differences in physical properties of each
species and use it as basis in discrimination. This experiment showed the
categorization ability the other way around from the first experiment; that is, the
transfer of concept from general to specific.
The Human vs Macaques and M. fascicularis vs Other Macaques
experiments showed that M. fascicularis were able to develop concept using
informations from similarity and difference of physical properties to put objects
similarity to put objects into one group. One object may relate to others based on
relations that could not be detected easily by sensory properties. Thus, in this case,
the subjects need the ability to associate different conceptualized mental images to
discriminate stimuli lacking in physical clues. The class that is created does not
rely on perceivable features and may reflect a more conceptual understanding of
category belongingness. This kind of concept assisted higher level of
categorization. To test whether M. fascicularis were able to create such a more
abstract concept, I did Human vs Non-Human experiment. In two previous
experiments, there are physical similarities between all sample and matching
stimuli. In Human vs Non-Human experiment, I dismissed physical similarities
within one category; that is, Non-Human class consisted of photos of animals
such as mammal, bird, reptile and amphibian which were different in many ways.
At first I tested them with colored stimuli. Our subjects showed high performance
in discriminating between human and non-human classes. Since there are no
similarity within non-human category, I may suspect that the subjects use color
information to do categorization. Still, if I took color off the stimuli, the subjects
discriminated non-human from human. I assumed that the monkeys created a new
concept, that is concept of non-human that differ from concept of human. Thus,
the result clearly showed that the subjects could perform higher level of
CONCLUSION
These studies showed that M. fascicularis were able to discriminate
several species of animals. They did it by creating concepts that were based on
common features of the stimuli. Thus, I concluded that M. fascicularis were able
REFERENCES
Baron-Cohen S, Ring HA, Wheelwright S, Bullmore ET, Brammer MJ, Simmons
A, and Williams SCR. 1999. Social intelligence in the normal and autistic
brain: an fMRI study. European J. of Neurosci. 11: 1891-1898.
Behl-Chadha. 1996. Basic level and superordinate-like categorical representations
in early infancy. Cognition 60 :105-141.
Dasser V. 1987. Slides of Group Members as Representations of the Real Animals
(Macaca fascicularis). Ethology. 76: 65–73.
Demaria C, Thierry B. 1988. Responses to Animal Stimulus Photographs in
Stumptailed Macaques (Macaca arctoides). Primates 29: 237-244.
de Waal FBM, van Hooff, Netto W. 1976. An Ethological Analysis of Types of
Agonistic Interaction in a Captive Group of Java-monkeys (Macaca
fascicularis). Primates 17: 257-290.
Dittrich W. 1994. How monkeys see others : Discrimination and recognition of
monkeys' shape. Behav. Processes 33: 139-154.
Eimas PE and Quinn PC. 1994. Studies on the formation of perceptually based basic-level categories in young infants. Child Development65:903–917. Fabre-Thorpe M. 2010. Visual categorization: accessing abstraction in
non−human primates. Phil. Trans. R. Soc. Lond. B 358: 1215-1223.
Farroni T, Csibra G, Simion F, Johnson MH. 2002. Eye contact detection in
humans from birth. Proc. Natl. Acad. Sci. 99: 9602–9605.
Freedman DJ, Riesenhuber M, Poggio T, Miller EK. 2002. Visual categorization
and the primate prefrontal cortex: neurophysiology and behavior. J
Neurophysiol 88:929-941.
Freedman DJ, Riesenhuber M, Poggio T, Miller EK. 2003. A comparison of
primate prefrontal and inferior temporal cortices during visual
categorization. The Journal of Neuroscience 23: 5235-5246.
Fujita K. 1987. Species recognition by five macaque monkeys. Primates 28:
353-366.
Pigtail Monkeys. Primates 32: 133-140.
Fujita K and Watanabe K. 1995. Visual Preference for Closely Related Species by
Sulawesi Macaques. Am. J. Primatol. 37: 253-261.
Fujita K, Watanabe K, Widarti TH, Suryobroto B. 1997. Discrimination of
macaques by macaques: the case of Sulawesi species. Volume 38: 233-245.
Hampson RE, Pons TP, Stanford TR, Deadwyler SA. 2004. Categorization in the
monkey hippocampus: A possible mechanism for enconding information
into memory. Proc. Natl. Acad. Sci 101: 3184-3189.
Herrnstein RJ and Loveland DH. 1964. Complex visual concept in the pigeon.
Science 146 : 549-551.
Inoue T, Hasegaway T, Takara S, Lukatz B, Mizuno M, Aou S. 2008.
Categorization of biologicallsignificant objects, food and gender, in rhesus
monkeys I.. Behavioral study. Neuroscience Research 61: 70-78.
Jitsumori M and Matsuzawa T. 1991. Picture perception in monkeys and pigeons:
transfer of rightside-up versus upside-down discrimination of photographic
objects across conceptual categories. Primates 32: 473-482.
Pascalis O and Bachevalier J.1998. Face recognition in primates : a cross-species
study. Behav. Processes 43: 87-96.
Quinn P. 2002. Category representation in young infants. Current Directions of
Psychol.l Sci. 11:66-70.
Quinn PC, Eimas PD, Rosenkrantz SL. 1993. Evidence for representations of
perceptually similar natural categories by 3-month-old and 4-month-old
R Development Core Team. 2010. R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, ISBN
3-900051-07-0. http://www.R-project.org/. [August 10, 2010].
Rao SC, Rainer G, Miller EK. 1997. Integration of What and Where in the
Primate Prefrontal Cortex. Science 276 :821-824.
Reynold GS. 1975. A Primer of Operant Conditioning. Illinois: Scott, Foresman
and Company.
Rosch E, Mervis CB, Gray WD, Johnson DM, Boyes-Braem P. 1976. Basic object
in natural categories. Cogn Psychol 8: 382-439.
Santos LR, Hauser MD, Spelke ES. 2001.Recognition and categorization of
biologically significant objects by rhesus monkeys (Macaca mulatta): the
domain of food. Cognition 82 :127–155.
Tanaka M. 2001. Discrimination and categorization of photographs of natural
objects by chimpanzees (Pan troglodytes). Animal Cognition 4:201-211.
Tomonaga M. 2007. Visual search for orientation of faces by a chimpanzee (Pan
troglodytes): face-specific upright superiority and the role of facial
configural properties. Primates 48:1–12.
Venables WN and Ripley, B. D. 2002. Modern applied statistics with S. Fourth
edition. USA: Springer.
Vonk J and MacDonald. 2002. Natural concept formation in a juvenile gorilla
(Gorilla gorilla gorilla) at three levels of abstraction. J. Exp. Analysis of
Behav. 78:315-332.
Vonk J and MacDonald. 2004. Levels of abstraction in Orang Utan (Pongo abelii)
Categorization. J. Comparative Psychol. 118:3-13.
Yoshikubo S. 1985. Species discrimination and concept formation by rhesus
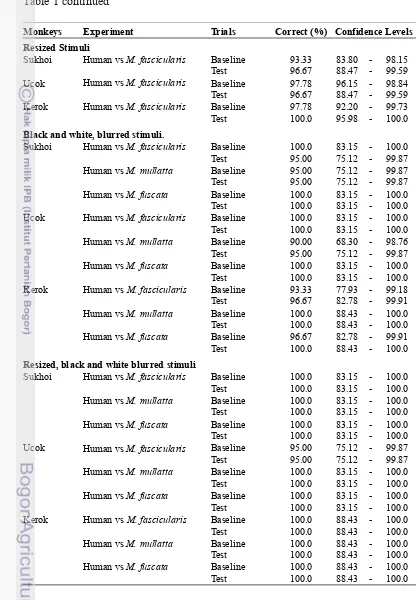
Appendix 1 Monkeys performance in Human vs Macaques experiments
Appendix 2 Monkeys performance in M. fascicularis vs other Macaques experiments
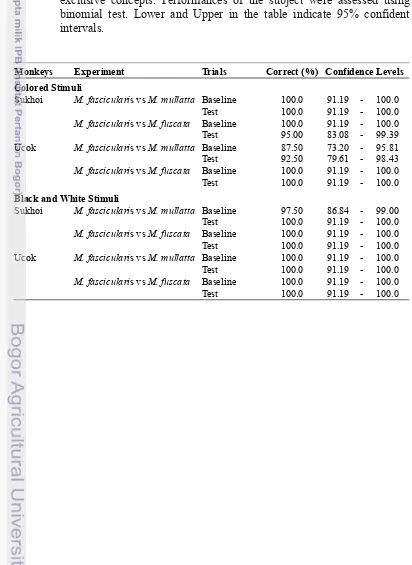
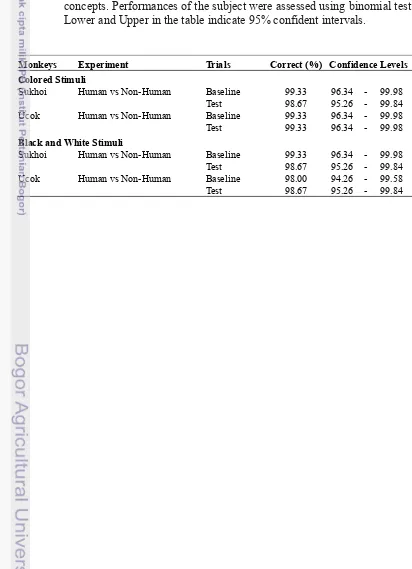
Appendix 3 Monkeys performances in Human vs Non-Human experiments
Tabel 3 Monkeys performances in categorizing human and non-human animal in Human vs Non-Human experiments. In every trials, correct response was defined as monkey chose matching stimulus in association with sample stimulus. Responses in test and baseline trials of all sessions were pooled to get the performance rates. Based on result of the training phase, we set 90% rate as threshold to define that the subject could discriminate human and non-human animal as mutually exclusive concepts. Performances of the subject were assessed using binomial test. Lower and Upper in the table indicate 95% confident intervals.
Monkeys Experiment Trials Correct (%) Confidence Levels Colored Stimuli
Sukhoi Baseline 99.33 96.34 - 99.98
Test 98.67 95.26 - 99.84
Ucok Baseline 99.33 96.34 - 99.98
Test 99.33 96.34 - 99.98
Black and White Stimuli
Sukhoi Baseline 99.33 96.34 - 99.98
Test 98.67 95.26 - 99.84
Ucok Baseline 98.00 94.26 - 99.58
Test 98.67 95.26 - 99.84
Human vs Non-Human
Human vs Non-Human
Human vs Non-Human
Appendix 4 Test of the similarity of the monkey performances between baseline and test trials
Tabel 4 Test of the similarity of the monkey performances (shown in Appendix 1 to Appendix 3) between baseline and test trials (GLMM, Venables and Ripley 2002). Values indicate statistical probability that performances in test trials were the same as in baseline trials. All values are higher than 0.05, it means the monkeys performances in both baseline and test trials are the same.
e. Blurred, Small, Black and White stimuli 0.57 1.00
2. Macaca fascicularis vs other macaques
1. M. fascicularis vs M. mulatta 2. M. fascicularis vs M. mulatta