PENGEMBANGAN PRODUKSI INOKULAN FUNGI MIKORIZA
ARBUSKULA BERBASIS BAHAN ALAMI DAN
PEMANFAATANNYA UNTUK PRODUKSI BIBIT JATI
(Tectona grandis L.f)
ABIMANYU DIPO NUSANTARA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi yang berjudul Pengembangan Produksi Inokulan Fungi Mikoriza Arbuskula Berbasis Bahan Alami dan Pemanfaatannya Untuk Produksi Bibit Jati (Tectona grandis L.f) adalah karya saya sendiri dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Juli 2011
ABIMANYU DIPO NUSANTARA. Developing arbuscular mycorrhiza fungi inoculants based on bio-material and its utilization for teak (Tectona grandis L.) seedling production. Supervised by CECEP KUSMANA, IRDIKA MANSUR, LATIFAH KOSIM DARUSMAN, and SOEDARMADI HARDJO-SOEWIGNYO.
Arbuscular mycorrhiza fungi (AMF) is a bioresources which is involved in various biogeochemical processes of carbon and soil nutrients. They influence plant fitness as well as terrestrial ecosystem stability. Mismanagement of land resources can make disappearance of AMF propagules from the soil. It is necessary to restore fungal propagules through inoculation of AMF in order to ensure the optimal function of terrestrial ecosystem. This dissertation composes of five experiments with main objective develop inoculant production model of AMF based on bio-material and its utilization for Tectona grandis L.f. seedlings production. Research results show that production of AMF inoculant is determined by AMF species and source of nutrient. Vermicompost and bone mill are bio-material that have potential to substitute fertilizer for the production of
AMF Glomus etunicatum inoculant. Good G. etunicatum inoculant can be
produced in the greenhouse by considering substrate sterilization, moisture content of the substrate, and bio-material used. Effectivity of each biomaterial is influenced by its nutrient content, particle diameter size and dosage. The finer the size of biomaterial the more effective source to produce G. etunicatum inoculant. The optimal dosage of bio-materials is equal to 50 mg P kg-1 of substrate. Solomon teak seedlings produced from tissue culture can be grown using polybag or bare root system with application of G. etunicatum inoculant and vermicompost. Inoculant of G. etunicatum in combination with vermicompost increase significantly teak seedling growth and chemical and biological characteristics of growth medium in the bare root system. Application mycorrhiza inoculant is recommended for producing teak seedlings from tissue culture to ensure teak seedling survivability in the field.
ABIMANYU DIPO NUSANTARA. Pengembangan Produksi Inokulan Fungi Mikoriza Arbuskula Berbasis Bahan Alami dan Pemanfaatannya Untuk Produksi Bibit Jati (Tectona grandis L.f). Di bawah bimbingan CECEP KUSMANA, IRDIKA MANSUR, LATIFAH KOSIM DARUSMAN, dan SOEDARMADI HARDJOSOEWIGNYO.
Fungi mikoriza arbuskula (FMA) telah hadir pada masa ketika permukaan bumi belum ditumbuhi tanaman darat. Fungi ini sangat penting artinya karena terlibat dalam berbagai daur biogeokimia unsur karbon dan hara sehingga menjamin kebugaran tanaman dan kemantapan ekosistem daratan. Fungi mikoriza arbuskula bertahan hidup dalam bentuk propagul yaitu spora, hifa ekstraradikal, tanah dan akar terkolonisasi. Bencana alam dan salah urus sumberdaya alam dapat meniadakan propagul dari permukaan tanah sehingga pengembalian propagul melalui pemanfaatan inokulan perlu dilakukan untuk menjamin ekosistem daratan tetap dapat berfungsi optimal untuk kesejahteraan seluruh mahluk hidup.
Disertasi ini disusun berdasarkan hasil lima percobaan, yang dirangkum menjadi tiga penelitian, dengan tujuan umum untuk mengembangkan model produksi inokulan fungi mikoriza arbuskula berbasis bahan alami untuk produksi bibit jati (Tectona grandis L.f.). Penelitian dilaksanakan di Laboratorium Silvikultur Fakultas Kehutanan IPB mengggunakan fungi mikoriza arbuskula
Glomus etunicatum, Acaulospora tuberculata, tanaman kudzu (Pueraria
phaseoloides Roxb) dan jati (Tectona grandis L.f) provenan Solomon, sumber
hara berupa larutan hara, SP36, tepung tulang, batuan fosfat dan vermikompos, serta zeolit dan tanah sebagai medium tumbuh. Data pengamatan diolah dengan analisis statistik model sidik ragam, korelasi dan regresi.
Terhadap Produksi Spora G. etunicatum. Penelitian ini terdiri atas dua percobaan yang bertujuan mendapatkan informasi mengenai interaksi sterilisasi, kadar air, dan sumber hara terhadap produksi inokulum G. etunicatum dan efektivitas simbiosisnya dengan tanaman kudzu. Hasil penelitian menunjukkan bahwa pembentukan struktur intraradikal G. etunicatum dan pertumbuhan kudzu lebih dipengaruhi oleh sumber P daripada oleh kadar air dan sterilisasi substrat. Tidak perlu dilakukan sterilisasi substrat untuk memproduksi inokulum G. etunicatum. Pemberian air dalam produksi inokulum cukup sampai sebanyak 50% dari kapasitas substrat memegang air. Vermikompos dan tepung tulang sapi dapat menggantikan peran pupuk buatan sebagai sumber hara. Kolonisasi dan sporulasi G. etunicatum dan efektivitas simbiosisnya dengan tanaman kudzu meningkat lebih tinggi jika menggunakan vermikompos dibandingkan dengan tepung telung sapi. Jumlah spora dan vesikel G. etunicatum tidak ditentukan oleh pertumbuhan awal tanaman kudzu. Takaran optimal vermikompos berukuran garis tengah butir
< β50 m ialah sebesar 150 – 224 mg per 175 g substrat.
Penelitian ketiga berjudul Efektivitas Metode Penyiapan Bibit, Inokulan Mikoriza dan Pupuk Terhadap Pertumbuhan dan Sifat Biologi Media Tumbuh Bibit Jati Solomon bertujuan menilai efektivitas interaksi metoda penyiapan bibit, inokulasi FMA G. etunicatum dan penambahan pupuk terhadap parameter pertumbuhan bibit jati dan parameter hayati media tumbuh bibit jati di kebun bibit. Hasil penelitian menunjukkan bahwa sistem akar telanjang menghasilkan indeks mutu bibit yang sama dengan polybag. Penggunaan mikoriza pada sistem akar telanjang bukan satu-satunya faktor yang menurunkan pertumbuhan bibit jati dibandingkan dengan pada system polybag. Sebaliknya, sistem akar telanjang menghasilkan media tumbuh dengan kadar hara dan aktivitas biologi yang lebih tinggi dibandingkan dengan sistem polybag. Perbanyakan bibit jati, melalui sistem akar telanjang dan polybag, tetap memerlukan pupuk dan vermikompos sebagai pupuk alternatif yang sama baiknya dengan pupuk buatan.
etunicatum dan vermikompos lebih efektif meningkatkan karakteristik kimia dan biologi medium tumbuh dalam sistem akar telanjang. Penggunaan inokulan mikoriza tetap dianjurkan untuk memproduksi bibit jati akar telanjang asal kultur jaringan untuk menjamin daya tahan hidupnya di lapangan.
@ Hak Cipta milik IPB, tahun 2011
Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian,
penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh
MIKORIZA ARBUSKULA BERBASIS BAHAN ALAMI DAN
PEMANFAATANNYA UNTUK PRODUKSI BIBIT JATI
(Tectona grandis L.f)
ABIMANYU DIPO NUSANTARA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Pengetahuan Kehutanan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi Pembimbing: a. Dr. Ir. Supriyanto, DEA
Staf Pengajar pada Departemen Silvikultur Tropika, Fakultas Kehutanan, Institut Pertanian Bogor b. Dr. Sri Wilarso Budi, M.Sc
Staf Pengajar pada Departemen Silvikultur Tropika, Fakultas Kehutanan Institut Pertanian Bogor
2. Ujian Terbuka Tanggal 27 Juli 2011 Penguji Luar Komisi Pembimbing:
a. Prof. Dr. Ir. Iskandar Zulkarnaen Siregar, M.Sc Staf Pengajar pada Departemen Silvikultur Tropika, Fakultas Kehutanan Institut Pertanian Bogor
b. Dr. Ir. Happy Widiastuti, M.S
Berbasis Bahan Alami dan Pemanfaatannya Untuk Produksi Bibit Jati (Tectona grandis L.f).
Nama : Abimanyu Dipo Nusantara
NRP : E061030132
Program Studi : Ilmu Pengetahuan Kehutanan
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Cecep Kusmana, M.S. Ketua
Dr. Ir. Irdika Mansur M.For.Sc. Prof. Dr. Ir. Latifah K. Darusman, M.Si.
Anggota Anggota
Prof. Dr. Ir. Soedarmadi Hardjosoewignyo, M.Sc Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Ilmu Pengetahuan Kehutanan
Prof. Dr. Ir. Fauzi Febrianto, M.S Dr. Ir. Dahrul Syah, M.Sc.Agr.
Ucapan syukur dipanjatkan kepada Tuhan YME atas segala nikmat-Nya sehingga disertasi yang berjudul “Pengembangan Produksi Inokulan Fungi Mikoriza Arbuskula Berbasis Bahan Alami dan Pemanfaatannya Untuk Produksi Bibit Jati (Tectona grandis L.f)” dapat terselesaikan. Pengembangan inokulan fungi mikoriza arbuskula merupakan kegiatan yang penting artinya untuk menjamin penyediaan dan pengembalian propagul fungi mikoriza arbuskula ke ekosistem daratan sehingga manfaatnya berguna untuk kesejahteraan umat manusia. Disertasi ini disusun sebagai salah satu syarat untuk memperoleh gelar Doktor pada Sekolah Pascasarjana Institut Pertanian Bogor.
Ungkapan rasa terima kasih yang sebesar-besarnya dan penghargaan setinggi-tingginya dengan tulus disampaikan kepada yang terhormat Prof. Dr. Ir. Cecep Kusmana, M.S., Dr. Irdika Mansur, M.For.Sc., Prof. Dr. Ir. Latifah K. Darusman, M.Si., dan Prof. Dr. Ir. Soedarmadi Hardjosoewignyo, M.Sc, selaku komisi pembimbing, atas segala bimbingan dan arahannya sehingga disertasi ini dapat terselesaikan dengan baik. Ucapan terima kasih juga disampaikan kepada
yang terhormat Dr. Ir. Rinekso, M.Sc., Dr. Deddy Duryadi, M.Sc., Dr. Ir. Supriyanto, DEA., Dr. Ir. Sri Budi Wilarso, M.Sc, Prof. Dr. Ir. Imam
Wahyudi, M.Sc., Prof. Dr. Ir. Iskandar Zulkarnaen Siregar, M.Sc, dan Dr. Ir. Happy Widiastuti, M.S sebagai penguji luar komisi pada ujian prelim, ujian
tertutup, dan ujian terbuka yang telah memberikan masukan mendasar terhadap keseluruhan isi disertasi ini.
Setiyadi, M.Sc., Dr. Ir. Hermawan Kresno Dipojono, M.S.E.E., Dr. Ir. Nanang Hariyanto, M.Sc., Dr. Kartini Kramadibrata, Drs. Alamsyah Harahap, M.Lib.Sc., Ir. Aso Kusuma, M.Eng., Sydharama Rudianto, Dr. Ir. Panca Dewi Manuhara Karti M.Si., Dr. Ir. Mochamad Chozin, M.Sc., Dr. Ir. Widodo Haryoko, M.S., Dr. Ir. Teguh Adi Prasetyo, M.S.c, atas segala perhatian dan bantuannya sehingga penulis dapat menyelesaikan tugas belajar di IPB.
Ucapan terimakasih penulis sampaikan pula kepada Kepala Laboratorium Silvikultur Fakultas Kehutanan IPB, Laboratorium Analisis Tanah dan Tanaman SEAMEO-BIOTROP, dan Laboratorium Biologi Tanah Fakultas Pertanian Universitas Bengkulu yang telah mengijinkan penggunaan seluruh fasilitas yang tersedia selama penulis melakukan penelitian. Kepada seluruh pengelola dan staf Laboratorium Bioteknologi Hutan dan Lingkungan, Pusat Penelitian Sumberdaya Hayati dan Bioteknologi, IPB, atas segala penerimaan dan perhatian selama penulis studi di IPB. Ucapan terima kasih juga disampaikan kepada seluruh pengurus Forum Wacana yang selalu berupaya untuk meringankan beban mahasiswa pascasarjana dan rekan-rekan lain yang tidak dapat disebutkan satu persatu atas segala perhatian dan bantuannya.
Ucapan terima kasih juga disampaikan kepada pimpinan Universitas Bengkulu dan pimpinan Fakultas Pertanian Universitas Bengkulu yang telah mengijinkan penulis untuk melanjutkan studi S3 di IPB, serta pimpinan Program Pascasarjana Ilmu Lingkungan Fakultas Pertanian Universitas Bengkulu atas segala perhatiannya selama penulis menyelesaikan studi S3 di IPB.
Ungkapan terima kasih dan kasih yang mendalam penulis sampaikan kepada istriku Dr. Ir. Rr. Yudhy Harini Bertham, M.P., anakku Ika Farida Wisnuwardhani S.P, dan Tri Ratna Anggraini Wisnumurthi S.H, menantuku Hilman Budiana, cucuku Hanifa Fatma Sabilla dan Damar Haikal Ibrahim atas segala kesabaran, pengorbanan, pengertian dan doanya.
Penulis dilahirkan pada tanggal 25 Desember 1956 di Purworejo, sebagai anak ketiga dari enam bersaudara dari Bapak Drs. Sru Adji Surjadi (alm) dan Ibu Hajjah Soemarni. Pendidikan SD sampai SMA diselesaikan di Jember, Jawa Timur. Gelar sarjana S1 bidang Kesuburan Tanah penulis peroleh pada tahun 1981 dari Jurusan Ilmu Tanah, Fakultas Pertanian Universitas Jember. Gelar sarjana S2 bidang Mikrobiologi Tanah penulis peroleh pada tahun 1994 dari Jurusan Ilmu Tanah Fakultas Pertanian Universitas Gadjah Mada, Yogyakarta. Pada tahun 2003 penulis diterima sebagai mahasiswa program S3 pada Program Studi Ilmu Pengetahuan Kehutanan, Sekolah Pascasarjana, Institut Pertanian Bogor.
Publikasi ilmiah yang merupakan bagian tidak terpisahkan dari disertasi ini telah di publikasikan pada jurnal ilmiah nasional dengan judul (i) “Peran substrat alami, kadar air, dan sterilisasi dalam produksi spora melalui simbiosis Pueraria
javanica dan Glomus etunicatum” pada tahun 2007 di Jurnal Akta Agrosia
yaitu (i) “Pengaruh sterilisasi bahan, kadar air, dan sumber fosfor terhadap pertumbuhan tanaman inang P. javanica dan produksi spora CMA G. etunicatum” pada tahun 2007 di Seminar Nasional Mikoriza II: Percepatan Sosialisasi Teknologi Mikoriza Untuk Mendukung Revitalisasi Kehutanan, Pertanian dan Perkebunan, (ii) “Assessing the effectiveness of seedling preparation method and mycorrhizal inoculants for enhancing teak seedling (Tectona grandis Linn. F) growth and soil biological activities” pada tahun 2010 di International Conference on Earth and Space Sciences, dan (iii) “Efektivitas vermikompos untuk memproduksi inokulan FMA Claroideoglomus etunicatum (Gerdemann & Becker, Schüßler & Walker) dan biomassa tanaman kudzu” dan “Teknik Produksi Inokulan FMA, Reformulasi, dan Monitoring” pada tahun 2011 di Workshop dan Seminar Nasional Mikoriza: Pupuk dan Pestisida Hayati Pendukung Pertanian Berkelanjutan Yang Ramah Lingkungan.
xvii
DAFTAR TABEL ………..……… xix
DAFTAR GAMBAR ……….………... xxiii
PENDAHULUAN ………..………... 1
Latar Belakang ………..………... 1
Tujuan Penelitian ………..………... 7
Hipotesis ………..………... 7
Manfaat ………..……….. 8
Kebaruan ... 8
Strategi Penelitian ………..……….. 9
TINJAUAN PUSTAKA ………..……….. 11
Definisi Mikoriza ………..………... 11
Mikoriza Arbuskula ………..………... 12
Produksi Inokulum FMA ………..………... 16
Interaksi Bibit Jati – Bahan Organik – Fungi Mikoriza Arbuskula …. 23 SELEKSI MIKORIZA ARBUSKULA DAN BAHAN BIO-ANORGANIK UNTUK MEMPRODUKSI INOKULAN MIKORIZA 33 Abstrak ………..………... 33
Abstract ………..……….. 33
Pendahuluan ………..………... 34
Bahan dan Metode ………... 35
Hasil dan Pembahasan ………..………... 40
Simpulan ………..……... 54
UKURAN DAN TAKARAN TEPUNG TULANG SAPI MENENTUKAN PRODUKSI INOKULAN MIKORIZA Glomus etunicatum NPI-126 (Becker & Gerdemann) ………... 55
Abstrak ………..………... 55
Abstract ………..……….. 55
Pendahuluan ………..………... 56
Bahan dan Metode ………... 57
Hasil dan Pembahasan ………..………... 60
xviii
Abstrak ………..………... 75
Abstract ………..……….. 75
Pendahuluan ………..………... 75
Bahan dan Metode ………... 78
Hasil dan Pembahasan ………..………... 82
Simpulan ………..……... 98
UKURAN DAN TAKARAN VERMIKOMPOS MENENTUKAN PRO-DUKSI INOKULUM MIKORIZA Glomus etunicatum DAN SIMBIOSIS-NYA DENGAN TANAMAN KUDZU ... 99
Abstrak ………..………... 99
Abstract ………..……….. 99
Pendahuluan ………..……….…….. 100
Bahan dan Metode ………..……. 101
Hasil dan Pembahasan ………..…………..……. 104
Simpulan ………..……...……. 112
EFEKTIVITAS METODE PENYIAPAN BIBIT, INOKULAN MIKORIZA DAN PUPUK TERHADAP PERTUMBUHAN DAN SIFAT BIOLOGI MEDIA TUMBUH BIBIT JATI SOLOMON ………... 113
Abstrak ………..……….…….. 113
Abstract ………..……….. 113
Pendahuluan ………..……….…….. 113
Bahan dan Metode ………..……. 116
Hasil dan Pembahasan ………..…………..……. 120
Simpulan ………..……...……. 144
PEMBAHASAN UMUM ………..………...……. 145
SIMPULAN DAN SARAN ………..………. 157
xix 1 Kadar hara tepung tulang, batuan fosfat, dan hiponeks merah ... 38 2 Nilai F hitung kolonisasi mikoriza pada akar kudzu umur 6 dan 12 MST
dan jumlah spora dalam media tumbuh kudzu umur 12 MST ………… 42 3 Interaksi sumber fosfor dan jenis fungi mikoriza arbuskula terhadap
kolonisasi mikoriza pada umur 6 dan 12 MST dan jumlah spora per 100 g media tumbuh kudzu umur 12 MST ...
42
4 Nilai F hitung bobot kering akar, pucuk, total dan nisbah tajuk akar kudzu umur 12 MST ...
44
5 Interaksi sumber fosfor dan jenis fungi mikoriza arbuskula terhadap bobot kering akar, pucuk dan total tanaman kudzu umur 12 MST (mg) ...
45 6 Rerata kolonisasi akar dan jumlah spora mikoriza arbuskula dalam
media tumbuh kudzu umur 12 MST ...
61 7 Takaran optimal tepung tulang untuk menghasilkan kolonisasi akar,
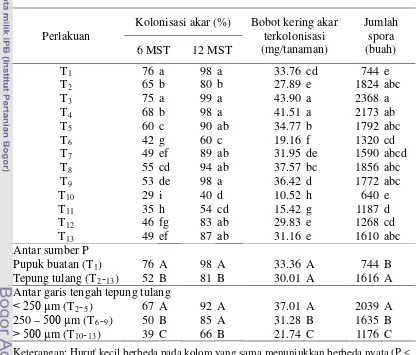
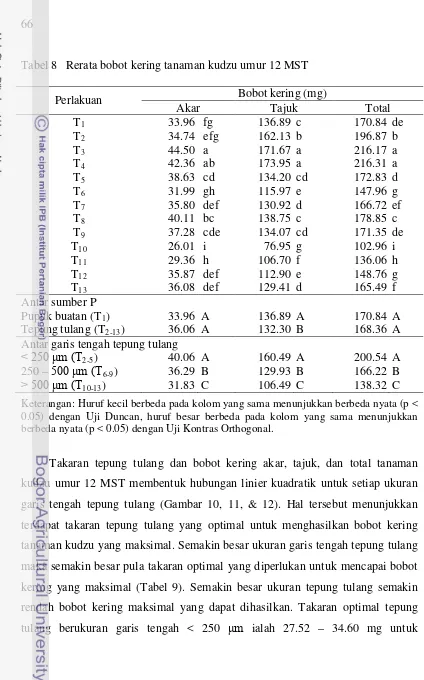
bobot kering akar terkolonisasi, jumlah spora G. etunicatum untuk setiap ukuran garis tengah tepung tulang ... 65 8 Rerata bobot kering tanaman kudzu umur 12 MST ... 66 9 Takaran optimal (mg) tepung tulang dan bobot kering maksimal (mg)
tanaman kudzu umur 12 MST yang dihasilkan oleh setiap pada ukuran garis tenga tepung tulang ... 68 10 Karakteristik fisikokimia sumber fosfor ………... 79 11 Rekapitulasi nilai F hitung pengaruh sterilisasi, kadar air, dan sumber P
terhadap jumlah vesikel, kolonisasi akar, dan jumlah spora pada umur 6 dan 12 MST ...
82 12 Interaksi substrat, kadar air, dan sumber fosfor terhadap kolonisasi (%) G.
etunicatum dalam akar kudzu pada umur 6 dan 12 MST ...
83
13 Pengaruh sumber fosfor terhadap jumlah spora (buah) per 100 g inokulan pada umur 6 dan 12 MST dan jumlah vesikel dalam akar tanaman kudzu pada umur 12 MST ...
84
14 Interaksi substrat, kadar air, dan sumber fosfor terhadap jumlah vesikel (buah) dalam akar tanaman kudzu pada umur 12 MST ...
85
15 Nilai F hitung pengaruh sterilisasi, kadar air, dan sumber P terhadap bobot kering total dan NPA tanaman kudzu umur 6 dan 12 MST ... 86 16 Interaksi sterilisasi substrat, kadar air, dan sumber P terhadap bobot kering
total tanaman kudzu (mg) pada umur 6 MST ...
86
17 Interaksi sterilisasi substrat, kadar air, dan sumber fosfor terhadap NPA tanaman kudzu pada umur 6 MST ...
xx
19 Nilai F hitung pengaruh sterilisasi, kadar air, dan sumber P terhadap serapan hara N, P dan Ca oleh tanaman P. phaseoloides umur 12 MST
89 20 Pengaruh sumber fosfor terhadap serapan hara N, P dan Ca pada
tanaman kudzu pada umur 12 MST ...
90
21 Rerata komponen pertumbuhan tanaman kudzu dan komponen inokulum G. etunicatum pada umur 12 MST ...
104 22 Rekapitulasi nilai F hitung tinggi bibit jati umur 2 – 26 MST ... 122 23 Rekapitulasi nilai F hitung garis tengah batang bibit jati umur 2 – 26
MST ... 123 24 Rekapitulasi nilai F hitung bobot kering bibit, panjang akar primer,
kolonisasi mikoriza arbuskula, dan aktivitas enzim fosfatase asam dan alkalin di akar bibit jati umur 4 MST ...
128
25 Interaksi metode penyiapan bibit, inokulasi mikoriza, dan pupuk terhadap bobot kering, panjang akar primer, dan aktivitas enzim
fosfatase alkalin akar bibit jati umur 4 MST ... 129
26 Rekapitulasi nilai F hitung pengaruh metode penyiapan bibit, inokulasi mikoriza, dan pemberian pupuk terhadap bobot kering bibit jati umur 26
MST ... 130
27 Interaksi metode penyiapan bibit, inokulasi mikoriza, dan pupuk terhadap bobot kering bibit bibit jati umur 26 MST ...
130 28 Rekapitulasi nilai F hitung pengaruh metode penyiapan bibit, inokulasi
mikoriza, dan pemberian pupuk terhadap mutu bibit jati umur 26 MST
131
29 Interaksi metode penyiapan bibit dan inokulasi mikoriza terhadap bobot kering akar lateral dan diameter batang bibit jati umur 26 MST ...
131 30 Rekapitulasi nilai F hitung pengaruh metode penyiapan bibit, inokulasi
mikoriza, dan pemberian pupuk terhadap serapan hara (mg) makro dan mikro bibit jati umur 26 MST ... 132 31 Interaksi metode penyiapan bibit dan inokulasi mikoriza terhadap
serap-an unsur hara makro N, P dserap-an K bibit jati umur 26 MST ...
133 32 Interaksi metode penyiapan bibit, inokulasi mikoriza, dan pupuk
ter-hadap serapan unsur hara mikro Fe, Mn, dan Zn bibit jati umur 26 MST ………... 133 33 Rerata kadar N dan P dan kolonisasi mikoriza pada akar bibit jati umur
26 MST ...
134 34 Interaksi metode penyiapan bibit, inokulasi mikoriza, dan pupuk
ter-hadap kolonisasi mikoriza pada akar bibit jati umur 26 MST ...
xxi media tumbuh bibit jati umur 26 MST ... 135 36 Interaksi metode penyiapan bibit, inokulasi mikoriza, dan pupuk
ter-hadap total fungi, respirasi, dan aktivitas fosfatase alkalin media tumbuh
bibit jati umur 26 MST ... 136
37 Rekapitulasi nilai F hitung pengaruh metode penyiapan bibit, inokulasi mikoriza, dan pemberian pupuk terhadap karakteristik kimia media
tumbuh bibit jati umur 26 MST ... 137
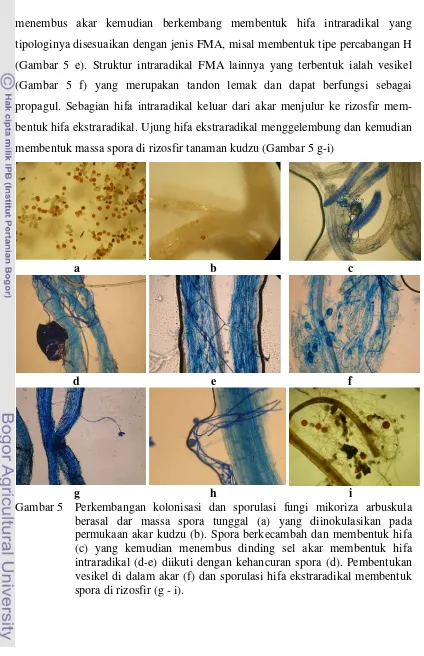
xxiii 1 Alur kegiatan penelitian ... 10 2 Tipologi spora G. etunicatum (kiri) dan A. tuberculata (kanan) ... 36 3 Tipologi hifa G. etunicatum (kiri) dan A. tuberculata (kanan) (400x) ... 37 4 Tipologi vesikel G. etunicatum (kiri) dan A. tuberculata (kanan) (400x) 37 5 Perkembangan kolonisasi dan sporulasi fungi mikoriza arbuskula
berasal dar massa spora tunggal (a) yang diinokulasikan pada permukaan akar kudzu (b). Spora berkecambah dan membentuk hifa (c) yang kemudian menembus dinding sel akar membentuk hifa intraradikal (d-e) diikuti dengan kehancuran spora (d). Pembentukan vesikel di dalam akar (f) dan sporulasi hifa ekstraradikal membentuk spora di rizosfir (g - i) ... 41 6 Hubungan antara takaran tepung tulang dengan kolonisasi G.
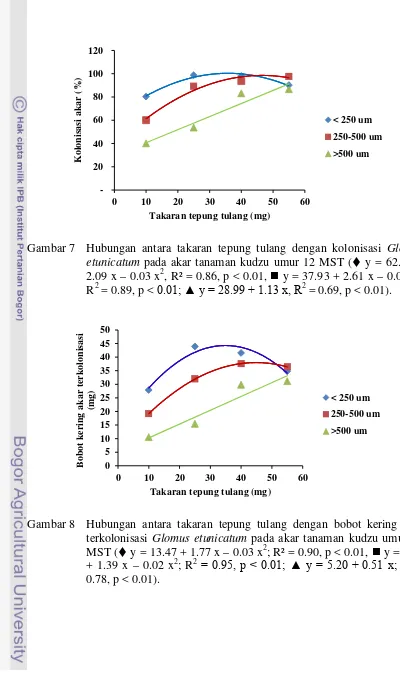
etunica-tum pada akar tanaman kudzu umur 6 MST ...
62 7 Hubungan antara takaran tepung tulang dengan kolonisasi G.
etunica-tum pada akar tanaman kudzu umur 12 MST ...
63 8 Hubungan antara takaran tepung tulang dengan bobot kering akar
ter-kolonisasi G. etunicatum tanaman kudzu pada umur 12 MST ………... 63 9 Hubungan antara takaran tepung tulang dengan jumlah spora G.
etuni-catum pada medium tumbuh tanaman kudzu umur 12 MST ...
64 10 Hubungan antara takaran tepung tulang dengan bobot kering akar
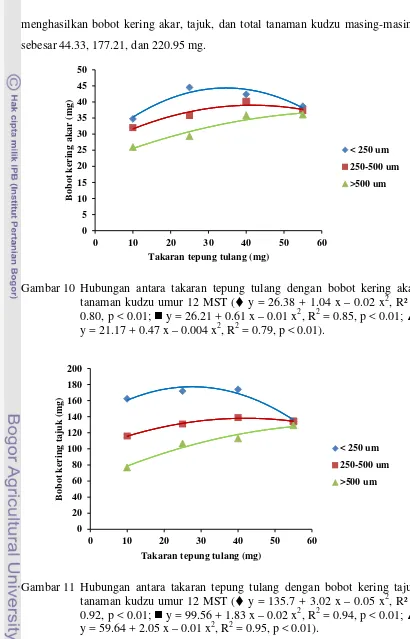
tanaman kudzu pada umur 12 MST ... 67 11 Hubungan antara takaran tepung tulang dengan bobot kering tajuk
tanaman kudzu pada umur 12 MST ... 67 12 Hubungan antara takaran tepung tulang dengan bobot kering total
tanaman kudzu pada umur 12 MST ...
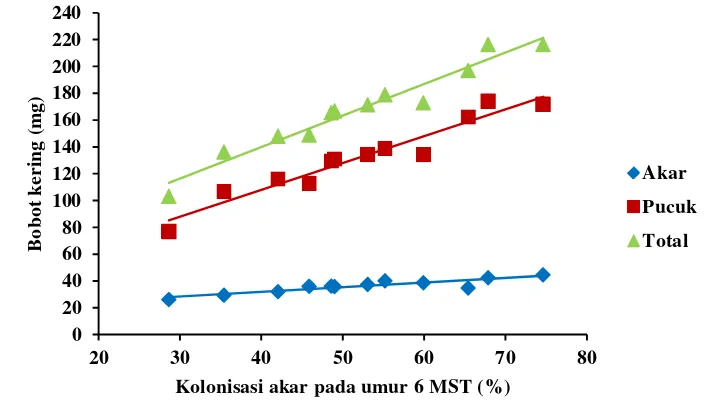
68 13 Hubungan antara kolonisasi FMA di akar kudzu umur 6 MST dengan
bobot kering tanaman kudzu umur 12 MST ...
69 14 Hubungan antara kolonisasi (%) pada akar tanaman kudzu umur 6 dan
12 MST dengan jumlah spora G. etunicatum ...
70 15 Hubungan antara takaran vermikompos dengan bobot kering akar
tanaman kudzu umur 12 MST ...
105 16 Hubungan antara takaran vermikompos dengan bobot kering tajuk
tanaman kudzu umur 12 MST ...
106 17 Hubungan antara takaran vermikompos dengan bobot kering total
tanaman kudzu umur 12 MST ...
xxiv
19 Hubungan antara takaran vermikompos dan bobot kering akar terkoloni-sasi FMA G. etunicatum umur 12 MST ... 108 20 Hubungan antara kolonisasi dengan jumlah spora G. etunicatum ... 108 21 Aklimatisasi bibit jati Solomon hasil perbanyakan melalui kultur jaringan 116 22 Penanaman bibit jati dalam polybag (kiri) dan akar telanjang (kanan) .... 117 23 Keragaan bibit jati Solomon umur 26 MST pada sistem akar telanjang
yang diinokulasi mikoriza (A& B), tidak diinokulasi mikoriza (D & E), dipupuk NPK (A & D) dan vermikompos (B & E) ... 121 24 Keragaan bibit jati Solomon umur 26 MST pada sistem polybag yang
diinokulasi mikoriza (A&B), tidak diinokulasi mikoriza (D & E), diberi pupuk NPK (A & D) dan vermikompos (B & E) ... 121 25 Pengaruh inokulasi mikoriza terhadap tinggi bibit jati umur 2 – 26
minggu setelah tanam ... 124 26 Pengaruh metode penyiapan bibit dan inokulasi mikoriza terhadap
tinggi bibit jati umur 2 – 26 MST (B = akar telanjang, P = polybag, + M = diinokulasi mikoriza, - M = tanpa mikoriza) ...
124 27 Pengaruh pupuk terhadap tinggi bibit jati umur 2 – 26 MST ... 125 28 Pengaruh inokulasi mikoriza dan pemberian pupuk terhadap tinggi bibit
jati umur 2 – 26 MST (M = mikoriza, TM = tanpa mikoriza) ...
126 29 Pengaruh metode penyiapan bibit dan inokulasi mikoriza terhadap
diameter batang bibit jati umur 2 – 26 MST ...
126 30 Pengaruh pemberian pupuk terhadap diameter bibit jati umur 2 – 26
MST ………. 127
31 Pengaruh inokulasi mikoriza dan pemberian pupuk terhadap diameter batang bibit jati umur 2 – 26 MST ...
Latar Belakang
Mikoriza arbuskula (MA) merupakan simbiosis tertua di permukaaan bumi, (Remy et al. 1994; Bonfante & Genre 2008) merupakan simbiosis yang paling sering ditemui di muka bumi, ditelaah, dan dimanfaatkan untuk peningkatan produktivitas sumber daya alam hayati dan nir-hayati (Smith & Read 2008; Gianinazzi et al. 2010). Fitobion yang terlibat dalam simbiosis MA ialah sekitar 80 – 90% tanaman darat baik yang memiliki akar atau tidak (Wang & Qui 2006) atau sekitar 73% bangsa tanaman berbunga (Brundrett 2009). Mikobion yang terlibat ialah fungi obligat warga filum Glomeromycota (Schü ler et al. 2001) yang memiliki empat bangsa (ordo) (Glomerales, Diversisporales, Paraglomerales, dan Archaeosporales), 11 suku (famili), 18 marga dan sekitar 300 jenis yang berhasil dikenali (Schüßler & Walker 2010).
Fungi MA hidup bersama dengan komunitas jasad hidup lainnya yang ada di rizosfer. Agar mampu hidup di rizosfer, sebuah habitat yang kompetitif dan dinamis, FMA harus mampu memproduksi banyak propagul dalam bentuk hifa ekstraradikal dan spora. Propagul FMA dapat punah karena kegagalan manusia menjaga keswalanjutan (sustainability) ekosistem (Barrios 2007; Douds & Johnson 2007, Gilbert 2009; Gianinazzi et al. 2010; Mendes-Filho et al. 2010). Menjadi tanggung jawab umat manusia untuk menjaga dan melestarikan propagul FMA di alam agar manfaat positifnya dapat diwariskan kepada generasi yang akan datang. Upaya yang dapat dilakukan diantaranya ialah stimulasi aktivitas FMA pribumi (indigenous) efektif mengggunakan praktek budidaya yang tepat misalnya budidaya organik, penggunaan bahan kimia pertanian sebijak mungkin, inokulasi FMA, penanaman bibit bermikoriza (Douds & Johnson 2007; Gosling et al. 2007), atau dengan penggunaan stimulan tertentu (Setiadi 2011 - komunikasi pribadi).
jika ada ternyata kemampuannya juga sangat terbatas. Hal tersebut menunjukkan produksi inokulan merupakan teknologi yang disembunyikan oleh produsen inokulan FMA dan para peneliti kurang berminat menekuni teknologi produksi inokulan. Oleh sebab itu, menjadi penting artinya untuk menguasai teknologi produksi inokulan FMA agar tidak tercipta ketergantungan terhadap produk dari luar negeri.
Inokulan FMA dapat diproduksi dengan metode konvensional atau metode modern yang memerlukan teknologi dan ketrampilan tinggi. Sekalipun telah ditemukan berbagai metode modern untuk memproduksi inokulan FMA (Douds 2002; Bhowmik & Singh, 2004; Selvaraj & Kim 2004; Lee & George 2005; Voets et al. 2005) namun metode konvensional yang lebih murah dan sederhana prosedurnya masih tetap menjadi andalan sebagian besar produsen inokulan FMA di dunia (Gianinazzi & Vosátka 2004; Feldmann et al. 2009; IJdo et al. 2011; Siddiqui & Kataoka 2011). Metode konvensional dapat dilaksanakan dengan teknik kultur pot terbuka menggunakan jenis FMA tunggal atau campuran, jenis tanaman inang, substrat, dan wadah tertentu yang diletakkan di rumah kaca atau lapangan. Teknik kultur terbuka merupakan teknologi produksi inokulan FMA yang lebih membumi dan dapat dikerjakan oleh masyarakat luas dibandingkan dengan teknik lainnya dan dapat dipadukan dengan kegiatan lain misalnya dalam produksi bibit.
menjamin ketersediaan pupuk dan pestisida kapanpun dibutuhkan oleh petani. Kemampuan memproduksi inokulan FMA dengan demikian menjamin kemandirian dan kedaulatan petani untuk melaksanakan budidaya tanaman tepat waktu tidak ditentukan oleh ketersediaan pupuk dan pestisida yang seringkali langka atau harganya melonjak tajam pada musim tanam. Inokulan FMA juga cukup diberikan sekali saja yaitu pada saat tanam namun menghasilkan pengaruh dalam jangka panjang. Hal ini dapat menekan investasi untuk pengadaan pupuk dan pestisida sehingga petani mampu menghasilkan produk pertanian yang aman, bermutu tinggi, namun dengan biaya yang lebih rendah. Produk pertanian yang bebas bahan kimia juga memperoleh harga premium. Kondisi demikian dapat menjamin produk pertanian Indonesia menjadi lebih kompetitif dibandingkan produk pertanian dari luar negeri.
dan organik. Bahan yang diinginkan ialah yang mudah diperoleh dengan harga murah dan efektif sebagai sumber hara dalam produksi massal inokulum FMA sekaligus memacu respon tanaman terhadap inokulan FMA.
Inokulasi FMA pada tanaman pertanian, hortikultura, dan kehutanan memerlukan inokulan bermutu tinggi dalam jumlah yang cukup. Mutu inokulan merupakan gambaran baik buruknya sebuah inokulan yang dapat didekati dari sisi fungi, tanaman, dan tanaman. Potensi propagul, atau jumlah spora dan hifa, infektif merupakan indikator mutu inokulan ditinjau dari sisi fungi (Feldmann & Idczak 1992; Feldmann et al. 2009; IJdo et al. 2011). Perubahan morfologi, fisiologi, dan biokimia yang ditunjukkan dalam bentuk kemampuan tanaman merespon cekaman hayati dan nir-hayati telah dilaporkan merupakan indikator respon tanaman pertanian terhadap inokulasi FMA (Khalil et al. 1994; Bressan & Vasconcellos 2002; Andrade et al. 2010; Smith et al. 2010). Indikator-indikator tanah seperti biomassa tanah dan kadar karbon total juga telah sering dikaitkan dengan aktivitas FMA pada tanaman pertanian (Rillig et al. 2001; Bago et al. 2003; Barrios 2007; Douds & Johnson 2007; Hamel 2007). Namun demikian, hasil penelitian juga menunjukkan keeratan hubungan antar indikator fungi-tanaman-tanah pada tanaman pertanian tidak selalu berlaku untuk pohon buah-buahan maupun tanaman hutan (Bâ et al. 2000). Sejauh ini belum pernah ditentukan kriteria dan indikator inokulan terbaik, khususnya untuk peningkatan pertumbuhan tanaman kehutanan ditinjau dari gatra (aspect) fungi, tanaman dan tanah. Oleh karena itu perlu dilakukan penelitian mengenai indikator mutu inokulan pada tanaman kehutanan ditinjau dari sisi fungi, tanaman, dan medium tumbuh.
persyaratan dan tujuan pengelolaan bahan tanaman (Ritchie 1984) yang ditunjukkan oleh pertumbuhan yang baik dan kokoh pada kondisi lapangan yang kurang mendukung dan memiliki laju pertumbuhan yang tinggi pasca penanaman (Duryea 1985; Johnson & Cline 1991; Mason 2004). Bibit jati mampu tumbuh kokoh dan sehat jika mendapatkan faktor tumbuh (air, hara, cahaya matahari) yang cukup dan perlindungan terhadap serangan hama dan penyakit.
Bibit jati umumnya diperbanyak dipersemaian secara generatif maupun vegetatif menggunakan polybag sebagai wadah dan medium tumbuh berupa tanah yang dicampur dengan substrat anorganik, misalnya pasir, atau organik misalnya sekam padi, kompos, atau serbuk gergaji. Polybag merupakan bahan yang sulit terdegradasi sehingga berpotensi mencemari lingkungan. Bibit dalam polybag diangkut dari persemaian bersama medium tumbuh dan wadahnya sehingga memerlukan volume ruang yang besar ketika diangkut ke lapangan. Pemindahan medium tanah juga berpotensi merusak lingkungan karena tanah harus terus diambil dari tempat lain. Keharusan membeli tanah dan wadah serta kebutuhan ruangan yang lebih besar selama pengangkutan dapat meningkatkan biaya produksi sehingga meningkatkan harga jual bibit. Informasi demikian menunjukkan perlu dilakukan upaya untuk mendapatkan teknik perbanyakan bibit jati yang murah biayanya namun tidak berpotensi merusak lingkungan.
merupakan alternatif pemecahan karena harganya cenderung naik sehingga berpengaruh terhadap harga jual bibit.
Penggunaan inokulan FMA dan pupuk organik yang dapat diproduksi sendiri merupakan alternatif yang lebih baik dibandingkan pupuk buatan untuk mengatasi hambatan pertumbuhan pada bibit jati akar telanjang. Rizosfer jati telah dilaporkan merupakan habitat beberapa marga FMA yang dominan yaitu Glomus, Gigaspora, Acaulospora dan Scutellospora (Maryadi 2001; Husna et al. 2006). Hasil penelitian menunjukkan penggunaan FMA dapat meningkatkan pertumbuhan bibit jati (Rohayati 1999; Irianto et al. 2003; Turjaman et al. 2003) atau tidak menghasilkan pengaruh yang nyata (Purnawan 2005) bergantung kepada provenan bibit jati, jenis FMA, lokasi pertumbuhan, dan teknik produksi bibit yang digunakan. Informasi demikian menunjukkan perlu dilakukan seleksi terhadap FMA yang akan diinokulasikan pada bibit jati di persemaian. Inokulasi FMA terseleksi bukan saja dapat menghemat biaya produksi bibit namun juga menjadikan bibit jati tumbuh lebih kokoh pada saat dipindah ke lapangan (Rajan et al. 2000). Reformulasi inokulan FMA, misalnya dengan menambahkan pupuk organik atau buatan, dilaporkan menghasilkan pengaruh yang berbeda-beda bergantung kepada jenis FMA, jenis dan takaran pupuk yang ditambahkan, dan provenan jati (Suwandi et
al. 2006; Ramadani 2008; Arif et al. 2009). Provenan jati yang digunakan pada
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Mendapatkan jenis FMA yang merespon positif bahan alami yang digunakan pada proses produksi inokulan FMA.
2. Mendapatkan prosedur pemanfaatan berbagai bahan alami dalam proses produksi inokulum FMA dengan menguji faktor sterilisasi, pengaturan pemberian air, dan sumber hara.
3. Mendapatkan bahan alami yang efektif untuk memproduksi inokulan FMA dan menghasilkan kerapatan spora yang tinggi menggunakan tanaman inang kudzu (Pueraria phaseoloides Roxb).
4. Menguji efektivitas sumber hara ditinjau dari sisi karakteristik kimia (kelarutan dan kadar hara), karakteristik fisik (ukuran garis tengah butir), dan takaran untuk memproduksi inokulan FMA.
5. Mendapatkan inokulan FMA yang efektif untuk meningkatkan pertumbuhan dan menyuburkan medium tumbuh bibit jati pada skala persemaian.
Hipotesis
Hipotesis yang akan diuji dalam penelitian ini ialah sebagai berikut :
1. Satu jenis FMA akan dapat memberikan respon positif terhadap bahan alami yang digunakan dalam produksi inokulan FMA.
2. Sterilisasi substrat, pengaturan kadar air substrat, dan pemilihan sumber hara perlu dilakukan untuk memaksimalkan produksi spora FMA.
3. Satu sumber hara yang efektif dapat diperoleh untuk memproduksi inokulan FMA dan menghasilkan kerapatan spora > 1000 buah per 100 g substrat. 4. Efektivitas bahan alami yang digunakan dalam produksi inokulum FMA
ditentukan oleh karakteristik kimia (kelarutan dan kadar hara), fisik (ukuran bahan), dan takaran bahan.
Manfaat Penelitian
Luaran dari penelitian ini diharapkan bermanfaat bagi peneliti dan pengguna FMA khususnya produsen bibit jati atau tanaman lain yang bernilai ekonomi tinggi. Rincian manfaat penelitian ini ialah sebagai berikut:
1. Memberikan pemahaman holistik mengenai faktor-faktor yang berpengaruh terhadap dan indikator keberhasilan prosedur produksi inokulan FMA.
2. Menghasilkan prosedur produksi inokulan FMA yang lebih sederhana berdasarkan sterilisasi substrat, pengaturan volume air penyiraman, dan karakteristik sumber hara.
3. Memberikan pemahaman produksi bibit jati berbasis sumber daya hayati dan nir-hayati yang tidak merusak lingkungan.
4. Membuka peluang usaha baru berupa produksi pupuk hayati berbasis bahan alami yang murah harganya, efektif meningkatkan pertumbuhan tanaman, dan ramah lingkungan sehingga melepaskan ketergantungan petani terhadap pupuk buatan dan produk dari luar negeri.
Kebaruan
1. Penggunaan bahan alami (bio-material) anorganik dan organik untuk mempercepat kolonisasi dan sporulasi FMA.
2. Penggunaan sumber hara organik dan inokulan FMA untuk mempercepat pertumbuhan bibit jati Solomon.
3. Penilaian kesuburan kimia dan biologi medium tumbuh bibit jati di persemaian.
4. Penggunaan metode pewarnaan akar menggunakan bahan yang aman bagi pengguna, murah harganya, dan prosedurnya lebih singkat.
Strategi Penelitian
Gambar 1. Alur kegiatan penelitian Penelitian III
Efektivitas metode penyiapan bibit, inokulan mikoriza dan pupuk terhadap pertumbuhan dan kesuburan medium tumbuh bibit jati
Solomon
inokulan FMA dan inokulan hasil reformulasi yang mampu (i) mempercepat respon bibit jati terhadap inokulan tersebut, (ii)
menghasilkan bibit jati yang bermutu tinggi pada skala persemaian, dan (iii) menyuburkan medium tumbuh bibit jati
Penelitian II
Interaksi sterilisasi, kadar air, dan sumber P dalam produksi inokulum Glomus etunicatum
Interaksi sterilisasi, kadar air, dan sumber P dalam produksi inokulum Glomus etunicatum Penentuan ukuran dan takaran vermikompos untuk
produksi inokulum G. etunicatum
Penelitian I
Seleksi mikoriza arbuskula dan bahan alami bio-anorganik untuk memproduksi inokulan mikoriza
Seleksi mikoriza arbuskula dan bahan alami bio-anorganik untuk memproduksi inokulan
mikoriza
Penentuan ukuran dan takaran tepung tulang untuk produksi inokulum G. etunicatum
Definisi Mikoriza
Sesuai dengan akar katanya yaitu myces dan rhiza, Frank (1885) untuk pertamakalinya mengajukan definisi yang bersifat kebendaan yaitu mikoriza sebagai fungi-akar. Frank pada saat itu meyakini mikoriza diperlukan untuk urusan keharaan bagi kedua mitra simbiosis. Istilah simbiosis (symbiotismus) dalam definisi mikoriza mengacu kepada konsep yang dikemukakan pertamakalinya oleh Frank pada tahun 1877 dan kemudian dikembangkan oleh de Bary pada tahun 1887 sebagai hubungan antara dua jasad hidup terlepas hasilnya apakah negatif (parasitime, sering disinonimkan dengan patogenisme) atau positif (mutualisme) (Smith & Read 2008). Oleh karena itu kemaujudan mikoriza kemudian ditelaah berdasarkan aspek fungsional dalam penyerapan berbagai substansi dari dalam tanah (Harley & Smith 1983) sehingga meningkatkan kebugaran salah satu atau kedua mitra yang bersimbiosis (Read 1999) karena terbentuknya antar-muka
spesifik untuk keperluan pertukaran bahan dan energi (Pfeffer et al. 2001).
Brundrett (2004) mendefinisikan asosiasi mikoriza sebagai simbiosis yang tidak selalu bersifat mutualis namun dapat setimbang ataupun eksploitatif, yang ditunjukkan dengan esensialitasnya untuk salah satu atau kedua mitra yang bersimbiosis, dapat terbentuk pada akar dan atau organ lain yang bersentuhan dengan substrat, fungsinya bekerja serentak dengan perkembangan tanaman-fungi. Hubungan kemitraan dalam asosiasi mikoriza merentang lebar dari yang negatif (parasit) (Bever 2002), netral (Aquino & Cassiolato 2002) atau positif (Hart & Reader 2002) bergantung pada berbagai faktor yang terlibat dalam asosiasi mikoriza (Johnson et al. 1997). Smith dan Read (2008) menegaskan adanya ciri khas dari simbiosis mikoriza, yaitu (i) terbentuk antara fungi, tanaman tingkat tinggi dan tingkat rendah, serta bakteri, (ii) memiliki struktur spesifik yang tidak dijumpai pada simbiosis jasad hidup-tanaman lainnya, misalnya arbuskula, vesikel, hifa, dan sebagainya, dan (iii) perkembangan, keberadaan, dan fungsinya ditentukan oleh kerjasama antar jasad yang bersimbiosis dan kondisi lingkungan.
tumbuh, dibentuk oleh fungi dengan tanaman dan jasad renik rizosfir, fungsinya ditentukan oleh interaksi jasad hidup yang bersimbiosis dan faktor lingkungan, sifatnya mulai dari mutualis sampai parasit, tugas utamanya ialah mempertukarkan energi dan bahan, dan berkembang serentak dengan perkembangan mitra simbiosisnya.
Mikoriza Arbuskula
Mikoriza merupakan asosiasi bersisi ganda yang terdiri atas beraneka kategori morfologi, fungsi dan evolusi (Brundrett 2002, 2004) dan dikelompokkan menjadi tiga yaitu (i) mikoriza arbuskula, (ii) ektomikoriza, dan (iii) mikoriza orchid, ericoid, dan mikoheterotrof (Brundrett 2004; Smith & Read 2008). Diantara ketiga mikoriza tersebut, asosiasi mikoriza arbuskula (MA) yang paling sering dijumpai keberadaan dan interaksinya di alam, ditelaah, dan dimanfaatkan untuk peningkatan produktivitas tanaman dan pembenahan lingkungan.
Asosiasi MA dibentuk oleh fungi obligat dengan sekitar 80 – 90% tanaman darat (Wang & Qui 2006) atau sekitar 73% bangsa tanaman berbunga (Brundrett 2009), baik yang memiliki akar maupun tidak (Read et al. 2000; Pressel et al. 2010). Asosiasi MA ditengarai telah ada semenjak 600 juta tahun lalu, atau mungkin lebih tua lagi sekitar 1 milyar tahun, yaitu dengan ditemukannya struktur mikroskopis serupa arbuskula dalam akar fosil tanaman purba Rhynia gwynne-vaughanii) dan gametofit (Aglaophyton major) tingkat rendah tidak berakar dan tidak berdaun (Taylor et al. 2009) dari jaman Ordovician (Pirozynski & Malloch 1975; Pirozynski & Dalpe 1989; Simon et al 1993; Remy et al. 1994) sampai Carboniferous (Strullu-Derrien & Strullu 2007; Strullu-Derrien et al. 2009). Hasil penelitian palaeobotani tersebut juga didukung oleh hasil analisis molekuler (Redecker et al. 2000; Heckman et al. 2001).
merupakan hasil modifikasi daun (Krings et al. 2011). Kolonisasi FMA tersebut tetap teramati pada tanaman Lycopsidae modern (Winther & Friedman 2008; Sudová et al. 2011). Berdasarkan keyakinan hubungan kemitraan FMA dengan tanaman modern (Parniske 2008; Smith & Read 2008) maka dapat diduga bahwa hubungan kemitraan demikian berlaku pada tanaman Lycopsidae pada jaman Carboniferous tersebut (Wagner & Taylor 1981). Hasil penelitian Taber & Trappe (1982) menunjukkan FMA ditemukan mengolonisasi daun tanaman jahe modern, namun tidak membentuk arbuskula dan belum terbukti adanya kemitraan yang saling menguntungkan. Temuan demikian memperkuat konsep yang dikemukakan sebelumnya (Brundrett 2004; Smith & Read 2008) bahwa bukan jenis organ tanaman yang menentukan asosiasi MA namun fungsi lingkungan yang dihasilkan oleh organ tanaman yang bersangkutan.
Mengacu pada perannya dalam pertukaran hara maka FMA diyakini merupakan fasilitator serapan hara tanaman purba ketika untuk pertamakalinya mengolonisasi daratan (Simon et al. 1993; Remy et al. 1994; Taylor et al. 1995). Pada jaman itu tanaman darat berasal dari tanaman air yang mendarat di muka bumi yang masih gersang. Tanaman itu tidak memiliki akar dan oleh karena itu memerlukan bantuan fungi mikoriza untuk mendapatkan hara dan air. Tanaman bermikoriza kemudian berevolusi membentuk struktur, organisasi internal, cara reproduksi, dan tumbuh pada habitat yang spesifik, sebagian diantaranya tetap bersimbiosis dengan MA dan sebagian lainnya tidak (Helgason & Fitter 2009). Sebaliknya, struktur FMA tidak mengalami perubahan struktur dan fungisnya. Jadi, berdasarkan sejarah evolusinya dapat disimpulkan tanaman memerlukan FMA agar mampu tumbuh baik dan produktif pada suatu lingkungan tumbuh tertentu.
kawasan dekat kutub (Frenot et al. 2005). Namun demikian, baru sekitar 300 jenis FMA yang berhasil diiidentifikasi karakternya, padahal menurut Morton et al. (1994) jumlah jenis FMA dapat mencapai 2700.
Beragamnya hasil akhir simbiosis MA, negatif sampai positif, disebabkan ada spesifitas fungsional antara FMA dan tanaman inang (Bressan 2002a,b; Hart & Reader 2002, 2005; Klironomos 2003; Leake et al. 2004; Feddermann et al. 2010) dan karakteristik lingkungan simbiosis (Gamage et al. 2004; Hamel 2007; Helgason & Fitter 2009). Interaksi demikian pada akhirnya berpengaruh pada struktur komunitas tanaman di alam (Klironomos et al. 2000; van der Heijden & Sanders 2002) dan karakter lingkungan simbiosis (Gianinazzi et al. 2010). Keragaman jenis dan pengaruh demikian perlu dijadikan bahan pertimbangan sebelum jasa baik simbiosis dimanfaatkan untuk peningkatan produktivitas tanaman dan lingkungan.
Jaringan HE di dalam tanah segera terbentuk setelah terjadinya kolonisasi akar. Hifa ekstraradikal berfungsi ganda yaitu untuk angkutan hara dan air, produksi spora, agregasi tanah, dan perlindungan tanaman inang dari serangan patogen. Peran HE dalam angkutan hara, khususnya P anorganik, penting artinya karena HE mampu menjangkau sampai di luar mintakat pengurasan (depletion zone) yang tidak dapat dijangkau oleh atau tidak tersedia untuk akar tanaman (Zhu et al. 2001). Hifa ekstraradikal, karena garis tengahnya yang jauh lebih kecil dibandingkan dengan garis tengah akar, mampu menembus pori mikro untuk mendapatkan air yang tidak dapat dijangkau oleh akar sehingga tanaman bermikoriza menjadi lebih adaptif menghadapi cekaman kekeringan. Sumbangan MA terhadap serapan P anorganik dan air dipengaruhi oleh jenis FMA, tanaman, dan lingkungan yang mengindikasikan kesesuaian fungsional di antara FMA dan tanaman tidak selalu berkaitan dengan kolonisasinya (Pearson & Jakobsen 1993; Burleigh et al. 2002).
Berdasarkan analisis genetika molekuler diketahui hanya ada satu fungi obligat yang membentuk asosiasi MA, yaitu Glomeromikota, yang berbeda asal usul dan karakternya dengan fungi lainnya (Askomikota dan Basidiomikota) (Schüßler et al. 2001). Glomeromikota memiliki empat bangsa (ordo) (Glomerales, Diversisporales, Paraglomerales, dan Archaeosporales), 11 suku (famili) dan 18 marga (genus) (Schüßler & Walker 2010). Pengetahuan tentang tatanama Glomeromycota terus mengalami penyempurnaan sejak tahun 2001, sebagai contoh bangsa Glomerales tadinya hanya beranggotakan suku Glomeraceae kemudian berubah menjadi dua suku yaitu Glomeraceae dan Claroideoglome-raceae (Schüßler & Walker 2010). Berdasarkan tatanama tersebut G. etunicatum berubah menjadi Claroideoglomus etunicatum. Informasi selengkapnya mengenai tatanama Glomeromycota tersedia pada www.amf-phylogeny.com.
makro (Govindarajulu et al. 2005; Li et al. 2006) dan mikro (Purakayastha et al. 2001; Nogueira & Cardoso 2002; Andrade et al. 2010), dan yang bersifat nir-hara misalnya produksi metabolit sekunder (Yao et al. 2003; Fester & Hause 2007). Fungi MA telah dilaporkan terlibat dalam proses rizodeposisi yang memperkaya tanah dengan substrat karbon sehingga meningkatkan aktivitas hayati tanah (Rillig
et al. 2001) dan agregasi tanah (Rillig et al. 2002; Rillig & Mummey 2006).
Sekalipun belum dilaporkan secara spesifik, seperti halnya FMA lainnya, patut diduga G. etunicatum terlibat dalam proses demikian.
Pengaruh positif simbiosis G. etunicatum menjadikan tanaman lebih tahan terhadap cekaman hayati, misalnya serangan patogen (Yao et al. 2003; Hao et al. 2005; Watanarojanaporn et al. 2011) dan hama (Wooley & Paine 2007); dan cekaman nirhayati, misalnya lebih tahan terhadap toksisitas logam berat (Wu et al. 2010) dan senyawa hidrokarbon (Gao et al. 2010), kekeringan (Wu et al. 2007), kegaraman (Ghorbanli et al. 2004), dan kepadatan tanah (Miransari et al. 2007). Simbiosis G. etunicatum juga berdampak terhadap morfologi akar tanaman (Bressan & Vasconcellos 2002) sehingga mempercepat pertumbuhan awal bibit tanaman berkayu yang ditumbuhkan pada tanah pasca penambangan (Flores-Aylas et al. 2003; Santos et al. 2008) atau memfasilitasi tanaman legum bersimbiosis dengan rizobia (Shrihari et al. 2000; Siviero et al. 2008).
Produksi Inokulum FMA
stimulasi aktivitas FMA pribumi (indigenous) efektif mengggunakan praktek budidaya yang tepat, inokulasi FMA, atau penanaman langsung bibit bermikoriza (Douds & Johnson 2007). Inokulasi mikoriza pada tanaman yang ditumbuhkan pada tanah terdegradasi terbukti berhasil memulihkan kesuburan tanah dan komunitas jasad hidup pada suatu ekosistem (Herrera et al. 1993; Jeffries et al. 2003; Gianinazzi et al. 2010). Untuk itu, penting artinya menguasai pengetahuan tentang produksi propagul atau inokulum dan tehnik inokulasi FMA serta faktor-faktor yang berpengaruh terhadap produksi propagul.
Inokulan FMA telah dipasarkan dengan berbagai nama, bentuk, dan formulasi untuk digunakan dalam upaya peningkatan produktivitas tanaman dan perbaikan ekosistem. Inokulan FMA dapat diproduksi menggunakan berbagai teknik yang masing-masing memiliki keunggulan dan kelemahan dalam hal rancangan, komersialisasi, dan ranah aplikasinya. Pada dasarnya terdapat tiga tehnik produksi inokulum yang umum digunakan yaitu teknik berbasis substrat, nir-substrat, dan in vitro (IJdo et al. 2011; Siddiqui & Kataoka 2011). Teknik berbasis substrat merupakan teknik produksi klasik yang telah digunakan di banyak negara dan merupakan tehnik yang murah namun efektif untuk mem-produksi inokulum secara massal.
Teknik nir-substrat (hidroponik dan aeroponik) (Jarstfer & Sylvia 1999; Singh & Tilak 2001) dan teknik in-vitro (Karandashov et al. 1999; Pawloswska et
al. 1999; Fortin et al. 2002) dikembangkan dengan tujuan untuk memproduksi
kentang hasil propagasi mikro (Voets et al. 2005). Produksi inokulum dapat dipacu dengan penambahan bakteri rizosfer perangsang pertumbuhan (Azospirillum sp.,
Azotobacter chroococcum, Pseudomonas fluorescens, P. Striata) atau khamir
(Saccharomyces cerevisiae) (Bhowmik & Singh 2004). Teknik in vitro belum umum digunakan di Indonesia mengingat komponen teknologinya memerlukan biaya yang tinggi dan operator yang terampil. Oleh sebab itu perlu dicari alternatif teknologi yang berbiaya murah namun mampu memproduksi spora dan inokulum FMA dalam jumlah yang banyak.
Teknik berbasis substrat, khususnya substrat tanah, merupakan teknik klasik yang telah lama digunakan untuk memproduksi inokulum berbagai jenis FMA mengingat biayanya yang murah dan kemudahan pelaksanaannya. Substrat yang digunakan pada umumnya berupa (1) tanah (Sylvia & Schenck 1983; Douds & Schenck 1990a,b; Sieverding 1991) atau lempung (clay) (Plenchette et al. 1982; Feldmann & Idczak 1992), (2) bahan mineral misalnya pasir (Millner & Kitt 1992; Lee & George 2005; Neumann & George 2005), inolite dan zeolit (Setiadi 2002) dan vermiculite (Douds et al. 2006), perlite (Lee & George 2005), (3) bahan organik misalnya kompos (Douds et al. 2005, 2006) atau gambut (Ma et al. 2007), atau campuran tanah dengan salah satu atau kedua bahan tersebut. Tehnik berbasis tanah kemudian diadaptasi untuk meningkatkan jumlah propagul di lapangan atau lahan pertanian (on-farm) (Douds et al. 2005, 2010).
1983, Millner & Kitt 1992). Spora hasil kultur penangkaran kemudian dikelompokkan berdasarkan karakter morfologi dan masing-masing diperbanyak dengan kultur tunggal. Inokulasi FMA dapat dilakukan dengan menempelkan spora tunggal ke permukaan akar muda atau meletakkan propagul di permukaan medium tumbuh pada bagian perakaran bibit.
Tanaman yang dapat digunakan untuk memproduksi inokulum FMA ialah
Allium cepa, Chloris guyana, Cenchrus ciliaris, Panicum maximum, Paspalum
notatum, Pueraria phaseoloides, Sorghum halepense, Trifolium subterraneum,
atau Zea mays (Chellappan et al. 2001; Setiadi 2002). Tanaman tersebut sering digunakan karena memiliki beberapa kelebihan, misalnya berumur pendek, memiliki system perakaran yang luas, dapat dikolonisasi sampai batas yang tinggi oleh berbagai jenis FMA, dan toleran terhadap kadar fosfor (P) rendah (IJdo et al. 2011). Kelebihan lainnya ialah resisten terhadap patogen, akar yang terkolonisasi umumnya berubah menjadi kuning dari putih (khususnya pada bawang perai dan jagung), dan toleran terhadap lonjakan suhu (Millner & Kitt 1992). Perbanyakan dilakukan rumah kaca, ruang berpendingin, pot yang diletakkan di lapangan atau yang lebih besar lagi pada bedengan persemaian (Gaur & Adholeya 2002, Douds et al. 2005, 2006). Optimasi kondisi pertumbuhan diperlukan mengingat pembentukan dan perkembangan FMA serta simbosisnya dengan tanaman inang ditentukan oleh tiga faktor utama yaitu jenis FMA, jenis tanaman, dan lingkungan simbiosis (Smith & Read 2008).
keberadaan senyawa kimia tertentu, misalnya pupuk (Onguene & Habte 1995), pestisida (Sukarno et al. 2000), dan hasil metabolisme tanaman inang yang dieksudasikan ke rizosfer misalnya khitin (Gryndler et al. 2003) atau strigolakton, 5-deoxy-strigol (Akiyama et al. 2005). Keberadaan bahan-bahan amandemen anorganik (zeolit, liat, perlit dan sebagainya) dan organik juga ikut menentukan laju produksi spora dan kolonisasi FMA ke akar tanaman (Saito 1990; Nishio & Okano 1991; Ezawa et al. 2002).
Unsur hara, khususnya P, berpengaruh langsung dan berpengaruh tidak langsung terhadap FMA melalui respon tanaman terhadap ketersediaan hara, misalnya dengan mengubah pertumbuhan akar atau fotosintesis. Kadar, bentuk, dan kelarutan sumber hara P merupakan faktor penting bagi pembentukan dan perkembangan MA yang paling sering dibahas. Kadar hara P dilaporkan berkorelasi negatif (Amijee et al. 1989; Koide & Li 1990), tidak berkorelasi (Tawaraya et al. 1996), atau berkorelasi positif (Carrenho et al. 2001) terhadap mikorizasi bergantung pada jenis FMA, jenis tanaman, bentuk dan kelarutan sumber P-nya (Cardoso 1996; Tawaraya et al. 1996; Nikolaou et al. 2002; Bhadalung et al. 2005). Pembentukan simbiosis mikoriza mencapai maksimum jika kadar P dalam tanah tidak > 50 mg kg-1 (50 ppm) (Ishii 2004) dan kadar hara
P sebesar β0 M optimal untuk produksi inokulum Glomus etunicatum pada sistem berbasis pasir menggunakan tanaman inang jagung (Millner & Kitt 1992). Perkembangan hifa marga Glomus berkorelasi negatif dengan kadar P namun sporulasi hifa tidak berkorelasi dengan kadar P (Tawaraya et al. 1996). Produksi propagul Glomus intraradices pada rizosfer jagung meningkat tajam pada kondisi tanpa pasokan P dan sebaliknya jika dipasok unsur P (Gaur & Adholeya 2000). Sporulasi G. etunicatum, dilaporkan berkorelasi positif dengan kadar hara P (Carrenho et al. 2001). Spora G. etunicatum dilaporkan tetap melimpah sekalipun tanah mineral asam Lampung Timur diberi pupuk P takaran tinggi (200 kg ha -1 SP36) (Yusnaini et al. 2004).
tinggi dilaporkan mengubah produksi keme-limpahan, daya mengolonisasi, dan efektivitas propagul FMA (Johnson 1993; Bhadalung et al. 2005). Penggunaan fosfat berkelarutan rendah, misalnya batuan fosfat dan tepung tulang, sekalipun diberikan dengan takaran yang tinggi ternyata efektif untuk memelihara FMA dan meningkatkan pertumbuhan tanaman (Cardoso 1996; Nikolaou et al. 2002). Sylvia & Schenck (1983) melaporkan G. etunicatum merespon negatif terhadap superfosfat namun merespon positif terhadap P organik. Nisbah N/P substrat, jaringan tanaman, atau sumber P juga dilaporkan berpengaruh terhadap aras kolonisasi dan sporulasi FMA (Douds & Schenck 1990a,b; Blanke et al. 2005). Dapat disimpulkan bahwa tidak ada hubungan yang jelas antara hara P dengan berbagai jenis mikoriza arbuskula di alam.
negatif terhadap FMA (Roldan & Albaladejo 1993). Pengaruh pupuk kandang berkadar hara P tinggi terhadap kerapatan hifa lebih ditentukan oleh tekstur tanahnya, pengaruhnya lebih tinggi pada tanah bertekstur lempung daripada tanah geluh berpasir (Kabir et al. 1997). Hasil penelitian menunjukkan FMA mampu menyerap P organik dari senyawa fitat dalam medium tumbuh (Widiastuti 2004). Berbagai laporan tersebut menunjukkan bahan organik berpotensi meningkatkan kemelimpahan dan sporulasi FMA dalam tanah namun demikian hasil akhirnya ditentukan oleh jenis FMA, jenis tanaman inang, jenis bahan organik, derajat dekomposisi, kadar hara bahan organik khususnya fosfat, dan karakteristik tanah.
Perlakuan terhadap substrat atau pupuk, misalnya sterilisasi, juga dapat berpengaruh terhadap perkembangan FMA. Sterilisasi merupakan tindakan yang dilakukan untuk meniadakan jasad renik pengganggu. Sterilisasi pada produksi inokulum FMA pada dasarnya hanya dilakukan untuk meniadakan isolat FMA lain yang tidak diproduksi dan tidak bertujuan untuk meniadakan sama sekali jasad renik lain yang ada dalam sistem produksi inokulum. Sterilisasi dilaporkan berpengaruh tidak nyata terhadap kolonisasi mikoriza pada tanaman sorgum (Cavender et al. 2003) namun menurunkan kolonisasi dan kerapatan spora FMA pada rizosfir tanaman tomat (Manian et al. 1995). Namun demikian belum ada informasi perlu tidaknya dilakukan sterilisasi untuk sumber hara P lainnya, misal tepung tulang.
semakin tinggi nilai KRM (Guissou et al. 1998; Ba et al. 2000). Tanaman yang bersimbiosis dengan FMA pada umumnya memiliki nilai NTA yang lebih rendah dibandingkan dengan yang tidak bermikoriza (Piccini et al. 1988; Khalil et al. 1994). Nisbah tajuk akar tajuk (NTA) merupakan gambaran translokasi karbon dari bagian atas tanaman ke akar. Nilai NTA yang mengecil akibat perubahan dinamika rizosfir mencerminkan meningkatnya translokasi karbon ke akar dan merupakan salah satu mekanisme tanaman untuk mendapatkan hara yang menjadi pembatas. Translokasi karbon ke akar diwujudkan dalam bentuk peningkatan biomassa akar sehingga meningkatkan permukaan yang dapat dikolonisasi FMA.
Uraian di atas mengindikasikan adanya potensi berbagai sumber P anorganik berkelarutan rendah dan P organik dengan derajat dekomposisi tertentu untuk digunakan dalam menyusun formulasi inokulan FMA. Namun demikian, perlu disadari bahwa banyak faktor yang terlibat dalam interaksi antara berbagai jenis sumber P dengan pembentukan dan perkembangan simbiosis MA dengan tanaman pada suatu lingkungan tertentu. Para peneliti mikoriza di Indonesia jarang sekali melakukan pengujian keterkaitan antara faktor yang mempengaruhi pertumbuhan tanaman dengan aktivitas FMA. Keberhasilan inokulasi FMA umumnya hanya dinilai berdasarkan indikator tanaman, misalnya kenaikan bobot kering tanaman dan bobot kering hasil, serapan hara – khususnya fosfor (Rossiana & Supriatun 2003; Simarmata & Herdiani 2003), dan beberapa diantaranya terhadap perubahan sifat-sifat tanah (Bertham 2006). Masih jarang yang melakukan studi korelatif antara indikator tanaman, FMA (kolonisasi dan jumlah spora) dan karakteristik tanah.
Interaksi Bibit Jati- Bahan Organik-Fungi Mikoriza Arbuskula
sekalipun dari hutan tanaman yang dikelola secara intensif, tetap tidak dapat memenuhi permintaan dari konsumen. Kondisi demikian, ditambah kemudahan penguasaan silvikultur jati, telah menarik minat masyarakat luas untuk ikut menanam dan memperdagangkan jati.
Jati (Tectona grandis L.f.) merupakan salah satu pohon penghasil kayu utama dunia. Hutan alam jati pada umumnya banyak terdapat di India, Myanmar, Thailand, dan Laos dengan luasan sekitar 23 juta ha. Jati juga tumbuh pada hutan tanaman pada sedikitnya 36 negara di kawasan tropik dengan luasan sekitar 5.7 juta ha (Bhat & Ma 2004). Hutan tanaman jati mencakup 75% dari hutan tanaman berkayu utama dunia (FAO 2001), dengan penyebaran 43% di India, 31% di Indonesia, 7% di Thailand, 6% di Myanmar, dan 5% di Afrika tropik. Jati diyakini sudah tumbuh di Jawa selama beberapa abad dan telah beradaptasi dengan lingkungan setempat dan kemudian secara alami menghasilkan keturunan yang ada sekarang ini (White 1991; Ball 1999; Pandey & Brown 2000). Pada awalnya jati di Indonesia diyakini berasal dari India, namun hasil analisis molekuler menunjukkan jati yang tumbuh di P. Jawa sesungguhnya berasal dari Thailand utara (Verhaegen et al. 2010). Di Indonesia, khususnya di Pulau Jawa, jati umumnya tumbuh pada elevasi 100 – 700 m dari permukaan laut. Tinggi pohon dapat mencapai 39 – 45 m pada kondisi yang sesuai dan kayunya dapat dipanen pada umur 60 tahun. Dari Pulau Jawa sendiri setiap tahun berhasil dipanen sekitar 820.000 m3, angka tersebut merupakan 44% dari total volume kayu yang diproduksi di P. Jawa namun merupakan 80% total pendapatan dari produksi kayu nasional (Bailey & Haryanto 2005).
genetik sama dengan induknya. Penggunaan bahan tanam yang berasal dari perbanyakan vegetatif, konvensional ataupun modern, dapat mengurangi daur produksi jati dan menghasilkan pohon yang mewarisi sifat unggul tetuanya.
Perbanyakan vegetatif pada dasarnya ialah upaya memperbanyak bahan tanaman atau bibit dengan menggunakan bagian vegetatif tanaman seperti batang, akar, dan daun atau tajuk (Hartmann et al. 1997). Perbanyakan vegetatif bibit jati dapat dilakukan secara konvensional misalnya stek, penempelan (okulasi), dan penyambungan (Mahfudz et al. 2003) atau teknologi modern menggunakan kultur jaringan (Herawan 2003). Sekalipun teknik kultur jaringan mampu menghasilkan bibit dalam waktu yang lebih singkat dan keseragaman yang lebih tinggi namun harga jual bibit lebih mahal dibandingkan teknik konvensional (Herawan 2003).
Bibit jati, hasil perbanyakan generatif atau vegetatif, umumnya dipelihara dalam wadah yang berisi media tumbuh tertentu di persemaian. Persemaian jati, atau tanaman hutan lainnya, berskala kecil sampai menengah, baik yang dikelola secara individual ataupun oleh sekelompok masyarakat, telah berkembang sebagai aktivitas ekonomi yang penting artinya di Indonesia. Namun demikian, produksi bibit jati atau tanaman hutan lainnya pada skala demikian seringkali menghasilkan bibit yang bermutu rendah karena kurangnya kemampuan mendapatkan sumber daya genetik yang bermutu tinggi, pemahaman mengenai praktek budidaya tanaman yang tepat, tenaga ahli untuk memelihara dan menilai mutu bibit (Harrison et al. 2008). Sumber benih yang tidak jelas, komposisi media yang keliru, pemberian pupuk yang tidak tepat jenis dan jumlah, dan faktor lingkungan tumbuh merupakan faktor utama yang menjadi penyebab rendahnya mutu bibit yang dihasilkan oleh produsen bibit berskala kecil sampai menengah (Harrison et al. 2008).
bengkok karena pertumbuhannya dibatasi oleh volume wadah (Arnold 1992). Akar demikian cenderung tetap tumbuh melingkar dan hanya sebagian kecil yang tumbuh radial ketika ditanam di lapangan sehingga menurunkan vigor dan kekokohan mekanis serta meningkatkan kematian bibit di lapangan (Arnold & Young 1991; Beeson & Newton 1992; Woodall & Ward 2002; Tsakaldimi & Ganatsas 2006). Modifikasi wadah, misalnya berbentuk segi empat atau berlubang di bagian bawahnya atau dari bahan yang mudah ditembus akar ternyata tidak selalu memuaskan hasilnya. Pertumbuhan akar yang lebih baik hanya terdapat pada bagian samping wadah akan tetapi deformasi akar tetap terjadi pada bagian bawah wadah (Warren & Blazich 1991; Beeson & Newton 1992). Pemangkasan akar merupakan salah satu cara untuk mengoreksi buruknya pertumbuhan akar pada bibit dalam wadah.
Pemangkasan akar mengganggu serapan hara dan air dan meningkatkan infeksi patogen sehingga mempengaruhi pertumbuhan tanaman. Tanaman kemudian meresponnya dengan cara meningkatkan pertumbuhan akar lateral dan menurunkan pertumbuhan tajuk atau menurunkan nisbah tajuk akar (Geisler & Ferree 1984). Sistem perakaran yang tumbuh dengan struktur yang bagus, yaitu dengan jumlah akar lateral fibrous yang tinggi, merupakan salah satu indikator dari bibit yang bermutu tinggi. Hasil penelitian menunjukkan pemangkasan akar berpengaruh negatif terhadap pertumbuhan tajuk dan tinggi bibit namun tidak mempengaruhi garis tengah batang, dengan kata lain meningkatkan kekokohan bibit yang merupakan salah satu indikator mutu bibit (Awang & De Chavez 1993). Laju pemulihan diri melalui peningkatan pertumbuhan akar lateral tersebut dipengaruhi oleh faktor genetik tanaman, lingkungan pertumbuhan, dan budidaya tanaman (Hartmann et al. 1997; Ludwig-Müller et al. 2005; Husen & Pal 2007).
terkena cekaman lengas, mudah terganggu oleh penanganan yang tidak sempurna selama pengangkutan dan penanaman di lapangan, dan peka terhadap persaingan dengan akar gulma (Fujimori 2001). Oleh sebab itu bibit akar telanjang lebih cocok untuk tanaman yang bersifat tumbuh lambat pasca perkecambahan (Fujimori 2001). Agar tumbuh lebih baik di lapangan sebaiknya bibit akar telanjang diberi perlakuan tambahan selama di persemaian, misalnya dengan pemupukan atau diinokulasi dengan FMA..
Penggunaan pupuk buatan untuk meningkatkan pertumbuhan bibit merupakan hal yang sudah biasa dilakukan oleh produsen bibit. Suhartati dan Nursyamsi (2006) melaporkan pemberian 50 g pupuk NPK per tanaman meningkatkan tinggi dan garis tengah batang bibit jati masing-masing sebesar 30% dan 9%. Penggunaan pupuk buatan bukan merupakan pilihan tunggal mengingat harganya yang cenderung semakin meningkat dan seringkali menghilang ketika diperlukan petani, memerlukan air untuk melarutkannya, dan karakteristik kimianya yang dapat berubah jika diberikan pada tanah-tanah bermasalah. Oleh karena itu perlu dicarikan alternatif penggantinya, yaitu dalam bentuk bahan alami yang dapat diproduksi sendiri misalnya kompos dan vermikompos atau yang banyak dijumpai dalam kehidupan sehari-hari misalnya tepung tulang.
batang bibit jati asal Cepu masing-masing sebesar 50 – 69% dan 28 – 41% (Irianto et al. 2003). Sifat baik dari bahan organik demikian bukan bermakna bahwa bahan organik dapat sepenuhnya menggantikan tanah yang digunakan sebagai medium tumbuh selama ini. Penggunaan bahan organik yang tidak terkontrol mutunya dapat menimbulkan immobilisasi atau ketidak seimbangan hara, keracunan logam berat atau senyawa organik beracun, sehingga berakibat buruk terhadap pertumbuhan bibit. Sifat ragawinya yang sarang menyebabkan bibit mudah roboh dan menderita kekurangan air atau hara.
Penggunaan FMA merupakan alternatif yang lebih baik dibandingkan penggunaan pupuk buatan. Fungi MA telah diketahui menghasilkan beraneka manfaat sehingga tanaman mampu mengatasi cekaman hayati dan nirhayati (Smith & Read 2008; Gianinazzi et al. 2010; Smith et al. 2010). Manfaat nyata inokulasi FMA dalam produksi bibit ialah meningkatkan daya tumbuh bibit ketika ditanam di lapangan. Bibit tidak bermikoriza ketika dipindah ke lapangan dari pesemaian seringkali tumbuh kerdil dan daunnya lebih banyak yang nekrosis dibandingkan dengan bibit bermikoriza (Landis & Amaranthus 2009).
melaporkan terdapat interaksi nyata antara jenis FMA, pupuk, dan medium tumbuh terhadap pertumbuhan bibit jati asal Cepu. Sangaji (2004) melaporkan inokulan FMA diperkaya dengan tepung tulang berpengaruh nyata meningkatkan pertumbuhan bibit jati namun berpengaruh tidak nyata terhadap kolonisasi FMA dan jumlah spora dalam media tumbuh. Suwandi et al. (2006) melaporkan pemberian 1.5 – 3 g inokulan FMA (tidak disebutkan jenisnya) dikombinasikan dengan pemberian 2 – 4 g pupuk NPK nyata meningkatkan tinggi, garis tengah batang, dan panjang akar bibit jati yang ditumbuhkan dalam polybag yang di Kampar, Riau. Arif et al. (2009) melaporkan inokulum G. etunicatum dan Glomus sp. yang diperkaya dengan vermikompos sampai 40% meningkatkan bobot kering bibit jati muna masing-masing sebesar 529 dan 500% dibandingkan kontrol. Informasi demikian menunjukkan adanya kombinasi spesifik antara provenan bibit jati, jenis FMA, dan pupuk organik atau bahan alami sebagaimana telah dilaporkan pada tanaman lain. Namun demikian, sejauh ini belum pernah diteliti interaksi demikian pada bibit jati provenan lain, misalnya provenan Solomon yang belakangan ini banyak diperdagangkan masyarakat.
sebagai berikut (a) pohon induk asalan, (b) pohon plus dari hutan alam atau hutan tanaman, (c) tegakan benih teridentifikasi, (d) tegakan benih terseleksi, (e) areal produksi benih, (f) tegakan benih provenan, (g) persemaian generasi pertama, (h) persemaian klon generasi pertama (Hendromono 2003).
Karakteristik morfologi yang umum digunakan untuk menilai mutu bibit ialah tinggi, garis tengah batang, nisbah kekokohan bibit (NKB), nisbah tajuk akar (NTA), morfologi dan massa akar (Duryea 1985; van den Driessche 1994; Claussen 1996; Mohamed et al. 1998; South 2000) dan potensi regenerasi akar (jumlah, panjang, jumlah tajuk akar, luas permukaan dan volume) (Tinus 1996), dan indeks mutu bibit (IMB) (Dickson et al. 1960). Tinggi dan garis tengah batang bibit merupakan indikator mutu bibit yang paling mudah diukur sekalipun keduanya tidak selalu tepat menggambarkan kemampuan bibit tumbuh baik di lapangan. Tinggi bibit dipandang merupakan gambaran kecepatan tumbuh bibit pada suatu lingkungan pertumbuhan tertentu. Mantovan (2002) melaporkan tinggi bibit umur 20 hari berkorelasi positif dengan pertumbuhan tanaman Prosopis flexuosa umur 3 dan 7 tahun. Loha et al. (2006) melaporkan rerata tinggi bibit di persemaian berkorelasi positif dengan pertumbuhan tanaman Cordia africana Lam. umur 4 dan 8 bulan. Zheng et al. (2009) melaporkan tinggi bibit berkorelasi positif dengan garis tengah batang tanaman Trigonobalanus doichangensis umur 4 tahun. Namun demikian, penampilan bibit di persemaian tidak selalu menggambarkan pertumbuhan sesungguhnya di lapangan (Mantovan 2002). Bibit yang bergaris tengah batang besar lebih tahan terhadap pembengkokan dan kerusakan mekanis selama pengangkutan atau penamaman, daya hidup dan volume kayu yang lebih tinggi (Johnson & Cline 1991). Ukuran garis tengah batang seringkali berkaitan erat dengan ukuran akar. Bibit dengan garis tengah batang yang lebih besar umumnya memiliki sistem perakaran yang lebih besar pula (Fujimori 2001). Garis tengah batang merupakan penduga ukuran sistem perakaran yang lebih baik dibandingkan tinggi tanaman.