i
PERTUKARAN GAS CO
2STUDI KASUS DI SELAT NASIK, BELITUNG DAN ESTUARI
DONAN, CILACAP
UDARA-LAUT DI PERAIRAN
PESISIR:
A F D A L
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
ii
PERNYATAAN MENGENAI TESIS DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul Pertukaran gas CO2 Udara–Laut di Peraian Pesisir: Studi Kasus di Selat Nasik Belitung dan Estuari Donan Cilacap adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juli 2011
iii
ABSTRACT
AFDAL. Air-Sea CO2 Gas Exchange In Coastal Waters: A Case Study In Nasik Strait, Belitung and Donan Estuary, Cilacap. Under supervised by RICHARDUS F. KASWADJI, ALAN F. KOROPITAN.
Marine carbonate system plays an important role in the air-sea CO2 gas exchange. Aim of the present study is to investigate the air-sea flux of CO2 in Nasik Strait, Belitung and Donan Estuary, Cilacap. Field observation was carried out during April and June, 2010, where the observed parameters consisted of temperature, salinity, pH, dissolved inorganic carbon (DIC), total alkalinity (TA), primary productivity of phytoplankton and nutrients (phosphate and silicate). Particularly the partial pressure of CO2 (pCO2) in sea surface, it was calculated using ABIOTIC model of the ocean carbon cycle model intercomparison project phase-2. Analysis results of the marine carbonate system showed that generally Nasik Strait waters and Donan Estuary act as a source (release) of CO2 to the atmosphere. The CO2 flux in Donan Estuary, mangrove waters, coral reef waters and coastal waters (non mangrove and coral reef) vary between 6.76–7.72, 3.06– 3.19, 0.96–1.45 and 2.77–2.98 mmolC.m-2.d-1, respectively. The present study found that the CO2 uptake by phytoplankton (photosynthesis) is not significantly affect the CO2 flux. In this case, the decomposition of particulate organic carbon tends to give significant contribution to the CO2
Keywords: CO
flux.
iv
AFDAL. Pertukaran Gas CO2 Udara-Laut di Perairan Pesisir: Studi Kasus di Selat Nasik, Belitung dan Estuari Donan, Cilacap. Dibimbing oleh RICHARDUS F. KASWADJI, ALAN F. KOROPITAN.
Penelitian dengan topik pertukaran gas CO2 udara-laut di perairan pesisir: studi kasus di Selat Nasik, Belitung dan Estuari Donan, Cilacap ini dilaksanakan dengan tujuan untuk mengkaji fluks CO2 udara–laut di perairan Selat Nasik Belitung dan perairan Estuari Donan Cilacap. Penelitian ini diharapkan dapat memberikan kontribusi bagi perkembangan ilmu pengetahuan kelautan di Indonesia, khususnya mengenai peranan perairan pesisir tropis dalam mengontrol fluks CO2
Pengambilan contoh air dilakukan di perairan Selat Nasik, Kabupaten Belitung pada bulan April 2010 dan di perairan Estuari Donan, Cilacap pada bulan Juni 2010. Di perairan Selat Nasik, pengukuran dan pengambilan contoh air laut dilakukan di 3 stasiun pada lokasi yang berbeda yaitu: Stasiun 1 pada lokasi di sekitar ekosistem mangrove, Stasiun 2 pada lokasi sekitar terumbu karang dan Stasiun 3 pada lokasi yang tidak ada pengaruh mangrove dan terumbu karang (perairan laut pesisir). Pengambilan sampel dilakukan 2 kali dalam sehari yaitu pada waktu pagi (06:35–08:15) dan siang hari (12:00–13:25) untuk Stasiun 1 dan 2, sedangkan pada Stasiun 3 pengambilan sampel dilakukan sebanyak 3 kali (pagi, siang dan malam hari (19:00) untuk melihat sejauh mana peranan produktivitas primer fitoplankton dalam menurunkan CO
.
2 dalam air laut. Di perairan Estuari Donan Cilacap, pengukuran dan pengambilan contoh air laut di lakukan pada 5 stasiun dari muara menuju sungai. Parameter yang diukur adalah suhu, salinitas, produktivitas primer, fosfat, silikat, CO2 atmosfir dan sistem CO2 (pH, total alkalinitas, DIC dan tekanan parsial CO2
Hasil menunjukkan bahwa pH di perairan Selat Nasik berkisar antara 7,71–8,03. Di perairan Estuari Donan, Cilacap pH berkisar antara 7,65–7,95 dengan rata-rata 7,83±0,117. Konsentrasi DIC di perairan Selat Nasik berkisar antara 1912,15–2119,03 µmol/kg. Di perairan Estuari Donan konsentrasi DIC berkisar antara 1842,25–1957,82 µmol/kg dengan rata-rata 1901,57±49,76 µmol/kg. Total alkalinitas di perairan Selat Nasik berkisar antara 2130,28– 2247,28 µmol/kg. Di perairan Estuari Donan Cilacap total alkalinitas berkisar antara 1954,27–2032,44 µmol/kg dengan rata-rata 1901,57±49,76 µmol/kg. Tekanan parsial CO
). Pengukuran total alkalinitas, produktivitas primer dan nutrien dilakukan di laboratorium produktivitas primer dan kimia hara Pusat Penelitian Oseanografi-LIPI Jakarta. Pengukuran konsentrasi DIC dilakukan di laboratorium pengujian pusat aplikasi teknologi isotop dan radiasi BATAN, pasar jumat Jakarta.
v
vi
© Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
vii
PERTUKARAN GAS CO
2STUDI KASUS DI SELAT NASIK, BELITUNG DAN ESTUARI
DONAN, CILACAP
UDARA-LAUT DI PERAIRAN
PESISIR:
A F D A L
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Kelautan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
viii
ix Judul Tesis : Pertukaran Gas CO2 Nama : Afdal
Udara-Laut di Perairan Pesisir: Studi Kasus di Selat Nasik, Belitung dan Estuari Donan, Cilacap
NRP : C551080171
Disetujui Komisi Pembimbing
Ketua
Dr. Ir. Richardus F. Kaswadji, M.Sc
Anggota
Dr. Alan F. Koropitan, S.Pi, M.Si
Diketahui
Ketua Program Studi Ilmu Kelautan
Dekan Sekolah Pascasarjana
Dr. Ir. Neviaty P. Zamani, M.Sc Dr. Ir. Dahrul Syah, M.Sc.Agr
x
Puji syukur kehadirat Allah SWT atas segala rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan karya ilmiah (tesis) dengan judul “Pertukaran Gas CO2
Dalam kesempatan ini penulis menyampaikan terima kasih yang sebesar-besarnya kepada semua pihak yang telah membantu dalam penyusunan karya ilmiah ini, terutama kepada:
Udara–Laut di Perairan Pesisir: Studi Kasus di Selat Nasik, Belitung dan Estuari Donan, Cilacap” .
1. Bapak Dr. Ir. Richardus F. Kaswadji, M.Sc dan Bapak Dr. Alan F. Koropitan, S.Pi, M.Si selaku Ketua Komisi Pembimbing dan anggota komisi pembimbing yang telah penuh perhatian dan kesabaran dalam memberikan bimbingan dan arahan hingga penyusunan tesis ini selesai.
2. Bapak Dr. Ir. Tri Prartono, MSc selaku penguji luar komisi yang telah banyak memberikan saran dan masukan dalam penulisan tesis ini.
3. Bapak Dr. Ir. Iwayan Nurjaya, M.Sc selaku perwakilan dari Program Studi IKL yang telah memberikan koreksian dan saran dalam penulisn tesis ini. 4. Ibu Dr. Suhartati M. Nasir dan Ibu Lily M.G. Panggabean, MSc., yang telah
mengikutkan penulis dalam kegiatan penelitian di perairan Selat Nasik, Belitung dan Estuari Donan, Cilacap.
5. Bapak Sumidjo Hadi Riyono (Teknisi Laboratorium Produktivitas Primer, P2O-LIPI) atas bantuannya selama penulis kuliah di pascasarjana IPB.
6. Ibu Ristin (Peneliti Puslitbang Teknologi Isotop dan Radiasi, BATAN, Jakarta) atas bantuannya dalam analisis DIC.
7. Isteri tercinta Ade Wirma Yulia dan anak-anakku tersayang Nabil Zahra Aldenda dan Gahzy Firjatullah Aldenda yang telah memberikan perhatian dan dukungan dalam menyelesaikan tesis ini.
8. Ibunda tercinta Wisnimar atas do’a dan dukungannya.
9. Kembaranku Afdil, SPt yang telah memberikan bantuan moril dan materil selama kuliah di pascasarjana IPB.
10.Saudara-saudaraku Dra. Delwita, MPd, Dra. Delinoviar, Pentri Desmon, Nelma Eriyanti, Toni Irfan dan Ade Suryani, SPd yang telah memberikan dukungan selama penulis kuliah di Pascasarjana IPB.
11.Rekan-rekan mahasiswa program studi Ilmu Kelautan angkatan 2008 (Ratno, Agung, Sabam, Sayyid, Ayu) dan pihak-pihak lain yang telah membantu kelancaran dalam penyusunan tesis ini.
Penulis menyadari bahwa tesis ini belum sempurna, oleh karena itu penulis mengharapkan kritik dan saran untuk perbaikan tesis ini. Penulis berharap tesis ini dapat bermanfaat bagi kita semua.
Bogor, Juli 2011
xi
RIWAYAT HIDUP
Penulis dilahirkan di Bukittinggi pada tanggal 16 Maret 1979 dari Ayah Alm. Djalius dan Ibu Wisnimar. Penulis merupakan anak ke enam dari delapan bersaudara. Mempunyai istri bernama Ade Wirma Yulia dan dikaruniai sepasang buah hati bernama Nabil Zahra Aldenda dan Gahzy Firjatullah Aldenda.
Pendidikan SD, SMP dan SMA ditempuh di Kecamatan Tilatang Kamang Kabupaten Agam Sumatera Barat. Pada tahun 1997 penulis meneruskan pendidikan sarjana di Universitas Andalas pada Fakultas Matematika dan Ilmu Pengetahuan Alam, Program Studi Biologi dan lulus pada tahun 2002. Pada tahun 2003 Penulis diterima sebagai staf peneliti di laboratorium plankton dan produktivitas primer, Pusat Penelitian Oseanografi - LIPI. Semenjak tahun 2008 penulis menjalani studi di Program Studi Ilmu Kelautan pada Program Pascasarjana IPB.
xii
DAFTAR LAMPIRAN... xvii
1 PENDAHULUAN... 1
1.1 Latar Belakang... 1
1.2 Pendekatan masalah... 2
1.3 Tujuan dan Manfaat Penelitian... 3
2 TINJAUAN PUSTAKA... 4
2.1 Sistem karbonat laut... 4
2.2 Siklus karbon dan pertukaran CO2 udara-laut... 6
2.2.1 Laut global... 6
3.1 Waktu dan Lokasi Penelitian... 15
3.2 Bahan dan Alat Penelitian... 17
3.3 Prosedur Penelitian... 17
3.3.1. Sistem CO2... 17
3.3.2. Produktivitas Primer... 18
3.4 Analisis data... 18
3.4.1 Sistem CO2... 18
3.4.2 Pertukaran CO2 udara-laut... 19
3.4.3 Produktivitas Primer... 20
4 HASIL DAN PEMBAHASAN... 21
4.1 Sistem CO2... 21
4.1.1 Selat Nasik... 21
4.1.2 Estuari Donan... 24
4.2 Produktivitas Primer... 27
4.2.1 Selat Nasik... 27
4.2.2 Estuari Donan... 28
4.3 Pertukaran CO2 udara-laut... 29
xiii
4.3.2 Estuari Donan... 30
4.4 Pembahasan... 31
4.4.1 Selat Nasik... 31
4.4.2 Estuari Donan... 36
5 KESIMPULAN DAN SARAN... 40
DAFTAR PUSTAKA... 41
xiv
DAFTAR TABEL
Halaman
xv
DAFTAR GAMBAR
Halaman
1. Skema pendekatan masalah... 3 2. Siklus karbon global. Panah menunjukkan fluks (dalam PgC/th) antara
atmosfir dan darat, dan atmosfir dan laut. Angka dalam kotak menunjukkan stok (PgC). Panah hitam = fluks alami; Panah merah = fluks antropogenik (Sarmiento and Gruber, 2002)... 7 3. Skema pompa fisis dan sirkulasi termohalin yang membawa CO2, yang
menggambarkan interkoneksi perairan laut dunia (Hansell and Carlson, 2001) ... 8 4. Lokasi penelitian di perairan Selat Nasik, Belitung, April 2010... 15 5. Lokasi penelitian di perairan Estuari Donan Cilacap, Juni
2010... 16 6. Nilai pH di perairan Selat Nasik pada pengamatan pagi, siang dan
malam hari, April 2010... 21 7. Konsentrasi DIC di perairan Selat Nasik pada pengamatan pagi, siang
dan malam hari, April 2010... 22 8. Konsenrasi total alkalinitas di perairan Selat Nasik pada pengamatan
pagi, siang dan malam hari... 23 9. Tekanan parsial CO2 di perairan Selat Nasik pada pengamatan pagi,
siang dan malam hari... 24 10. Distribusi nilai pH di perairan Estuari Donan Cilacap, Juni 2010... 25 11. Distribusi konsentrasi DIC di perairan Estuari Donan Cilacap, Juni 2010 25 12. Distrbusi Total Alkalinitas di perairan Estuari Donan Cilacap, Juni
2010... 26 13. Distribusi tekanan parsial CO2 di perairan Estuari Donan Cilacap, Juni
2010... 27 14. Laju fotosintesis (GPP), respirasi, dan produksi primer bersih (NPP) di
perairan Selat Nasik, April 2010. [A=pagi (6:35–8:15), B=siang (12:00–13:25)]... 28 15. Laju fotosintesis (GPP), respirasi dan produksi primer bersih (NPP) di
xvi
16. Fluks CO2 di perairan Selat Nasik, April 2010... 30 17. Fluks CO2 di perairan Estuari Donan Cilacap, Juni 31
2010...
18. Skema siklus karbon pada Stasiun 1 (perairan sekitar mangrove) pada pengamatan pagi (A) dan siang hari (B). FCO2, GPP, NPP, Respirasi dan Dekomposisi dalam satuan mmolC/m2/hari... 32 19. Skema siklus karbon pada Stasiun 2 (perairan sekitar terumbu karang)
pada pengamatan pagi (A) dan siang hari (B). FCO2, GPP, NPP, Respirasi dan Dekomposisi dalam satuan mmolC/m2/hari... 34 20. Skema siklus karbon pada Stasiun 3 (perairan laut) pada pengamatan
pagi (A) dan siang hari (B). FCO2, GPP, NPP, Respirasi dan Dekomposisi dalam satuan mmolC/m2/hari... 36 21. Skema siklus karbon di perairan Estuari Donan. FCO2, GPP, NPP,
xvii
DAFTAR LAMPIRAN
Halaman
1. Paramater fisika-kimia dan sistem CO2 di perairan Selat Nasik... 46
2. Data produktivitas primer perairan Selat Nasik... 47
3. Paramater fisika-kimia dan sistem CO2 di perairan Estuari Donan... 47
1 PENDAHULUAN
1.1. Latar Belakang
Pemanasan global merupakan isu yang paling banyak mendapat perhatian akhir-akhir ini. Gas rumah kaca diantaranya, CO2, CH4, dan N2O merupakan penyebab terjadinya pemanasan global (IPCC 2001). Hal ini diyakini bahwa dengan meningkatnya kadar gas-gas rumah kaca di atmosfir bumi menyebabkan kenaikan suhu global serta perubahan pola curah hujan (IPCC 2007). CO2 sebagai gas rumah kaca utama mendapat perhatian yang lebih besar di seluruh dunia, karena keterlibatannya dalam siklus biogeokimia wilayah pesisir dan laut terbuka (Takahashi et al., 2002; Borges, 2005; Borges et al., 2005).
Perairan wilayah pesisir memainkan peranan utama dalam siklus biogeokimia di laut meskipun hanya mempunyai luas <7% luas lautan dunia dan mempunyai volume <0,5% volume lautan global (Gattuso et al., 1998). Hal ini disebabkan oleh perairan pesisir mempunyai laju produksi primer baru secara signifikan lebih tinggi daripada di lautan terbuka akibat besarnya pasokan nutrien dari sungai dan proses upwelling, dan tingginya proses dekomposisi material organik (Chen dan Borges, 2009).
Penelitian sebelumnya menunjukkan bahwa perairan pesisir, khususnya perairan sekitar mangrove dan estuari merupakan net source CO2 ke atmosfir (Chen dan Borges, 2009), demikian juga dengan perairan sekitar terumbu karang (Gattuso et al., 1996). Beberapa peneliti lainnya menyatakan bahwa perairan pesisir di daerah temperate dan lintang tinggi berperan sebagai sink CO2 dari atmosfir, sedangkan perairan pesisir di daerah subtropis dan tropis berperan sebagai source CO2
Namun penelitian lainnya juga mencatat ada perairan pesisir di daerah tropis yang berperan sebagai sink CO
ke atmosfir (Cai et al., 2003; Wang and Cai, 2004; Borges, 2005).
2
terutama di perairan Indonesia masih langka, dan masih banyak dibutuhkan untuk menilai secara kuantitatif peran laut dan pesisir dalam siklus karbon global.
1.2. Pendekatan Masalah
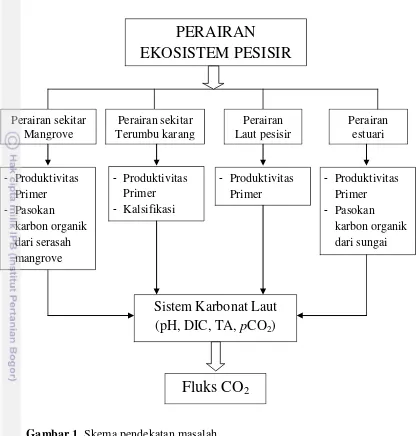
Skema pendekatan masalah ditampilkan dalam Gambar 1. Pertukaran CO2 antara atmosfir dan laut, yang diatur oleh proses-proses fisik dan biologis tidak terdistribusi secara merata terhadap ruang dan waktu, sehingga perbedaan kharakteristik pada ekosistem perairan pesisir dan laut yang menyebabkan faktor-faktor yang mengontrol fluks CO2 udara-laut pada masing-masing ekosistem (perairan sekitar mangrove, perairan sekitar terumbu karang, laut peisisr dan estuari) juga berbeda.
Perairan sekitar mangrove dan estuari dicirikan oleh tingginya pasokan karbon organik baik yang berasal dari sungai maupun dari ekosistem mangrove yang akan menyumbang CO2 ke kolom perairan melalui proses dekomposisi, sedangkan penyerapan CO2 dilakukan oleh fitoplankton melalui proses fotosintesis. Perairan sekitar terumbu karang dicirikan oleh adanya proses kalsifikasi yang menyumbang CO2 ke dalam kolom perairan, sedangkan penyerapan CO2 melalui proses fotosintesis dilakukan oleh fitoplankton, zooxanthella dan makroalga. Di perairan laut, fitoplakton memegang peranan utama dalam penyerapan CO2.
Gambar 1. Skema pendekatan masalah
1.3Tujuan dan Manfaat Penelitian
Penelitian ini dilaksanakan dengan tujuan mengkaji fluks CO2 udara–laut di perairan Selat Nasik Belitung dan perairan Estuari Donan Cilacap. Penelitian ini diharapkan dapat memberikan kontribusi bagi perkembangan ilmu pengetahuan kelautan di Indonesia, khususnya mengenai peranan perairan pesisir tropis dalam mengontrol fluks CO2.
4
2 TINJAUAN PUSTAKA
2.1 Sistem karbonat laut
Karbon dioksida (CO2) termasuk gas yang reaktif dan banyak terdapat dalam air laut. Keberadaan karbon dioksida di laut pada umumnya berasal dari udara melalui proses difusi, aktivitas metabolisme dari organisme laut terutama respirasi dan hasil penguraian zat organik oleh mikroorganisme. Total jumlah karbon di laut diperkirakan 50 kali lebih besar dibandingkan jumlah karbon yang ada di atmosfir, dan pertukaran karbon laut dan atmosfir terjadi dalam skala waktu beberapa ratus tahun (IPCC, 2001).
Karbon dioksida yang terlarut di dalam air laut ditemukan dalam tiga bentuk anorganik yang berbeda, yaitu CO2 bebas (≈0,5%), bikarbonat (86,5%) dan ion karbonat (13%), penjumlahan dari ketiganya disebut sebagai dissolved inorganic carbon atau yang dikenal dengan DIC atau ƩCO 2 (Zeebe and Wolf-Gladrow, 2001). Dalam berbagai literatur DIC juga disimbolkan dengan TCO2 dan CT (Dickson et al. 2007). Persamaannya adalah sebagai berikut:
DIC ≡ƩCO2 ≡ CT = [CO2] + [HCO3-] + [CO32-]... (1) Selanjutnya menurut Zeebe and Wolf-Gladrow (2001), di dalam air laut, CO2 berpartisipasi dalam reaksi kesetimbangan yang dikenal sebagai sistem karbonat. Keseimbangan antara CO2
CO
terlarut, bikarbonat, ion karbonat, dan ion H+ memberikan suatu sistem pengaturan pH di laut. Reaksi keseimbangan ini dapat ditulis sebagai berikut:
Persamaan dalam sistem karbonat melibatkan enam variabel yaitu [CO2], [HCO3-], [CO32-], [H+], DIC, dan Alkalinitas karbonat, ketika dua dari enam variabel diketahui, maka semua komponen lainnya dapat dihitung (Zeebe and Wolf-Gladrow, 2001; Dickson et al. 2007). Variabel yang cukup penting dalam sistem karbonat adalah total alkalinitas (TA) yang berkaitan erat dengan keseimbangan muatan dalam air laut. Dickson (1981) dalam Dickson et al. (2007) mendefinisikan TA sebagai "jumlah mol ion hidrogen yang setara dengan kelebihan akseptor proton (basa yang terbentuk dari asam lemah dengan konstanta disosiasi (K) ≤ 10 - 4,5 pada suhu 25°C) atas donor proton (asam dengan K > 10 - 4,5) dalam 1 kilogram sampel. Persamaannya adalah sebagai berikut:
TA = [HCO3-] + 2[CO32-] + [B(OH)4-] + [OH-] + [HPO42-] + 2[PO43-] + [SiO(OH)3-] + [NH3] + [HS-] – [H+] – [HSO4-] – [HF] – [H3PO4]... (3) Sistem karbonat di laut di pengaruhi oleh proses-proses metabolisme organisme seperti fotosintesis, respirasi, kalsifikasi dan dekomposisi, dimana perubahan pada sistem karbonat tersebut paling baik digambarkan oleh perubahan konsentrasi DIC dan TA (Zeebe and Wolf-Gladrow, 2001). Proses fotosintesis akan mengurangi konsentrasi DIC. Selanjutnya pembentukan CaCO3 dalam proses kalsifikasi menyebabkan turunnya konsentrasi DIC dan TA. Untuk setiap mol CaCO3 yang terbentuk atau diendapkan dari satu mol karbon dan satu mol ion positif Ca2+
Disamping proses-proses metabolisme, sistem karbonat dalam air laut juga dipengaruhi oleh suhu, salinitas dan tekanan dimana suhu, salinitas dan tekanan akan mempengaruhi nilai konstanta kesetimbangan (pK) (Zeebe and Wolf-Gladrow, 2001). Perubahan dari nilai pK menyebabkan perubahan dalam proporsi relatif CO
akan mengarah kepada penurunan DIC dan TA dengan rasio 1:2. Hasil dari berbagai proses tersebut juga akan mempengaruhi spesies karbonat dan pH air laut.
6
Contoh lain ketergantungan sistem karbonat terhadap suhu dan tekanan adalah air yang didinginkan dari suhu 25 sampai 0oC dan kemudian tenggelam dari permukaan (S = 35, dan p = 1 atm) ke laut dalam (S = 35 dan p = 300 atm pada kedalaman 3 km). Dalam contoh ini hanya ada hipotetis abiotik, karena proses biologis seperti produksi primer dan kalsifikasi yang merubah DIC dan TA di laut diabaikan. Penurunan suhu dari 25 sampai 0oC menyebabkan peningkatan besar dalam nilai-nilai pK, sedangkan perubahan tekanan dari 1 sampai 300 atm hanya menyebabkan peningkatan nilai pK yang kecil.
2.2Siklus karbon dan pertukaran CO2 udara-laut 2.2.1 Laut global
5,4 59,6 60 1,9 1,7
Sink perubahan daratan lahan
70,6 70 21,9 20
fluks dari sungai 50 `0,8
39
90,2 101 1,6
11
0,2
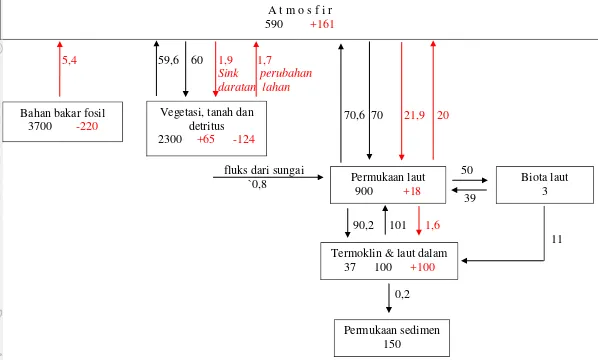
Gambar 2. Siklus karbon global. Panah menunjukkan fluks (dalam PgC/th) antara atmosfir dan darat, dan atmosfir dan laut. Angka dalam kotak menunjukkan stok (PgC). Panah hitam = fluks alami; Panah merah = fluks antropogenik (Sarmiento and Gruber, 2002).
A t m o s f i r 590 +161
Vegetasi, tanah dan detritus 2300 +65 -124
Permukaan laut 900 +18
Biota laut 3 Bahan bakar fosil
3700 -220
Termoklin & laut dalam 37 100 +100
8
Fluks CO2 udara-laut global merupakan fungsi terintegrasi dari pompa daya larut dan penyerapan biologis, proses-proses yang telah berjalan selama ribuan tahun (Hasell and Carlson, 2001). Kedua proses ini bertindak meningkatkan konsentrasi CO2 di dalam interior laut. Pompa fisis dibangkitkan oleh sirkulasi balik laut yang lamban dan lebih mudah terlarutnya CO2 di air dingin daripada di air hangat. Massa air yang dingin dan rapat (dense) di laut lintang tinggi, terutama di Atlantik Utara dan Laut Selatan (Southern Ocean), menyerap CO2 atmosfirik sebelum tenggelam (sink) ke interior laut. Air yang tenggelam ini akan diimbangi oleh transpor vertikal (upwelling) di bagian laut lainnya. Air yang naik ke atas ini akan menjadi hangat ketika mencapai permukaan sehingga CO2 menjadi kurang dapat larut dan sebagian diantaranya akan terlepas kembali ke atmosfir (melalui sebuah proses yang disebut pelepasgasan atau outgassing).
Gambar 3. Skema pompa fisis dan sirkulasi termohalin yang membawa CO2
Proses biologi yang ikut memandu distribusi regional dan musiman dari fluks CO
, yang menggambarkan interkoneksi perairan laut dunia (Hansell and Carlson, 2001).
2 adalah produksi primer kotor (GPP) oleh fitoplankton laut yang telah diperkirakan oleh Sarmiento and Gruber (2002) sebesar 50 PgC/th. Sebagiannya (39 PgC/th) dikembalikan ke DIC melalui respirasi autotrofik, dan sisanya menjadi produksi primer bersih yang diperkirakan sekitar 11 PgC/th (Sarmiento
and Gruber, 2002). Hasil karbon organik kemudian dikonsumsi oleh zooplankton atau menjadi detritus. Beberapa karbon organik dilepaskan dalam bentuk terlarut (DOC) dan oksidasi oleh bakteri dengan produksi DOC bersih yang masuk ke reservoir samudera. Penenggelaman partikel organik karbon (POC) yang terdiri dari organisme yang telah mati dan detritus bersama-sama dengan transfer vertikal DOC menciptakan suatu fluks karbon organik yang mengarah ke bawah dari permukaan samudera yang dikenal sebagai "produksi ekspor". Perkiraan untuk produksi ekspor global berkisar antara 10-20 PgC/th (Falkowski et al., 1998). Hanya sebagian kecil (±0,2 PgC/th) produksi ekspor yang mengendap pada sedimen (Sarmiento and Gruber, 2002).
2.2.2 Perairan Pesisir
Perairan pesisir didefinisikan sebagai wilayah yang membentang dari pantai ke landas kontinen, termasuk lereng benua dan inner estuari, sisa masa perairan laut disebut sebagai laut terbuka (Borges, 2005). Komponen perairan pesisir terdiri dari 6 ekosistem pesisir utama yaitu perairan estuari, mangrove, salt marsh, terumbu karang, sistem upwelling, dan continental shelf. Perairan pesisir merupakan daerah yang mempunyai siklus biogeokimia paling aktif di biosfer karena menerima masukan bahan organik dan nutrien yang besar dari daratan, dan adanya pertukaran materi dan energi dalam jumlah yang besar dengan laut terbuka (Gattuso et al 1998). Perairan pesisir menyumbang sekitar 20% dari total produksi bahan organik samudera, 90% dari total sedimen laut, dan menyumbang 30% dan 50% dari total produksi dan akumulasi partikel karbon anorganik (Gattuso et al., 1998; Wollast, 1998).
10
penyerapan CO2 oleh samudera yang diperkirakan 1,6 Pg C/th (Takahashi et al. 2002).
Chen (2004) mengemukakan bahwa secara umum perairan pesisir cenderung untuk menyerap CO2 pada musim dingin, sebagai konsekuensi dinginnya air permukaan dan dari proses biologi. Pada musim panas dan musim gugur, proses pemanasan menyebabkan proses respirasi dan dekomposisi organisme laut melepaskan kembali CO2 ke atmosfir, demikian juga dengan bakteri yang terlibat dalam proses produksi CH4 (gas metan) serta dalam produksi biologis dimetil sulfida (DMS) juga melepas gas rumah kaca ke atmosfir.
Penelitian lainnya menunjukkan bahwa perairan pesisir adalah source CO2 ke atmosfir (Ver et al. 1999). Hal ini didasarkan pada ketidakseimbangan antara angkutan sungai total sekitar 0,4 PgC/th dengan karbon organik yang terkubur di dasar perairan sekitar 0,14 PgC/th, jadi ada perbedaan 0,26 PgC/th yang mungkin dikembalikan ke atmosfir. Selanjutnya Fasham et al. (2001) melaporkan bahwa perairan pesisir adalah source CO2 ke atmosfir, dimana fluks CO2 udara laut sekitar 0,5 PgC/th. Mackenzie et al. (2000) menyatakan bahwa sebelum kegiatan antropogenik, perairan pesisir global adalah sistem autotrofik bersih dengan ekspor bersih dari karbon organik ke sedimen laut terbuka adalah 20 TmolC/th. Namun, hasil penelitian mereka berikutnya menyimpulkan bahwa meskipun zona pesisir yang dekat dengan daratan adalah source CO2 (8,4 x 1012 molC/th), zona pesisir yang jauh dari daratan adalah sink CO2 (28,4 x 1012 molC/th). Akibatnya perairan pesisir ditemukan sebagai net sink CO2 sekitar 20 x 1012 molC/th (0,24 PgC/th).
disimpan pada dasar perairan (Chen et al., 2003). Studi terbaru berdasarkan pengukuran C13, C14 dan N15 menunjukkan bahwa sebagian besar dari transportasi material organik ke perairan pesisir berasal dari daratan (Chen et al., 2003). Transportasi bersih DOC ke perairan pesisir adalah 0,60 PgC/th, dimana 0,32 PgC/th berasal dari darat, 0,28 PgC/th sisanya diproduksi di perairan pesisir dan mewakili 35 persen dari produksi karbon organik baru, atau 27 persen dari total produksi karbon baru (Cai et al., 2003).
2.2.2.1 Perairan Estuari
Cameron dan Pritchard (1963) mendefinisikan perairan estuari sebagai suatu badan perairan yang semi tertutup yang memiliki hubungan bebas dengan laut terbuka, dan di mana air laut diencerkan oleh air tawar yang berasal dari daratan. Batas hulu dari perairan estuari adalah batas pengaruh pasang surut (sungai pasang surut), di mana arus air dan proses sedimentasi menjadi sangat berbeda dibandingkan sungai, batas bawah dari perairan estuari adalah batas geografis pantai sesuai dengan mulut sungai. Perairan estuari mempunyai keragaman yang besar dalam hal geomorfologi, geokimia dan luas permukaan daerah aliran sungai, debit air tawar, dan pengaruh pasang surut, sehingga sangat mempengaruhi siklus biogeokimia karbon dan nutrien, stratifikasi vertikal, dan waktu tinggal air tawar (Borges 2005). Perairan estuari menerima bahan terlarut dan partikulat dalam jumlah besar, terutama karbon organik dan anorganik, dan nutrien yang dibawa oleh-sungai. Ini adalah sistem yang sangat dinamis yang dicirikan oleh gradien kuat dari senyawa biogeokimia, proses produksi dan degradasi bahan organik yang tinggi, dan adanya proses sedimentasi dan resuspensi yang intensif (Borges 2005).
12
yang berkaitan dengan perbedaan dalam kontribusi relatif kecepatan transfer gas terhadap turbulensi antarmuka perairan, kecepatan angin, arus air, dan topografi (Borges et al., 2004)
Berdasarkan hasil review dari Borges (2005) menunjukkan bahwa hampir semua perairan estuari adalah sumber bersih CO2 ke atmosfir. Di antara perairan estuari di Eropa, Scheldt dan Randers Fjord berada di batas tinggi dan rendah untuk fluks CO2 udara-laut. Estuari Randers Fjord adalah tipe estuari microtidal yang dicirikan oleh stratifikasi permanen yang kuat, sedangkan estuari Scheldt adalah estuari macrotidal yang ditandai oleh kolom air tercampur permanen. Meskipun secara keseluruhan Randers Fjord adalah sistem heterotrofik bersih, lapisan tercampur adalah autotrofik bersih sedangkan lapisan bawah adalah sangat heterotrofik. Emisi CO2 dari estuari Scheldt hampir 15 kali lebih tinggi dibanding Randers Fjord, meskipun NEP nya hanya 2 kali lebih rendah di Scheldt dibanding estuari Randers Fjord. Hal ini berhubungan dengan tingginya produksi bahan organik pada lapisan tercampur dan degradasi di lapisan bawah. Dengan cara ini CO2 hasil degradasi tidak langsung kembali ke atmosfir. Disamping itu lamanya waktu tinggal air tawar di estuari Scheldt (30-90 hari) dibanding estuari Randers Fjord (5-10 hari) menyebabkan pengayaan DIC di perairan estuari Scheldt lebih intensif.
2.2.2.2 Perairan Sekitar Mangrove
(2005) mengemukakan bahwa disamping proses dekomposisi material organik, dinamika DIC di perairan sekitar mangrove juga dipengaruhi oleh air poros dalam sedimen yang kaya dengan DIC, TA dan CO2 dan miskin oksigen yang dipompa oleh arus pasang surut.
Fluks CO2 udara-laut pada ekosistem mangrove global telah diperkirakan oleh Jennerjahn dan Ittekkot (2002). Jika ekspor bahan organik dari sistem perairan yang berdekatan dimasukkan maka fluks CO2 udara laut di perairan sekitar mangrove adalah 18,7 molC/m2/th. Jika ekspor bahan organik dengan sistem perairan yang berdekatan tidak dimasukkan maka fluksnya akan berkurang sekitar 50% (9,8 molC/m2/th). Dalam perhitungan ini diasumsikan bahwa fluks CO2 udara-laut hanya berhubungan dengan degradasi bahan organik yang berasal dari ekosistem mangrove, sedangkan pasokan karbon dari tempat lain seperti ekspor DIC dari sistem perairan yang berdekatan, emisi CO2 dari sedimen, dan masukan allochthonous tidak diperhitungkan.
2.2.2.3 Perairan Sekitar Terumbu Karang
Terumbu karang merupakan ekosistem perairan tropis dan sub tropis yang terdiri dari struktur karbonat yang didominasi oleh karang Scleractinia dan mikroalga (Borges, 2005). Ekosistem terumbu karang mewakili sekitar 2% dari luas permukaan continental shelf dan menyumbang sekitar 83% dan 50% dari produksi dan akumulasi partikel karbon anorganik ekosistem pesisir (Milliman, 1993). Terumbu karang berkembang pada tingkat kekeruhan rendah, perairan yang oligotrofik dengan suhu tahunan minimum 18oC dan mempunyai tingkat metabolisme karbon organik dan kalsifikasi yang tinggi (Gattuso et al., 1998). Meskipun perairan terumbu karang mempunyai laju fotosintesis dan respirasi yang tinggi, produksi ekosistem bersih di perairan tersebut mendekati nol (Gattuso et al., 1998). Proses kalsifikasi dan fotosintesis merupakan proses utama yang mempengaruhi siklus karbon di perairan sekitar terumbu karang. Fiksasi CO2 oleh produksi ekosistem bersih (NEP) biasanya rendah tetapi tingkat kalsifikasi tinggi sehingga menyebabkan proses kalsifikasi melepas CO2
Ca
ke perairan sekitarnya, sebagaimana dalam persamaan berikut:
2+
14
Konsentrasi CO2 meningkat sebesar 0,6 mol untuk setiap pengendapan 1 mol kalsium karbonat (CaCO3) dalam standar air laut (salinitas=35, suhu=25, TA=2370 µmol/kg, pCO2=365 ppm) (Gattuso et al., 1999). Rasio antara produksi CO2 dengan presipitasi CaCO3 pada umumnya tergantung pada keseimbangan termodinamika khususnya suhu dan salinitas (Ware et al., 1992; Frankignoulle et al., 1994).
Berdasarkan perkiraan global dari kalsifikasi bersih dan NEP, Ware et al. (1992) menghitung potensi pelepasan CO2 ke perairan sekitarnya dari kesetimbangan metabolisme organik dan kalsifikasi berkisar antara 3-11,3 mmolC/m2/th. Fluks CO2 udara laut di perairan sekitar terumbu karang sangat tergantung pada waktu tinggal massa air, bentuk geomorfologi terumbu karang (karang tepi, penghalang, atau sistem terumbu karang atol) dan pola arus air laut di perairan sekitarnya. Di samping proses metabolisme yang terjadi dalam sistem terumbu karang dan waktu tinggal dari massa air, fluks CO2 udara-laut juga dimodulasi oleh ΔpCO2 air laut yang masuk.
3 METODE PENELITIAN
3.1 Lokasi dan Waktu Penelitian
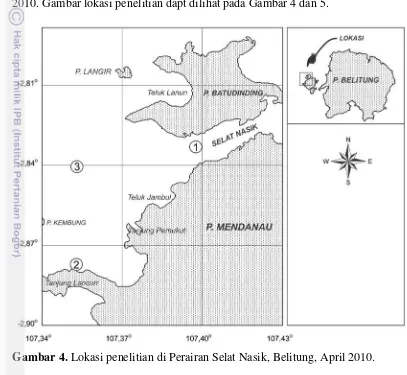
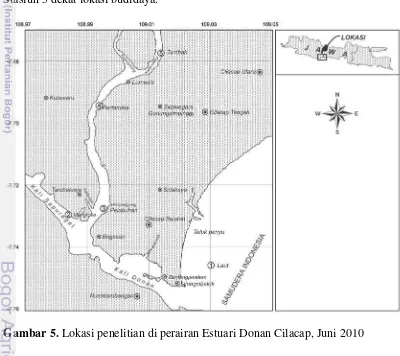
Penelitian telah dilakukan di perairan Selat Nasik Kabupaten Belitung pada bulan April 2010 dan di perairan Estuari Donan Cilacap pada bulan Juni 2010. Gambar lokasi penelitian dapt dilihat pada Gambar 4 dan 5.
Gambar 4. Lokasi penelitian di Perairan Selat Nasik, Belitung, April 2010.
16
fitoplankton dalam menurunkan CO2 dalam air laut, karena di perairan laut fitoplankton merupakan satu-satunya organisme yang menyerap CO2 sedangkan di perairan sekitar mangrove dan terumbu karang ada organisme lain yang menyerap CO2 diantaranya lamun, makroalga, dan zooxanthella sehingga untuk melihat peranan fitoplankton dalam penyerapan CO2
Gambar 5. Lokasi penelitian di perairan Estuari Donan Cilacap, Juni 2010
hanya dilakukan di perairan laut.
3.2 Bahan dan Alat Penelitian
Bahan yang digunakan dalam penelitian meliputi : 1. Bahan kimia untuk analisis karbon, terdiri dari HgCl2
2. Bahan kimia untuk analisis Produktifitas Primer fitoplankton terdiri dari: MnSO
sebagai pengawet atau untuk menghentikan aktifitas biologi, HCl dan NaOH untuk analisis alkalinitas dan DIC.
4, NaI-Azida, H2SO4
3. Bahan kimia untuk analisis fosfat dan silikat: ammonium molibdate, asam sulfat, asam ascorbate, kalium antimonyl tartrat, methol sulfit dan asam oksalat.
dan Natrium Thiosulfat.
Tabel 1. Peralatan yang digunakan dalam penelitian.
No. Nama Alat Kegunaan
1 CTD Pengukuran suhu dan salinitas
2 Nansen water sampler Pengambilan sampel air
3 pH meter Pengukuran pH
4 CO2 meter Pengukuran CO2
5
atmosfir
Light meter Pengukuran intensitas cahaya
6 Cool Box Untuk penyimpanan sampel air
7 Alat saring dan Vacum pump Membantu proses penyaringan sampel
8 Kertas saring CNM 0,45μm Untuk menyaring sampel air
9 Mikro titrator Untuk titrasi
10 Alat-alat gelas
11 Spectrophotometer Pengukuran fosfat dan silikat
3.3 Prosedur penelitian
3.3.1 Sistem CO2
18
3.3.2 Produktifitas primer
Pengambilan sampel air dilakukan dengan menggunakan VanDorn water sampler pada lapisan permukaan (0–1 meter). Pada waktu sampling, sampel air langsung dipindahkan ke plastic container (jerigen) 5–10 liter. Sampel kemudian
dialirkan melalui ‘plankton gauze’ (mesh size 200 μm) ke dalam 2 botol terang
dan 1 botol gelap. Untuk menentukan kandungan O2 pada permulaan inkubasi dilakukan dengan metode titrasi Winkler. Pada saat yang bersamaan, 1 botol terang + 1 botol gelap diinkubasi di bawah sinar matahari selama ±4 jam. Proses inkubasi dilakukan dengan cara menggantungkan botol pada pelampung sehingga botol terendam pada lapisan permukaan. Setelah inkubasi, konsentrasi oksigen yang terdapat dalam botol gelap dan botol terang tersebut diukur kembali dengan metode titrasi Winkler.
3.4 Analisis data
3.4.1 Sistem CO
Sistem CO 2
2 di perairan dapat dikaji melalui empat parameter yang dapat diukur, yaitu DIC, total alkalinitas (TA), pH dan pCO2 (tekanan parsial CO2) (Lewis dan Wallace, 1997). Pada studi ini DIC diukur menggunakan metode “Titrasi” (Giggenbach & Goguel, 1989), dengan prinsip mendasarkan pada
perubahan pH setelah ditambahkan HCl dan NaOH. DIC didapatkan dari penjumlahan HCO3- dan CO3
2-Keterangan:
A dan B = Volume HCl yang digunakan untuk menurunkan pH. C dan D = Volume NaOH yang digunakan untuk menaikkan pH. Vs = Volume sampel air laut yang dianalisa
Hasil pengukuran DIC dengan metode ini kemudian dikoreksi dengan hasil pengukuran certified sample/Certified Refference Material (RCM) dari Marine Physical Laboratory, University of California, San Diego.
dalam satuan μmol/kg.
... (5)
Nilai RCM = 2021±0,65 µmol/kg, sedangkan hasil pengukuran RCM oleh BATAN = 1657,20 µmol/kg, jadi nilai koreksinya adalah 364,45 µmol/kg.
Total Alkalinitas diukur dengan menggunakan metode “Titrasi”
(Grasshoff, 1976). Prosedurnya meliputi: Kedalam 50 ml sampel air laut ditambahkan 5 ml HCl 0,025 M dan dididihkan selama ±5 menit, kemudian didinginkan dalam water bath. Setelah dingin kedalam sampel ditambahkan 3–5 tetes bromothymol blue sebagai indikator, kemudian sampel dititrasi dengan NaOH 0,02 M, selama titrasi kedalam sampel dialirkan gas bebas CO2 (Nitrogen atau Helium). Proses titrasi dihentikan setelah sampel bewarna biru, dan volume NaOH yang terpakai dicatat dan dimasukkan kedalam rumus berikut:
Keterangan:
V = Volume HCl dan NaOH t = Molaritas HCl dan NaOH Vb = Volume sampel
Tekanan parsial CO2 (pCO2) kolom air dihitung dengan menggunakan Model OCMIP (Ocean Carbon Cycle Model Intercomparison Project) yang dikembangkan oleh Orr et al. (1999). Parameter yang digunakan dalam model ini adalah: suhu, salinitas, kisaran pH, pH insitu, DIC, alkalinitas, fosfat dan silikat. pCO2 atmosfir diukur pada waktu pengambilan sampel dengan menggunakan CO2
3.4.2 Pertukaran CO meter.
2
Secara umum fluks atau pertukaran aliran gas CO udara-laut
2 antara udara dan laut dihitung dengan menggunakan rumus berikut:
Fluks CO2 = K. α. Δ pCO2 air–atm ... (8) ΔpCO2 = pCO2 air-pCO2 atm ... (9)
Keterangan:
K = kecepatan transfer gas (fungsi dari kecepatan angin)
α = koefisien daya larut (fungsi dari suhu dan salinitas)
20
ΔpCO2air–atm = perbedaan (selisih) antara tekanan parsial CO2 permukaan air dengan atmosfir
Berdasarkan nilai pCO2 laut dan atmosfir dapat ditentukan apakah suatu perairan penyerap (sink) atau pelepas (source) CO2. Suatu perairan berperan sebagai ‘source’ atau pelepas CO2 ke udara/atmosfir jika nilai pCO2-nya lebih tinggi dari nilai atmosfir (nilai positif) karena akan terjadi aliran CO2dari air ke atmosfir. Sebaliknya berperan sebagai penyerap/‘sink’ CO2dari atmosfir jika nilai pCO2air-nya lebih rendah dari pCO2atm
3.4.3 Produktifitas Primer
(nilai negatif).
Pengukuran produktifitas primer dilakukan dengan menggunakan botol gelap dan botol terang dengan metode winkler modifikasi azida (Strickland & Parsons, 1968; APHA, 1980). Laju fotosintesis dan respirasi dalam satuan mgC/l/jam dihitung dengan menggunakan rumus:
LB (ppmO2) – DB (ppmO2
0,375 = faktor konversi dari oksigen ke karbon LB = Botol terang (Light bottle), Kandungan O
RQ = Respiration Quotient (1) N = Lama inkubasi (4 jam)
4
HASIL DAN PEMBAHASAN
4.1 Sistem CO2 4.1.1 Selat Nasik
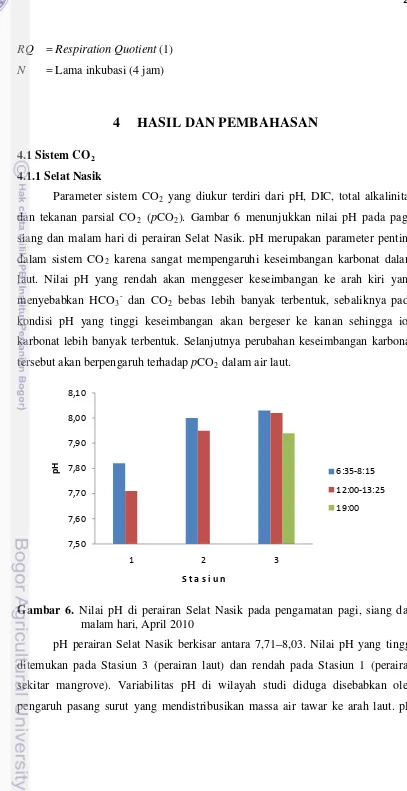
Parameter sistem CO2 yang diukur terdiri dari pH, DIC, total alkalinitas dan tekanan parsial CO2 (pCO2). Gambar 6 menunjukkan nilai pH pada pagi, siang dan malam hari di perairan Selat Nasik. pH merupakan parameter penting dalam sistem CO2 karena sangat mempengaruhi keseimbangan karbonat dalam laut. Nilai pH yang rendah akan menggeser keseimbangan ke arah kiri yang menyebabkan HCO3- dan CO2 bebas lebih banyak terbentuk, sebaliknya pada kondisi pH yang tinggi keseimbangan akan bergeser ke kanan sehingga ion karbonat lebih banyak terbentuk. Selanjutnya perubahan keseimbangan karbonat tersebut akan berpengaruh terhadap pCO2
Gambar 6. Nilai pH di perairan Selat Nasik pada pengamatan pagi, siang dan malam hari, April 2010
dalam air laut.
pH perairan Selat Nasik berkisar antara 7,71–8,03. Nilai pH yang tinggi ditemukan pada Stasiun 3 (perairan laut) dan rendah pada Stasiun 1 (perairan sekitar mangrove). Variabilitas pH di wilayah studi diduga disebabkan oleh pengaruh pasang surut yang mendistribusikan massa air tawar ke arah laut. pH
22
yang tinggi cenderung ditemukan pada saat pasang dan rendah pada saat surut. Hal ini disebabkan oleh masuknya massa air laut dengan pH yang tinggi pada saat pasang sehingga perairan pesisir didominasi oleh massa air laut, sedangkan pada saat surut perairan pesisir didominasi oleh air tawar dengan pH yang rendah. Secara umum pH yang tinggi ditemukan pada pagi hari dan rendah pada siang hari. Khusus di Stasiun 3 pH yang rendah ditemukan pada malam hari. Rendahnya pH pada malam hari disebabkan oleh aktivitas organisme yang melakukan respirasi pada malam hari. Karbon dioksida yang dihasilkan dalam proses respirasi bereaksi dengan air laut menghasilkan H2CO3 yang bersifat asam sehingga pH air laut menjadi turun.
Gambar 7 menunjukkan konsentrasi DIC pada pagi, siang dan malam hari di perairan Selat Nasik. Konsentrasi DIC menggambarkan total konsentrasi CO2
Gambar 7. Konsentrasi DIC di perairan Selat Nasik pada pengamatan pagi, siang dan malam hari, April 2010.
dalam air laut. Konsentrasi DIC di perairan Selat Nasik berkisar antara 1912,15– 2119,03 µmol/kg. Secara umum konsentrasi DIC yang tinggi ditemukan pada Stasiun 1 yang dekat dengan ekosistem mangrove dan yang paling rendah pada Stasiun 2 di perairan sekitar terumbu karang. Secara umum konsentrasi DIC pada pagi hari lebih tinggi dibanding siang hari.
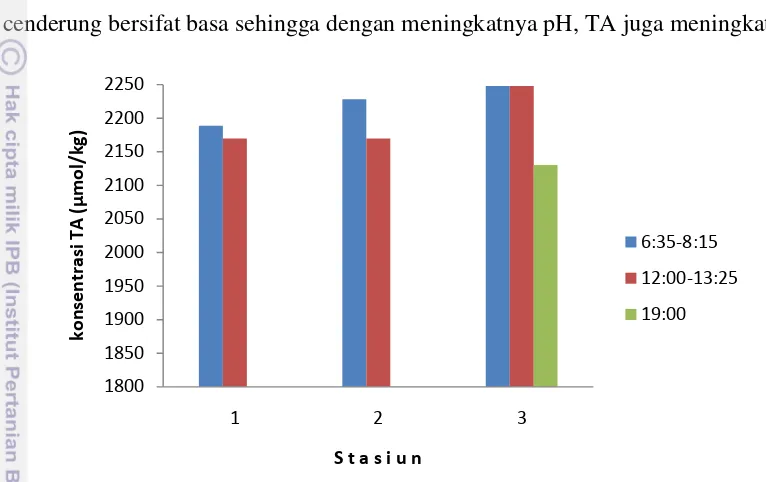
Gambar 8 menunjukkan konsentrasi total alkalinitas (TA) pada pagi, siang dan malam hari di perairan Selat Nasik. TA menggambarkan perbedaan muatan antara kation-kation dan anion-anion konservatif dalam air laut, sehingga TA akan
berubah dengan perubahan ion-ion konservatif yang bervariasi terhadap salinitas. Total alkalinitas di perairan Selat Nasik berkisar antara 2130,28–2247,28 µmol/kg. Sebaran TA di perairan selat Nasik mirip dengan sebaran pH dan salinitas, dimana secara umum total alkalinitas yang tinggi ditemukan pada Stasiun 3 dan rendah pada Stasiun 1. Hal ini disebabkan oleh alkalinitas yang cenderung bersifat basa sehingga dengan meningkatnya pH, TA juga meningkat.
Gambar 8. Konsentrasi total alkalinitas di perairan Selat Nasik pada pengamatan pagi, siang dan malam hari.
Berdasarkan Gambar 7 dan 8 terlihat bahwa pada Stasiun 2 di perairan sekitar terumbu karang, konsentrasi DIC cenderung rendah sedangkan TA relatif lebih tinggi. Hal ini disebabkan oleh faktor-faktor yang mempengaruhi konsentrasi DIC dan TA berbeda meskipun di perairan sekitar terumbu karang kedua parameter tersebut dipengaruhi oleh laju kalsifikasi (TA lebih banyak diserap dibanding DIC). Konsentrasi DIC berubah oleh perubahan CO2 akibat keseimbangan udara dan laut atau akibat fotosintesis dan respirasi, sedangkan TA tidak berubah (Zeebe and Wolf-Gladrow, 2001). Rendahnya konsentrasi DIC di Stasiun 2 diduga karena laju fotosintesis oleh fitoplankton, zooxanthella dan makroalga lebih dominan dibanding laju kalsifikasi.
24
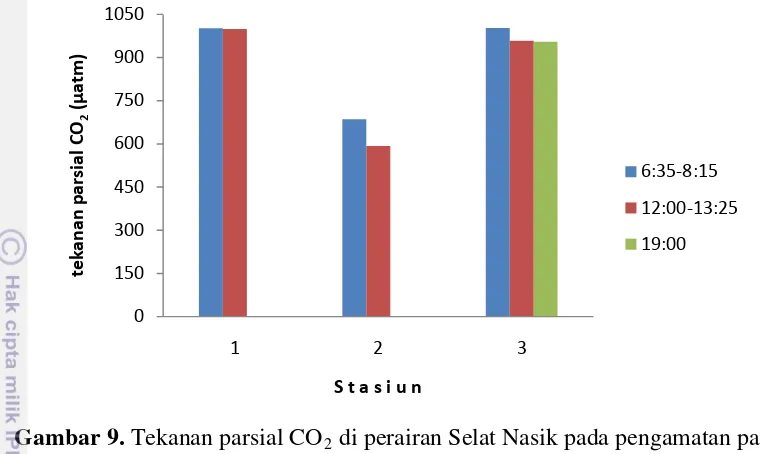
Gambar 9. Tekanan parsial CO2 di perairan Selat Nasik pada pengamatan pagi, siang dan malam hari.
Gambar 9 menunjukkan tekanan parsial CO2 (pCO2) di perairan Selat Nasik pada pengamatan pagi, siang dan malam hari. Perbedaan tekanan parsial CO2 antara kolom air dan atmosfir akan menentukan peranan perairan tersebut sebagai penyerap atau pelepas CO2. Tekanan parsial CO2 kolom air di perairan Selat Nasik berkisar antara 591,80–1002,01 µatm. Nilai tekanan parsial CO2 dalam kolom air ini lebih tinggi dibanding tekanan parsial CO2 di udara yang berkisar antara 307–409 µatm dengan rata-rata 373,38±25,95 µatm. Distribusi tekanan parsial CO2 dalam kolom air mirip dengan sebaran DIC yaitu cenderung lebih tinggi di Stasiun 1 dan rendah di Stasiun 2.
4.1.2 Estuari Donan
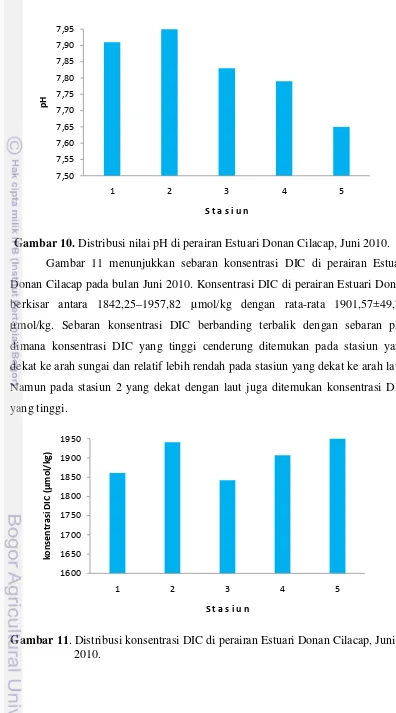
Sebaran parameter sistem CO2 di perairan Estuari Donan Cilacap disajikan dalam bentuk grafik (Gambar 10–13). Gambar 10 menunjukkan sebaran pH di perairan Estuari Donan Cilacap pada bulan Juni 2010. Nilai pH perairan Estuari Donan Cilacap relatif lebih rendah dibanding perairan Selat Nasik yaitu berkisar antara 7,65–7,95 dengan rata-rata 7,83±0,117. pH yang tinggi cenderung ditemukan pada stasiun yang dekat dengan laut atau mulut sungai dan semakin rendah pada stasiun bagian hulu sungai. Rendahnya nilai pH pada stasiun bagian hulu sungai berhubungan dengan rendahnya salinitas (23 psu) karena besarnya massa air tawar yang berasal dari darat.
Gambar 10. Distribusi nilai pH di perairan Estuari Donan Cilacap, Juni 2010.
Gambar 11 menunjukkan sebaran konsentrasi DIC di perairan Estuari Donan Cilacap pada bulan Juni 2010. Konsentrasi DIC di perairan Estuari Donan berkisar antara 1842,25–1957,82 µmol/kg dengan rata-rata 1901,57±49,76 µmol/kg. Sebaran konsentrasi DIC berbanding terbalik dengan sebaran pH, dimana konsentrasi DIC yang tinggi cenderung ditemukan pada stasiun yang dekat ke arah sungai dan relatif lebih rendah pada stasiun yang dekat ke arah laut. Namun pada stasiun 2 yang dekat dengan laut juga ditemukan konsentrasi DIC yang tinggi.
26
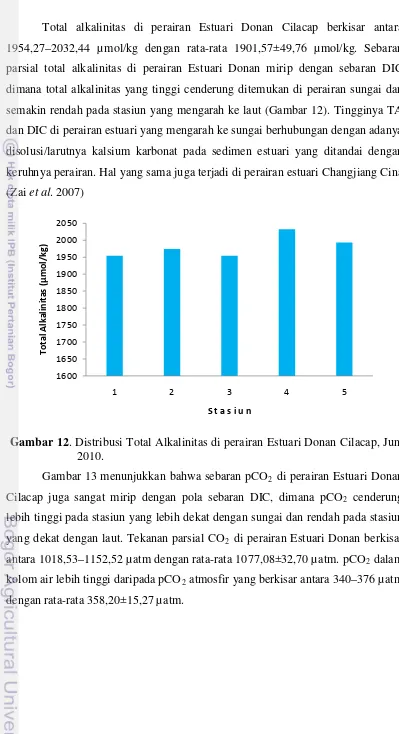
Total alkalinitas di perairan Estuari Donan Cilacap berkisar antara 1954,27–2032,44 µmol/kg dengan rata-rata 1901,57±49,76 µmol/kg. Sebaran parsial total alkalinitas di perairan Estuari Donan mirip dengan sebaran DIC dimana total alkalinitas yang tinggi cenderung ditemukan di perairan sungai dan semakin rendah pada stasiun yang mengarah ke laut (Gambar 12). Tingginya TA dan DIC di perairan estuari yang mengarah ke sungai berhubungan dengan adanya disolusi/larutnya kalsium karbonat pada sedimen estuari yang ditandai dengan keruhnya perairan. Hal yang sama juga terjadi di perairan estuari Changjiang Cina (Zai et al. 2007)
Gambar 12. Distribusi Total Alkalinitas di perairan Estuari Donan Cilacap, Juni 2010.
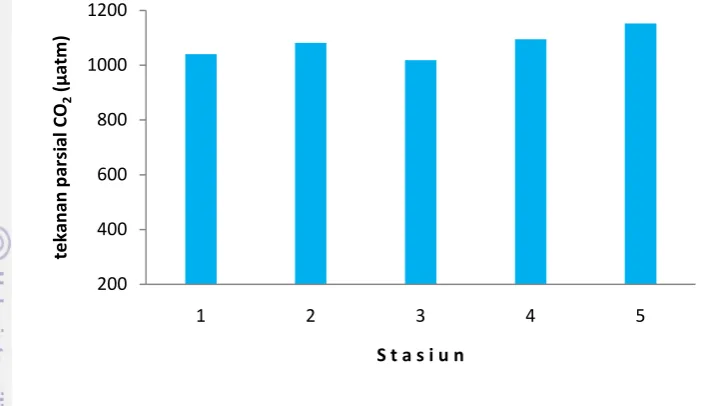
Gambar 13. Distribusi tekanan parsial CO2
4.2Produktifitas Primer
di perairan Estuari Donan Cilacap, Juni 2010.
4.2.1 Selat Nasik
Produktifitas primer perairan Selat Nasik cukup bervariasi antar lokasi dan waktu penelitian. Laju produksi primer kotor/fotosintesis (GPP), respirasi, dan produksi primer bersih (NPP) disajikan dalam Gambar 14. Secara umum aktifitas biologi di perairan Selat Nasik lebih tinggi pada Stasiun 1 (perairan sekitar mangrove). Hal ini ditunjukkan oleh nilai laju fotosintesis, respirasi dan produktifitas primer bersih yang lebih tinggi pada Stasiun 1. Laju fotosintesis dan respirasi di perairan Selat Nasik masing-masing berkisar antara 0,069–0,094 mgC/m2/hari dan 0,082–0,092 mgC/m2/hari. Fitoplankton pada Stasiun 1 cenderung bersifat autotrofik (laju fotosintesis lebih tinggi daripada laju respirasi), sedangkan pada Stasiun 2 dan 3 cenderung bersifat heterotrofik (laju respirasi lebih tinggi dibanding laju fotosintesis). Secara umum laju fotosintesis dan respirasi meningkat pada siang hari.
28
Gambar 14. Laju fotosintesis (GPP), respirasi, dan produksi primer bersih (NPP) di perairan Selat Nasik, April 2010. [A=pagi (6:35–8:15), B=siang (12:00–13:25)]
4.2.2 Estuari Donan
Sebaran laju fotosintesis (GPP), repirasi dan produktifitas primer bersih (NPP) di perairan Estuari Donan disajikan dalam Gambar 15.
Gambar 15. Laju fotosintesis (GPP), respirasi dan produksi primer bersih (NPP) di perairan Estuari Donan, Cilacap, Juni 2010.
Sebaran produktifitas primer di perairan Estuari Donan cukup berfluktuasi. Secara umum perairan Estuari Donan bersifat autotrofik dimana laju fotosintesis lebih tinggi dibanding laju respirasi. Sebaran laju fotosintesis di perairan Estuari Donan berbanding terbalik dengan sebaran DIC dan pCO2. Berdasarkan Gambar
15 terlihat bahwa semakin ke arah laut laju fotosintesis dan respirasi semakin tinggi, namun laju fotosintesis selalu lebih tinggi dibanding laju respirasi kecuali di Stasiun 4. Perairan di Stasiun 4 cenderung bersifat heterotrofik karena laju respirasi lebih tinggi dibanding fotosintesis. Hal ini diduga karena stasiun empat mempunyai suhu yang relatif lebih tinggi (29,6oC). Suhu dapat mempengaruhi fotosintesis di laut baik secara langsung maupun tidak langsung. Pengaruh suhu secara langsung yakni untuk mengontrol reaksi kimia enzimatik dalam proses fotosintesis, sedangkan pengaruh secara tidak langsung yakni dalam merubah struktur hidrologi kolom perairan yang dapat mempengaruhi distribusi fitoplankton (Tomascik et al. 1997). Suhu yang tinggi dapat menaikkan laju maksimum fotosintesis (Pmax). Secara umum, laju fotosintesis fitoplankton meningkat dengan meningkatnya suhu perairan, namun laju respirasi meningkat lebih cepat dibanding fotosintesis pada suhu tinggi. Laju fotosintesis dan respirasi di perairan Estuari Donan masing-masing berkisar antara 0,022–0,070 mgC/m2/hari dan 0,015–0,045 mgC/m2/hari.
4.3 Pertukaran CO2 udara-laut 4.3.1 Selat Nasik
30
Gambar 16. Fluks CO2 di perairan Selat Nasik, April 2010.
Emisi CO2 di perairan Selat Nasik berkisar antara 0,96 mmolC/m2/hari atau 0,35 molC/m2/th sampai 3,19 mmolC/m2/hari atau 1,17 molC/m2/th. Secara umum emisi CO2 yang tinggi ditemukan pada Stasiun 1 di sekitar perairan mangrove dan rendah pada Stasiun 2 di sekitar perairan terumbu karang. Tingginya fluks CO2 di perairan sekitar mangrove disebabkan oleh tingginya konsentrasi DIC dan tekanan parsial CO2 dalam kolom perairan dan rendahnya pH.
4.3.2 Estuari Donan
Gambar 17 menunjukkan bahwa perairan Estuari Donan pada pengamatan Juni 2010 juga berperan sebagai pelepas (source) CO2 ke atmosfir yang ditunjukkan oleh fluks CO2 positif. Fluks CO2 di perairan Estuari Donan tidak begitu berfluktuasi, namun cenderung lebih tinggi pada Stasiun 2 dan Stasiun 5. Stasiun 2 adalah stasiun yang berada dekat ekosistem mangrove, sedangkan stasiun 5 berada pada bagian paling hulu dari lokasi penelitian.
Gambar 17. Fluks CO2 di perairan Estuari Donan Cilacap, Juni 2010.
Fluks CO2 perairan Estuari Donan berkisar antara 6,76–7,72 mmolC/m2/hari atau 2,47–2,72 molC/m2/th dengan rata-rata 7,14±0,43 mmolC/m2/hari atau 2,54±0,11 molC/m2/th. Nilai fluks CO2 di perairan Estuari Donan ini relatif lebih tinggi dibanding perairan Selat Nasik.
4.4 Pembahasan
4.4.1 Selat Nasik
Hasil di atas menunjukkan bahwa secara umum perairan Selat Nasik (perairan sekitar mangrove, terumbu karang dan laut) adalah source CO2 ke atmosfir. Hasil ini menegaskan penelitian sebelumnya yang menyatakan bahwa perairan laut dan pesisir di daerah tropis adalah source CO2 ke atmosfir (Cai et al., 2003; Wang and Cai, 2004; Borges, 2005; Fagan and Mackenzie, 2007; Chen and Borges, 2009). Fluks CO2 yang tinggi di perairan Selat Nasik ditemukan pada Stasiun 1 yang berada di sekitar ekosistem mangrove yang disebabkan oleh tingginya tekanan parsial CO2 dalam kolom perairan. Tingginya tekanan parsial CO2 dalam kolom air di perairan sekitar mangrove, dipengaruhi oleh tingginya konsentrasi DIC dan rendahnya pH dan salinitas. Tingginya konsentrasi DIC diperairan sekitar mangrove diduga disebabkan oleh pasokan karbon organik dan anorganik baik yang berasal dari ekosisitem mangrove maupun yang berasal dari luar ekosistem mangrove yang dibawa oleh arus pasang surut.
32
Gambar 18 menunjukkan siklus karbon pada Stasiun 1 (perairan sekitar mangrove) yang ditampilkan secara sederhana dari kondisi sesungguhnya yang rumit. Pembahasan mengenai siklus karbon di perairan ekosistem mangrove ini dibatasi hanya pada lapisan permukaan, karena pertukaran CO2 udara-laut terjadi pada lapisan permukaan. Kajian siklus karbon dalam penelitian ini lebih ditekankan kepada peranan fitoplankton dalam penyerapan CO2. Fitoplankton mengambil nutrien dan CO2 melalui proses fotosintesis, laju dimana proses ini terjadi disebut produktivitas primer. Fitoplankton pada waktu yang sama juga melakukan respirasi yang meningkatkan konsentrasi CO2 dalam kolom air. Peningkatan konsentrasi CO2 dalam kolom air juga disebabkan oleh adanya proses dekomposisi (diremineralisasi) karbon organik menjadi CO2
Gambar 18. Skema siklus karbon pada Stasiun 1 (perairan sekitar mangrove) pada pengamatan pagi (A) dan siang hari (B). FCO2, GPP, NPP, Respirasi dan Dekomposisi dalam satuan mmolC/m2
Siklus karbon di perairan sekitar mangrove menunjukkan bahwa dekomposisi karbon organik diperkirakan memberikan sumbangan paling besar terhadap fluks CO
/hari.
2 ke atmosfir yaitu 3,121 mmolC/m2/hari pada pagi hari dan 2,988 mmolC/m2/hari pada siang hari. Kemampuan fitoplankton dalam menyerap CO2 sangat kecil yaitu 0,02 mmolC/m2/hari pada pagi hari dan 0,015 mmolC/m2/hari pada siang hari. Hal ini hampir sama dengan yang dikemukakan oleh Gattuso (1998) bahwa fluks CO2 udara-laut yang heterotrofik bersih di perairan sekitar mangrove di dorong oleh tingginya pasokan sedimen yang sebagian besarnya adalah bahan organik yang berasal dari ekosistem mangrove, sedangkan produksi primer biasanya rendah tergantung kepada geomorfologi, waktu tinggal massa air, kekeruhan, dan pasokan nutrient. Menurut Bouillon and CO2 atm
DIC Fitoplankton POC
CO2 atm
DIC Fitoplankton POC
Boskher (2006), masukan karbon organik ke perairan sekitar mangrove bisa bersifat autochtonous (berasal dari perairan itu sendiri) atau allochtonous (berasal dari luar). Masukan karbon organik yang autotochtonous berupa detritus mangrove dan mikrofitobentos, sedangkan yang allochtonous berupa fitoplankton, material yang berasal dari ekosistem lamun, dan karbon organik yang berasal dari daratan. Karbon organik yang masuk ke perairan sekitar mangrove kemudian mengalami dekomposisi dan remineralisasi menjadi karbon anorganik. Selanjutnya Borges et al. (2003) menyatakan bahwa pengayaan DIC di perairan sekitar mangrove juga disebabkan oleh masukan air poros yang kaya CO2 pada saat surut. Pasang surut memompa air poros yang ada dalam sedimen yang kaya akan karbon organik (Bouillon et al. 2008). Jennerjahn dan Ittekkot (2002) memperkirakan bahwa ekosistem mangrove menyumbang >10% dari total karbon organik sungai yang masuk ke laut.
Berdasarkan nilai fotosintesis dan respirasi menunjukkan bahwa fitoplankton di perairan sekitar mangrove cenderung bersifat autotrofik karena laju fotosintesis lebih tinggi dibandingkan laju respirasi, namun tidak berpengaruh signifikan terhadap penurunan emisi CO2
Nilai
ke atmosfir. Hal ini disebabkan oleh laju dekomposisi material organik baik yang berasal dari fitoplankton sendiri ataupun dari sumber autochtonous dan allochtonous, lebih dominan dibanding laju produktifitas primer fitoplankton.
sifat-34
sifat kimia dalam mendegradasi bahan organik dan ditunjang oleh volume air yang lebih kecil dan waktu tinggal yang lebih lama.
Fluks dan tekanan parsial CO2 yang paling rendah ditemukan pada Stasiun 2 yaitu di perairan sekitar terumbu karang. Rendahnya fluks CO2 pada Stasiun 2 disebabkan oleh rendahnya konsentrasi DIC. Konsentrasi DIC di perairan sekitar terumbu karang secara signifikan lebih rendah dibandingkan dengan perairan sekitar mangrove dan laut baik pada pagi maupun siang hari. Hal ini disebabkan oleh proses metabolisme terumbu yang menyebabkan pergeseran dalam sistem karbon anorganik yang menyebabkan perubahan besar dan arah gradien pCO2
Gambar 19 menunjukkan siklus karbon di perairan sekitar terumbu karang. Siklus karbon di perairan sekitar terumbu karang relatif lebih kompleks dibandingkan perairan sekitar mangrove dan laut, karena banyaknya organisme yang terlibat dengan siklus karbon, diantaranya proses fotosintesis oleh fitoplankton, zooxanthella dan makroalga, proses respirasi dan kalsifikasi oleh terumbu karang dan dekomposisi oleh bakteri, namun dalam Gambar 19 hanya dibatasi pada lapisan permukaan yang hanya melibatkan fitoplankton dalam penyerapan CO
antara air laut dan atmosfer.
2
Gambar 19. Skema siklus karbon pada Stasiun 2 (perairan sekitar terumbu karang) pada pengamatan pagi (A) dan siang hari (B). FCO2, GPP, NPP, Respirasi dan Dekomposisi dalam satuan mmolC/m2
Berdasarkan gambar di atas terlihat bahwa penyerapan CO /hari.
2 oleh fitoplankton relatif kecil yaitu 0,009 mmolC/m2/hari pada pagi hari dan 0,010 mmolC/m2/hari pada siang hari. Fitoplankton di perairan sekitar terumbu karang CO2 atm
DIC Fitoplankton POC
CO2 atm
DIC Fitoplankton POC
cenderung bersifat heterotrofik karena laju respirasi lebih tinggi dibanding fotosintesis. Tingginya laju respirasi fitoplankton di perairan sekitar terumbu karang seharusnya meningkatkan fluks CO2 ke atmosfir, karena proses respirasi akan menyumbang CO2 kedalam kolom perairan dan meningkatkan tekanan parsial CO2. Namun hal ini tidak terlihat di perairan sekitar terumbu karang Selat Nasik karena fluks CO2 di perairan sekitar terumbu karang lebih rendah dibanding perairan sekitar mangrove dan laut dan fluks CO2 di perairan sekitar terumbu karang Selat Nasik lebih rendah dibanding emisi CO2 global dari sistem terumbu karang yang diperkirakan oleh Borges (2005) yaitu sekitar 2 mmolC/m2/hari.
Rendahnya fluks CO2 di perairan sekitar terumbu karang Selat Nasik diduga disebabkan oleh tingginya konsumsi CO2 oleh makroalga. Seperti yang dikemukakan oleh Gattuso et al., (1996) bahwa terumbu tepi di bawah pengaruh tekanan manusia telah bergeser dari dominasi karang ke dominasi makroalga, seperti yang terjadi di terumbu Shiraho, pulau Ryukyu. Hal ini akan menyebabkan peningkatan produksi ekosistem bersih dan penurunan kalsifikasi dan memungkinkan pergeseran peran ekosistem terumbu karang dari source ke sink untuk CO2
Fluks CO atmosfir.
36
(1992) menunjukkan bahwa proses kalsifikasi berpengaruh terhadap sistem karbon anorganik air laut dan menyebabkan terumbu karang adalah sedikit source CO2 ke atmosfir.
Gambar 20. Skema siklus karbon pada Stasiun 3 (perairan laut) pada pengamatan pagi (A) dan siang hari (B). FCO2, GPP, NPP, Respirasi dan Dekomposisi dalam satuan mmolC/m2/hari.
Gambar 20 menunjukkan bahwa di perairan laut Selat Nasik laju dekomposisi memberikan sumbangan paling besar terhadap fluks CO2 yaitu 2,973 mmolC/m2/hari pada pagi hari dan 2,802 mmolC/m2/hari pada siang hari. Fitoplankton merupakan satu-satunya organisme yang menyerap CO2 di perairan laut, namun penyerapan CO2 oleh fitoplankton di perairan laut Selat Nasik sangat kecil yaitu 0,0001 mmolC/m2/hari pada pagi hari dan meningkat pada siang hari menjadi 0,0003 mmolC/m2/hari sehingga produktivitas primer memberikan kontribusi yang kecil terhadap pengurangan fluks CO2 ke atmosfer. Penyerapan dan pelepasan CO2 di perairan laut Selat Nasik diduga lebih dipengaruhi oleh konsentrasi DIC dan faktor fisis (suhu permukaan laut). Seperti yang dikemukakan oleh Wang et al. (2006) bahwa konsentrasi DIC dan suhu permukaan laut memainkan peranan utama dalam mengontrol fluks CO2
Berdasarkan hasil di atas menunjukkan bahwa perairan Estuari Donan adalah source atau pelepas CO
di perairan laut tropis.
4.4.2 Estuari Donan
2 ke atmosfir. Secara umum fluks CO2 di perairan Estuari Donan lebih tinggi dibanding perairan Selat Nasik. Tingginya fluks CO2 di perairan Estuari Donan disebabkan oleh tingginya kecepatan angin pada lapisan permukaan (2,50 m/s) dan tingginya tekanan parsial CO2 dalam kolom air. Source CO2 atm
DIC Fitoplankton POC
CO2 atm
DIC Fitoplankton POC
atau pelepasan CO2ke atmosfir ini disebabkan oleh tekanan parsial CO2 (pCO2) dalam kolom air lebih tinggi dibandingkan tekanan parsial CO2 di atmosfir sehingga terjadi aliran gas CO2dari air laut ke atmosfir. Tingginya pCO2 kolom air berhubungan dengan konsentrasi DIC, salinitas dan laju fotosintesis fitoplankton. Tekanan parsial CO2 meningkat dengan meningkatnya konsentrasi DIC dan menurun dengan meningkatnya salinitas dan laju fotosintesis. Sebaran pCO2 di Estuari Donan sangat mirip dengan sebaran DIC, pCO2 dan konsentrasi DIC yang tinggi cenderung ditemukan pada stasiun yang dekat ke arah sungai dan relatif lebih rendah pada stasiun yang dekat ke arah laut. Hal yang serupa juga terjadi di perairan estuari Changjiang Cina (Chen et al., 2008), Godavari India (Bouillon et al., 2003) dan Chilka India (Gupta et al., 2008).Namun pada stasiun 2 yang dekat dengan laut juga ditemukan konsentrasi DIC yang tinggi. Hal ini disebabkan karena stasiun 2 berada pada lokasi yang berada dekat dengan ekosistem mangrove. Tingginya konsentrasi DIC pada lokasi ini diduga karena adanya pasokan karbon organik dan anorganik dari sungai dan ekosistem mangrove. Seperti yang dikemukakan oleh Cai and Wang (1998) bahwa perairan estuari menerima pasokan DIC yang berasal dari eksternal dan internal estuari. Pasokan eksternal berasal dari air tawar dari sungai, air laut dari pantai, tidal-flush, dan perairan sekitarnya. Sedangkan pasokan internal berasal dari degradasi material organik (repirasi aerobik dan fotodegradasi).
38
Gambar 21. Skema siklus karbon di perairan Estuari Donan. FCO2, GPP, NPP, Respirasi dan Dekomposisi dalam satuan mmolC/m2/hari.
Gambar 21 menunjukkan bahwa laju dekomposisi memberikan sumbangan paling besar terhadap fluks CO2 di perairan Estuari Donan yaitu 7,14 mmolC/m2/hari. Laju dekomposisi material organik di perairan Estuari Donan jauh lebih tinggi dibanding perairan Selat Nasik, demikian juga dengan laju produktivitas primer fitoplankton. Tingginya laju dekomposisi di perairan Estuari Donan diduga disebabkan oleh besarnya pasokan material organik dari sungai. Gattuso et al. (1998) mengemukakan bahwa tingginya beban nutrien yang masuk ke perairan estuari menyebabkan terjadinya peningkatan produksi ekosistem bersih, sedangkan degradasi karbon organik antropogenik menurunkan produksi ekosistem bersih. Secara keseluruhan, ekosistem estuari adalah sumber bahan organik, nutrien dan karbon anorganik untuk perairan sekitarnya dan berpotensi sebagai sumber CO2 ke atmosfir (Borges 2005).
Sebaran laju fotosintesis di perairan Estuari Donan menunjukkan bahwa laju fotosintesis cenderung meningkat ke arah mulut sungai (laut). Hal ini berbanding terbalik dengan sebaran DIC dan pCO2. Meningkatnya laju fotosintesis ke arah laut disebabkan karena jumlah material organik terestrial yang tersedia untuk dekomposisi telah berkurang, dan kondisi cahaya yang lebih baik setelah terjadi pengendapan partikel tersuspensi. Meningkatnya aktifitas biologis pada stasiun yang lebih dekat ke arah laut menyebabkan pCO2 lebih rendah. Selanjutnya, karena ada pencampuran dengan air laut yang lebih rendah pCO2, maka pCO2 air permukaan dengan cepat turun ke dekat titik jenuh. Lebih jauh, air CO2 atm
permukaan dapat menjadi net autotrofik jika CO2 lebih banyak dikonsumsi oleh produksi biologis (Chen et al., 2008).
40
5 KESIMPULAN DAN SARAN
Berdasarkan hasil di atas dapat disimpulkan bahwa perairan Selat Nasik, Belitung dan Estuari Donan, Cilacap mempunyai tingkat kejenuhan yang tinggi terhadap CO2 sehingga berperan sebagai pelepas (source) CO2 ke atmosfir. Perairan estuari menyumbang emisi CO2 paling besar diikuti oleh perairan sekitar ekosistem mangrove, laut pesisir (non mangrove dan terumbu karang) dan perairan sekitar terumbu karang. Dalam studi ini penyerapan CO2 oleh fitoplankton melalui proses fotosintesis tidak berpengaruh signifikan terhadap fluks CO2. Dalam kasus ini, laju dekomposisi karbon organik partikel cenderung memberikan pengaruh yang signifikan terhadap fluks CO2.
DAFTAR PUSTAKA
APHA 1980. Standard Methods for The Examination of Water and Wastewater. APHA, AWWA, WPCF. 15 th eds.: 388–389.
Bates, N.M., 2002. Seasonal variability of the effect of coral reefs on seawtare CO2 and air-sea CO2 exchange. Limnol Oceanogr, 47:43–52.
Borges, A.V., J.P. Vanderborght, L.S. Sciettecatte, F. Gazeau, S. Ferron-Smith, B. Dellie and M. Frankignoulle. 2004. Variability of the gas transfer velocity of CO2 in macrotidal estuary (the Scheldt). Estuaries, 27:593– 603.
Borges, A.V., 2005. Do we have enough pieces of the jigsaw to integrate CO2 fluxes in the coastal ocean? Estuaries, 28 (1):3–27.
Borges, A. V., B. Delille and M. Frankignoulle, 2005. Budgeting sinks and sources of CO2 in the coastal ocean: Diversity of ecosystems counts. Geophys Res Lett 32:L14601. doi:10.1029/2005GL023053
Borges, A.V., S. Djenidi, G. Lacroix, J. Theate, B. Delille, and M. Frankignoulle, 2003. Atmospheric CO2 flux from mangrove surrounding waters. Geophys Res Lett, 30(11):1558. doi:10.1029/2003GL017143
Bouillon, S., A.V. Borges, E. Castaneda-Moya, K. Diele, T. Dittmar, N.C. Duke, E. Kristensen, S.Y. Lee, C. Marchand, J.J. Middlburg, V.H. Rivera-Monroy, T.J. Smith III, and R.R. Twilley, 2008. Mangrove production and carbon sink: A revision of global budget estimates. Glob Biogeochem Cyc, 22: GB2013, doi:10.1029/2007GB003052.
Bouillon, S., F. Dehairs, B. Velimirov, G. Abril, and A.V. Borges. 2007. Dynamics of organic and inorganic carbon across contiguous mangrove and seagrass systems (Gazi Bay, Kenya). J Geophys Res, (112): G02018, doi:10.1029/2006JG000325
Bouillon, S. and H.T.S. Boschker, 2006. Bacterial carbon sources in coastal sediments: a cross-system analysis based on stable isotope data of biomarkers. Biogeosciences, 3:175–185.
Bouillon, S., M. Frankignoulle, F. Dehairs, B. Velimirov, A. Eiler, G. Abril, H. Etcheber and A.V. Borges, 2003. Inorganic and organic carbon biogeochemistry in the Gautami Godavari estuary (Andhra Pradesh, India) during pre-monsoon: the local impact of extensive mangrove forest. Glob Biogeochem Cyc, 17(4):1114.
Caffrey, J.M. 2004. Factors controlling net ecosystem metabolism in U.S. estuaries. Estuaries, 27:90–101.
42
Cai, W.J., Z.A. Wang, and Y. Wang, 2003. The role of marsh-dominated heterotrophic continental margins in transport of CO2 between the atmosphere, the land–sea interface and the ocean. Geophys Res Lett, 30:1849.
Cameron, W.M. and D.W. Pritchard, 1963. Estuaries, P. 306 – 324. In M. N. Hill (eds). The sea, Volume 2. John Wiley & Sons. New York.
Chen, C.T.A., W. Zhai, and M. Dai, 2008. Riverine input and air-sea CO2 exchanges near the Changjiang (Yangtze River) estuary: Status quo and implication on possible future changes in metabolic status. Continent Shelf Res, 28:1476–1482.
Chen, C.T.A. and A. V. Borges, 2009. Reconciling views on carbon cycling in the coastal ocean: Continental shleves as sinks and near-shore ecosystem as sources of atmospheric CO2. Deep-Sea Res II, 56:578–590.
Chen C.T.A., K.K. Liu and R. Macdonald, 2003. Continental margin exchanges, p. 53 – 97. In M.J.R. Fasham (ed.). Ocean biogeochemistry: A synthesis of the Joint Global Ocean Flux Study (JGOFS). Springer-Verlag. Berlin. Germany.
Chen, C.T.A., 2004. Exchange of carbon in the coastal seas. In: Field, C.B., M.R. Raupach (Eds.), The Global Carbon Cycle: Integrating Human, Climate and the Natural World. SCOPE, Washington, DC, pp. 341–351.
Dickson, A.G., C.L. Sabine, and J.R. Christian, 2007. Guide to best practices for ocean CO2 measurements. Pices special publication 3. IOCCP Report 8: 176.
Fagan, K.E and F.T. Mackenzie, 2007. Air-sea CO2 exchange in a subtropical estuarine-coral reef system, Kaneohe Bay, Oahu, Hawaii. Mar Chem, 106:174–191.
Falkowski, P.G., R.T. Barber and V. Smetacek, 1998. Biogeochemical controls and feedbacks on ocean primary production. Science, 281:200-206. Fasham, M.J.R., B.M. Balin, C. Bowles (Eds), 2001. A new vision of ocean
biogeochemistry after a decade of the Joint Global Ocean Flux Study (JGOFS). Ambio Special Report:4–31.
Feely R.A., C.L. Sabine, T. Takahashi, and R. Wanninkhof, 2001. Uptake and storage of carbon dioxide in the ocean. Oceanography. Vol. 4: 17 – 32. Frankignoulle, M., C. Canon, and J.P. Gattuso, 1994. Marine calcification as a
source of carbon dioxide: Positive feedback to increasing atmospheric CO2. Limnol Oceanogr, 39:458–462.
Gattuso, J.P., M. Frankignoulle and S.V. Smith, 1999. Measurement of community metabolism and significance in the coral reef CO2 source-sink debate. PNAS 96(23):1317–1322.