KERAGAMAN GENETIK CYTOCHROME B
PADA BURUNG MAMBRUK (Goura sp.)
Oleh:
Lasriama Siahaan G04400032
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
ABSTRAK
LASRIAMA SIAHAAN. Keragaman Genetik Cytochrome b pada Burung Mambruk (Goura sp.). Dibimbing oleh DEDY DURYADI SOLIHIN dan DJOKO WALUYO.
Burung Mambruk (Goura sp.) merupakan satwa endemik Indonesia dengan status “Vulnerable” yang tersebar di daerah Pulau Irian Jaya dan beberapa daerah Papua New Guinea. Usaha konservasi terhadap spesies ini akan berhasil jika karakteristik morfologi, keragaman molekuler dan genetik dapat diketahui dengan pasti. Tujuan penelitian ini untuk menganalisis keragaman genetik cytochrome b parsial dengan metode Polimerase Chain Reaction. Hasil perunutan dari amplifikasi pasangan primer M101 dan M102 pada cytochrome b parsial sepanjang 382 nukleotida (menyandikan 127 asam amino) disejajarkan (multiple aligment) dengan bantuan perangkat lunak Genetyx-Win versi 3.0 dan Clustal-X, selanjutnya dianalisis dengan program MEGA versi 3.0.
Hasil analisis dari 382 nukleotida yang dibandingkan terdapat 42 situs nukleotida yang beragam dengan rata-rata kejadian substitusi transisi 0.05 dan rata-rata substitusi transversi 0.01. Perubahan bersifat non-sinonimus 7.08% (10 situs asam amino) dan perubahan bersifat sebesar 23.62% (30 situs asam amino). Jarak genetik nukleotida cytochrome b (metode p-distance) didapat bahwa nilai paling kecil adalah 0.26% dan nilai yang paling besar 7.07% dengan rata-rata sebesar 5.26%. Hasil rekonstruksi filogenetik dengan metode Neighbor Joining menunjukkan bahwa G.cristata lebih berkerabat dekat dengan G.scheepmakeri daripada G.victoria.
Kata kunci: Goura sp., Cytochrome b, substitusi, perubahan
ABSTRACT
LASRIAMA SIAHAAN. Genetic Diversity of Cytochrome b of Crowned-pigeons (Goura sp.), Supervised by DEDY DURYADI SOLIHIN and DJOKO WALUYO.
Crowned-pigeon (Goura sp.) was one of endemic animal in Indonesia with “vulnerable" status which spread at Papua New Guinea and Irian Jaya Island. Conservation effort to this species will succeed if morphology characteristic and genetic diversity be known. The main purpose of this research is to analyze nucleotide variability of cytochrome b partial using Polymerase Chain Reaction method. The PCR product amplified by primer M101 and M102, then multiple aligment using Genetyx-Win 3.0™ and Clustal-X™, furthermore analyzed with MEGA 3.0™.
The result of analyse from 382 nucleotide of cytochrome b partial encoding 127 amino acid. Findings 42 sites of nucleotide variable with average of transitional substitutions 0.05 and average transversions substitutions is 0.01. Value non-synonymous mutations is 7.08% (10 amino acid sites) whereas synonymus mutations is 23.62% (30 amino acid sites). Value of genetic distance of nucleotide cytochrome b (method p-distance) range from 0.26% - 7.07% with average 5.26%. Filogenetic reconstruction using Neighbor Joining method indicate that G.cristata were closer to G.scheepmakeri compare with G.victoria.
mitokondria tidak hanya ditemukan pada kelompok burung, hal sama yang terjadi pada kelompok Rodensia dan Ikan (Kocher et al. 1989). Komposisi nukleotida-2 dari triplet kodon paling tidak beragam karena komposisi A (17.3%) sama, demikian pula dengan G (14.2%) untuk Goura sp. dalam penelitian ini.
Basa dari triplet kodon yang paling banyak mengalami perubahan substitusi adalah basa ke-3 sebesar 22.04% (28 situs), hal ini sesuai dengan pendapat Sorenson (2003) dimana kejadian substitusi pada basa ke-3 dari triplet kodon lebih banyak terjadi.
Perubahan penerjemahan asam amino terjadi karena adanya substitusi nukleotida transisi (perubahan antar basa purin atau antar basa pirimidin) dan substitusi transversi (perubahan dari basa purin menjadi basa pirimidin atau sebaliknya). Transisi pada basa ke-3 tidak menyebabkan perubahan asam amino dan transversi basa ke-3 tidak selalu menyebabkan perubahan asam amino.
Perubahan basa triplet kodon yang menyebabkan terjadinya perubahan asam amino non-sinonim adalah perubahan basa ke-1 sebesar 3.15% (4 situs asam amino), basa ke-2 sebesar 1.57% (2 situs asam amino), basa ke-3 sebesar 1.57% (2 situs asam amino), basa ke-1 dan 3 sebesar 1.57% (2 situs asam amino). Sedangkan perubahan asam amino sinonim terdapat pada perubahan basa ke-3 sebesar 20.47% (26 situs asam amino), basa ke-1 sebesar 3.15% (4 situs asam amino). Perubahan basa ke-1 umumnya menyebabkan perubahan asam amino. Kecuali pada asam amino Leu (leusin), walaupun transisi dan transversi terjadi pada basa ke-1 dan ke-3 tidak akan menyebabkan perubahan translasi asam aminonya.
Dari 127 asam amino yang diamati terdapat 68.50% (87 situs) merupakan asam amino kekal (nukleotidanya tidak mengalami substitusi), 7.87% (10 situs) perubahan bersifat non-sinonim (nukleotida mengalami substitusi dan asam amino berubah) dan 23.62% (30 situs) perubahan bersifat sinonim (nukleotida mengalami substitusi dan asam amino tetap).
Walaupun telah terjadi perubahan pada 42 situs nukleotida namun hanya 10 situs nukleotida beragam yang dapat mengubah asam amino (10 asam amino). Sepuluh situs asam amino yang berubah terjadi secara transisi dan transversi basa ke-1 (4 situs asam amino yaitu L (leusina) menjadi F (fenilalanina), M (metionina) menjadi L (leusina), T (treonina)
menjadi A (alanina) dan F (fenilalanina) menjadi L (leusina); transisi basa ke-2 (2 situs asam amino yaitu F (fenilalanina) menjadi S (serina) dan M (metionina) menjadi T (treonina); transversi basa ke-3 (2 situs asam amino yaitu C (sisteina) menjadi W (triptofan); serta perpaduan transisi basa ke-1 dan transversi basa ke-3 (2 situs asam amino yaitu S (serina) menjadi P (prolina) dan L (leusina) menjadi F (fenilalanina). Sedangkan situs nukleotida yang mengalami substitusi lebih banyak (30 situs asam amino) terjadi secara sinonim bukan non-sinonim sehingga tidak mengubah asam aminonya.
Kelompok burung (spesies, genera dan famili) memiliki rata-rata jarak genetik yang lebih kecil daripada jarak takson yang sama antara mamalia, amphibi dan reptil. Secara umum kelompok burung memiliki jarak genetik intraspesies yang sangat kecil (Stanley & Harrison 1999). Jarak genetik (p-distance) berdasarkan jumlah nukleotida yang berbeda, memperlihatkan jarak genetik intraspesies
Goura sp. yang paling rendah adalah 0.26% (G. scheepmakeri dengan G. cristata) dan paling tinggi sebesar 7.07% (G. victoria GenBank
dengan G. victoria hasil penelitian) dengan rata-rata sebesar 5.26%. Rata-rata-rata jarak genetik antar spesies yang dibandingkan adalah 11.54% dan jarak genetik rata-rata famili Columbidae (data dari GenBank) sebesar 9.79%. Jarak genetik ini mendukung hasil penelitian Johnson & Clayton (2000) yaitu didapatkan bahwa perbedaan nukleotida Cyt b antar spesies Columbiformes sekitar 0.97-17.07%.
Hasil ini juga mengasumsikan kekerabatan yang dekat di antara Goura sp. bahkan antar spesies Columbidae. Kekerabatan genetik antara G. cristata dengan G. scheepmakeri
(dengan nilai bootstrap 80%) lebih dekat dengan morfologi yang lebih mirip jika dibandingkan dengan G. victoria. Burung Mambruk dari jenis G. scheepmakeri berada di antara G. cristata dan G. victoria.
KATA PENGANTAR
Persembahan syukur kepada Allah Bapa atas anugerahNya, sehingga penulis
dapat menyelesaikan karya ilmiah ini. Tema yang dipilih dalam penelitian yang
dilaksanakan sejak bulan September 2004 ini adalah ”Keragaman Genetik
Cytochrome
b
pada Burung Mambruk (
Goura
sp.).
Terima kasih penulis ucapkan kepada Bapak Dr. Ir. Dedy Duryadi Solihin, DEA
dan Bapak drh. Djoko Waluyo, M.S selaku pembimbing dan penyandang dana dalam
penelitian ini. Disamping itu, penghargaan penulis sampaikan kepada Taman Burung
Taman Mini Indonesia Indah (TMII) yang telah membantu dalam hal penyediaan contoh
darah burung Mambruk. Tak lupa penulis sampaikan kepada Bapak Heri yang telah
membantu dalam masalah tehnik laboratorium serta rekan-rekan kerja di Laboratorium
Biologi Molekuler Pusat Studi Ilmu Hayati IPB, Ibu Rini, K’ Chule, K’Evi, Virgo atas
saran, dukungan dan kerjasama yang terjalin selama bekerja di Laboratorium.
Terimakasih juga penulis ucapkan kepada saudara terkasih Ike, T’ Lucien, K’ Regina, K’
Anna, B’ Andrew, Pemuda GKI Pengadilan Bogor atas segala bentuk dorongan dan
perhatiannya, Lies dan Biologi angkatan 37’ atas kebersamaannya. Segala cinta dan
terima kasih penulis ungkapkan kepada Bapak, Oma, Nardus, Herman, Okki, Mindo atas
doa, dukungan moral, material, kesabaran dan kasih sayangnya.
Berharap karya ilmiah ini bermanfaat.
Bogor, Maret 2006
PADA BURUNG MAMBRUK (
Goura
sp.)
LASRIAMA SIAHAAN
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul : Keragaman Genetik Cytochrome b pada Burung Mambruk (Goura sp.) Nama : Lasriama Siahaan
NRP : G04400032
Menyetujui,
Pembimbing I, Pembimbing II,
Dr. Ir. Dedy Duryadi Solihin, DEA drh. Djoko Waluyo, M.S NIP. 131415134 NIP. 130350056
Mengetahui,
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor
Dr. Ir. Yonny Koesmaryono, M.S NIP. 131473999
Penulis dilahirkan di Pangururan, Sumatera Utara pada tanggal 02 Mei 1983 dari
ayah Tigor Siahaan dan Ibu Tiarasi Simbolon. Penulis merupakan anak pertama dari
lima bersaudara.
Pada tahun 2000 penulis lulus dari SMUN 1 Pangururan dan pada tahun yang
sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB. Penulis
memilih Departemen Biologi, Fakultas Matematika dan Ilmu Pengetetahuan Alam.
DAFTAR ISI
Halaman
DAFTAR TABEL………..…….………... vi
DAFTAR GAMBAR………..…….………... vi
DAFTAR LAMPIRAN………. vi
PENDAHULUAN……….……… 1
BAHAN DAN METODE………. 2
Bahan……….……… 2
Metode……….……….. 2
Pengambilan sampel……….………. 2
Isolasi dan Purifikasi DNA………...……….. 2
Uji kualitas DNA……….………. 2
Amplifikasi Daerah Sitokrom b (Cyt b)……….……... 2
Perunutan Nukleotida dan Analisis Data……….……... 3
HASIL ……….………...………. 3
Amplifikasi Daerah Cytochrome b (Cyt b)…………..………... 3
Analisis Perunutan Nukleotida Gen Sitokrom b Parsial…... 3
PEMBAHASAN ………. 6
SIMPULAN DAN SARAN………. 8
DAFTAR PUSTAKA……….. 8
DAFTAR TABEL
Halaman
1 Rataan Komposisi Nukleotida Cyt b Parsial………... 4
2 Situs Kodon Penyandi Beserta Asam Amino………. 4
3 Jumlah dan Posisi Basa dari Triplet Kodon Beragam……….. 5
4 Rataan Tansisi Basa ke-1,2,3 dan Rataan Transversi………. 5
5 Jarak Genetik Nukleotida Cyt b Parsial……….. 6
DAFTAR GAMBAR Halaman 1 DNA Mitokondria Avian (Gallus gallus)………... 2
2 Profil Fragmen Produk PCR Cyt b Burung Mambruk………... 3
3 Situs Pengenalan Primer dan Hasil Penjajaran Nukleotida………... 3
4 Rekonstruksi Filogenetik Nukleotida Cyt b Parsial……… 6
DAFTAR LAMPIRAN Halaman 1 Peta Penyebaran Goura sp………... 11
2 Hasil Perunutan Nukleotida Cyt b (382 Nukleotida) Goura sp……… 13
3 Matriks Perbedaan Nukleotida dan Jarak Genetik Goura sp…………... 15
4 Penjajaran Berganda Nukleotida Cyt b Parsial Goura sp……… 16
5 Penjajaran Berganda Asam Amino Cyt b Parsial Goura sp……… 18
6 Jumlah Penggunaan Kodon pada Cyt b Parsial Goura sp………. 19
PENDAHULUAN
Sekitar 17% populasi burung di dunia ada di Indonesia, termasuk burung yang jenisnya endemik. Salah satu burung endemik Indonesia adalah burung Mambruk atau Dara mahkota (Goura sp.). Sibley dan Ahlquist (1991) mengelompokkan Goura sp. dalam superordo: Passerimorphae; ordo: Columbiformes; famili : Columbidae (Pigeons dan Doves); sub-famili: Gourinae; genus: Goura. Genus ini memiliki tiga spesies antara lain: Goura victoria (Mambruk raja/ Mambruk kembang) yang terbesar dalam kelompok pigeon (Perrins & Middleton 1985),
G. cristata (Mambruk polos/Mambruk kelabu) dan G. scheepmakeri (Mambruk besar/Mambruk ungu). Secara umum G. cristata (Western crowned-pigeon) hanya tersebar di daerah Kepala Burung Irian Jaya (Lampiran 1a), G. scheepmakeri (Southern crowned-pigeon) tersebar di daerah selatan Irian Jaya dan Papua New Guinea (Lampiran 1b). sedangkan G. victoria (Victoria crowned-pigeon) tersebar di bagian utara Irian Jaya dan Papua New Guinea (Lampiran 1c).
Burung Mambruk merupakan satwa yang dilindungi oleh pemerintah Indonesia yang tercantum dalam undang-undang Peraturan Perlindungan Binatang Liar 1931 (tertulis semua jenis dari genus Goura) dan Peraturan pemerintah No. 7 Tahun 1999 untuk semua jenis dari genus
Goura (Noerdjito & Maryamto 2001). Burung Mambruk termasuk dalam status rawan (“vulnerable”) artinya spesies ini akan menjadi genting (“endangered”) dimasa mendatang populasinya yang terus menurun sebagai akibat dari daerah habitatnya yang semakin sempit, diburu untuk dijadikan sebagai satwa peliharaan karena nilai jualnya cukup tinggi dan untuk dikonsumsi sebagai sumber protein bagi penduduk setempat. Berat rata-rata burung Mambruk dapat mencapai 1800-2400 gram dengan tinggi 66-79 cm, sehingga memungkinkan untuk dijadikan sebagai sumber protein hewani (Kilmaskossu 2001).
Upaya mempertahankan dan melestarikan suatu organisme memerlukan data informasi lengkap seperti morfologi, sifat biologi, ekologi persarangan, dan musim perkembangbiakan (Kilmaskossu 2001), keragaman genetik dan determinasi jenis kelamin (Duryadi 2002). Usaha-usaha penangkaran pada kondisi alami, semi alami atau budidaya (pemeliharaan intensif)
perlu untuk menunjang konservasi burung ini agar dapat mencegah kepunahan lebih lanjut.
Informasi genetik yang telah didapat mengenai genom kelompok burung (avian) telah berkembang dengan pesat. Berdasarkan kandungan protein putih telur, Sibley dan Ahlquist (1991) menyimpulkan bahwa genom kelompok burung umumnya terdiri atas 60-70% kopi linier, 13-20% ruas pengulangan intermedian dan 16-20% ruas berulang yang tinggi. Data ini telah digunakan untuk keperluan eksplorasi keragaman genetik dan sejarah evolusi asal usul kelompok burung.
Penelitian Johnson dan Clayton (2000), menguji keabsahan perbandingan filogenetik famili Columbiformes dengan membandingkan DNA inti, yaitu β-fibrinogen
intron-7 (FIB7) dan mitokondria (cytochrome
b/ Cyt b), hasilnya menunjukkan perbedaan nukleotida antar spesies Columbiformes sekitar 0.97-17.07% untuk Cyt b dan 0.27-7.03% untuk FIB7. Ini mengindikasikan bahwa Cyt b berkembang 5.6 kali lebih cepat dari FIB7. Uji pada daerah non-coding DNA mitokondria yaitu D-loop didapatkan bahwa substitusi transversi lebih sedikit, walaupun demikian data ini masih dapat membantu untuk determinasi kejadian mutasi untuk mtDNA.
KERAGAMAN GENETIK CYTOCHROME B
PADA BURUNG MAMBRUK (Goura sp.)
Oleh:
Lasriama Siahaan G04400032
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
ABSTRAK
LASRIAMA SIAHAAN. Keragaman Genetik Cytochrome b pada Burung Mambruk (Goura sp.). Dibimbing oleh DEDY DURYADI SOLIHIN dan DJOKO WALUYO.
Burung Mambruk (Goura sp.) merupakan satwa endemik Indonesia dengan status “Vulnerable” yang tersebar di daerah Pulau Irian Jaya dan beberapa daerah Papua New Guinea. Usaha konservasi terhadap spesies ini akan berhasil jika karakteristik morfologi, keragaman molekuler dan genetik dapat diketahui dengan pasti. Tujuan penelitian ini untuk menganalisis keragaman genetik cytochrome b parsial dengan metode Polimerase Chain Reaction. Hasil perunutan dari amplifikasi pasangan primer M101 dan M102 pada cytochrome b parsial sepanjang 382 nukleotida (menyandikan 127 asam amino) disejajarkan (multiple aligment) dengan bantuan perangkat lunak Genetyx-Win versi 3.0 dan Clustal-X, selanjutnya dianalisis dengan program MEGA versi 3.0.
Hasil analisis dari 382 nukleotida yang dibandingkan terdapat 42 situs nukleotida yang beragam dengan rata-rata kejadian substitusi transisi 0.05 dan rata-rata substitusi transversi 0.01. Perubahan bersifat non-sinonimus 7.08% (10 situs asam amino) dan perubahan bersifat sebesar 23.62% (30 situs asam amino). Jarak genetik nukleotida cytochrome b (metode p-distance) didapat bahwa nilai paling kecil adalah 0.26% dan nilai yang paling besar 7.07% dengan rata-rata sebesar 5.26%. Hasil rekonstruksi filogenetik dengan metode Neighbor Joining menunjukkan bahwa G.cristata lebih berkerabat dekat dengan G.scheepmakeri daripada G.victoria.
Kata kunci: Goura sp., Cytochrome b, substitusi, perubahan
ABSTRACT
LASRIAMA SIAHAAN. Genetic Diversity of Cytochrome b of Crowned-pigeons (Goura sp.), Supervised by DEDY DURYADI SOLIHIN and DJOKO WALUYO.
Crowned-pigeon (Goura sp.) was one of endemic animal in Indonesia with “vulnerable" status which spread at Papua New Guinea and Irian Jaya Island. Conservation effort to this species will succeed if morphology characteristic and genetic diversity be known. The main purpose of this research is to analyze nucleotide variability of cytochrome b partial using Polymerase Chain Reaction method. The PCR product amplified by primer M101 and M102, then multiple aligment using Genetyx-Win 3.0™ and Clustal-X™, furthermore analyzed with MEGA 3.0™.
The result of analyse from 382 nucleotide of cytochrome b partial encoding 127 amino acid. Findings 42 sites of nucleotide variable with average of transitional substitutions 0.05 and average transversions substitutions is 0.01. Value non-synonymous mutations is 7.08% (10 amino acid sites) whereas synonymus mutations is 23.62% (30 amino acid sites). Value of genetic distance of nucleotide cytochrome b (method p-distance) range from 0.26% - 7.07% with average 5.26%. Filogenetic reconstruction using Neighbor Joining method indicate that G.cristata were closer to G.scheepmakeri compare with G.victoria.
mitokondria tidak hanya ditemukan pada kelompok burung, hal sama yang terjadi pada kelompok Rodensia dan Ikan (Kocher et al. 1989). Komposisi nukleotida-2 dari triplet kodon paling tidak beragam karena komposisi A (17.3%) sama, demikian pula dengan G (14.2%) untuk Goura sp. dalam penelitian ini.
Basa dari triplet kodon yang paling banyak mengalami perubahan substitusi adalah basa ke-3 sebesar 22.04% (28 situs), hal ini sesuai dengan pendapat Sorenson (2003) dimana kejadian substitusi pada basa ke-3 dari triplet kodon lebih banyak terjadi.
Perubahan penerjemahan asam amino terjadi karena adanya substitusi nukleotida transisi (perubahan antar basa purin atau antar basa pirimidin) dan substitusi transversi (perubahan dari basa purin menjadi basa pirimidin atau sebaliknya). Transisi pada basa ke-3 tidak menyebabkan perubahan asam amino dan transversi basa ke-3 tidak selalu menyebabkan perubahan asam amino.
Perubahan basa triplet kodon yang menyebabkan terjadinya perubahan asam amino non-sinonim adalah perubahan basa ke-1 sebesar 3.15% (4 situs asam amino), basa ke-2 sebesar 1.57% (2 situs asam amino), basa ke-3 sebesar 1.57% (2 situs asam amino), basa ke-1 dan 3 sebesar 1.57% (2 situs asam amino). Sedangkan perubahan asam amino sinonim terdapat pada perubahan basa ke-3 sebesar 20.47% (26 situs asam amino), basa ke-1 sebesar 3.15% (4 situs asam amino). Perubahan basa ke-1 umumnya menyebabkan perubahan asam amino. Kecuali pada asam amino Leu (leusin), walaupun transisi dan transversi terjadi pada basa ke-1 dan ke-3 tidak akan menyebabkan perubahan translasi asam aminonya.
Dari 127 asam amino yang diamati terdapat 68.50% (87 situs) merupakan asam amino kekal (nukleotidanya tidak mengalami substitusi), 7.87% (10 situs) perubahan bersifat non-sinonim (nukleotida mengalami substitusi dan asam amino berubah) dan 23.62% (30 situs) perubahan bersifat sinonim (nukleotida mengalami substitusi dan asam amino tetap).
Walaupun telah terjadi perubahan pada 42 situs nukleotida namun hanya 10 situs nukleotida beragam yang dapat mengubah asam amino (10 asam amino). Sepuluh situs asam amino yang berubah terjadi secara transisi dan transversi basa ke-1 (4 situs asam amino yaitu L (leusina) menjadi F (fenilalanina), M (metionina) menjadi L (leusina), T (treonina)
menjadi A (alanina) dan F (fenilalanina) menjadi L (leusina); transisi basa ke-2 (2 situs asam amino yaitu F (fenilalanina) menjadi S (serina) dan M (metionina) menjadi T (treonina); transversi basa ke-3 (2 situs asam amino yaitu C (sisteina) menjadi W (triptofan); serta perpaduan transisi basa ke-1 dan transversi basa ke-3 (2 situs asam amino yaitu S (serina) menjadi P (prolina) dan L (leusina) menjadi F (fenilalanina). Sedangkan situs nukleotida yang mengalami substitusi lebih banyak (30 situs asam amino) terjadi secara sinonim bukan non-sinonim sehingga tidak mengubah asam aminonya.
Kelompok burung (spesies, genera dan famili) memiliki rata-rata jarak genetik yang lebih kecil daripada jarak takson yang sama antara mamalia, amphibi dan reptil. Secara umum kelompok burung memiliki jarak genetik intraspesies yang sangat kecil (Stanley & Harrison 1999). Jarak genetik (p-distance) berdasarkan jumlah nukleotida yang berbeda, memperlihatkan jarak genetik intraspesies
Goura sp. yang paling rendah adalah 0.26% (G. scheepmakeri dengan G. cristata) dan paling tinggi sebesar 7.07% (G. victoria GenBank
dengan G. victoria hasil penelitian) dengan rata-rata sebesar 5.26%. Rata-rata-rata jarak genetik antar spesies yang dibandingkan adalah 11.54% dan jarak genetik rata-rata famili Columbidae (data dari GenBank) sebesar 9.79%. Jarak genetik ini mendukung hasil penelitian Johnson & Clayton (2000) yaitu didapatkan bahwa perbedaan nukleotida Cyt b antar spesies Columbiformes sekitar 0.97-17.07%.
Hasil ini juga mengasumsikan kekerabatan yang dekat di antara Goura sp. bahkan antar spesies Columbidae. Kekerabatan genetik antara G. cristata dengan G. scheepmakeri
(dengan nilai bootstrap 80%) lebih dekat dengan morfologi yang lebih mirip jika dibandingkan dengan G. victoria. Burung Mambruk dari jenis G. scheepmakeri berada di antara G. cristata dan G. victoria.
KATA PENGANTAR
Persembahan syukur kepada Allah Bapa atas anugerahNya, sehingga penulis
dapat menyelesaikan karya ilmiah ini. Tema yang dipilih dalam penelitian yang
dilaksanakan sejak bulan September 2004 ini adalah ”Keragaman Genetik
Cytochrome
b
pada Burung Mambruk (
Goura
sp.).
Terima kasih penulis ucapkan kepada Bapak Dr. Ir. Dedy Duryadi Solihin, DEA
dan Bapak drh. Djoko Waluyo, M.S selaku pembimbing dan penyandang dana dalam
penelitian ini. Disamping itu, penghargaan penulis sampaikan kepada Taman Burung
Taman Mini Indonesia Indah (TMII) yang telah membantu dalam hal penyediaan contoh
darah burung Mambruk. Tak lupa penulis sampaikan kepada Bapak Heri yang telah
membantu dalam masalah tehnik laboratorium serta rekan-rekan kerja di Laboratorium
Biologi Molekuler Pusat Studi Ilmu Hayati IPB, Ibu Rini, K’ Chule, K’Evi, Virgo atas
saran, dukungan dan kerjasama yang terjalin selama bekerja di Laboratorium.
Terimakasih juga penulis ucapkan kepada saudara terkasih Ike, T’ Lucien, K’ Regina, K’
Anna, B’ Andrew, Pemuda GKI Pengadilan Bogor atas segala bentuk dorongan dan
perhatiannya, Lies dan Biologi angkatan 37’ atas kebersamaannya. Segala cinta dan
terima kasih penulis ungkapkan kepada Bapak, Oma, Nardus, Herman, Okki, Mindo atas
doa, dukungan moral, material, kesabaran dan kasih sayangnya.
Berharap karya ilmiah ini bermanfaat.
Bogor, Maret 2006
PADA BURUNG MAMBRUK (
Goura
sp.)
LASRIAMA SIAHAAN
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul : Keragaman Genetik Cytochrome b pada Burung Mambruk (Goura sp.) Nama : Lasriama Siahaan
NRP : G04400032
Menyetujui,
Pembimbing I, Pembimbing II,
Dr. Ir. Dedy Duryadi Solihin, DEA drh. Djoko Waluyo, M.S NIP. 131415134 NIP. 130350056
Mengetahui,
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor
Dr. Ir. Yonny Koesmaryono, M.S NIP. 131473999
Penulis dilahirkan di Pangururan, Sumatera Utara pada tanggal 02 Mei 1983 dari
ayah Tigor Siahaan dan Ibu Tiarasi Simbolon. Penulis merupakan anak pertama dari
lima bersaudara.
Pada tahun 2000 penulis lulus dari SMUN 1 Pangururan dan pada tahun yang
sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB. Penulis
memilih Departemen Biologi, Fakultas Matematika dan Ilmu Pengetetahuan Alam.
DAFTAR ISI
Halaman
DAFTAR TABEL………..…….………... vi
DAFTAR GAMBAR………..…….………... vi
DAFTAR LAMPIRAN………. vi
PENDAHULUAN……….……… 1
BAHAN DAN METODE………. 2
Bahan……….……… 2
Metode……….……….. 2
Pengambilan sampel……….………. 2
Isolasi dan Purifikasi DNA………...……….. 2
Uji kualitas DNA……….………. 2
Amplifikasi Daerah Sitokrom b (Cyt b)……….……... 2
Perunutan Nukleotida dan Analisis Data……….……... 3
HASIL ……….………...………. 3
Amplifikasi Daerah Cytochrome b (Cyt b)…………..………... 3
Analisis Perunutan Nukleotida Gen Sitokrom b Parsial…... 3
PEMBAHASAN ………. 6
SIMPULAN DAN SARAN………. 8
DAFTAR PUSTAKA……….. 8
DAFTAR TABEL
Halaman
1 Rataan Komposisi Nukleotida Cyt b Parsial………... 4
2 Situs Kodon Penyandi Beserta Asam Amino………. 4
3 Jumlah dan Posisi Basa dari Triplet Kodon Beragam……….. 5
4 Rataan Tansisi Basa ke-1,2,3 dan Rataan Transversi………. 5
5 Jarak Genetik Nukleotida Cyt b Parsial……….. 6
DAFTAR GAMBAR Halaman 1 DNA Mitokondria Avian (Gallus gallus)………... 2
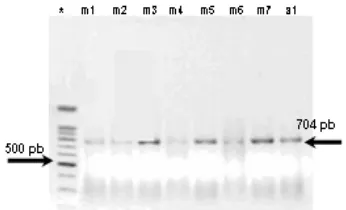
2 Profil Fragmen Produk PCR Cyt b Burung Mambruk………... 3
3 Situs Pengenalan Primer dan Hasil Penjajaran Nukleotida………... 3
4 Rekonstruksi Filogenetik Nukleotida Cyt b Parsial……… 6
DAFTAR LAMPIRAN Halaman 1 Peta Penyebaran Goura sp………... 11
2 Hasil Perunutan Nukleotida Cyt b (382 Nukleotida) Goura sp……… 13
3 Matriks Perbedaan Nukleotida dan Jarak Genetik Goura sp…………... 15
4 Penjajaran Berganda Nukleotida Cyt b Parsial Goura sp……… 16
5 Penjajaran Berganda Asam Amino Cyt b Parsial Goura sp……… 18
6 Jumlah Penggunaan Kodon pada Cyt b Parsial Goura sp………. 19
PENDAHULUAN
Sekitar 17% populasi burung di dunia ada di Indonesia, termasuk burung yang jenisnya endemik. Salah satu burung endemik Indonesia adalah burung Mambruk atau Dara mahkota (Goura sp.). Sibley dan Ahlquist (1991) mengelompokkan Goura sp. dalam superordo: Passerimorphae; ordo: Columbiformes; famili : Columbidae (Pigeons dan Doves); sub-famili: Gourinae; genus: Goura. Genus ini memiliki tiga spesies antara lain: Goura victoria (Mambruk raja/ Mambruk kembang) yang terbesar dalam kelompok pigeon (Perrins & Middleton 1985),
G. cristata (Mambruk polos/Mambruk kelabu) dan G. scheepmakeri (Mambruk besar/Mambruk ungu). Secara umum G. cristata (Western crowned-pigeon) hanya tersebar di daerah Kepala Burung Irian Jaya (Lampiran 1a), G. scheepmakeri (Southern crowned-pigeon) tersebar di daerah selatan Irian Jaya dan Papua New Guinea (Lampiran 1b). sedangkan G. victoria (Victoria crowned-pigeon) tersebar di bagian utara Irian Jaya dan Papua New Guinea (Lampiran 1c).
Burung Mambruk merupakan satwa yang dilindungi oleh pemerintah Indonesia yang tercantum dalam undang-undang Peraturan Perlindungan Binatang Liar 1931 (tertulis semua jenis dari genus Goura) dan Peraturan pemerintah No. 7 Tahun 1999 untuk semua jenis dari genus
Goura (Noerdjito & Maryamto 2001). Burung Mambruk termasuk dalam status rawan (“vulnerable”) artinya spesies ini akan menjadi genting (“endangered”) dimasa mendatang populasinya yang terus menurun sebagai akibat dari daerah habitatnya yang semakin sempit, diburu untuk dijadikan sebagai satwa peliharaan karena nilai jualnya cukup tinggi dan untuk dikonsumsi sebagai sumber protein bagi penduduk setempat. Berat rata-rata burung Mambruk dapat mencapai 1800-2400 gram dengan tinggi 66-79 cm, sehingga memungkinkan untuk dijadikan sebagai sumber protein hewani (Kilmaskossu 2001).
Upaya mempertahankan dan melestarikan suatu organisme memerlukan data informasi lengkap seperti morfologi, sifat biologi, ekologi persarangan, dan musim perkembangbiakan (Kilmaskossu 2001), keragaman genetik dan determinasi jenis kelamin (Duryadi 2002). Usaha-usaha penangkaran pada kondisi alami, semi alami atau budidaya (pemeliharaan intensif)
perlu untuk menunjang konservasi burung ini agar dapat mencegah kepunahan lebih lanjut.
Informasi genetik yang telah didapat mengenai genom kelompok burung (avian) telah berkembang dengan pesat. Berdasarkan kandungan protein putih telur, Sibley dan Ahlquist (1991) menyimpulkan bahwa genom kelompok burung umumnya terdiri atas 60-70% kopi linier, 13-20% ruas pengulangan intermedian dan 16-20% ruas berulang yang tinggi. Data ini telah digunakan untuk keperluan eksplorasi keragaman genetik dan sejarah evolusi asal usul kelompok burung.
Penelitian Johnson dan Clayton (2000), menguji keabsahan perbandingan filogenetik famili Columbiformes dengan membandingkan DNA inti, yaitu β-fibrinogen
intron-7 (FIB7) dan mitokondria (cytochrome
b/ Cyt b), hasilnya menunjukkan perbedaan nukleotida antar spesies Columbiformes sekitar 0.97-17.07% untuk Cyt b dan 0.27-7.03% untuk FIB7. Ini mengindikasikan bahwa Cyt b berkembang 5.6 kali lebih cepat dari FIB7. Uji pada daerah non-coding DNA mitokondria yaitu D-loop didapatkan bahwa substitusi transversi lebih sedikit, walaupun demikian data ini masih dapat membantu untuk determinasi kejadian mutasi untuk mtDNA.
Gambar 1 DNA mitokondria avian (Gallus gallus).
Kurangnya informasi mengenai burung Mambruk ini baik secara bioekologi, morfologi, molekuler atau genetik, untuk keperluan data di lapangan menjadi salah satu kesulitan bagi usaha-usaha konservasi dan peningkatan populasinya. Data hasil penelitian ini diharapkan dapat dijadikan acuan dalam penelusuran asal usul dan perbedaan dari ketiga spesies burung Mambruk yang ada.
Penelitian ini bertujuan menganalisis keragaman genetik cytochrome b burung Mambruk dengan merunut sebagian situs gen penyandi Cyt b.
Penelitan ini dimulai pada bulan September 2004 hingga November 2005 di Laboratorium Biologi Molekuler, Pusat Studi Ilmu Hayati (PSIH), IPB, Bogor.
BAHAN DAN METODE
Bahan
Bahan yang digunakan dalam penelitian ini adalah contoh darah burung Mambruk sebanyak 8 ekor yakni 7 ekor koleksi Taman Burung Taman Mini Indonesia Indah (TMII) dan 1 ekor koleksi Laboratorium Biologi Molekuler PSIH, IPB, Bogor.
Metode
Pengambilan Sampel. Sampel darah diambil dari sayap tepatnya di bagian vena vulgaris
menggunakan syringe 1cc/ml yang sebelumnya telah diisi dengan ¾ volume 10% etylene tetra
acetic acid (EDTA) sebagai anti koagulan, kemudian disimpan pada suhu -20oC.
Isolasi dan purifikasi DNA. DNA total hasil isolasi dipurifikasi dengan metode purifikasi fenol yang telah dimodifikasi. Dalam tabung eppendorf 1.5 ml dimasukkan sampel darah sebanyak 250-500 µl dan ditambahkan dengan lysis buffer 1x volume dikocok sampai homogen. Campuran disentrifugasi pada 6500 rpm selama 1 menit. Pada endapan ditambahkan 200 µl rinse buffer,
dan divortex. Setelah itu 500 µl digestion buffer ditambahkan pada tabung, dikocok sampai homogen lalu diinkubasi pada suhu 55oC sampai semalam (± 16 jam). Setelah inkubasi, 500 µl larutan fenol ditambahkan pada supernatan, dikocok selama 20 menit, lalu disentrifugasi pada 13.000 rpm selama 3 menit. Supernatan dipindahkan ke dalam tabung baru dan ditambah dengan Chloroform isoamil alcohol (CIAA) sebanyak 500 µl dikocok hingga homogen serta disentrifugasi pada 13.000 rpm selama 3 menit. Supernatan yang terbentuk dipindahkan ke dalam tabung baru dan ditambahkan etanol absolut sebanyak 2x volume, disentrifugasi pada 13.000 rpm selama 3 menit hingga terdapat endapan putih. Endapan tersebut dicuci dengan menambahkan alkohol 70% 1x volume dengan sentrifugasi 13.000 rpm selama 3 menit. Endapan kemudian dikeringkan pada suhu ruang. Selanjutnya pada endapan ditambahkan Tris EDTA (TE), disentrifugasi pelan dan diinkubasi pada suhu 37oC selama 15 menit lalu disimpan dalam freezer.
Uji kualitas DNA. DNA total yang telah dipurifikasi kemudian dielektroforesis pada gel agarose 1.2% menggunakan larutan buffer 1xTBE (89 mM Tris, 89 mM Asam Borat dan 2 mM EDTA, pH 8.0) dalam piranti submarine
electrophoresis (Hoefer). DNA
divisualisasikan menggunakan UV Transluminator ( λ=260 nm).
Amplifikasi daerah Cytochrome b. Hasil ekstraksi berupa DNA total digunakan sebagai cetakan untuk mengamplifikasi daerah Cyt b. Sepasang primer digunakan untuk mengamplifikasi daerah Cyt b yaitu M101 5’-CAA ATC CTC ACA GGC CTA TTC CTA GC -3’ (forwad), dan M102 5’-TAG GCG AAT AGG AAA TAT CAT TCG GGT TGA T-3’ (reverse). Pasangan primer ini mengamplifikasi dengan kondisi pre-denaturasi pada suhu 94oC selama 3 menit,
16.7 kb
3
denaturasi pada suhu 94oC selama 30 detik, anealing pada suhu 54oC selama 1 menit, elongasi pada suhu 72oC selama 2 menit. Siklus ini diulang sebanyak 35 kali, selanjutnya post-elongasi pada suhu 72oC selama 5 menit. PCR dilakukan dengan menggunakan mesin GeneAmp® PCR System 2400 (Perkin-Elmer).
Komposisi untuk tiap reaksi PCR terdiri dari DNA total 2-5 µl (10-100 ng), 10x buffer PCR mix 5 µl, 25 mM MgCl2 2 µl, 10 mM dNTP mix
1 µl, 25 pM primer M101 dan M102 masing-masing 2 µl, dan 5 unit/µl Tag DNA polymerase
0.2 µl dan air steril hingga volume total 50 µl.
Perunutan nukleotida dan analisis data.
DNA Cyt b produk PCR dirunut dengan menggunakan mesin pengurut DNA otomatis ABI Prism versi 3.4.1 (USA). Data perunutan yang diperoleh disejajarkan (alignment) dengan menggunakan perangkat lunak Genetix-Win versi 3.0 dan Clustal-W (Thompson et al. 1994). Kemudian basa-basa asam amino gen penyandi
Cyt b diterjemahkan dengan menggunakan
Vertebrate mitochondrial translation code yang terdapat dalam MEGA versi 3.0.
Sebagai pembanding dalam penelitian ini digunakan nukleotida lengkap Gallus gallus
(Chicken) dari famili Phasianidae (nomor akses NC007237, Desjardins & Morais 1990 ) dan spesies famili Columbidae (Lampiran 6) yaitu:
Alectroenas madagascarensis dari famili Columbidae (Madagascar blue-pigeon) (AF483344, Shapiro et al. 2002), Didunculus strigirostris dari famili Columbidae (tooth-billed pigeon) (AF483343, Shapiro et al. 2002),
Leptotila verreauxi fulviventri dari famili Columbidae (white-tipped doves) (AF279704, Clayton et al. 2003), Ptilinopus superbus dari famili Columbidae (fruit-doves) (AF483329, Shapiro et al. 2002), Columba livia dari famili Columbidae (Rock Pigeon/common pigeon) (AF182694, Johnson & Clayton 2000), Leptotila megalura dari famili Columbidae (white-faced dove) (AF182697, Johnson & Clayton 2000).
Analisis rekonstruksi filogenetik menggunakan perangkat lunak MEGA versi 3.0 (Kumar et al. 2004) dengan menggunakan
bootstrapped Neighbor-Joining dengan 1000 kali pengulangan.
HASIL
Hasil identifikasi pengamatan morfologi setiap sampel dari Taman Burung/TMII (n=7) didapatkan 1 ekor G. victoria (m1); 2 ekor G.
cristata (m2, m3); 4 ekor G. scheepmakeri
(m4, m5, m6, m7) dan 1 ekor G. victoria (a1) koleksiLaboratorium Biologi Molekuler.
Amplifikasi daerah Cyt b
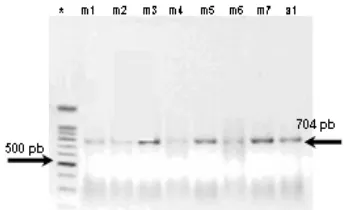
Hasil amplifikasi Cyt b menggunakan pasangan primer M101 dan M102 adalah sepanjang 704 bp (Gambar 2) dari 1045 nukleotida (nt) Cyt b utuh G. cristata (nomor akses AF182709) terletak pada situs ke-35 sampai dengan situs yang ke-738 (Johnson & Clayton 2000).
* Marker 50 bp; cetak tebal produk PCR yang dirunutkan yaitu m3 (G. cristata); m7 (G. scheepmakeri); a1 (G. victoria).
Gambar 2 Hasil elektroforesis pada gel agarose fragmen produk PCR (704 bp) dari
Cyt b burung Mambruk.
Analisis perunutan nukleotida Cyt b parsial
Delapan contoh darah yang diamplifikasi, empat di antaranya dirunut DNAnya (sequencing) dan satu tidak digunakan sebagai data karena banyak terdapat N. Fragmen produk PCR sepanjang 704 bp yang dirunut dengan mesin perunut otomatis ABI Prism
versi 3.4.1 didapat 382 nukleotida (nt) yang jelas, tidak ambigius dan baik urutannya (Lampiran 2). Setelah disejajarkan (alignment) dengan nukleotida Cyt b Gallus gallus dari
GenBank dan spesies lain dari famili Columbidae sebagai pembanding, didapat hasil bahwa fragmen sepanjang 382 nt (Lampiran 4) ini terletak pada situs ke-242 sampai dengan situs ke-623 Cyt b utuh G. cristata GenBank
(Gambar 3).
Primer M101 26 bp
Primer M102 31 bp
704 bp Sit b (1045 nt)
tRNA-Thr ND5
382 nt *
... produk PCR situs ke 35-738 _ _ _ hasil perunutan situs ke 242-623
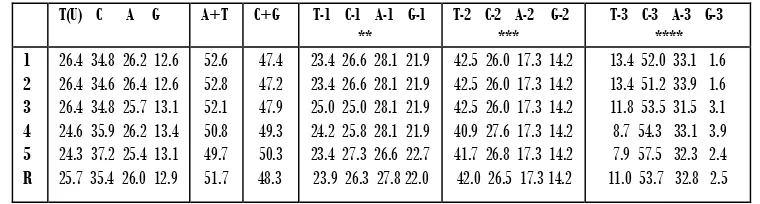
Hasil penjajaran sepanjang 382 nukleotida paling banyak ditemukan fragmen tersebut adalah nukleotida C (35.4%), diikuti dengan A (26.0%), T (25.7%) dan yang paling sedikit adalah G (12.9%). Komposisi pasangan nukleotida A+T lebih tinggi pada G. cristata (52.8%), dan paling rendah pada G. victoria 49.7% dengan rataan 51.7%, sedangkan untuk komposisi pasangan nukleotida C+G lebih tinggi pada G. victoria
(50.3%) dan paling rendah pada G. cristata
(47.2%) dengan rataan 48.3% (Tabel 1).
Fragmen sepanjang 382 nukleotida ini menyandikan 127 asam amino yaitu mulai dari asam amino ke 81 sampai dengan asam amino ke 207 (situs acuan adalah asam amino hasil translasi
Cyt b utuh G. Cristata GenBank). Komposisi basa ke-1 pada triplet kodon (nukleotida-1) paling banyak ditemukan adalah nukleotida A (28.1%) diikuti oleh nukleotida C (27.3%), T (25.0%) dan G (22.7%). Komposisi basa ke-2 dari triplet kodon (nukleotida-2) paling banyak (Tabel 1) adalah nukleotida T (42.5%) diikuti oleh nukleotida C (26.0%), A (17.3%), G (14.2%). Sedangkan untuk komposisi basa ke-3 dari triplet kodon (nukleotida-ke-3) yang paling banyak (Tabel 1) adalah nukleotida C (53.7%) diikuti dengan nukleotida A (32.8%), T (11.0%), dan G (2.5%) (Tabel 1).
Tabel 1 Rataan komposisi nukleotida Cyt b parsial Goura sp. hasil penelitian, G. cristata GenBank,
dan G. victoriaGenBank
T(U) C A G A+T C+G T-1 C-1 A-1 G-1 **
T-2 C-2 A-2 G-2 ***
T-3 C-3 A-3 G-3 **** 1 2 3 4 5 R
26.4 34.8 26.2 12.6 26.4 34.6 26.4 12.6 26.4 34.8 25.7 13.1 24.6 35.9 26.2 13.4 24.3 37.2 25.4 13.1 25.7 35.4 26.0 12.9
52.6 52.8 52.1 50.8 49.7 51.7 47.4 47.2 47.9 49.3 50.3 48.3
23.4 26.6 28.1 21.9 23.4 26.6 28.1 21.9 25.0 25.0 28.1 21.9 24.2 25.8 28.1 21.9 23.4 27.3 26.6 22.7 23.9 26.3 27.8 22.0
42.5 26.0 17.3 14.2 42.5 26.0 17.3 14.2 42.5 26.0 17.3 14.2 40.9 27.6 17.3 14.2 41.7 26.8 17.3 14.2 42.0 26.5 17.3 14.2
13.4 52.0 33.1 1.6 13.4 51.2 33.9 1.6 11.8 53.5 31.5 3.1 8.7 54.3 33.1 3.9 7.9 57.5 32.3 2.4 11.0 53.7 32.8 2.5
1.G. cristata*; 2. G. cristata._m3; 3.G. scheepmakeri._m7; 4. G. victoria_a1; 5. G. victoria*; R.Avg;
* GenBank; ** basa ke-1 dari triplet kodon; ** basake-2 dari triplet kodon *; **** basa ke-3 dari triplet kodon.
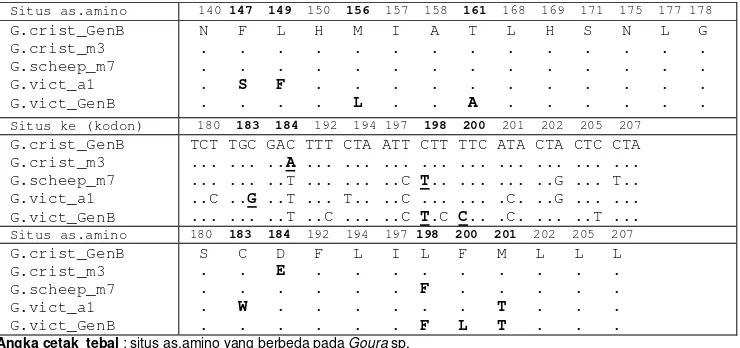
Hasil penjajaran (382 nt) nukleotida G. victoria GenBank, G. cristata GenBank, Goura sp. hasil penelitian terdapat 42 situs nukleotida beragam yang terdapat dalam 40 situs asam amino (Tabel 2). Situs nukleotida
yang beragam merupakan hasil kejadian substitusi basa ke-3 (paling banyak) diikuti dengan substitusi basa ke-1, basa ke-2 dan basa ke-1 dan 3 (Tabel 3).
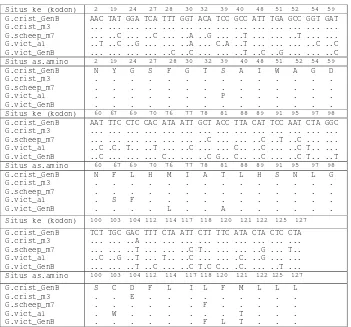
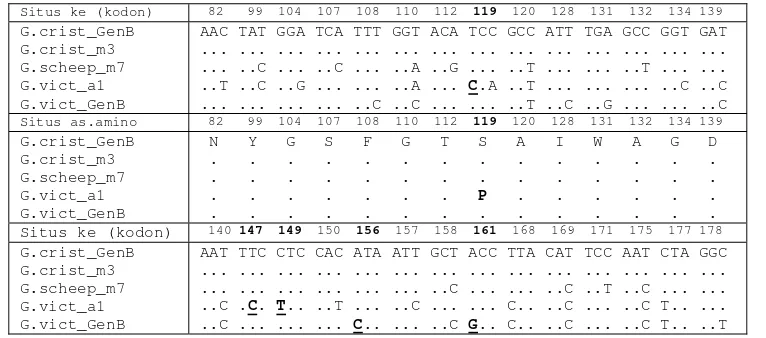
Tabel 2 Situs kodon penyandi beserta asam amino yang mengalami perubahan pada Cyt b parsial dari
G. cristata GenBank, G. victoria GenBank, Goura sp. hasil penelitian
Situs ke (kodon) 82 99 104 107 108 110 112 119 120 128 131 132 134 139 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
AAC TAT GGA TCA TTT GGT ACA TCC GCC ATT TGA GCC GGT GAT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..C ... ..C ... ..A ..G ... ..T ... ... ..T ... ... ..T ..C ..G ... ... ..A ... C.A ..T ... ... ... ..C ..C ... ... ... ... ..C ..C ... ... ..T ..C ..G ... ... ..C
Situs as.amino 82 99 104 107 108 110 112 119 120 128 131 132 134 139 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
N Y G S F G T S A I W A G D . . . . . . . . . . . P . . . . . . . . Situs ke (kodon) 140 147 149 150 156 157 158 161 168 169 171 175 177 178 G.crist_GenB
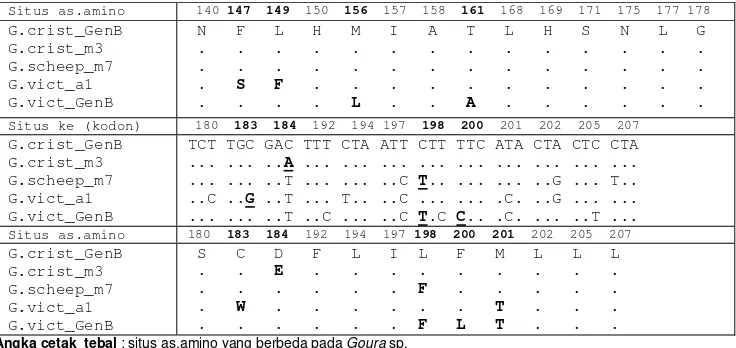
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
5
Situs as.amino 140 147 149 150 156 157 158 161 168 169 171 175 177 178 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
N F L H M I A T L H S N L G . . . . . . . . . S F . . . . . . . . L . . A . . . .
Situs ke (kodon) 180 183 184 192 194 197 198 200 201 202 205 207 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
TCT TGC GAC TTT CTA ATT CTT TTC ATA CTA CTC CTA ... ... ..A ... ... ... ... ... ... ... ... ... ... ... ..T ... ... ..C T.. ... ... ..G ... T.. ..C ..G ..T ... T.. ..C ... ... .C. ..G ... ... ... ... ..T ..C ... ..C T.C C.. .C. ... ..T ...
Situs as.amino 180 183 184 192 194 197 198 200 201 202 205 207 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
S C D F L I L F M L L L . . E . . . . . . . . . . . F . . . . . . W . . . . . . T . . . . . . . F L T . . .
Angka cetak tebal : situs as.amino yang berbeda pada Goura sp.
Huruf tebal, garis bawah : perubahn non-sinonimus (nukleotida berubah dan as.amino berubah).
Kodon ke-82 sama dengan asam amino ke-2 hasil penjajaran Kodon ke-207 sama dengan asam amino ke-127 hasil penjajaran
Tabel 3 Jumlah dan posisi basa dari triplet kodon
Cyt b parsial (42 situs) G. cristata GenBank, G. victoria GenBank dan
Goura sp. hasil penelitian
Basa beragam Jumlah situs nukleotida Jumlah Situs asam amino Ke-1 Ke-2 Ke-3 Ke-1 dan 3
8 2 28 4 8 (6.29%) 2 (1.57%) 28 (22.04%) 2 (1.57%)
Total 42 situs 40 situs
Kejadian substitusi transisi dan transversi memberi kontribusi pada keragaman nukleotida (Tabel 4). Kejadian substitusi transisi lebih besar pada tingkat spesies daripada nilai transversi. Sedangkan untuk tingkat genus umumnya nilai transversi lebih besar (Kocher et al. 1989). Nilai transisi terbesar 0.06 (G. victoria GenBank dan G. victoria hasil penelitian tehadap G. cristata GenBank; G. victoria GenBank terhadap G. victoria hasil penelitian) dan nilai terkecil 0.00 antara G. cristata GenBank terhadap G. cristata
hasil penelitian dimana rata-rata kejadian substitusi transisi sebesar 0.05. Sedangkan nilai transversi terbesar 0.01 dan terkecil adalah 0.00 dengan rata-rata 0.01.
Tabel 4 Rataan transisi basa ke-1,2,3 (di bawah diagonal) dan rataan transversi basa ke- 1, 2, 3 (diatas diagonal) pada
Goura sp. hasil penelitian, G. cristata GenBank dan G. victoria GenBank
1 2 3 4 5
1]G.crist_GenB 2]G.crist_m3 3]G.scheep_m 4]G.vict_a1 5]G.vict_GenB
0.00 0.01 0.01 0.00 0.00 0.01 0.01 0.01 0.03 0.03 0.01 0.01 0.06 0.05 0.05 0.01 0.06 0.05 0.05 0.06
Berdasarkan jumlah perbedaan nukleotida didapatkan jarak genetik intraspesies Goura
Tabel 5 Jarak genetik nukleotida Cyt b parsial (382 nt) G. cristata GenBank , G. victoria GenBank, Goura sp. hasil penelitian dan
Gallus gallus sebagai pembanding
1 2 3 4 5 6 1]Gal.gallus* 2]G.crist* 3]G.crist-m3 4]G.sche-m7 5]G.vict-a1 6]G.vict*
81 82 84 79 82 0.2120 1 15 25 23 0.2147 0.0026 15 25 23 0.2199 0.0393 0.0393 24 23 0.2068 0.0654 0.0654 0.0628 27 0.2147 0.0602 0.0602 0.0602 0.0707
* data GenBank
Jarak genetik (dibawah diagonal) dengan metode P-distance dan matriks perbedaan jumlah nukleotida (diatas diagonal).
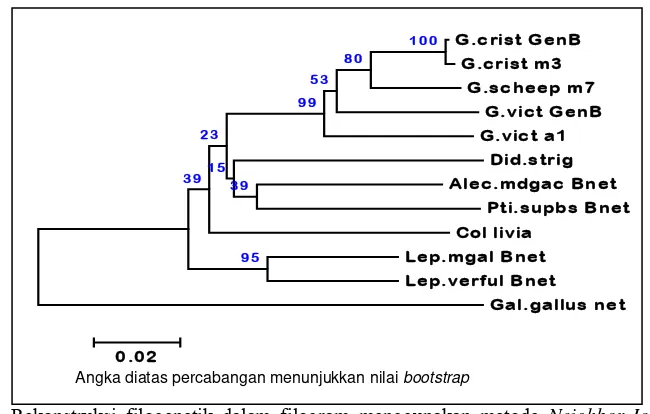
Filogram hasil rekonstruksi filogenetik nukleotida Cyt b parsial (Gambar 4) menunjukkan bahwa G. cristata GenBank, G. victoriaGenBank; Goura sp. hasil penelitian mengelompok dalam satu percabangan yang didukung dengan nilai bootstrap 99%. G. cristata GenBank dan G. cristata hasil penelitian berada dalam percabangan yang sama yang didukung dengan nilai bootstrap
100%. Selanjutnya G. cristata berada dalam cabang yang bersebelahan dengan G. scheepmakeri didukung dengan nilai
bootstrap 80%.
Gambar 4 Rekonstruksi filogenetik dalam filogram menggunakan metode Neighbor-Joining dari nukleotida Cyt b parsial (382 nt) Goura sp. hasil penelitian, spesies Columbidae dan
Gallus gallus sebagai pembanding.
PEMBAHASAN
Lokus cytochrome b menyimpan nilai yang berharga untuk analisis filogenetik pada tingkat spesies, dan famili pada kelompok burung (Bretagnolle et al. 1998; Randi 1996), karena ukuran nukleotida Cyt b setiap spesies memiliki variasi yang cukup tinggi. Menurut Johnson & Clayton (2000), penjajaran nukleotida Cyt b parsial tidak terdapat insersi atau delesi (indel), sehingga dianggap stabil dalam hal ukurannya (jumlah nukleotida). Pada G. cristata ukuran
gen penyandi Cyt b adalah sebesar 1045 nt dan menyandikan 346 asam amino (Johnson & Clayton 2000).
Pada Goura sp. nukleotida yang paling banyak ditemukan adalah C (35.4%), diikuti dengan A (26.0%), T (25.7%), dan yang paling sedikit adalah G (12.9%). Proporsi jumlah ini sesuai dengan pendapat Kocher et al. (1989) bahwa untuk kelompok Burung dan Ikan, nukleotida yang paling banyak adalah C diikuti dengan A, T dan G. Untuk setiap komposisi basa triplet kodon (nukleotida-1; nukleotida-2; nukleotida-3) menunjukkan bahwa nukleotida G selalu memiliki nilai paling rendah. Rendahnya nilai komposisi nukleotida G pada DNA
G .c rist G e n B G .c ris t m3
G.s c he e p m7 G .v ic t G e nB
G .v ic t a 1 Did.s trig A le c .mdga c B ne t
Pti.supbs B ne t Col liv ia
L e p.mga l B ne t L e p.v e rfu l B ne t
G a l.ga llu s n e t
1 0 0
9 5
8 0
5 3
9 9
3 9 3 91 5
2 3
0 .0 2
8
Perbedaan kodon bias tersebut didasarkan pada perhitungan jumlah efektif kodon (Nc)
dengan rentang indeks 60-20. Jika nilainya 60 maka semua kodon (di luar stop kodon) digunakan dengan frekuensi yang sama, sedangkan jika nilainya 20 yaitu hanya satu kodon digunakan untuk setiap asam amino. Dimana nilai efektif nukleotida terbesar adalah 60 dan terkecil adalah 20. Komposisi kodon (Lampiran 6) diterjemahkan berdasarkan
Vertebrate mitochondrial translation code yang terdapat dalam MEGA versi 3.0 (Kumar et al.
2004).
SIMPULAN DAN SARAN
Telah terjadi perubahan situs nukleotida pada Goura sp. sebesar 0.26%-7.07% dari 382 nukleotida namun yang berubah hanya 10 situs asam amino saja. Dari perubahan nukleotida yang ada didapat G. cristata lebih dekat kekerabatannya dengan G. scheepmakeri
daripada G. victoria.
Untuk penelitian selanjutnya perlu dilakukan perunutan produk PCR dengan situs yang berbeda untuk mendapatkan data yang lebih akurat, informatif, variatif dari daerah geografis yang diperkirakan sebagai daerah pertemuan Goura sp.
DAFTAR PUSTAKA
Benstead P, Dutson G. 2005. Western Crowned-pigeon - BirdLife Species Factsheet.http://www.birdlife.org/datazone/ search/species_search.html?action=SpcHTM Details.asp&sid=2754&m=0.[29/11/2005]. Benstead P, Dutson G. 2005. Victoria Crowned- pigeon - BirdLife Species Factsheet.http://www.birdlife.org/datazone/ search/species_search.html?action=SpcHT MDetails.asp&sid=2755&m=0.[29/11/2005] Benstead P, Dutson G. 2005. Southern Crowned- pigeon - BirdLife Species Factsheet.http://www.birdlife.org/datazone/ search/species_search.html?action=SpcHT MDetails.asp&sid=2756&m=0.[29/11/2005] Bretagnolle V, Attie C, Pasquet E. 1998. Cytochrome b-evidence for
validity and phylogenetic relationship of pseudobulweria and Bulweria. The Auk
115(1); 188-195
Clayton DH, Bush SE, Goates BM, Johnson KP. 2003. Host defense
reinforces host- parasite cospeciation.
Proc. Natl. Acad. Sci. USA. 100 (26):15694-15699.
Desjardins, P and R. Morais. 1990. Sequence and gene organization of the chicken
mitochondrial genome. A novel gene order in higher vertebrates.
J.Mol. Biol. 212(4): 599-634.
Duryadi D, 2002. Penentuan Sex Burung-burung Monomorf dengan Penanda Molekuler. Laporan Project Que Grant Tahun Ke-III. Jurusan Biologi.
Bogor. Institut Pertanian Bogor. Johnson KP, Clayton DH. 2000. Nuclear and
Mitochondrial Genes Contain Similar Phylogenetic Signal for Pigeons and Doves (Aves: Columbiformes). Mol. Phylogenet. Evol 14: 141-151.
Kilmaskossu A. 2001. Ekologi Persarangan, Musim Perkembangan dan Kajian Awall Keragamanan Morfogenetik Mambruk Polos (G.cristata). [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Kocher TD et al. 1989. Dynamics of
mitochondrial DNA evolution in animals: Amplifications and sequencing with conserved primers. J. Proc. NAtl. Acad. Sci. USA. 86:6196-6200
Kumar S, Tamura K, and Nei M. 2004. MEGA 3.0: Integrated software for Molecular Evolutionary Genetics Analysis and sequence alignment. Bioinformatics
5:150-163.
Noerdjito M, Maryamto I. 2001. Jenis-jenis Hayati yang Dilindungi Perundang-undangan Indonesia. Cibinong. Balitbang Zoologi Puslitbang Biologi-Lipi dan The Nature Conservacy.
Perrins CM, Middleton ALA. 1985. The Encyclopedia of Birds. New York. Fact on File Inc.
Randi E. 1996. A Mitochondrial cytochrome b phylogeny of the Alectrois partridges [abstrak]. Di dalam. J.Mol. Phyl. Evol. 6
Shapiro B et al. 2002. Flight of the dodo.
Science 295 (5560): 1683.
Stanley SE and Harrison RG. 1999. Cytochrome
b evolution in birds and mamals: An Evaluation of the avian constraint Hypotesis. Mol. Biol. Evol. 16(11): 1575-1585.
Sorenson MD. 2003. Avian mtDNA Primers. Boston University.
Lampiran 1 Peta Penyebaran Goura sp. (http://www.birdlife.org)
1a. Daerah penyebaran Goura cristata
(Western crowned-pigeon).
1c. Daerah penyebaran Goura victoria
Lampiran 2 Hasil perunutan nukleotida Sit b. Goura sp. hasil penelitian, G.victoria (Genbank ), G.cristata (Genbank) dan Gallusgallus sepanjang 382 nt
Gal_gallus
TGAAACACAG GAGTAATCCT CCTCCTCACA CTCATAGCCA CCGCCTTTGT GGGCTATGTT CTCCCATGGG GCCAAATATC ATTCTGAGGG GCCACCGTTA TCACAAACCT ATTCTCAGCA ATTCCCTACA TTGGACACAC CCTAGTAGAG TGAGCCTGAG GGGGATTTTC AGTCGACAAC CCAACCCTTA CCCGATTCTT CGCTTTACAC TTCCTCCTCC CCTTTGCAAT CGCAGGTATT ACTATCATCC ACCTCACCTT CCTACACGAA TCAGGCTCAA ACAACCCCCT AGGCATCTCA TCCGACTCTG ACAAAATTCC ATTTCACCCA TACTACTCCT TCAAAGACAT TCTGGGCTTA ACTCTCATAC TCACCCCATT CC
G.crist_GenB
TGAAACACAG GAGTCATCCT CCTACTAACC CTAATAGCAA CCGCCTTCGT AGGGTATGTC CTACCCTGAG GACAGATATC ATTTTGAGGT GCCACAGTTA TTACCAACCT ATTCTCCGCC ATCCCCTACA TCGGCCAAAC CATTGTCGAA TGAGCCTGAG GTGGCTTCTC AGTAGATAAT CCCACACTAA CACGATTCTT CACCCTCCAC TTCCTCCTCC CCTTCATAAT TGCTGGCCTC ACCATCATCC ACCTCACATT CTTACATGAA TCCGGCTCAA ACAATCCACT AGGCATCTCT TCCAACTGCG ACAAAATCCC ATTCCACCCC TACTTTTCCC TAAAAGACAT TCTTGGCTTC ATACTAATAC TCCTCCCCCT AA
G.crist_m3
TGAAACACAG GAGTCATCCT CCTACTAACC CTAATAGCAA CCGCCTTCGT AGGGTATGTC CTACCCTGAG GACAGATATC ATTTTGAGGT GCCACAGTTA TTACCAACCT ATTCTCCGCC ATCCCCTACA TCGGCCAAAC CATTGTCGAA TGAGCCTGAG GTGGCTTCTC AGTAGATAAT CCCACACTAA CACGATTCTT CACCCTCCAC TTCCTCCTCC CCTTCATAAT TGCTGGCCTC ACCATCATCC ACCTCACATT CTTACATGAA TCCGGCTCAA ACAATCCACT AGGCATCTCT TCCAACTGCG AAAAAATCCC ATTCCACCCC TACTTTTCCC TAAAAGACAT TCTTGGCTTC ATACTAATAC TCCTCCCCCT AA
G.vict_a1
TGAAATACAG GAGTCATCCT CCTACTAACC CTAATAGCAA CCGCCTTCGT AGGGTACGTC CTACCCTGAG GGCAGATATC ATTTTGAGGA GCCACAGTTA TTACCAACCT ATTCCCAGCT ATCCCCTACA TCGGCCAAAC CATTGTCGAA TGAGCCTGAG GCGGCTTCTC AGTAGACAAC CCCACACTAA CACGATTCTC CACCTTCCAT TTCCTCCTCC CCTTCATAAT CGCTGGCCTC ACCATCATCC ACCTCACATT CCTACACGAA TCCGGCTCAA ACAACCCATT AGGCATCTCC TCCAACTGGG ATAAAACCCC ATTCCACCCC TACTTTTCCT CAAAAGACAT CCTTGGCTTC ACACTGATAC TCCTCCCCCT AA
G.vict_GenB
Lampiran 2 Lanjutan
G.scheep_m7
Lampiran 1 Peta Penyebaran Goura sp. (www.birdlife.org)
G.victoria
G. cristata
1b Daerah penyebaran Goura cristata (Western crowned-pigeon).
Lampiran 1 Lanjutan
G. scheepmakeri
G. scheepmakeri
Lampiran 3 Matriks perbedaan nukleotida (diatas diagonal) dan jarak genetik (dibawah diagonal) dari Goura sp. hasil penelitian, G.victoriaGenBank dan
G.cristata(GenBank, spesies famili Columbidae dan Gallus gallus sebagai pembanding
1 2 3 4 5 6 7 8 9 10 11 12
1]Gal.gallus_net
2]G.crist_GenB
3]G.crist_m3
4]G.scheep_m7
5]G.vict_a1
6]G.vict_GenB
7]Did.strig
8]Alec.mdgac_Bnet
9]Pti.supbs_Bnet
10]Col_livia
11]Lep.mgal_Bnet
12]Lep.verful_Bnet
81 82 84 79 82 78 73 75 77 71 71
0.2120 1 15 25 23 42 40 37 44 36 36
0.2147 0.0026 15 25 23 42 41 38 44 37 37
0.2199 0.0393 0.0393 24 23 42 40 38 45 41 40
0.2068 0.0654 0.0654 0.0628 27 45 45 41 45 38 41
0.2147 0.0602 0.0602 0.0602 0.0707 45 42 44 43 42 40
0.2042 0.1099 0.1099 0.1099 0.1178 0.1178 40 44 44 47 47
0.1911 0.1047 0.1073 0.1047 0.1178 0.1099 0.1047 36 39 45 43
0.1963 0.0969 0.0995 0.0995 0.1073 0.1152 0.1152 0.0942 50 44 42
0.2016 0.1152 0.1152 0.1178 0.1178 0.1126 0.1152 0.1021 0.1309 38 42
0.1859 0.0942 0.0969 0.1073 0.0995 0.1099 0.1230 0.1178 0.1152 0.0995 23
Gal.gallus_net TGA AAC ACA GGA GTA ATC CTC CTC CTC ACA CTC ATA GCC ACC GCC TTT GTG GGC TAT GTT CTC CCA TGG [ 69] G.crist_GenB ... ... ... ... ..C ... ... ..A ..A ..C ..A ... ..A ... ... ..C ..A ..G ... ..C ..A ..C ..A [ 69] G.crist_m3 ... ... ... ... ..C ... ... ..A ..A ..C ..A ... ..A ... ... ..C ..A ..G ... ..C ..A ..C ..A [ 69] G.scheep_m7 ... ... ... ... ..C ... ... ..A ..A ..C ..A ... ..A ... ... ..C ..A ..G ..C ..C ..A ..C ..A [ 69] G.vict_a1 ... ..T ... ... ..C ... ... ..A ..A ..C ..A ... ..A ... ... ..C ..A ..G ..C ..C ..A ..C ..A [ 69] G.vict_GenB ... ... ... ... ..C ... ... ..A ..A ..C ..A ... ..A ... ... ..C ..A ..G ... ..C ..A ..C ..A [ 69] Did.strig ... ... ... ... ..C ... ..A ..A ..A ..C ..A ... ..A ..T ... ..C ..A ..A ... ..C ..A ..C ..A [ 69] Alec.mdgac_Bnet ... ... ..G ... ..C ... ... ... ..A ..C ..A ... ..A ..T ... ..C ..C ..A ..C ..C ..A ..C ..A [ 69] Pti.supbs_Bnet ... ... ... ... A.C ..T ... ... ..G ..C ..A ... ..A ... ... ..C ..C ..A ..C ..C ..A ..C ..A [ 69] Col_livia ... ... ... ... ..C G.. ... ..A ..A ..C ..T ... ... ..T ..A ..C ..A ..A ... ..C ..A ..C ..A [ 69] Lep.mgal_Bnet ... ... ... ... ..C ... ... ..A ..A ..C ..A ... ..A ... ... ..C ..A ..A ..C ..C ..G ... ..A [ 69] Lep.verful_Bnet ... ... ... ... ..C ... ... ..A ..A ..C ..A ... ..A ... ... ..C ... ..G ..C ..C ..A ..C ..A [ 69]
Lanjutan Lampiran 4
Gal.gallus_net ACC CTA GTA GAG TGA GCC TGA GGG GGA TTT TCA GTC GAC AAC CCA ACC CTT ACC CGA TTC TTC GCT TTA [207] G.crist_GenB ... A.T ..C ..A ... ... ... ..T ..C ..C ... ..A ..T ..T ..C ..A ..A ..A ... ... ... A.C C.C [207] G.crist_m3 ... A.T ..C ..A ... ... ... ..T ..C ..C ... ..A ..T ..T ..C ..A ..A ..A ... ... ... A.C C.C [207] G.scheep_m7 ... A.T ..C ..A ... ..T ... ..T ..C ..C ... ..A ..T ..T ..C ..A ..A ..A ... ... ... A.C C.C [207] G.vict_a1 ... A.T ..C ..A ... ... ... ..C ..C ..C ... ..A ... ... ..C ..A ..A ..A ... ... .C. A.C ..C [207] G.vict_GenB ... A.C ..C ..A ..G ... ... ..T ..C ..C ... ..A ... ... ..C ..A ..A ..A ... ... ... A.C C.C [207] Did.strig ... A.C ..C ..A ... ..T ... ..A ... ... ... ..A ... ... ..C ..A ..C ..A ... ... ... A.C C.T [207] Alec.mdgac_Bnet ... A.T ..C ..A ... ..T ... ... ..G ... ... ..A ... ... ..C ..A T.A ..A ... ... ... A.C C.T [207] Pti.supbs_Bnet ... ..T ..C ..A ... ..T ... ... ..C ... ... ..A ... ... ..C ..A ..A ..A ... ... ... A.. C.C [207] Col_livia ... ..C ..T ..A ... ... ... ..C ... ... ..C ..A ..T ... ..T ..A T.A ..A ... ... ... A.C C.T [207] Lep.mgal_Bnet ... ... ..T ..A ... ... ... ..C ... ..C ..C ..A ..T ... ..C ..A ..A ..A ... ... ... A.C ..G [207] Lep.verful_Bnet ... ... ..C ..A ... ... ... ..T ... ... ..C ..A ..T ... ..C ..A ..A ..A ... ..T ... A.C ... [207]
Gal.gallus_net TCA AAC AAC CCC CTA GGC ATC TCA TCC GAC TCT GAC AAA ATT CCA TTT CAC CCA TAC TAC TCC TTC AAA [345] G.crist_GenB ... ... ..T ..A ... ... ... ..T ... A.. .GC ... ... ..C ... ..C ... ..C ... .TT ... C.A ... [345] G.crist_m3 ... ... ..T ..A ... ... ... ..T ... A.. .GC ..A ... ..C ... ..C ... ..C ... .TT ... C.A ... [345] G.scheep_m7 ... ... ... ..A ... ... ... ..T ... A.. .GC ..T ... ..C ... ..C ... ..C ... .TT ... C.A ... [345] G.vict_a1 ... ... ... ..A T.. ... ... ..C ... A.. .GG ..T ... ..C ... ..C ... ..C ... .TT ... ..A ... [345] G.vict_GenB ... ... ... ..A T.. ..T ... ..T ... A.. .GC ..T ... ..C ... ..C ... ..C ... .T. ... C.A ... [345] Did.strig ... ... ... ..A ... A.. ... ..C ... A.. .G. ..T ... ..C T.. ..C ... ..C .G. .T. ... C.A ... [345] Alec.mdgac_Bnet ... ... ... ... ... ... ... ..T ... A.. .G. ... ... ..C ... ..C ... ..C ... .T. ... C.A ... [345] Pti.supbs_Bnet ... ... ... ... ... ... ... ..T ... A.. .GC ... ... ..C ... ..C ... ..T ... .TT ..T C.A ... [345] Col_livia ... ... ... ..A ... ... ... A.C ... A.. .GC ..T ... ..C ... ..C ... ..C ... .T. ... C.A ... [345] Lep.mgal_Bnet ... ... ... ..A ... ... ... ..C ... A.. .GC ... ... ..C ... ..C ... ..C ... .T. ... C.A ... [345] Lep.verful_Bnet ... ... ... ..A ... ... ... ..C ... A.. .GC ... ... ..C ... ..C ... ..C ... .T. ..T C.A ... [345]
Lampiran 5 Penjajaran berganda asam amino dari Cyt b parsial (127 aa) G.cristata GenBank, G.victoria GenBank, Goura sp. hasil penelitian, spesies famili Columbidae dan Gallus gallus sebagai pembanding.
Gal.gallus_net WNTGVILLLT LMATAFVGYV LPWGQMSFWG ATVITNLFSA IPYIGHTLVE WAWGGFSVDN PTLTR [ 65] G.crist_GenB ... ... ... ... ...Q.I.. ... ... [ 65] G.crist_m3 ... ... ... ... ...Q.I.. ... ... [ 65] G.scheep_m7 ... ... ... ... ...Q.I.. ... ... [ 65] G.vict_a1 ... ... ... ...P. ...Q.I.. ... ... [ 65] G.vict_GenB ... ... ... ... ...Q.I.. ... ... [ 65] Did.strig ... ... ... ... ...Q.I.. ... ... [ 65] Alec.mdgac_Bnet ... ... ... ... ...Q.I.. ... ... [ 65] Pti.supbs_Bnet ....I... ... ... ... V....Q.... ... ... [ 65] Col_livia ...V.... ... ... ... V....Q.... ... ... [ 65] Lep.mgal_Bnet ... ... ... ... ...Q.... ... ... [ 65] Lep.verful_Bnet ... ... ... ... ...Q.... ... ... [ 65]
UUC10.4(1.70)
Leu UUA 1.4(0.42) UUG 0.0(0.00)
UCC 4.2(3.15)
UCA 2.8(2.10) UCG 0.0(0.00)
UAC 2.4(1.60)
* UAA 0.0(0.00) UAG 0.0(0.00)
UGC 0.8(2.00)
Trp UGA 4.8(1.85) UGG 0.4(0.15) Leu CUU 0.8(0.24)
CUC 7.8(2.36) CUA 9.4(2.85) CUG 0.4(0.12)
Pro CCU 0.0(0.00)
CCC 6.0(2.93) CCA 2.2(1.07) CCG 0.0(0.00)
His CAU 0.6(0.30)
CAC 3.4(1.70) Gln CAA 1.0(1.00) CAG 1.0(1.00)
Arg CGU 0.0(0.00)
CGC 0.0(0.00) CGA 1.0(4.00) CGG 0.0(0.00) Ile AUU 3.0(0.55)
AUC 8.0(1.45) Met AUA 4.4(2.00)
AUG 0.0(0.00)
Thr ACU 0.0(0.00) ACC 5.8(2.07) ACA 5.2(1.86)
ACG 0.2(0.07)
Asn AAU 1.2(0.40) AAC 4.8(1.60) Lys AAA 2.0(2.00)
AAG 0.0(0.00)
Ser AGU 0.0(0.00) AGC 0.0(0.00) * AGA 0.0(0.00)
AGG 0.0(0.00) Val GUU 1.0(0.67)
GUC 3.0(2.00) GUA 2.0(1.33) GUG 0.0(0.00)
Ala GCU 1.4(0.90) GCC 3.8(2.45) GCA 1.0(0.65) GCG 0.0(0.00)
Ala GAU 1.2(0.86) GAC 1.6(1.14) Glu GAA 2.2(2.00) GAG 0.0(0.00)
Gly GGU 1.4(0.51) GGC 6.2(2.25) GGA 2.2(0.80) GGG 1.2(0.44) * Rata-rata kodon 127
angka cetak tebal: besar penggunaan kodon pada Goura sp.
Tabel 3 Kodon penyandi dan asam amino yang disandikan yang mengalami mutasi pada Cyt b parsial dari G.cristata GenBank, G.victoria GenBank, dan Goura sp. hasil penelitian.
Situs ke (kodon) 2 19 24 27 28 30 32 39 40 48 51 52 54 59 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
AAC TAT GGA TCA TTT GGT ACA TCC GCC ATT TGA GCC GGT GAT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..C ... ..C ... ..A ..G ... ..T ... ... ..T ... ... ..T ..C ..G ... ... ..A ... C.A ..T ... ... ... ..C ..C ... ... ... ... ..C ..C ... ... ..T ..C ..G ... ... ..C Situs as.amino 2 19 24 27 28 30 32 39 40 48 51 52 54 59 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
N Y G S F G T S A I W A G D . . . . . . . . . . . P . . . . . . . . Situs ke (kodon) 60 67 69 70 76 77 78 81 88 89 91 95 97 98 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
AAT TTC CTC CAC ATA ATT GCT ACC TTA CAT TCC AAT CTA GGC ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..C ... ... ..C ..T ..C ... ... ..C .C. T.. ..T ... ..C ... ... C.. ..C ... ..C T.. ... ..C ... ... ... C.. ... ..C G.. C.. ..C ... ..C T.. ..T Situs as.amino 60 67 69 70 76 77 78 81 88 89 91 95 97 98 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
N F L H M I A T L H S N L G . . . . . . . . . S F . . . . . . . . L . . A . . . . Situs ke (kodon) 100 103 104 112 114 117 118 120 121 122 125 127 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
TCT TGC GAC TTT CTA ATT CTT TTC ATA CTA CTC CTA ... ... ..A ... ... ... ... ... ... ... ... ... ... ... ..T ... ... ..C T.. ... ... ..G ... T.. ..C ..G ..T ... T.. ..C ... ... .C. ..G ... ... ... ... ..T ..C ... ..C T.C C.. .C. ... ..T ... Situs as.amino 100 103 104 112 114 117 118 120 121 122 125 127 G.crist_GenB
G.crist_m3 G.scheep_m7 G.vict_a1 G.vict_GenB
Ordo : Columbiformes Famili : Phasianidae Spesies : Gallus gallus
Nama Umum: Chicken
Ordo : Columbiformes Famili : Columbidae
Spesies : Alectroenas madagascarensis
Nama umum: Madagascar blue-pigeon
Ordo : Columbiformes Famili : Columbidae
Spesies : Didunculus strigirostris
Nama umum: Tooth-billed pigeon
Ordo : Columbiformes Famili : Columbidae Spesies : Columba livia
Lanjutan Lampiran 7
Ordo : Columbiformes Famili : Columbidae Spesies : Leptotila verreauxi fulviventri
Nama umum: White-tipped doves
Ordo : Columbiformes Famili : Columbidae Spesies : Ptilinopus superbus
Nama umum: Fruit-doves
KERAGAMAN GENETIK CYTOCHROME B
PADA BURUNG MAMBRUK (Goura sp.)
Oleh:
Lasriama Siahaan G04400032
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
ABSTRAK
LASRIAMA SIAHAAN. Keragaman Genetik Cytochrome b pada Burung Mambruk (Goura sp.). Dibimbing oleh DEDY DURYADI SOLIHIN dan DJOKO WALUYO.
Burung Mambruk (Goura sp.) merupakan satwa endemik Indonesia dengan status “Vulnerable” yang tersebar di daerah Pulau Irian Jaya dan beberapa daerah Papua New Guinea. Usaha konservasi terhadap spesies ini akan berhasil jika karakteristik morfologi, keragaman molekuler dan genetik dapat diketahui dengan pasti. Tujuan penelitian ini untuk menganalisis keragaman genetik cytochrome b parsial dengan metode Polimerase Chain Reaction. Hasil perunutan dari amplifikasi pasangan primer M101 dan M102 pada cytochrome b parsial sepanjang 382 nukleotida (menyandikan 127 asam amino) disejajarkan (multiple aligment) dengan bantuan perangkat lunak Genetyx-Win versi 3.0 dan Clustal-X, selanjutnya dianalisis dengan program MEGA versi 3.0.
Hasil analisis dari 382 nukleotida yang dibandingkan terdapat 42 situs nukleotida yang beragam dengan rata-rata kejadian substitusi transisi 0.05 dan rata-rata substitusi transversi 0.01. Perubahan bersifat non-sinonimus 7.08% (10 situs asam amino) dan perubahan bersifat sebesar 23.62% (30 situs asam amino). Jarak genetik nukleotida cytochrome b (metode p-distance) didapat bahwa nilai paling kecil adalah 0.26% dan nilai yang paling besar 7.07% dengan rata-rata sebesar 5.26%. Hasil rekonstruksi filogenetik dengan metode Neighbor Joining menunjukkan bahwa G.cristata lebih berkerabat dekat dengan G.scheepmakeri daripada G.victoria.
Kata kunci: Goura sp., Cytochrome b, substitusi, perubahan
ABSTRACT
LASRIAMA SIAHAAN. Genetic Diversity of Cytochrome b of Crowned-pigeons (Goura sp.), Supervised by DEDY DURYADI SOLIHIN and DJOKO WALUYO.
Crowned-pigeon (Goura sp.) was one of endemic animal in Indonesia with “vulnerable" status which spread at Papua New Guinea and Irian Jaya Island. Conservation effort to this species will succeed if morphology characteristic and genetic diversity be known. The main purpose of this research is to analyze nucleotide variability of cytochrome b partial using Polymerase Chain Reaction method. The PCR product amplified by primer M101 and M102, then multiple aligment using Genetyx-Win 3.0™ and Clustal-X™, furthermore analyzed with MEGA 3.0™.
The result of analyse from 382 nucleotide of cytochrome b partial encoding 127 amino acid. Findings 42 sites of nucleotide variable with average of transitional substitutions 0.05 and average transversions substitutions is 0.01. Value non-synonymous mutations is 7.08% (10 amino acid sites) whereas synonymus mutations is 23.62% (30 amino acid sites). Value of genetic distance of nucleotide cytochrome b (method p-distance) range from 0.26% - 7.07% with average 5.26%. Filogenetic reconstruction using Neighbor Joining method indicate that G.cristata were closer to G.scheepmakeri c