MOCHAMAD TAUFIQ RIDWAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Cecropia peltata L. di
Karst Gunung Cibodas, Bogor: Derajat Invasi, Asosiasi Spesies Diagnostik, dan Manfaatnya bagi Manusia serta Lingkungan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Oktober 2016
Mochamad Taufiq Ridwan
RINGKASAN
MOCHAMAD TAUFIQ RIDWAN. Cecropia peltata L. di Karst Gunung
Cibodas, Bogor: Derajat Invasi, Asosiasi Spesies Diagnostik, dan Manfaatnya bagi Manusia serta Lingkungan. Dibimbing oleh MUHADIONO dan IWAN HILWAN.
Cecropia peltata termasuk spesies asing invasif yang penting untuk
dipelajari. Tujuan penelitian ini yaitu untuk menguraikan: 1) derajat invasi C. peltata dan pengaruhnya terhadap vegetasi kawasan karst Gunung Cibodas, 2)
spesies diagnostik lokasi terinvasi C. peltata, 3) spesies yang mampu berasosiasi
dengan C. peltata, 4) perbandingan jumlah artikel yang meneliti C. peltata dengan
spesies lain, 5) perbandingan manfaat dan kerugian yang ditimbulkan C. peltata.
Penelitian ini dilaksanakan pada Maret-Mei 2015 di Kawasan Karst Gunung Cibodas, Kecamatan Ciampea, Kabupaten Bogor. Pencuplikan data vegetasi dan tanah dilakukan secara purposive mempertimbangkan faktor topografi (punggung
dan lereng bukit) dan kondisi (terinvasi dan tak terinvasi). Teknik pencuplikan vegetasi menggunakan kombinasi transek dan plot bersarang berukuran 400 (pohon), 100 (tiang), 25 (pancang), 4 m2 (tumbuhan bawah dan semai).
Derajat invasi C. peltata diestimasi dari kerapatan dan penutupan di
masing-masing lokasi, sedangkan pengaruhnya terhadap vegetasi dianalisis dari keanekaragaman alfa (jumlah spesies, heterogenitas/indeks Shannon-wiener, dan kemerataan/indeks Pielou), keanekaragaman beta (level plot dan transek), kerapatan (individu ha-1) dan penutupan (m2 ha-1). Spesies diagnostik dideteksi
dengan analisis koefisien phi, sedangkan asosiasinya dengan C. peltata dianalisis
dengan khi kuadrat. Perbandingan manfaat dan kerugian yang ditimbulkan oleh C. peltata dilakukan dengan analisis sainstometri. Kutipan relatif dan indeks h
digunakan sebagai peubah bebas dalam analisis sainstometri.
Lokasi terinvasi C. peltata ditandai dengan kerapatan dan penutupan yang
lebih tinggi dibanding lokasi tak terinvasi. Kerapatan pancang, tiang dan pohon C. peltata mencapai kisaran 71-1368, 128-649, dan 42-98 individu ha-1, sedangkan
penutupan tiang dan pohon mencapai 2.5-10.69 dan 3.08-9.10 m2 ha-1. Invasi C.
peltata menurunkan keanekaragaman alfa dan beta tumbuhan bawah, semai dan
pancang vegetasi.
Spesies diagnostik lokasi terinvasi terdiri dari delapan spesies tumbuhan bawah (Clidemia hirta, Cryota mitis, Cyclosorus sp, Lygodium circinatum, Piper aduncum, Pseudophegopteris sp, S. wildenowii dan Thelypteris sp), dua semai
(Calliandra calothyrsus dan Croton triglium), dan tiga pancang (C. calothyrsus, Gyrinops sp, dan Cinnamomum iners). Dari 12 spesies tersebut, hanya C. siamea
yang berasosiasi dengan C. peltata. Invasi C. peltata lebih sedikit dilaporkan
dibanding spesies asing invasif lainnya. Selain itu, manfaat yang diperoleh dari C. peltata, terutama obat-obatan, lebih tinggi dibanding kerugian yang ditimbulkan.
Dengan demikian, populasi C. peltata yang tinggi menjadi peluang
pemanfaatannya bagi manusia dan lingkungan terutama di bidang medis.
Kata kunci : analisis sainstometri, Cecropia peltata, derajat invasi,
Advantages for Human and Environment. Supervised by MUHADIONO and IWAN HILWAN.
Cecropia peltata is important invasive alien species that should be studied.
The aims of this study were to determine : 1) invasion degree of C. peltata and its
effect on vegetation at karst area of Gunung Cibodas, 2) diagnostic species of invaded site, 3) species that could associate with C. peltata, 4) comparison of
scientific article number that studied invasion of C. peltata and other species, 5)
comparison of advantages and disadvantages of C. peltata. This study was
conducted on March-May 2015 at Karst Area of Gunung Cibodas Hill, Ciampea Subdistrict, Bogor District. Vegetation and soil data was purposively collected, on ridge and slope of the hill, and invaded and uninvaded of the site. A combination of line transect and plot with varying size : 400 (tree), 100 (pole), 25 (sapling), and 4 m2 (understory and seedling) were used to sample the vegetation data.
Invsion degree of C. peltata was estimated by its density and coverage in
each site, while its effect on vegetation was analyzed by alpha diversity (species richness, heterogeneity/Shannon-wiener index, evenness/Pielou index), beta diversity (plot and transect level), density (individual ha-1) and coverage (m2 ha-1).
Diagnostic species was detected by phi coefficient analysis, while its association with C. peltata was determined by chi-square analysis. Advantage and
disadvantage of C. peltata was compared using scientometric analysis. Relative
citation and h-index were used as independent variables in scientometric analysis.
Invaded site by C. peltata was identified by its higher density and coverage
than uninvaded site. Density range of sapling, pole, and tree of C. peltata were
71-1368, 128-649 and 42-98 individual ha-1, respectively. In addition, coverage
range of pole and tree were 2.5-10.69 and 3.08-9.10 m2 ha-1, respectively. C.
peltata invasion decreased alpha and beta diversity of understories, seedlings and
saplings.
Eight understory species (Clidemia hirta, Cryota mitis, Cyclosorus sp, Lygodium circinatum, Piper aduncum, Pseudophegopteris sp, S. wildenowii and Thelypteris sp) were identified as diagnostic species of invaded site. In addition,
two seedling species (Calliandra calothyrsus and Croton trigilium) and three
sapling species (C. calothyrsus, Gyrinops sp, and Cinnamomum iners), were also
identified as diagnostic species of invaded site. Among 12 diagnostic species, C. siamea was the only species associated with C. peltata. Invasion of C. peltata on
native vegetation was less reported by a few scientists in scientific articles than another invasive alien species. In addition, the advantages obtained from C. peltata to human and environment, especially medicines, were more reported by
many scientists in scientific articles than the disadvantages like invasion risk and parasites facilitation. Furthermore, high population of C. peltata in Karst Area
Gunung Cibodas Hill has tha advantages for human and environment, especially to be utilized in medicine.
Keywords: Cecropia peltata, diagnostic species, diversity, invasion degree,
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Tumbuhan
Cecropia peltata
L. DI KARST GUNUNG CIBODAS, BOGOR:
DERAJAT INVASI, ASOSIASI SPESIES DIAGNOSTIK, DAN
MANFAATNYA BAGI MANUSIA SERTA LINGKUNGAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Judul Tesis : Cecropia peltata L. di Karst Gunung Cibodas, Bogor: Derajat Invasi,
Asosiasi Spesies Diagnostik, dan Manfaatnya bagi Manusia serta Lingkungan
Nama : Mochamad Taufiq Ridwan NIM : G353130361
Disetujui oleh
Komisi Pembimbing
Dr Ir Muhadiono, MSc Ketua
Dr Ir Iwan Hilwan, MS Anggota
Diketahui oleh
Ketua Program Studi Biologi Tumbuhan
Dr Ir Miftahudin, MSi
Dekan Sekolah Pascasarjana
PRAKATA
Segala puji hanya milik Allah yang memberi segala karunia-Nya sehingga tesis yang berjudul “Cecropia peltata L. di Karst Gunung Cibodas, Bogor: Derajat
Invasi, Asosiasi Spesies Diagnostik, dan Manfaatnya bagi Manusia serta Lingkungan” ilmiah ini dapat diselesaikan. Tesis ini disusun berdasarkan hasil penelitian di kawasan Karst Gunung Cibodas, Kecamatan Ciampea, Kabupaten Bogor.
Penulis mengucapkan terima kasih sebesar-besarnya kepada dosen pembimbing yaitu Dr Ir Muhadiono MSc dan Dr Ir Iwan Hilwan MS. Di samping itu, penghargaan penulis sampaikan kepada Tb Dedi Supriadi dari Perum Perhutani KPH Bogor atas bantuan berupa informasi mengenai lokasi penelitian dan Dr Julisasi Tri Hadiah dari Pusat Konservasi Tumbuhan Kebun Raya Bogor atas bantuan identifikasi spesimen C. peltata. Penulis juga mengucapkan terima
kasih kepada Dr Sulistijorini MSi sebagai komisi penguji ujian tesis atas saran dan masukannya. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, serta seluruh keluarga, atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Oktober 2016
DAFTAR ISI
Spesies Pesaing bagi Spesies Asing Invasif 7
Spesies Diagnostik 8
Analisis Sainstometri 8
3 METODE 10
Pengaruh C. peltata terhadap Vegetasi Karst Cibodas 10
Deskripsi Lokasi 10
Pencuplikan Vegetasi dan Tanah 11
Analisis Data 11
Analisis Sainstometri C. peltata 16
3 HASIL 17
Pengaruh Invasi C. peltata terhadap Vegetasi Karst Gunung Cibodas 17
Struktur Vegetasi 17
Karakteristik Tanah 18
Pengaruh Invasi C. peltata 18
Derajat Invasi C. peltata 20
Pendugaan Karakter Floristik dengan Faktor Tanah 21
Komposisi Spesies setiap Strata 21
Spesies Diagnostik setiap Strata 22
Asosiasi C. peltata dengan Tumbuhan Lainnya 24
Analisis Sainstometri C. peltata 24
4 PEMBAHASAN 27
Derajat Invasi C. peltata dan Pengaruhnya terhadap Vegetasi 27
Asosiasi Spesies Diagnostik dengan C. peltata 28
DAFTAR TABEL
1. Kondisi iklim, kelerengan dan bukaan tajuk di lokasi penelitian
(rerata±galat baku) pada 2012-2014 10
2. Kontingensi 2 × 2 untuk menentukan komponen koefisien phi 14 3. Kontingensi 2 × 2 untuk menentukan komponen asosiasi 15 4. Indeks keanekaragaman dan kemerataan spesies setiap strata dan spesies
dominan di Kawasan Karst Gunung Cibodas 17
5. Keanekaragaman beta setiap level pencuplikan (plot, transek, dan lokasi) dan kerapatan. Huruf berbeda dalam kolom yang sama menunjukkan perbedaan nyata di P<0.05 uji lanjut Duncan 18
6. Penduga keanekearagaman dan kerapatan tumbuhan bawah dan semai
berdasarkan analisis regresi 21
7. Spesies diagnostik (dengan nilai koefisien phi) setiap strata di setiap
lokasi (nyata pada P<0.05) 23
DAFTAR GAMBAR
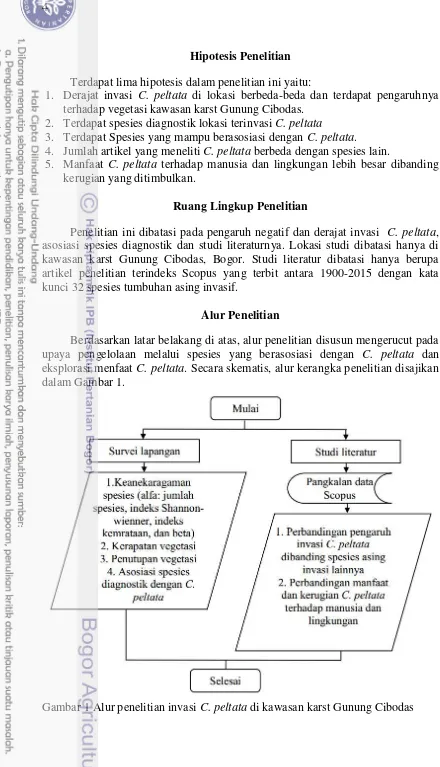
1. Alur penelitian invasi C. peltata di kawasan karst Gunung Cibodas 4
2. Kondisi vegetasi lereng selatan yang didominasi oleh C. peltata 10
3. Ilustrasi transek dan plot pencuplikan. a : 20 x 20 m untuk strata pohon diameter >20 cm, b : 10 x 10 m untuk tiang (diameter 10-20 cm), c : 5 x 5 m untuk pancang (diameter <10 cm dan tinggi >1.5 m), dan d: 2 x 2 m2 untuk semai (tinggi <1.5 m) dan tumbuhan bawah (herba dan semak).
: titik pencuplikan tanah 11
4. Interpolasi (garis penuh) dan ekstrapolasi (garis putus-putus) jumlah spesies (A); penutupan (B). Po: pohon; Ti: tiang; Pa: pancang; Se: semai; Tb: tumbuhan bawah. Huruf berbeda menunjukkan perbedaan nyata di setiap strata di P<0.05 uji lanjut Duncan; td: tidak dihitung 17
5. Rerata karakter fisika dan kimia tanah setiap lokasi. P: punggung, LU: lereng utara, LS: lereng selatan. Huruf berbeda menunjukkan perbedaan
nyata di P<0.05 uji lanjut Duncan 18
6. Rerata keanekaragaman alfa berdasarkan faktor topografi (□) dan invasi (■). L: lereng, P: punggung, T: terinvasi, TT: tak terinvasi, tn: tidak nyata, S: jumlah spesies, H’: indeks Shannon-wiener, E: indeks kemerataan. Perbedaan nyata antar perlakuan berdasarkan uji Z pada †P<0.1, *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001 19 7. Pengaruh invasi terhadap keanekaragaman beta, penutupan (m2 ha-1) dan
kerapatan (individu x 103 ha-1). Huruf berbeda menunjukkan perbedaan
nyata antar lokasi berdasarkan uji lanjut Duncan pada P<0.05. P:
punggung, LU: lereng utara, LS: lereng selatan 20 8. Rerata kerapatan (individu ha-1) dan penutupan (m2 ha-1) C. peltata. T:
terinvasi C. peltata, TT: tak terinvasi. Perbedaan nyata antar lokasi
berdasarkan uji T pada †P<0.1, *P<0.05, **P<0.01. tn: tidak nyata 21 9. Komposisi spesies setiap strata berdasarkan analisis ordinasi bipolar 22 10.Ilustrasi asosiasi positif antara C. calothyrsus dengan C. peltata, dan
asosiasi negatif antara C. siamea dan C. peltata. Keduanya nyata
berasosiasi pada P<0.05 uji khi kuadrat 24
11.Laju kutipan C. peltata dan L. camara per tahun (A). Total artikel dan
kutipan C. peltata 59 dan 1388, sedangkan L. camara 1066 dan 13234.
Persentase kutipan manfaat dan kerugian C. peltata. Total jumlah kutipan
DAFTAR LAMPIRAN
1. Korelasi (R) dan koefisien determinasi (adjusted R2) antara indeks nilai penting (INP) dan koefisien phi (Ф) tiap strata dan lokasi. Cetak tebal
menunjukkan taraf nyata di P<0.05 38
2. Rerata karakter fisika dan kimia tanah ketiga lokasi 39 3. Pendugaan keanekaragaman spesies dan kerapatan 40 4. Data INP dan komponennya, SDR, dan Phi tiap lokasi (cetak tebal nyata
di P<0.05) 41
5. Korelasi (R) dan pendugaan indeks h oleh jumlah artikel dan kutipan
1
PENDAHULUAN
Latar Belakang
Invasi tumbuhan asing berdampak negatif terhadap vegetasi dengan memengaruhi dinamika keanekaragaman tumbuhan (Mayor et al. 2012;
MacDougall et al. 2013). Hal tersebut terjadi akibat adanya dominasi spesies
asing invasif yang menekan spesies lainnya. Di samping itu, gangguan alami maupun yang disebabkan oleh manusia juga dapat memfasilitasi proses invasi oleh spesies asing terhadap ekosistem (Paudel & Battaglia 2015), dan interaksi keduanya menurunkan kekayaan spesies asli (Vellend et al. 2013).
Di Indonesia, ditemukan 1936 spesies tumbuhan asing invasif dan 17% dari spesies yang tercatat merupakan gulma (Tjitrosoedirdjo 2005). Beberapa spesies asing invasif tersebut dapat menimbulkan resiko invasi dan berdampak negatif terhadap ekosistem (Tjitrosoedirdjo 2005, Junaedi & Dodo 2014), misalnya Acacia nilotica. Spesies asing invasif ini menurunkan keanekaragaman
spesies di Taman Nasional Baluran (Caesariantika et al. 2011). Spesies ini juga
menyebabkan ketersediaan sumber makanan bagi herbivora berkurang.
Di lokasi lainnya, Acacia decurrens menjadi spesies dominan di Taman
Nasional Gunung Merapi, setelah terjadinya erupsi pada 2006 dan 2010 (Suryawan et al. 2015). Invasi tidak hanya terjadi oleh spesies asing invasif
berhabitus pohon seperti halnya Acacia, namun juga tumbuhan pemanjat seperti
mantangan (Merremia peltata). Invasi mantangan di Taman Nasional Bukit
Barisan Selatan menyebabkan turunnya keanekaragaman spesies tumbuhan bawah dan semai (Master et al. 2012).
Cecropia peltata yang dikenal di Jawa Barat dengan nama
Kikopong/Kibolong, (Sheil & Padmanaba 2011a) yang merupakan tumbuhan asing asal neotropis, termasuk daftar 100 spesies asing invasif versi IUCN (Lowe
et al. 2000). Spesies ini dilaporkan menyebar luas dan menggantikan Musanga cecropioides, spesies asli di Kamerun (McKey 1988). Spesies ini juga
mendominasi bagian utara dan timur Singapura setelah diintroduksi pada 1902 (Lok et al. 2010). Cecropia peltata diintroduksi di Indonesia pada 1943, dan
menjadi invasif di Jawa Barat (Webber et al. 2011; Conn et al. 2012), serta
menyebar dengan kisaran lebih dari 1290 km2 (Sheil & Padmanaba 2011a).
Cecropia peltata tersebar di dataran rendah (Jakarta) hingga dataran tinggi
(Gunung Gede) dan berbagai tipe tanah termasuk kawasan karst (Sheil & Padmanaba 2011a).
Tanah karst yang khas menghasilkan flora unik dan endemik (Clements et al. 2006). Saat ini kawasan karst terancam rusak akibat gangguan manusia.
Praktik eksploitasi sumber daya karst menurunkan populasi taksa endemik bahkan dapat menyebabkan kepunahan. Selain itu, kawasan karst merupakan ekosistem yang mengalami proses pemulihan sangat lambat setelah mengalami gangguan (Chen et al. 2012).
dan Melastoma ditemukan melimpah di karst Gunung Cibodas. Dua spesies Cecropia juga ditemukan di kawasan karst ini, yaitu C. palmata dan C. sundaica.
Kendati demikian, kedua spesies ini tidak menunjukkan aktivitas invasi (Satyanti & Kusuma 2010). Selain itu, kedua spesies tersebut tidak diketahui informasi kehadirannya saat ini (Conn et al. 2012).
Cecropia palmata diintroduksi dari Royal Botanic Gardens Kew dan tidak
dapat hidup lama setelah ditanam di Kebun Raya Bogor pada 1904, sedangkan C. sundaica tidak diketahui eksistensinya dalam pangkalan data seperti International Plant Name index (http://www.ipni.org). Hanya terdapat dua spesies Cecropia
yang eksis di Indonesia hingga saat ini yaitu C. peltata dan C. pachystaschya
dengan sinonim C. adenopus dan C. schiedeana (Sheil & Padmanaba 2011a,
Webber et al. 2011, Conn et al. 2012). Meskipun identifikasi spesies dari genus Cecropia bersifat problematis (Richardson & Rejmanèk 2011), beberapa studi
menunjukkan eksistensi C. peltata dibanding C. pachystaschya di sekitar Bogor
(Rachman & Balfas 1987 dalam Conn et al. 2012, Sheil & Padmanaba 2011,
Conn et al. 2012). Oleh karena itu, persebaran dan potensi invasi Cecropia di
sekitar Bogor, termasuk di antaranya Karst Gunung Cibodas, kemungkinan besar disebabkan oleh C. peltata.
Spesies ini memiliki karakteristik sebagai spesies asing invasif terutama potensi reproduksinya (Webber et al. 2011). Jutaan biji dapat dihasilkan pertahun
pemencaran biji hingga radius tiga kilometer dari suatu individu yang telah dewasa, seed bank yang awet hingga lima tahun, dan kemampuan untuk
membentuk belukar dan memperbanyak diri secara vegetatif dari batang.
Cecropia peltata dapat dikategorikan sebagai spesies asing invasif yang tersebar
luas dan mendominasi vegetasi setempat (Colautti & MacIsaac 2004, Conn et al.
2012). Spesies asing ini juga dilaporkan menggantikan spesies asli di berbagai daerah, namun derajat ancaman dan dampak invasinya terhadap vegetasi di Jawa Barat masih belum jelas diketahui terutama di kawasan karst (Sheil & Padmanaba 2011a).
Invasi C. peltata dan ancamannya terhadap ekosistem masih
diperdebatkan (Sheil & Padmanaba 2011a; Webber et al. 2011). Sheil dan
Padmanaba (2011b) berargumen bahwa bukti kerugian yang ditimbulkan oleh
Cecropia tak meyakinkan. Mereka tidak menemukan bukti jelas Cecropia
menyebabkan kerugian berdasarkan kriteria yang mereka tentukan (Sheil & Padmanaba 2011a). Walaupun C. peltata ditemukan (satu individu) di Taman
Nasional Gunung Gede Pangrango, spesies ini belum menginvasi hutan alam. Selain itu, Sheil dan Padmanaba (2011a) gagal mengidentifikasi ancaman C. peltata di samping menggantikan spesies asli (misal McKey 1988), dan penduduk
lokal belum menganggap spesies ini sebagai ancaman. Oleh karena itu, studi mengenai invasi C. peltata di Jawa Barat perlu dilakukan agar dapat menangani
dampak negatif yang ditimbulkan (Webber et al. 2011).
Salah satu upaya pengendalian invasi spesies asing invasif adalah pemanfaatan musuh alami ataupun pesaing. Tumbuhan invasif merupakan pesaing yang lebih baik dibanding tumbuhan non-invasif (Vilà & Weiner 2004). Oleh karena itu, diperlukan spesies non-invasif yang mampu hidup berasosiasi dengan spesies asing invasif dan berpotensi menekan pertumbuhannya.
penutupan tanah, mempercepat suksesi, habitat bagi frugivor (Sheil & Padmanaba 2011a dan referensi di dalamnya). Meskipun demikian, perbandingan antara resiko invasi dan manfaat C. peltata masih belum diketahui dan studi sainstometri
mengenai C. peltata perlu dilakukan untuk menguraikan potensi resiko dan
manfaatnya dari literatur ilmiah.
Perumusan Masalah
Fokus penelitian ini yaitu proses invasi C. peltata memengaruhi vegetasi
kawasan karst Gunung Cibodas dari beberapa aspek yakni derajat invasinya, asosiasi spesies diagostik dengan C. peltata, serta manfaatnya bagi manusia dan
lingkungan. Selain itu, eksistensi dan performansi C. peltata di lokasi penelitian
dapat menjadi acuan dalam penentuan derajat invasi dan pengaruhnya terhadap vegetasi. Kendati demikian, dari sekian banyak tumbuhan yang ada di lokasi penelitian, diduga ada beberapa spesies yang mampu berasosiasi dengan C. peltata, terutama spesies diagnostik lokasi terinvasi. Dengan demikian terdapat
lima permasalahan yang muncul yaitu:
1. Bagaimana kondisi derajat invasi C. peltata dan apa pengaruhnya terhadap
vegetasi kawasan karst Gunung Cibodas?
2. Apa saja spesies diagnostik di lokasi terinvasi C. peltata?
3. Spesies apa saja yang mampu berasosiasi dengan C. peltata?
4. Bagaimana perbandingan jumlah artikel yang meneliti C. peltata dengan spesies lain?
5. Bagaimana perbandingan manfaat dan kerugian yang ditimbulkan C. peltata
terhadap manusia dan lingkungan?
Tujuan Penelitian Penelitian ini bertujuan untuk menguraikan:
1. Derajat invasi C. peltata dan pengaruhnya terhadap vegetasi kawasan karst
Gunung Cibodas.
2. Spesies diagnostik lokasi terinvasi C. peltata
3. Spesies yang mampu berasosiasi dengan C. peltata.
4. Perbandingan jumlah artikel yang meneliti C. peltata dengan spesies lain.
5. Perbandingan manfaat dan kerugian yang ditimbulkan C. peltata terhadap
manusia dan lingkungan.
Hipotesis Penelitian Terdapat lima hipotesis dalam penelitian ini yaitu:
1. Derajat invasi C. peltata di lokasi berbeda-beda dan terdapat pengaruhnya
terhadap vegetasi kawasan karst Gunung Cibodas. 2. Terdapat spesies diagnostik lokasi terinvasi C. peltata
3. Terdapat Spesies yang mampu berasosiasi dengan C. peltata.
4. Jumlah artikel yang meneliti C. peltata berbeda dengan spesies lain.
5. Manfaat C. peltata terhadap manusia dan lingkungan lebih besar dibanding
kerugian yang ditimbulkan.
Ruang Lingkup Penelitian
Penelitian ini dibatasi pada pengaruh negatif dan derajat invasi C. peltata,
asosiasi spesies diagnostik dan studi literaturnya. Lokasi studi dibatasi hanya di kawasan karst Gunung Cibodas, Bogor. Studi literatur dibatasi hanya berupa artikel penelitian terindeks Scopus yang terbit antara 1900-2015 dengan kata kunci 32 spesies tumbuhan asing invasif.
Alur Penelitian
Berdasarkan latar belakang di atas, alur penelitian disusun mengerucut pada upaya pengelolaan melalui spesies yang berasosiasi dengan C. peltata dan
eksplorasi menfaat C. peltata. Secara skematis, alur kerangka penelitian disajikan
dalam Gambar 1.
2 TINJAUAN PUSTAKA
Definisi Spesies Asing Invasif
Spesies asing invasif, baik berupa introduksi antar daerah/pulau maupun yang berasal dari negara lain telah sejak lama diperkirakan menjadi salh satu penyebab yang cukup berpengaruh terhadap penurunan kekayaan keanekaragaman hayati. Menurut UN-CBD (The United Nations Convention on Biological Diversity), spesies asing invasif diartikan sebagai jenis introduksi
dan/atau penyebarannya di luar tempat penyebaran alaminya, baik dahulu maupun saat ini, mengganggu atau mengancam keanekaragaman hayati. Organisme ini bisa berupa hewan, tumbuhan, jamur dan miroorganisme. Introduksi spesies ini, selain mengancam sistem ekologi, pada gilirannya juga akan menyebabkan kerugian ekonomi yang tidak sedikit.secara langsung introduksi ini lebih banyak berpengaruh terhadap spesies dan ekosistem. Karena sifatnya yang mengalami pertumbuhan (growing), pada umumnya dampak yang muncul tidak dapat langsung terlihat atau disadari dan baru menjadi perhatian setelah timbulnya gangguan akibat penyebaran yang cepat dan menekan pertumbuhan populasi spesies lokal (Radiansyah et al. 2015).
Penyebaran spesies asing invasif dapat merubah seluruh sistem pada suatu ekosistem, sepertisistem hidrologi, siklus makanan dan proses ekosistem lainnya. Sebagai contoh introduksi Acacia nilotica, spesies asli Afrika, pertama kali
ditanam sebagai sekat bakar di Taman Nasional Baluran pada 1969 telah berubah menjadi spesies invasif. Dalam perkembangannya, spesies pohon tersebut tumbuh dan menyebar sangat cepat dan mendominasi savana yang merupakan kawasan konservasi banteng (Bos javanicus). Penyebaran yang sangat cepat dan masih sulit
untuk ditangani ini mengakibatkan semakin sempitnya wilayah padang pengembalaan banteng sehingga menekan pertumbuhan spesies tumbuhan pakan dan populasi spesies dilindungi ini serta hewan lainnya yang berasosiasi dengan savanna tersebut (Radiansyah et al. 2015).
Penyebaran spesies asing terjadi melalui berbagai cara, baik sengaja maupun tidak. Secara sengaja. Penyebaran dilakukan melalui perdagangan untuk berbagai kepentingan, pertukaran jenis antar negara, introduksi untuk kepentingan tertentu, misalnya spesies cepat tumbuh untuk reklamasi lahan, optimalisasi pemanfaatan makanan di suatu perairan, serta untuk kepentingan kepariwisataan. Secara tidak sengaja, penyebaran dapat terjadi karena adanya benih yang terbawa oleh manusia atau tumbuhan dan hewan melalui transportasi darat, laut maupun udara. Beberapa penyebab terjadinya introduksi spesies asing invasif antara lain: 1. Perdagangan. Introduksi tumbuhan dan/atau hewan merupakan bisnis besar
yang memanfaatkan kecenderungan sifat manusia yang menyukai hal-hal yang baru dan unik sehingga mereka mengintroduksi hewan atau tumbuhan yang belum pernah dilihat atau dikenal sebelumnya (asing).
Manipulasi ekosistem. Introduksi dilakukan ketika terjadi serangan organisme pengganggu di suatu ekosistem tertentu sehingga diperlukan musuh alami (predator) untuk menekan pertumbuhannya (Radiansyah et al.
2015)
Perhatian terhadap spesies asing invasif ini meningkat dengan disepakatinya UN-CBD oleh sejumlah besar negara di dunia, dalam Konferensi Tingkat Tinggi Bumi di Rio de Janeiro, Brazil, pada 3-14 Juni 1992, termasuk Indonesia yang kaya akan keanekaragaman hayati. Konvensi ini diharapkan dapat meningkatkan kerja sama internasional di bidang ilmu pengetahuan dan teknologi guna kepentingan generasi sekarang dan yang akan datang dalam rangka melestarikan keanekaragaman hayati dan memanfaatkan setiap unsurnya secara berkelanjutan. UN-CBD merupakan perjanjian kerjasama internasional yang bersifat mengikat memiliki tiga tujuan yaitu: 1) konservasi keanekaragaman hayati, 2) pemanfaatan secara lestari dari komponen-komponennya, 3) pembagian secara adil dan wajar dari manfaat yang dihasilkan oleh sumberdaya genetik. Oleh karena itu, tujuannya adalah untuk membangun strategi nasional dalam upaya konservasi dan memanfaatkan keanekaragaman hayati secara lestari, yang sering juga dipandang sebagai dokumen pembangunan berkelanjutan (Radiansyah et al. 2015).
Proses dan Penyebab Penyebaran
Pada dasarnya, secara alami tumbuhan dan hewan melakukan penyebaran ke daerah sekitarnya. Penyebaran tersebut disebabkan karena adanya proses pergerakan dan perkembangbiakan spesies tumbuhan, hewan serta adanya proses alam (geologi/vulkanik). Proses alam ini mengakibatkan terjadinya isolasi dari suatu atau beberapa ekosistem, termasuk spesies tumbuhan dan hewan yang hidup di dalamnya. Kondisi geografi Indonesia yang merupakan kepulauan telah membentuk berbagai jenis ekosistem yang berkembang secara unik dan membentuk keseimbangan baru yang berbeda satu dengan lainnya dan juga dengan ekosistem asalnya. Perkembangan dan penyebaran tumbuhan dan hewan menjadi terbatas karena adanya hambatan (barrier) alami seperti laut, sungai, lembah dan pegunungan telah membantu perkembangan ekosistem untuk mencapai suksesi klimaks. Dalam banyak kasus hambatan ini dapat diatasi oleh spesies hewan yang memiliki kemampuan daya jelajah atau mobilitas tinggi sehingga beberapa spesies tumbuhan, dapat menyebar sampai keluar habitat aslinya (Radiansyah et al. 2015).
suatu area yang pada awalnya tidak disadari dapat merusak ekosistem daerah yang bersangkutan yang kelak dapat berdampak lebih besar terhadap lingkungan, kesehatan, sosial dan ekonomi. Dalam beberapa laporandisampaikan bahwa spesies asing invasif dapat menimbulkan kerusakan lingkungan dan ekonomi besar yang diperburuk oleh terjadinya perubahan iklim, polusi, kehilangan habitat serta gangguan yang disebabkan oleh manusia (Radiansyah et al. 2015).
Pemahaman terhadap pola penyebaran dan tingkat invasi oleh spesies tumbuhan dan hewan akan sangat berpengaruh terhadap upaya pengelolaannya. Di samping itu penyebaran dan besaran dapak yang telah ditimbulkannya juga perlu diketahui untuk menetapkan prioritas. Pada umumnya, proses invasi terjadi secara bertahap dan relatif perlahan sehingga tidak banyak disadari. Invasi oleh spesies asing diawali dengan kehadiran spesies tersebut di suatu daerah, kemudian terjadi pertumbuhan dan adaptasi dengan lingkungan sekitar. Proses selanjutnya terjadi perkembangbiakan hingga dominasi dan pengambilalihan daerah jelajah/penyebaran spesies dominan terdahulu/setempat oleh spesies asing yang kemudian berubah menjadi spesies invasif (Radiansyah et al. 2015).
Secara umum, tingkatan proses invasi suatu spesies tumbuhan dan hewan dapat dibagi mulai dari pengangkutan, yaitu pergerakan suatu spesies dari tempat asal ke lokasi baru, sampai dengan penyebaran dan dampak yang ditimbulkan di lokasi baru. Catford (2009 dalam Radiansyah et al. 2015) membagi tingkat invasi
suatu spesies invasif ke dalam beberapa kategori tingkatan, yaitu: 1) transport, 2) introduksi, 3) kolonisasi, 4) naturalisasi , 5) penyebaran, dan 6) dampak. Dijelaskan pula bahwa tingkatan invasi tersebut ditentukan oleh hasil interaksi dan besar kecilnya ketiga faktor pendorong invasi, yang terdiri dari: 1) propagul (P), yaitu bagian dari tumbuhan seperti tunas atau anakan yang dapat hidup menjadi tumbuhan baru, 2) faktor abiotik (A), yaitu faktor kimia dan fisika dallam lingkungan, seperti cahaya, temperatur, air, gas di udara dan angin serta tanah, edafik hewan dan fisiografi, 3) faktor biotik (B), yaitu hal yang berkaitan dengan dihasilkan atau disebabkan oleh mahluk hidup (Radiansyah et al. 2015).
Spesies Pesaing bagi Spesies Asing Invasif
Penelitian sebelumnya menunjukkan kompetisi antara spesies asing invasif dan spesies asli (Lankau 2009, Denoth & Myers 2007, Lankau 2012). Beberapa spesies asli mampu berkompetisi dengan spesies asing invasif. Dampak spesies asing invasif Lythrum salicaria tidak cukup kuat untuk menekan spesies asli Sidalcea hendersonii di vegetasi mangrove Pasifik Baratlaut (Denoth & Myers
2007).
Spesies asing umumnya lebih kompetitif dibanding spesies asli karena kandungan alelopati yang dimilikinya (Callaway & Ridenour 2004). Meskipun demikian, terdapat speses asli yang mampu bertahan terhadap dampak negatif alelopati tersebut. Spesies asli Amerika utara Pilea pumila toleran terhadap
tingginya kandungan alelopati dalam tanah yang dikeluarkan spesies asal Eropa
Alliaria petiolata (Lankau 2012).
Selain itu, kemampuan kompetitif spesies asing dapat dihambat oleh aktivitas mikroba tanah (Lankau 2009). Alelopati yang dikeluarkan oleh A. petiolata dapat didegradasi oleh mikroba tanah sehingga mengurangi dampak
konteks lingkungan yang megurangi atau mengeliminasi penghambatan alelopati dapat menjadi informasi yang penting dalam keragaman spasial dalam keberhasilan invasi dan proses penanganannya (Lankau 2009).
Spesies Diagnostik
Identifikasi karakteristik atau spesies indikator/diagnostik merupakan aktivitas tradisional dalam ekologi dan biogeografi (Dufrêne & Legendre 1997). Studi berdasarkan kerja lapangan, mendeskripsikan suatu lokasi atau habitat, umumnya menyebut satu atau beberapa spesies yang mencirikan tiap habitat. Indicator Value atau IndVal, menjadi metode yang lebih sederhana dibanding Twinspan yang masih memerlukan pseudospesies (Dufrêne & Legendre 1997).
Dalm perkembangannya, identifikasi spesies diagnostik dapat ditaksir dengan koefisien phi (Ф, Chytrỳ et al. 2002). Koefisien phi lebih baik dibanding
IndVal karena bersifat simetris (bernilai positif dan negatif). Koefisien phi juga lebih baik dibanding indeks diagnostik lainnya (Chytrỳ et al. 2002). Meskipun
demikian, untuk data yang besar, lebih disarankan menggunakan indeks Ochiai (De Cáceres et al. 2008).
Analisis Sainstometri
Sainstometri adalah studi aspek kuantitatif sains dan teknologi sebagai suatu proses komunikasi (Mingers & Leydesdorff 2015). Sainstometri berbeda dari jenis metrik lainnya seperti bibliometri, webometri, infometri, dan almetri. Umumnya sainstometri berkaitan dengan analisis kutipan dalam literatur akademik. Selanjutnya, kutipan juga menyediakan sebuah kaitan antara publikasi sebelumnya dari pustaka tersebut dan kemunculan kutipan setelahnya. Pentingnya kutipan dalam hal ini dimulai dari ide Index Kutipan Sains (Science Citation Index, SCI) yang diusulkan oleh Eugene Garfield sebagai pangkalan data untuk
perhitungan kutipan (Garfield 1955).
Sumber utama kutipan di masa lalu yang dikenal hingga saat ini yaitu Thomson Reuters Institut for Scientific Information (ISI) Web of Science (WoS)
yang kemudian diikuti oleh Scopus dari penerbit Elsevier dan Google Scholar (Mingers & Leydesdorff 2015). Lin dan Fenner (2013) mengategorikan ketiganya ke dalam tipe dampak sumber kutipan yang paling signifikan. Masing-masing sumber kutipan tersebut memiliki keunggulan dan kelemahan. Google Scholar (GS) memiliki keunggulan yaitu cakupan output penelitian yang lebih besar, yang mana subjek non-sains Scopus dan WoS lemah. Selain itu, GS menghasilkan jumlah kutipan yang lebih besar untuk penelitian tertentu.
Beberapa penelitian menggunakan pendekatan sainstometri untuk menganalisis suatu bidang atau topik secara kuantitatif. Misalnya, Nadkarni et al.
(2011) menganalisis topik tajuk hutan sebagai suatu bidang sains yang sedang muncul. Laju kutipan tajuk hutan relatif lebih tinggi dibanding bidang geologi dan jumlah kutipan meningkat drastis dalam kurun waktu 1990-2010. Sainstometri juga dapat digunakan untuk mencari suatu istilah yang umum dalam suatu bidang sains. Pautasso (2016), mengamati perkembangan topik kesehatan hutan dan penyakit pohon. Pautasso menemukan fakta bahwa para peneliti cenderung menggunakan istilah “kesehatan pohon” dibanding “penyakit pohon”.
Sainstometri juga dapat diaplikasikan untuk beberapa tujuan yaitu: 1. Menguji tren temporal dalam jumlah publikasi topik tertentu
2. Menginvestigasi distribusi spasial fokus penelitian topik tertentu
3. Mempelajari jumlah relatif artikel yang berkaitan dengan berbagai area penelitian
4. Menganalisis sambutan artikel bagi generasi di masa depan 5. Menaksir produktivitas peneliti dan kualitasnya
6. Membandingkan performansi penelitian dan keragaman institusi dan negara 7. Mendokumentasikan pola dan penentu lamanya proses tinjau ulang yang
mendalam (peer review)
3 METODE
Pengaruh C. peltata terhadap Vegetasi Karst Cibodas Deskripsi Lokasi
Penelitian dilaksanakan di kawasan Karst Gunung Cibodas, Kecamatan Ciampea, Kabupaten Bogor, posisi astronomis yaitu 6o33’01.04”S-6o33’19.06”S
dan 106o41’38.63”T-106o40’39.08”T. Secara administrasi, kawasan terbagi
menjadi beberapa status kepemilikan dengan Perum Perhutani pemilik lahan terluas (84 ha). Karst Gunung Cibodas merupakan hutan produksi dan terdiri atas lereng (utara dan selatan) yang cukup luas dan punggung relatif lebih sempit. Lereng selatan pernah ditanami pohon mahoni dan kaliandra tahun 2002.
Tahun 2004, lereng selatan mengalami penebangan pohon dengan intensitas tinggi hingga menjadi lahan terbuka minim tutupan vegetasi. Selama survei pendahuluan, C. peltata ditemukan di lereng selatan dengan kerapatan yang
relatif cukup tinggi (Gambar 2). Secara visual, lereng selatan didominasi oleh spesies ini, sehingga secara kuantitatif, C. peltata diduga mendominasi lereng
selatan.
Gambar 2 Kondisi vegetasi lereng selatan yang didominasi oleh C. peltata
Dengan demikian, terdapat tiga lokasi berbeda berdasarkan topografi dan kondisi invasi yaitu punggung, lereng selatan (terinvasi) dan lereng utara (tak terinvasi). Elevasi (diukur dengan GPS) lokasi ini antara 155-355 mdpl. Batuan
induk Karst Gunung Cibodas terdiri atas formasi Bojongmanik terbentuk sejak masa Miosen Tengah. Kondisi iklim, kelerengan dan bukaan tajuk disajikan dalam Tabel 1. Data iklim diperoleh dari Stasiun Pengamatan Badan Meteorologi Klimatolog dan Geofisika (BMKG) Dramaga.
Tabel 1 Kondisi iklim, kelerengan dan bukaan tajuk di lokasi penelitian (rerata±galat baku) pada 2012-2014
Lokasi Curah hujan
(mm tahun-1) suhu udara (oC) kelembapan udara (%) Kelerengan (%) tajuk (%) Bukaan
Punggung
3935±256 25.8±0.46 83.1±4 29.42±3.26 66.11±9.53
Lereng utara 66.10±9.69 12.8±3.88
Pencuplikan Vegetasi dan Tanah
Observasi lapangan dilakukan pada Maret-Mei 2015. Pencuplikan vegetasi dilakukan secara purposive mempertimbangkan perbedaan topografi (punggung
dan lereng) dan kondisi (terinvasi dan tak terinvasi). Teknik pencuplikan vegetasi dilakukan dengan kombinasi transek dan plot kuadrat bersarang (Gambar 3).
Gambar 3 Ilustrasi transek dan plot pencuplikan. a : 20 x 20 m untuk strata pohon diameter >20 cm, b : 10 x 10 m untuk tiang (diameter 10-20 cm), c : 5 x 5 m untuk pancang (diameter <10 cm dan tinggi >1.5 m), dan d: 2 x 2 m2 untuk semai (tinggi <1.5 m) dan tumbuhan bawah (herba dan
semak). : titik pencuplikan tanah
Transek dibuat sepanjang 100 m berjumlah tiga untuk masing-masing lokasi sehingga total transek berjumlah 15. Setiap transek terdiri atas lima plot berukuran 20 x 20 m. Jumlah plot lereng utara, lereng selatan dan punggung masing-masing 15.
Data vegetasi dihitung setiap plot yaitu jumlah individu dan penutupan. Penutupan diukur dari diameter setinggi dada pada strata pohon, tiang dan pancang menggunakan jangka sorong/meteran. Cuplikan tanah diambil setiap transek dari permukaan hingga kedalaman 15 cm setelah bersih dari serasah. Sepuluh cuplikan tanah diambil dari tiap transek (Gambar 3), dikompositkan dan diambil sebanyak 0.5-1 kg untuk dianalisis. Karakter kimia dan fisika diukur yaitu pH, C-organik, N, P-tersedia, kation dapat ditukar (Ca, Mg, Na dan K) dan tekstur (liat, debu, dan pasir) yang dianalisis di Laboratorium Balai Besar Penelitian dan Pengembangan Tanah, Kementerian Pertanian.
Analisis Data
Data vegetasi yang dikoleksi setiap strata yaitu keanekaragaman spesies dan dominansi. Keanekaragaman spesies terdiri atas alfa dan beta. Jumlah spesies (alfa) diestimasi dengan estimator Chao2 menggunakan perangkat lunak EstimateS versi 9.1.0 (Colwell 2013).
Sampling completeness (SC) dihitung untuk mengestimasi apakah
pencuplikan sudah cukup mewakili total lokasi penelitian. Interpolasi dan ekstrapolasi jumlah spesies juga dihitung untuk mengestimasi pertambahan jumlah spesies sebanyak jumlah cuplikan (plot) hingga mencapai titik jenuh (asimtot). Sampling completeness (SC), keanekaragaman dan kemerataan spesies
dihitung menggunakan indeks Shannon-wiener dan Pielou (Magurran 2004) sebagai berikut:
SC = Sobs × 100%
H’ = - Σ N n × ln N n
E = H’
ln Sobs
Keterangan:
SC : Sampling completeness
Sobs : Jumlah spesies yang teramati
Sest : Jumlah spesies yang diestimasi
H’ : Indeks keanekaragaman Shannon-wiener E : Indeks kemerataan Pielou
n : Jumlah individu spesies i
N : Jumlah individu seluruh spesies
Keanekaragaman beta dihitung dengan partisi aditif untuk mengestimasi rerata jumlah spesies setiap level pencuplikan yang berbeda sebagai berikut:
Alfa1 : rerata jumlah spesies tiap plot Beta1 = Alfa2-Alfa1 Alfa2 : rerata jumlah spesies transek tiap lokasi Beta2 = Alfa3-Alfa2 Alfa3 : rerata jumlah spesies tiap lokasi (Wagner et al. 2000)
Summed Dominance Ratio (SDR) dihitung dari nilai rata-rata komponen
Indeks Nilai Penting (INP) yaitu kerapatan relatif (KR), frekuensi relatif (FR) dan penutupan relative (DR) untuk mengetahui dominansi spesies (Husch et al. 2003).
Perhitungan INP strata semai dan tumbuhan bawah hanya terdiri atas kerapatan dan frekuensi karena mengukur penutupan tajuk di lapangan sulit. SDR dihitung untuk setiap strata untuk seluruh lokasi sebagai berikut:
SDR = KR+FR+DR untuk pancang, tiang dan pohon Kerapatan mutlak total (individu ha-1)
FRi = Frekuensi mutlak (jumlah plot terisi ) spesies Frekuensi mutlak total (jumlah plot terisi ) i × 100%
DRi = Penutupan mutlak (m
2 ha-1) spesies i
Analisis ordinasi bipolar komunitas dari data hasil analisis vegetasi secara garis besar dapat dikelompokkan dari beberapa tahapan yaitu tabulasi data, pembuatan matrik nilai kemiripan dan ketidak miripan, penentuan sumbu X, penentuan sumbu Y, dan pembuatan grafik.
1. Tabulasi Data
Tabular data secara sederhana menunjukkan kehadiran dan ketidakhadiran dari spesies-spesies yang terdapat di dalam komunitas yang sedang diamati. Walaupun secara lebih terperinci dari struktur komunitas tumbuhan dapat mengadakan studi perbandinganyang dapat langsung menunjukkan nilai penting jenis di dalam komunitasnya. Data dari tabel ini selanjutnya digunakan kedalam perhitungan-perhitungan seperti pada tahap 2 dan seterusnya.
2. Pembuatan Matrik Nilai Kemiripan dan Ketidak Miripan
Dari data indeks nilai penting dapat dihitung besarnya nilai kemiripan dan ketidak miripan dua komunitas yang sedang diperbandingkan. Nilai kemiripan atau indeks similarity (IS) dari persamaan Bray-Curtis diperoleh dari perangkat lunak EstimateS. Nilai ketidak miripan atau indeks disimilarity (ID) dapat dihitung dengan rumus :
ID = 100 – IS
Selanjutnya nilai-nilai dari seluruh perbandingan komunitas yang ada disusun kedalam matrik.
3. Penentuan Sumbu X
a. Menetapkan titik A dengan syarat (a) memiliki jumlah total IS terkecil, dan (b) harus mempunyai paling tidak 3 buah nilai IS ≥ 50
b. Menentukan titik B dengan syarat (a) memiliki ID terbesar terhadap titik A, (b) harus mempunyai paling sedikit 3 buah nilai IS ≥ 50
c. Setelah titik A dan B diketahui, maka dapat diketahui nilai L yang merupakan nilai ketidaksamaan antara titik A dan titik B
d. Menentukan posisi titik yang lain pada ordinat X dengan menggunakan rumus Beal sebagai berikut :
x = L2+(dA)2 - (dB)2 2L
Keterangan:
L = Besarnya sumbu dari perpotongan dari nilai ketidak miripan A dan B dA = nilai ID dari titik A
4. Penentuan Sumbu Y
a. Menentukan harga ex2 dari masing-masing plot dengan rumus:
ex2 = (dA)2 - x2
b. Menentukan titik A’ atau titik acuan pertama ordinat Y dengan syarat (a) memiliki harga tertinggi ex2, (b) harus terletak dalam kisaran rata-rata 50%
nilai tengah ordinat X, dan (c) memiliki paling sedikit 3 IS ≥ 50
c. Menentukan titik B’ sebagai titik acuan kedua ordinat Y, dengan syarat (a) harus sedekat mungkin dengan titik acuan pertama sepanjang ordinat X, (b) harus mempunyai ID terbesar terhadap titik A’, (c) harus memiliki sedikitnya 3 IS ≥ 50, dengan kata lain titik ini adalah paling bsear ketidak miripannya terhadap titik A’
d. Setelah titik A’ dan B’ diketahui, maka dapat diketahui nilai ketidaksamaan kedua titik tersebut (L’) seperti menentukan nilai L.
e. Menentukan posisi titik yang lain pada ordinat Y dengan menggunakan rumus Beal:
y = L2+(dA)2-(dB)2 2L
5. Grafik dan Delineasi Pengelompokkan
Langkah selanjutnya adalah membuat delineasi pengelompokkan dan menterjemahkan kedalam sumbu gradient perubahan lingkungan yang diamati selama survei pengumpulan data. Untuk itu akan terlihat adanya pola perubahan komunitas yang berkesinambungan atau yang terputus mendadak, dimana hal tersebut di atas sangat erat berkaitan dengan faktor lingkungan yang sedang diamati, yang digambarkan sebagai sumbu X dan sumbu Y.
Spesies diagnostik ditentukan dari nilai koefisien phi (Ф) yang diperoleh dari data frekuensi (Chytrỳ et al. 2002). Kehadiran spesies diagnostik
mengindikasikan secara spesifik lokasi tertentu. Keofisien phi berkorelasi positif dengan INP (Lampiran 1), namun menawarkan uji statistik yang lebih jelas dibanding INP. Koefisien phi diperoleh data kehadiran-ketidakhadiran yang dirangkum dalam tabel kontingensi 2 × 2 (Tabel 2) yaitu:
Tabel 2 Kontingensi 2 × 2 untuk menentukan komponen koefisien phi Jumlah plot Di dalam grup plot Tidak terdapat di dalam grup plot
Terdapat spesies i np n-np
Tidak terdapat spesies i Np - np N - Np - n + np
berdasarkan tabel tersebut, maka persamaan koefisien phi yang diperoleh yaitu:
Ф = (N × np )- (n × Np)
dengan indikasi nyata lokasi target secara statistik yang diperoleh dari persamaan:
Z : nilai Z statistik, di mana titik kritis untuk tolak H0 (indikasi nyata lokasi
target) yaitu α=0.05>1.96
Asosiasi spesies diagnostik dengan C. peltata
Asosiasi antara C. peltata dan spesies lainnya dihitung di setiap strata dengan
persamaan khi kuadrat (χ2)melalui tabel kontingensi 2 × 2 (Tabel 3) yaitu:
Tabel 3 Kontingensi 2 × 2 untuk menentukan komponen asosiasi
C. peltata
Dengan tingkat kekuatan asosiasi yang diperoleh dari persamaan:
E(a) = (a + b) × (a + c) E(b) = (b + d) × (a + b)
b : Jumlah plot yang berisi spesies lainnya saja c : Jumlah plot yang berisi C. peltata saja
d : Jumlah plot yang tidak berisi C. peltata dan spesies lainnya
E(a) : Preferensi antara C. peltata dan spesies lainnya yang diharapkan, jika
a>E(a), maka terdapat preferensi antara C. peltata dan spesies lainnya
E(b) : Pengelakan antara spesies lainnya dan C. peltata yang diharapkan, jika
Data vegetasi setiap strata yaitu kerapatan (individu ha-1), penutupan/basal
area (m2 ha-1), jumlah spesies, dan keanekaragaman beta dihitung di setiap lokasi.
Perbedaan statistik interpolasi jumlah spesies antar strata berdasarkan kriteria konservatif selang kepercayaan minimal 95% yang tak bersinggungan (Colwell 2013). Karakter floristik dihitung untuk C. peltata hanya kerapatan dan
penutupan. Analisis ragam digunakan untuk menguji pengaruh perbedaan lokasi terhadap karakter floristik (keanekaragaman beta, kerapatan dan penutupan) setiap strata dan faktor tanah, serta uji lanjut Duncan jika terdapat perbedaan nyata pada α=0.05. Uji Z dilakukan untuk mengetahui pengaruh invasi (lereng selatan vs punggung dan lereng utara) dan topografi (punggung vs lereng) terhadap keanekaragaman alfa (jumlah spesies, indeks Shannon-wiener, dan indeks kemerataan Pielou). Data keanekaragaman alfa tersebut diperoleh dari interpolasi yang dijalankan pada perangkat lunak EstimateS. Pendugaan keragaman karakter floristik (y) oleh faktor tanah (x) menggunakan analisis regresi tunggal. Analisis ragam dan regresi menggunakan perangkat lunak SPSS versi 22.
Analisis Sainstometri C. peltata
Penelitian ini dilaksanakan pada 6 Januari 2016 menggunakan pangkalan data Scopus yang terdaftar melalui akun perpustakaan Institut Pertanian Bogor. Pencarian literatur menggunakan kata kunci 32 spesies asing invasif (termasuk C. peltata) yang disusun oleh ISSG (Invasive Species Specialist Group, Lowe et al.
2000). Pencarian tipe dokumen dibatasi hanya artikel penelitian dan ditulis antara 1990 hingga 2015. Kata kunci pencarian juga dibatasi hanya di dalam judul dan/atau abstrak dan/atau kata kunci artikel untuk menentukan spesies sebagai objek penelitian.
Jumlah kutipan digunakan untuk menganalisis secara kuantitatif topik ini. Kata kunci spesifik yang digunakan yaitu ((TITLE-ABS-KEY (“species”) AND TITLE-ABS-KEY(invasion) AND (LIMIT-TO(DOCTYPE,”ar”))) untuk membandingkan kutipan artikel dari topik invasi tiap spesies. Manfaat dan kerugian yang ditimbulkan oleh C. peltata kemudian dianalisis dengan pencariaan
kata kunci literatur yaitu ((TITLE-ABS-KEY (“Cecropia peltata” AND (LIMIT -TO(DOCTYPE,”ar”)). Manfaat dan kerugian ditentukan dari abstrak dan/atau artikel lengkap, apakah C. peltata bermanfaat bagi manusia dan lingkungan atau
justru menimbulkan kerugian. Hasil pencarian kemudian diunduh dan disusun dengan urutan: penulis, tahun publikasi, judul artikel, nama jurnal, volume, halaman, jumlah total kutipan, jumlah kutipan per tahun, dan indeks h. Kutipan
per tahun dibatasi dari periode 1970 hingga 2015. Perbandingan laporan invasi antar spesies dihitung dari kutipan relatif yaitu:
Kutipan relatif spesies i = Jumlah kutipan spesies i × 100 %
3 HASIL
Pengaruh Invasi C. peltata terhadap Vegetasi Karst Gunung Cibodas Struktur Vegetasi
Hasil survei menunjukkan adanya tiga spesies mendominasi kelima strata (Tabel 4). Meskipun demikian, koefisien keragaman C. peltata relatif lebih kecil
dibanding Calliandra sp, Syzygium lineatum dan Nephrolepis biserrata, yang
berarti nilai kerapatan, frekuensi, dan penutupan C. peltata lebih merata. Cecropia peltata mendominasi strata tiang dan pohon dengan nilai SDR tinggi (Tabel 4).
Selain itu, sampling completeness strata pancang dan tiang rendah, sehingga
hanya 56.44 % dan 41.18 % spesies tercatat selama observasi.
Tabel 4 Indeks keanekaragaman dan kemerataan spesies setiap strata dan spesies dominan di Kawasan Karst Gunung Cibodas
Strata Sobs Sest SC (%) H’ E Spesies dominan SDR (KK)
Tumbuhan bawah 48 58 82.76 2.40 0.62 Nephrolepis biserrata 22.40 (0.52)
Semai 33 55 60.00 2.73 0.78 Syzygium lineatum 12.88 (0.29)
Pancang 57 101 56.44 3.00 0.74 Calliandra callothyrsus 12.62 (0.57)
Tiang 7 17 41.18 0.96 0.49 Cecropia peltata 51.46 (0.38)
Pohon 5 8 62.50 0.76 0.47 Cecropia peltata 72.78 (0.08)
Sobs: jumlah spesies yang teramati; Sest: estimasi jumlah spesies (Chao2) total kawasan ; SC (Sampling completeness; H’: indeks keanekaragaman Shannon-Wiener; E: indeks kemerataan Pielou; SDR: Summed Dominance Ratio; KK: koefisien keragaman: simpangan baku/SDR
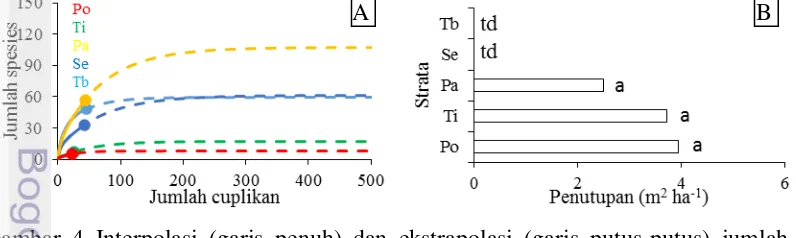
Kurva interpolasi dan ekstrapolasi menunjukkan jumlah spesies strata tiang dan pohon paling rendah, sedangkan pancang paling tinggi (Gambar 4A). Meskipun jumlah spesies pancang teramati lebih tinggi dibanding tumbuhan bawah dan semai (Tabel 4), namun kurva ekstrapolasi menunjukkan tidak ada perbedaan nyata antara ketiga strata (Gambar 4A). Tidak ada perbedaan nyata penutupan pancang, tiang dan pohon (Gambar 4B). Keanekaragaman beta dan penutupan pancang dan tumbuhan bawah lebih tinggi dibanding strata lain (Tabel 5).
Gambar 4 Interpolasi (garis penuh) dan ekstrapolasi (garis putus-putus) jumlah spesies (A); penutupan (B). Po: pohon; Ti: tiang; Pa: pancang; Se: semai; Tb: tumbuhan bawah. Huruf berbeda menunjukkan perbedaan nyata di setiap strata di P<0.05 uji lanjut Duncan; td: tidak dihitung
Tabel 5 Keanekaragaman beta setiap level pencuplikan (plot, transek, dan lokasi) dan kerapatan. Huruf berbeda dalam kolom yang sama menunjukkan perbedaan nyata di P<0.05 uji lanjut Duncan
Keanekaragaman beta
Kerapatan (x 103 m2 ha-1)
Plot Transek Lokasi
Tumbuhan bawah 5.64d 9.36c 7.33b 99.00c
Semai 3.26b 5.07b 8.00b 28.69b
Pancang 4.38c 8.94c 11.33b 4.39a
Tiang 1.15a 1.13a 1.04a 0.24a
Pohon 1.13a 0.86a 0.66a 0.05a
Karakteristik Tanah
Kondisi tanah di ketiga lokasi cukup beragam (Gambar 5). Kandungan karbon, nitrogen dan rasio karbon:nitrogen di punggung lebih rendah dibanding lereng. Persentase fraksi pasir lereng utara paling tinggi, sedangkan pH tanah lereng selatan paling tinggi. Kandungan hara di punggung lebih rendah dibanding lereng, kecuali magnesium (Gambar 5). Perbedaan faktor cukup beragam terhadap perbedaan fisika dan kimia tanah, namun topografi lebih memengaruhi perbedaan hara tanah. Data tanah selengkapnya disajikan pada Lampiran 2.
Gambar 5 Rerata karakter fisika dan kimia tanah setiap lokasi. P: punggung, LU: lereng utara, LS: lereng selatan. Huruf berbeda menunjukkan perbedaan nyata di P<0.05 uji lanjut Duncan
Pengaruh Invasi C. peltata
Terdapat dua kelompok perlakuan yaitu topografi (lereng vs punggung) dan invasi (terinvasi dan tak terinvasi C. peltata) yang diduga memengaruhi
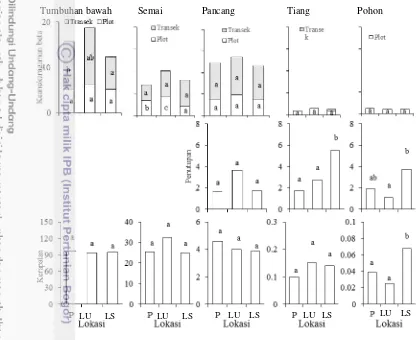
keanekaragaman alfa. Analisis interpolasi menunjukkan jumlah spesies strata tumbuhan bawah dan semai yang lebih rendah di lokasi terinvasi (Gambar 6). Indeks Shannon-wiener dan kemerataan Pielou pancang di lokasi terinvasi lebih rendah dibanding lokasi tak terinvasi C. peltata. Sebaliknya, jumlah spesies dan
indeks Shannon-wiener tiang justru dipengaruhi oleh perbedaan topografi. a
Keanekaragaman beta level transek tumbuhan bawah di punggung lebih tinggi dibanding lereng selatan dan semai level plot lebih tinggi di lokasi tak terinvasi. Keanekaragaman beta level plot dan transek pancang, tiang dan pohon tidak berbeda nyata antar lokasi (Gambar 7). Sebaliknya, tren berbeda terjadi pada penutupan dan kerapatan. Strata pohon memiliki penutupan dan kerapatan lebih tinggi di lereng selatan dibanding punggung dan lereng utara, sementara strata lainnya tidak berbeda nyata antar lokasi (Gambar 7).
Tumbuhan bawah Semai Pancang Tiang Pohon
Gambar 7 Pengaruh invasi terhadap keanekaragaman beta, penutupan (m2 ha-1)
dan kerapatan (individu x 103 ha-1). Huruf berbeda menunjukkan
perbedaan nyata antar lokasi berdasarkan uji lanjut Duncan pada
P<0.05. P: punggung, LU: lereng utara, LS: lereng selatan
Derajat Invasi C. peltata
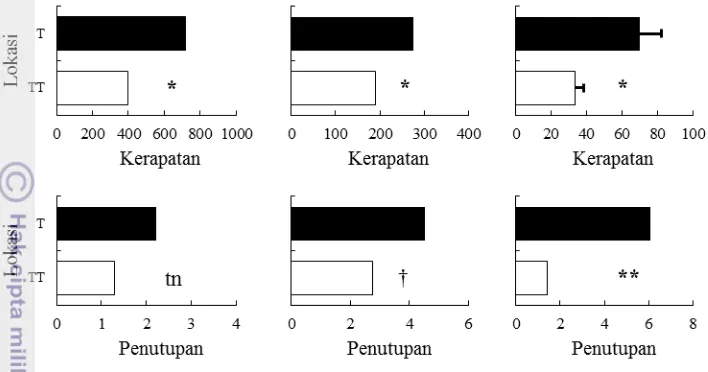
Derajat invasi C. peltata diestimasi dengan perhitungan kerapatan dan
penutupan. Berdasarkan lokasi (lereng utaran dan selatan, punggung), kerapatan dan penutupan C. peltata tidak berbeda nyata kecuali strata pohon. Kerapatan dan
penutupan pohon C. peltata lebih tinggi di lereng selatan dibanding punggung.
Sementara itu, satu individu strata pohon C. peltata hanya ditemukan di lereng
utara di mana bukaan tajuk lebih rendah dibanding punggung (P=0.005, uji lanjut
Duncan).
Invasi C. peltata dapat dideteksi dari nilai kerapatan dan penutupannya.
Invasi ditandai saat kerapatan pancang, tiang dan pohon C. peltata mencapai
LU LS LU LS
LU LS LU LS
kisaran 71-1368, 128-649, dan 42-98 individu ha-1. Selain itu, invasi ditandai saat
penutupan tiang dan pohon mencapai 2.5-10.69 dan 3.08-9.10 m2 ha-1 (Gambar 8).
Nilai SDR juga menunjukkan rerata kerapatan, frekuensi dan penutupan pohon C. peltata lebih dari 70 % (Tabel 4).
Gambar 8 Rerata kerapatan (individu ha-1) dan penutupan (m2 ha-1) C. peltata. T:
terinvasi C. peltata, TT: tak terinvasi. Perbedaan nyata antar lokasi
berdasarkan uji T pada †P<0.1, *P<0.05, **P<0.01. tn: tidak nyata
Pendugaan Karakter Floristik dengan Faktor Tanah
Keragaman karakter floristik dapat diduga oleh faktor tanah terutama pH, tekstur pasir dan kandungan magnesium (Tabel 6). Keanekaragaman alfa dan beta tumbuhan bawah dan semai diduga oleh faktor pH dan pasir sebanyak 54.6 % hingga 86.2 %, sementara kerapatan tumbuhan bawah diduga oleh magnesium sebanyak 82.1 %. Kerapatan dan penutupan C. peltata tidak cukup baik dibanding
faktor tanah untuk menduga keragaman karakter floristik (Lampiran 3).
Tabel 6 Penduga keanekearagaman dan kerapatan tumbuhan bawah dan semai berdasarkan analisis regresi
Peubah tak bebas (y) α Penduga (x) βa adj. R2 F P
Jumlah spesies tumbuhan bawah 55.26 pH -5.16 0.694 6.89 0.006
Jumlah spesies semai 34.82 pH -3.74 0.557 9.80 0.020
β-plot semaib -1.35 Pasir 0.68 0.546 9.42 0.022
β-transek tumbuhan bawahc 51.81 pH -1.23 0.862 44.90 0.001
Kerapatan tumbuhan bawah 135.60 Mg -0.92 0.821 33.10 0.001
a Beta baku, bβ-plot: keanekaragaman beta level plot; cβ-transek: keanekaragaman beta level transek
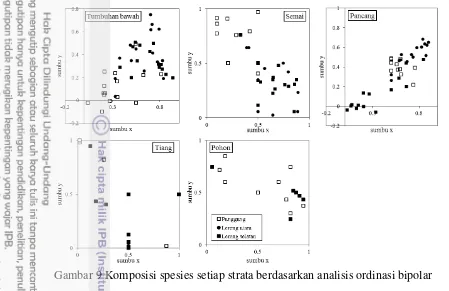
Komposisi Spesies setiap Strata
Secara kualitatif, berdasarkan analisis ordinasi bipolar, komposisi spesies tumbuhan di lokasi penelitian berbeda terutama strata tumbuhan bawah, semai dan pancang (Gambar 9). Komposisi tumbuhan bawah dan semai di punggung berbeda dengan lereng, sementara strata pancang berbeda di tiap lokasi. Hal tersebut menunjukkan bahwa komposisi penyusun strata pancang di satu lokasi berbeda dengan lokasi lainnya. Spesifitas strata pancang di lereng selatan menunjukkan representasi lokasi yang terinvasi C. peltata, yang mana kerapatan
spesies diagnostik, dapat menjadi kandidat spesies yang mampu berasosiasi dengan C. peltata.
Gambar 9 Komposisi spesies setiap strata berdasarkan analisis ordinasi bipolar
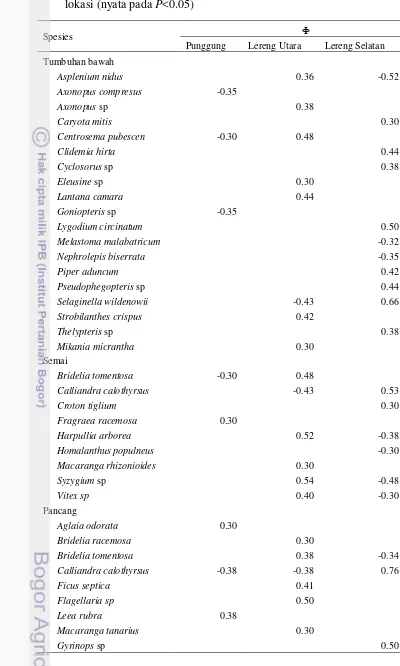
Spesies Diagnostik setiap Strata
Nilai koefisien phi yang berkisar -1 hingga +1, menunjukkan kesamaan kisaran nilai dengan korelasi Pearson dalam uji korelasi parametrik. Tabel 7 menunjukkan adanya variasi nilai koefisien phi. Beberapa spesies hanya memiliki nilai minus di suatu lokasi, namun tidak menjadi spesies diagnostik di lokasi lainnya. Sebaliknya, beberapa spesies menjadi spesies diagnostik di suatu lokasi, namun tetap ditemukan di lokasi lainnya walaupun dengan frekuensi kehadiran yang sedikit. Nilai koefisien phi selengkapnya ditampilkan di Lampiran 4.
Spesies lainnya yaitu Selaginella wildenowii, Asplenium nidus, Calliandra calothyrsus, Harpullia arborea, Vitex sp, Syzygium picnatum, Syzygium sp, Bridelia tomentosa, dan Cecropia peltata memiliki dua nilai koefisien phi yang
berlawanan (Tabel 7). Spesies-spesies tersebut menjadi spesies diagnostik di suatu lokasi dan tidak berkorelasi dengan lokasi lainnya. C. peltata menjadi spesies
diagnostik strata pohon di lereng selatan dan tidak berkorelasi dengan lereng utara.
Hal tersebut sesuai dengan nilai kerapatan dan penutupannya yang lebih tinggi di lereng selatan (Gambar 8). Berdasarkan lokasi, delapan spesies strata tumbuhan bawah (Clidemia hirta, Cryota mitis, Cyclosorus sp, Lygodium circinatum, Piper aduncum, Pseudophegopteris sp, S. wildenowii dan Thelypteris
sp), dua spesies semai (C. calothyrsus dan Croton triglium), dan tiga spesies
pancang (C. calothyrsus, Gyrinops sp, dan Cinnamomum iners) menjadi spesies
Tabel 7 Spesies diagnostik (dengan nilai koefisien phi) setiap strata di setiap lokasi (nyata pada P<0.05)
Spesies Ф
Punggung Lereng Utara Lereng Selatan Tumbuhan bawah
Calliandra calothyrsus -0.38 -0.38 0.76
Ficus septica 0.41
Flagellaria sp 0.50
Leea rubra 0.38
Macaranga tanarius 0.30
Tabel 7 Spesies diagnostik (dengan nilai koefisien phi) setiap strata di setiap lokasi (nyata pada P<0.05) (lanjutan)
Spesies Ф
Punggung Lereng Utara Lereng Selatan
Microcos hirsuta 0.30
Poikilospermum suaveolens 0.30
Cinnamomum iners 0.50
Syzygium picnatum 0.48 -0.30
Vitex sp -0.34 0.40
Tiang
Buchanania arborescens -0.30
Pohon
Cecropia peltata -0.45 0.42
Asosiasi C. peltata dengan Tumbuhan Lainnya
Seperti halnya koefisien phi yang bernilai positif dan negatif, maka asosiasi antar spesies yang diestimasi dengan khi kuadrat juga bernilai positif atau negatif.
C. calothyrsus (pancang) nyata berasosiasi positif dengan C. peltata (pohon),
sementara Cassia siamea (pancang) nyata berasosiasi negatif berdasarkan uji khi
kuadrat. Gambar 10 mengilustrasikan kondisi asosiasi kedua spesies tersebut dengan C. peltata.
Gambar 10 Ilustrasi asosiasi positif antara C. calothyrsus dengan C. peltata, dan
asosiasi negatif antara C. siamea dan C. peltata. Keduanya nyata
berasosiasi pada P<0.05 uji khi kuadrat
Analisis Sainstometri C. peltata
ellipticus (Tabel 8). Peubah Indeks h berkorelasi positif dengan jumlah artikel dan
kutipan (Lampiran 5). Lantana camara dan Lythrum salicaria merupakan spesies
yang aktivitas invasinya paling banyak dilaporkan berdasarkan jumlah kutipan dan indeks h yang paling tinggi. Sebaliknya, C. peltata merupakan spesies yang
paling sedikit dilaporkan dibanding spesies lainnya. Hasil pencarian ini menandakan invasi C. peltata terhadap spesies asli dan dampaknya terhadap
lingkungan yang sedikit.
Tabel 8 Ringkasan jumlah artikel dan kutipannya yang memublikasikan spesies tumbuhan asing invasif dan istilah “invasion” di pangkalan data Scopus Spesies Jumlah artikel Jumlah kutipan Kutipan relatif (%) indeks h
Acacia mearnsii 21 475 4.143 8
Melaleuca quinquenervia 29 417 3.637 12
Miconia calvescens 16 203 1.770 8
Schinus terebinthifolius 13 244 2.128 8
Spathodea campanulata 4 65 0.566 2
Sphagneticola trilobata 3 11 0.096 2
Tamarix ramosissima 26 966 8.426 16
Saat lingkup pencarian diperluas dengan menghapus istilah “invasion”, kutipan C. peltata juga lebih lambat dibanding L. camara (Gambar 11A). Lingkup
yang lebih luas ini mencakup tidak hanya kerugian yang ditimbulkan akibat invasi, tapi juga manfaatnya. Meskipun demikian, total jumlah kutipan L. camara
relatif lebih tinggi dibanding C. peltata. Perbandingan antara manfaat dan
kerugian yang ditimbulkan C. peltata cukup jauh. Dampak positif lebih tinggi
dibanding negatif yang ditimbulkan (Gambar 11B, Tabel 9).
Gambar 11 Laju kutipan C. peltata dan L. camara per tahun (A). Total artikel dan
kutipan C. peltata 59 dan 1388, sedangkan L. camara 1066 dan
13234. Persentase kutipan manfaat dan kerugian C. peltata. Total
jumlah kutipan manfaat dan kerugian 785 dan 23, dari 28 dan 4 artikel
Tabel 9 Manfaat dan kerugian C. peltata bagi manusia dan lingkungan
Manfaat Kerugian
Dampak bagi lingkungan Dampak bagi lingkungan Sumber makanan bagi herbivora
(kelelawar2, larva serangga3,
monyet black howler5)
Vektor infestasi Trichuris spp1 dan Controrchis spp4 terhadap monyet black howler, menggantikan Musanga cecropioides6 dan resiko
invasi7
Obat tradisional dan manfaat farmakologi
Antibakteri8, antidiabetes9,10
(kontrol glikemik), antifungi11,