UJI KETAHANAN 48 GALUR PADI TERHADAP PENYAKIT BLAST
(
Pyricularia oryzae
) RAS 001
YULITA FITRIANNA
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
YULITA FITRIANNA. Uji Ketahanan 48 Galur Padi terhadap Penyakit Blast (Pyricularia oryzae) Ras 001. Dibimbing oleh UTUT WIDYASTUTI dan HAJRIAL ASWIDINNOOR.
Penyakit blas yang disebabkan oleh cendawan Pyricularia oryzae Cav. merupakan salah satu penyakit utama yang menyerang padi di Indonesia. Penelitian ini bertujuan untuk menguji tingkat ketahanan galur-galur padi terhadap serangan penyakit blas dan mengidentifikasi keberadaan gen ketahanan Pi-ta pada galur padi yang tahan. Penelitian dilakukan dengan menggunakan padi varietas Kencana bali (peka) dan Cisadane (tahan) sebagai kontrol serta 48 galur padi hasil seleksi Program Pemuliaan di Departemen Agronomi dan Hortikultura IPB dengan metode injeksi menggunakan P. oryzae ras 001. Padi berumur 16 hari setelah tanam diinokulasikan dengan 1 ml suspensi spora (104-105 spora/ml) isolat P. Oryzae. Tanaman diinkubasi pada kondisi gelap lalu dipindah ke ruang terang dan dilakukan pengamatan hingga 7-9 hari setelah infeksi. Pengujian tahan blas dilakukan 2 tahap, yaitu penapisan I dilakukan sebanyak 3 ulangan dan penapisan II dilakukan sebanyak 2 ulangan untuk masing-masing galur terpilih dari penapisan I, dengan 10 tanaman pada tiap ulangan. Pengamatan hingga penapisan II menunjukkan adanya 13 galur padi tahan terhadap P. oryzae ras 001 yaitu IPB140-F-2-1, IPB115-F-4-2-2, IPB113-F-2-2, IPB107-F-7-3, IPB140-F-5, IPB149-F-2, IPB149-F-8, Martapura, IPB149-F-4, IPB149-F-5, IPB113-F-1, IPB102-F-92-1-1, IPB107-F-5-1 dengan persentase ketahanan ≥ 80 %. Dua pasang primer spesifik untuk gen Pi-ta didesain dari sekuen DNA dengan nomor aksesi AF207842 untuk mengamplifikasi fragmen DNA gen Pi-ta dengan metode PCR. Hasil identifikasi gen ketahanan Pi-ta menunjukkan bahwa 13 galur tahan maupun 2 galur rentan (18-4-2 dan IPB107-F-16E-6) mempunyai gen Pi-ta, bahkan pada Kencana Bali. Percobaan sebelumnya, menunjukkan bahwa Kencana Bali tidak memiliki gen ketahanan Pi-ta. Diduga bahwa varietas Kencana Bali mengalami delesi di daerah bagian belakang ekson 2. Hasil identifikasi gen Pi-ta juga mengindikasikan gen Pi-ta tidak spesifik berperan dalam mekanisme pertahanan tanaman pada galur-galur padi ini, sehingga diduga ada gen lain yang berperan dalam proses ketahanan tanaman padi terhadap serangan P. oryzae ras 001.
Kata kunci: Padi (Oryza sativa), blas, Pyricularia oryzae, resisten
ABSTRACT
YULITA FITRIANNA. Resistance Test toward of 48 Lines Rice to Blast Disease (Pyricularia oryzae) Race 001. Supervised by UTUT WIDYASTUTI and HAJRIAL ASWIDINNOOR.
Blast disease, caused by Pyricularia oryzae Cav., is one of the major fungal diseases infected rice (Oryza sativa L.) in Indonesia. The aim of this research was to examine the resistance of rice from the infection of P. Oryzae race 001 and to detect the existance of Pi-ta gene in resistant lines of rice. Resistance evaluation to blast disease (Pyricularia oryzae) on Cisadane (resistant), Kencana Bali (susceptible) as control, and 48 lines of rice was carried out in the green house by using injection inoculated method with P. oryzae race 001. Rice seedling were planted on sterilize soil, and sixteen days old of rice were soaked in the fungal spore suspension (105 spores/ ml) in the dark for 24 hours, and moved to the light. Rice disease was observered on 7-9th days after infection. There were 2 step screening of blast-resistant, which screening I performed a total of three replicates and screening II performed from selected plant in screening I a total of two replicates for each line of rice, with 10 plants in each replication. The result of second screening at green house showed that 13 lines resistant to P. oryzae 001 were IPB140-F-2-1, IPB115-F-4-2-2, IPB113-F-2-2, IPB107-F-7-3, IPB140-F-5, IPB149-F-2, IPB149-F-8, Martapura, IPB149-F-4, IPB149-F-5, IPB113-F-1, IPB102-F-92-1-1, IPB107-F-5-1 with the percentage of resistance ≥ 80 %. Two pairs of DNA primers specific to the dominant indica Pi-ta gene were designed from DNA sequence in gene bank with accession number AF207842 to amplify the Pi-ta DNA fragments by polymerase chain reaction (PCR) method. The results revealed that all of 13 resistant and 2 susceptible lines (IPB107-F-18-4-2 and IPB107-F-16E-6) of rice carrying Pi-ta genes, even in Kencana Bali. The previous experiment shown that Kencana Bali does not have Pi-ta resistance gene. It is expected that Kencana Bali has deletion of Pi-ta resistance gene in the end part of exon 2. Further more, those results also indicated that Pi-ta gene did not confer resistance to race 001 of Pyricularia oryzae. Resistance to P. oryzae race 001 might be controlled by different resistant gene.
UJI KETAHANAN 48 GALUR PADI TERHADAP PENYAKIT BLAST
(
Pyricularia oryzae
) RAS 001
YULITA FITRIANNA
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul
: Uji Ketahanan 48 Galur Padi terhadap Penyakit Blast (
Pyricularia
oryzae
) Ras 001
Nama
: Yulita Fitrianna
NIM
: G34062968
Disetujui :
Diketahui :
Tanggal lulus :
Pembimbing 1,
Dr. Ir. Utut Widyastuti, M.Si.
NIP 19640517 198903 2001
Pembimbing 2,
Dr. Ir. Hajrial Aswidinnoor, M.Sc.
NIP 19590929 198303 1008
Ketua Departemen Biologi,
Dr. Ir. Ence Darmo Jaya Supena, M.Si.
PRAKATA
Alhamdulillah, puji dan syukur penulis panjatkan kepada Allah SWT atas rahmat, nikmat dan inayah-Nya sehingga penulis dapat menyelesaikan karya ilmiah. Tema yang dipilih pada karya ilmiah ini ialah mengenai penyakit pada tanaman padi, dengan judul Uji Ketahanan 48 Galur Padi terhadap Penyakit Blast (Pyricularia oryzae) Ras 001. Karya ilmiah ini dilaksanakan pada bulan Februari 2010 sampai Maret 2011.
Terima kasih penulis sampaikan kepada Ibu Dr. Ir.Utut Widyastuti, M.Si. dan Bapak Dr. Ir. Hajrial Aswidinnoor, M.Sc. selaku pembimbing atas semua ilmu, kesabaran, bimbingan, nasihat dan saran yang diberikan selama ini. Terima kasih kepada Dr. Ir. Miftahudin, M.Si. selaku penguji atas setiap sarannya. Terima kasih kepada proyek I-MHERE atas nama Dr. Ir. Utut Widyastuti Suharsono, M.Si. yang telah membiayai penelitian ini. Terima kasih kepada direktur dan staf PPSHB atas fasilitas, sarana dan prasarana yang diberikan selama penulis melakukan penelitian. Terima kasih kepada Ibu Anggiani Nasution dan Pak Santoso atas ilmu yang telah diberikan. Terima kasih penulis ucapkan kepada bapak dan ibu di Depok atas semua dukungan, doa, cinta dan kasih sayangnya. Terima kasih kepada saudari-saudariku, kak Dina Riski dan kak Windy Hadiana, yang tak henti memberi semangat dan dukungan agar penelitian ini cepat terselesaikan. Terima kasih kepada keluarga besar Bapak H. Natsir atas semua doa dan kasih sayangnya. Terima kasih kepada teman-teman seperjuangan Biologi 43. Terima kasih kepada saudari-saudariku Pondok Sugih crew (sugihers) atas kebersamaan, keceriaan, semangat dan doanya. Terima kasih kepada rekan-rekan peneliti di Laboratorium Biorin dan Biologi Molekuler Seluler Tanaman (Indah Khayati, Hayatul Fajri, Laila Nur Syafitri, Iin Premitasari, Pak Muzuni, Pak Yustinus Ulung Anggraito, Pak Radite Tistama, Bu Saleha Hanum, Bu Ratna Yuniati, Mbak Ulfah Mushofa, Kak Ika Atifah Zahroh, Kak Goto Giok, Kak Luria Marlina Limbong, Bu Dini Tasuruni, Mbak Anita Theresia, Kak Nurul Fitriah, Kak Novita Andriany, Kak Medikca Tanjung, dan Kak Muchdar Davis) atas kerjasama, semangat, saran, bantuan, canda, kasih sayang, rasa kekeluargaan dan kebersamaan yang akan selalu terkenang di hati. Terima kasih kepada Pak Yulianto, Mbak Pepi Elvavina, Pak Abdul Mulya, Pak Adi Supardi, Pak Yusman, Pak Itar Suherman dan Pak Edi atas semua bantuan dan dukungannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2011
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 16 Mei 1988 sebagai anak ketiga dari tiga bersaudara pasangan bapak Yusuf Hadi dan Ibu Rohana. Penulis menyelesaikan pendidikan dasar di SDN Depok Baru 3 pada tahun 2000. Tahun 2003 penulis menyelesaikan pendidikan menengah pertama di SLTP Negeri 2 Depok. Dan pada tahun 2006 penulis menyelesaikan pendidikan sekolah menengah atas di SMA Negeri 1 Depok. Penulis diterima di Institut Pertanian Bogor di tahun yang sama melalui jalur Undangan Seleksi Masuk IPB (USMI).
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... viii
PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian ... 2
BAHAN DAN METODE Waktu dan Tempat ... 2
Bahan ... 2
Metode ... 2
HASIL Morfologi dan Perkecambahan Spora ... 3
Kemampuan Infeksi Penyakit Blas pada Padi ... 3
Identifikasi Keberadaan Gen Pi-ta ... 5
PEMBAHASAN Penyakit Blas Daun pada Padi ... 6
Identifikasi Keberadaan Gen Pi-ta ... 7
SIMPULAN ... 8
SARAN ... 8
DAFTAR PUSTAKA ... 9
DAFTAR TABEL
Halaman
1 Primer spesifik gen Pi-ta ... 3
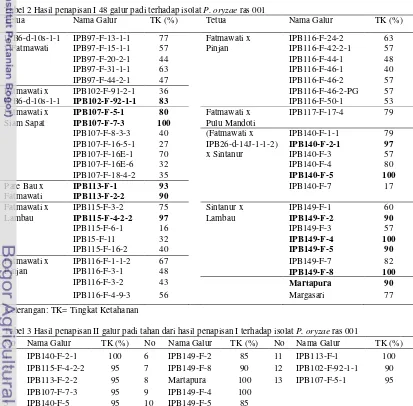
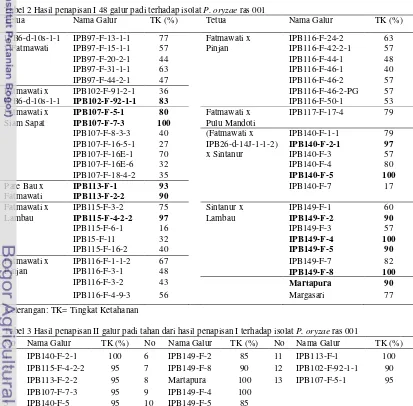
2 Hasil penapisan I 48 galur padi harapan terhadap isolat P. oryzae ras 001 ... 5
3 Hasil penapisan II galur padi harapan tahan dari hasil penapisan I ……….. 5
DAFTAR GAMBAR
Halaman 1 Perkecambahan spora P. grisea pada media kaca ... 42 Bercak blas pada daun padi ... 4
3 Hasil elektroforesis identifikasi gen Pi-ta ... 6
4 Daerah penempelan primer dan daerah dugaan delesi ... 8
DAFTAR LAMPIRAN
Halaman 1 Skala penyakit blas berdasarkan sistem evaluasi standar IRRI (1996) ... 122 Data morfologi dan panen galur padi tahan ... 13
PENDAHULUAN
Latar Belakang
Tanaman padi (Oryza sativa L.) di Indonesia banyak terserang oleh bermacam-macam penyakit (Semangun 1991). Salah satu penyakit yang sangat merugikan petani adalah penyakit blas. Penyakit blas yang disebabkan oleh cendawan Pyricularia oryzae Cav. masih merupakan masalah utama pada padi gogo (Rais et al. 2001). Penyakit blas sebelumnya diketahui hanya menyerang padi gogo (Semangun 1991), tetapi saat ini penyebaran penyakit blas sudah mulai menyerang pertanaman padi sawah (Amir et al. 2000). Serangan penyakit blas di Indonesia diperkirakan mencapai 19.629 hektar dari total luas lahan pertanaman padi Indonesia sebesar 12.833.578 hektar pada tahun 2009 (BPS 2010). Kehilangan hasil pada varietas yang peka seperti Bical dapat mencapai 50-90% (Rais et al. 2001).
Secara umum P. oryzae menyerang tanaman padi pada bagian daun, buku, leher, malai, cabang malai, bulir padi, kulit gabah (Chen 1993; Rais et al. 2001), dan collar daun (Scardaci et al. 1997). Gejala serangan penyakit blas mempunyai bentuk khas, gejala daun yang terserang oleh blas daun (leaf blast) berupa bercak berbentuk belah ketupat sampai elips dengan kedua ujungnya runcing. Bagian tengah bercak berwarna abu-abu atau putih keabu-abuan, pada bagian tepi berwarna coklat sampai coklat kemerahan dengan ukuran panjang bercak 1-1.5 cm dan lebar 0.3-0.35 cm (Ou 1985; Scardaci et al. 1997).
Bentuk, warna, dan ukuran bercak dapat beragam bergantung pada ketahanan varietas, umur tanaman, dan umur lesio. Bercak tidak akan berkembang dan tetap seperti titik kecil pada varietas yang tahan dan bercak akan berkembang sampai beberapa milimeter berbentuk bulat atau elips dengan tepi berwarna coklat pada varietas dengan reaksi sedang (Ou 1985). Blas daun kadang-kadang menyebabkan kematian keseluruhan pada tanaman yang masih muda sampai stadia pembentukan anakan atau tillering (Scardaci et al. 1997).
Menurut Amir et al. (2000) ras 001, 003, 033, dan 173 merupakan ras yang ada pada setiap musim tanam, artinya isolat ini memiliki penyebaran yang luas. Isolat ras 001 mempunyai tingkat patogenesitas yang paling rendah virulensinya, menyebar luas dan mampu bertahan lama di lapangan (Sari 2008; Utami 2000). Adapun penentuan ras baru dari patogen blas di Indonesia dilakukan dengan
menggunakan varietas diferensial yaitu Asahan, Cisokan, IR64, Krueng Aceh, Cisadane, Cisanggang, dan Kencana Bali (Mogi et al. 1991).
Studi genetika untuk mendapatkan gen resisten terhadap penyakit blas telah banyak dilakukan. Pada tanaman padi telah diidentifikasi sekitar 40 gen mayor yang mengendalikan ketahanan terhadap penyakit blas dan 5 gen resisten, Pi-b, Pi-ta, Piz-5(Pi-2), Piz-t, dan Pi9 telah berhasil diisolasi (Wang et al. 1999; Qu et al. 2006; Zhou et al. 2006). Menurut Ou (1985), gen-gen ketahanan tersebut (Pi) akan berinteraksi secara spesifik dengan ras-ras tertentu dari cendawan Pyricularia grisea.
Menurut Suwarno et al. (2001) sumber gen Pi banyak terdapat pada gene pool padi Indonesia baik yang berasal dari varietas budidaya maupun spesies padi liar. Dua puluh delapan genotip padi indica yang terdapat di Indonesia telah dianalisis oleh Santoso (2005) dan ditemukan bahwa banyak padi indica yang mempunyai gen ketahanan baik Pi-b maupun Pi-ta. Gen Pi-b dan Pi-ta merupakan 2 gen mayor resisten blas hasil introgresi dari kultivar padi indica. Kedua gen resisten tersebut mempunyai struktur gen yang mirip, yaitu NBS (nucleotide binding site) dan LRR (leucine rich repeat) dan di duga menyandikan protein sitoplasmik yang bertanggung jawab merespon gen avirulen AVR-Pita, memicu resisten ras spesifik (Wang et al. 1999; Bryan et al. 2000; Sari 2008).
Tujuan Penelitian
Penelitian ini bertujuan untuk menguji tingkat ketahanan galur-galur padi terhadap serangan penyakit Blas (Pyricularia oryzae) ras 001 dan mengidentifikasi keberadaan gen ketahanan Pi-ta pada galur padi yang tahan.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Februari 2010 – Maret 2011, bertempat di laboratorium BIORIN dan Rumah Kaca Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB), Institut Pertanian Bogor.
Bahan dan Alat
Bahan tanaman yang digunakan yaitu 48 galur padi hasil seleksi Program Pemuliaan di Departemen Agronomi dan Hortikultura IPB, varietas peka (Kencana Bali) dan varietas tahan (Cisadane) sebagai kontrol, dan isolat cendawan P. oryzae ras 001. Bahan kimia yang digunakan yaitu media oatmeal, Potato Dextrose Agar (setiap liter mengandung 39 g PDA dan supaya lebih padat ditambah 5 g agar), antibiotik kloramfenikol, aquades, larutan Tween 0.025% (v/v), nitrogen cair, buffer ekstrak 2% (w/v) (dengan komposisi 2% CTAB (w/v), 0.1 M Tris HCl pH 8.0, 1.5
M NaCl, 0.02 M EDTA), 2% β-mercapto etanol, kloroform, isoamil alkohol, phenol, Na acetat 3.2 M, alkohol absolut, RNAse 10 µg/µl, aquabides, gel agarosa konsentrasi 1% (b/v), larutan TAE 1X, loading dye, dan ethidium bromida 0.5 mg/L.
Alat yang digunakan yaitu autoklaf, laminar, mikroskop, hemasitometer, magnetik stirrer, mikropipet berbagai ukuran, sentrifuse Jouan BR-4i, shaker, cawan petri, jarum ose, mesin PCR, dan alat elektroforesis.
Metode penelitian
Uji Ketahanan Tanaman Padi
Penyiapan Tanaman Padi. Benih padi dicuci kemudian dikecambahkan dalam cawan petri yang berisi kertas merang selama kurang lebih empat hari lalu ditanam ke media tanah. Masing-masing galur ditanam sebanyak 10 benih dan dibuat sebanyak 3x ulangan.
Produksi Spora. Potongan miselium isolat cendawan P. oryzae ras 001 asal padi diinokulasikan pada media oatmeal (30 g oat/l, 20 g agar/l, 5 g sukrosa/l), dan diinkubasi selama 10-14 hari pada suhu ruang, lalu kultur dimaserasi atau dicuci secara aseptik untuk menghilangkan hifa aerial. Kultur disinari n-UV (300-400 nm) terus menerus selama
kurang lebih enam hari untuk menginduksi pembentukan spora. Sebanyak 1-3 ml air steril yang mengandung 0.025% (v/v) Tween 20 disiramkan pada permukaan koloni cendawan. Permukaan koloni cendawan diusap-usap dengan bantuan kaca objek steril untuk melepaskan spora dari konidiofor. Spora-spora dari isolat yang sama dipanen dan dikumpulkan kemudian dilakukan pengukuran kerapatan spora dengan hemasitometer hingga diperoleh suspensi spora 104-105 spora/ml (Chao & Ellingboe 1997).
Uji Ketahanan terhadap Infeksi. Benih padi yang telah berkecambah ditanam pada bak berukuran 25x35x15 cm yang diberi tray berisi campuran tanah dan pupuk dengan perbandingan 10 kg tanah, 2.5 gr urea, 1.5 gr TSP, dan 1.2 gr KCl. Tanah yang digunakan adalah tanah yang telah disterilkan. Masing-masing galur ditanam sebanyak 10 benih dan dibuat sebanyak 3x ulangan. Selanjutnya, padi berumur 16 hari setelah tanam (memiliki ±5 daun) diinokulasikan dengan 1 ml suspensi spora (104-105 spora/ml) isolat P. Oryzae melalui penyuntikan di bagian pelepah daun kelima. Sebagai kontrol negatif, padi disuntik menggunakan akuades (tidak diinfeksi). Selanjutnya semua tanaman diinkubasi pada kondisi gelap dan lembab selama 24 jam, kemudian dipindahkan ke ruang terang selama enam hari pada suhu 25-30˚C dan kelembaban 70-90 %. Pengamatan kemampuan infeksi terhadap inang berupa kemunculan bercak pada daun dilakukan setiap hari selama kurang lebih 7-9 hari setelah infeksi. Tingkat ketahanan diukur dengan menggunakan tabel skala penyakit yang ditentukan oleh IRRI (Lampiran 1). Uji ketahanan terhadap infeksi dilakukan sebanyak 2 tahap penapisan (Screening). Penapisan I dilakukan sebanyak 3 ulangan untuk masing-masing galur dengan 10 tanaman pada setiap ulangan. Setelah didapatkan galur padi dengan nilai persentase ketahanan terhadap blas ≥ 80% dilakukan tahap penapisan II yang dilakukan sebanyak 2 ulangan untuk masing-masing galur dengan 10 tanaman pada setiap ulangan. Persentase akhir ketahanan merupakan rata-rata persentase ketahanan dari kedua ulangan. Persentase
ketahanan ≥ 80% digolongkan menjadi tanaman resisten (tahan).
Identifikasi Keberadaan Gen Ketahanan Pi-ta pada Galur Padi Tahan Blas
– 0.2 g lalu ditambahkan 600 µL buffer ekstrak (dengan komposisi 2% Cetyl Trimethyl Ammonium Bromide [CTAB] (w/v), 0.1 M Tris HCl pH 8.0, 1.5 M NaCl, 0.02 M Ethylene Diamine Tetra acetic Acid [EDTA] ), dan mercapto etanol 0,2%. Campuran dikocok, diinkubasi selama 30 menit pada suhu 650C. Campuran dalam tabung eppendorf didinginkan di es selama 5 menit, kemudian ditambahkan 600 µL kloroform : isoamil alkohol (24:1). Tabung eppendorf kemudian disentrifuse pada 10000 rpm (g = ± 9.8 m/s2) dengan suhu 40C, selama 10 menit. Supernatan bagian atas (mengandung DNA) diambil dan dipindahkan pada tabung ependorf lain kemudian ditambahkan PCI (Phenol, Chloroform, dan Isoamil alkohol) dingin dengan perbandingan (25:24:1) sebanyak 500 µL, disentrifuse kembali pada 10000 rpm (g = ± 9.8 m/s2) dengan suhu 40C selama 5 menit. Supernatan diambil kemudian ditambahkan 2M NaOAC pH 5,2 (0.1x volume supernatan) dan EtOH 100% sebanyak 2x volume supernatan. Presipitasi dilakukan selama 30 menit dalam freezer. Pelet diambil dan dibilas dengan 500 µL etanol 70% lalu disentrifuse 5 menit dengan kecepatan 10000 rpm (g = ± 9.8 m/s2). Cairan dibuang dan pelet dikeringkan dengan vakum. Pelet dilarutkan ke dalam 20-50 µL aquades steril, diberi RNAse (10 µg/µl) sebanyak 0.2x volume aquades steril selama 30 menit pada suhu 370C. Tahap terakhir yaitu inaktif RNAse pada suhu 700C, selama 10 menit.
Uji Kualitas dan Kuantitas DNA. Uji kualitas dan kuantitas DNA dilakukan menurut metode Sambrook et al. (1989) dengan beberapa modifikasi. DNA padi yang berhasil diisolasi dielektroforesis pada gel agarosa dengan konsentrasi 1% (b/v) dalam buffer penyangga 1x TAE. Perbandingan DNA dengan loading dye adalah 5:2 µL. Elektroforesis dilakukan selama kurang lebih 30 menit pada tegangan 100 volt. Selanjutnya gel agarose hasil elektroforesis dimasukkan ke dalam larutan ethidium bromida (konsentrasi larutan 0.5 mg/L) selama 5 menit. Panjang pita DNA diamati dengan melihat pita DNA pada UV transiluminator.
Identifikasi Gen Pi-ta. Amplifikasi DNA guna mengidentifikasi gen ketahanan Pi-ta dilakukan dengan bantuan Polymerase Chain Reaction (PCR). Primer yang digunakan adalah Pi-ta ekson 1 dan Pi-ta ekson 2. Primer spesifik ini didesain dari sekuen DNA pada database dengan nomor kode aksesi AF207842. Primer spesifik Pi-ta ekson 1 digunakan untuk amplifikasi gen Pi-ta di
ekson 1 dengan ukuran produk PCR sebesar 418 bp. Sedangkan, primer spesifik Pi-ta ekson 2 digunakan untuk amplifikasi gen Pi-ta di ekson 2 dengan ukuran produk PCR sebesar 448 bp.
Tabel 1 Primer spesifik gen Pi-ta Primer spesifik Pi-ta ekson 1
forward 5’ACTGCTGGTGCCAAGAAGAT3’
reverse 5’GGCCATGCAGACGATAGAAT3’
Primer spesifik Pi-ta ekson 2
forward 5’CCCAGGATGACCTTGACACT3’
reverse 5’TGTGCCAAATCTTCATCCAA3’
Reaksi PCR mengikuti metode yang dilakukan Naqvi dan Chattoo (1996). PCR dilakukan dengan total reaksi 10µl mengandung 100 ng DNA genomik cetakan (1 µl), dNTPmix (dATP, dCTP, dGTP, dTTP) 0.2 mM (1 µl), masing-masing primer (forward dan reverse) 1 pmol (masing-masing sebanyak 0.4 µl), enzim taq DNA polymerase 1 unit (0.2 µl) dalam larutan buffer 1X (1 µl) dan ditambahkan air bebas ion hingga mencapai volume 10 µl. Amplifikasi PCR dilakukan sebanyak 30 siklus. Program PCR terdiri dari Pra-denaturasi pada suhu 940C selama 5 menit, dilanjutkan dengan 30 siklus yang terdiri atas denaturasi pada suhu 940C ,1 menit, annealing pada suhu 550C, 30 detik, dan extension pada suhu 720C, 1 menit. Tahap akhir proses PCR yaitu final extension pada suhu 720C selama 5 menit dan pendinginan pada suhu 720C selama 10 menit. Produk PCR kemudian dipisahkan dengan elektroforesis pada gel agarosa 1% dalam larutan buffer penyangga 1x TAE. DNA 1 kb (ladder) diletakkan di sumur pertama untuk mengukur panjang pita-pita DNA yang dihasilkan. Proses elektroforesis dilakukan selama 26 menit pada tegangan 100 volt. Kemudian diletakkan dalam larutan ethidium bromida (0.5 mg/L) selama 5 menit lalu dibilas dengan aquades. Hasil pita DNA pada agarose divisualisasi dengan cahaya UV. Ada tidaknya gen Pi dapat diketahui dengan melihat pita DNA yang terkait dengan fragment gen Pi yang dapat diamplifikasi dengan primer spesifik.
HASIL DAN PEMBAHASAN
HASIL
Konsentrasi rata-rata spora yang dihasilkan adalah 105 sampai 5.105 sel/ml suspensi. Hasil pengamatan spora dengan mikroskop menunjukkkan bahwa spora P. oryzae berbentuk menyerupai buah pir, memiliki 2 sekat, dan berwarna hialin (Gambar 1a). Guna mengetahui kemampuan infeksi spora P. oryzae yang diinokulasi pada tanaman padi maka dilakukan juga pengamatan perkecambahan spora yang membentuk apresorium. Pengamatan terhadap kemampuan perkecambahan spora memperlihatkan bahwa spora mulai berkecambah pada jam ke empat setelah panen, selanjutnya terbentuk tabung kecambah yang muncul dari bagian apikal atau basal dari sel spora (Gambar 1b). Satu spora dapat membentuk lebih dari satu tabung kecambah. Tabung kecambah selanjutnya membentuk struktur apresorium pada bagian ujungnya. Apresorium berbentuk bulat dan
membentuk tabung infeksi yang berfungsi sebagai struktur penetrasi ke tanaman inang (Gambar 1c).
Kemampuan Infeksi Penyakit Blas Pada Tanaman Padi
Varietas padi Kencana Bali merupakan varietas peka yang digunakan sebagai kontrol positif pada penelitian ini menunjukkan gejala penyakit blas pada bagian daun. Bercak pada varietas Kencana bali mulai tampak pada hari ke empat setelah infeksi dan bercak bertambah lebar setelah hari ke tujuh. Gejala blas daun yang teramati yaitu berupa bercak berbentuk elips dengan ujung runcing, tepi bercak berwarna coklat tua hingga hitam dengan sedikit warna kuning, bagian tengah berwarna abu-abu (Gambar 2). Bercak hanya berupa bintik-bintik hitam dan tidak terlihat melebar pada tanaman tahan (resisten).
Gambar 1 (a) Spora P. grisea pada jam ke-0 setelah panen, perbesaran 10x40; (b) Spora mulai berkecambah pada jam ke-4 setelah panen, perbesaran 10x10; (c) Apresorium pada jam ke-20 setelah panen perbesaran 40x10.
Varietas/ Galur Ketahanan 0 hsi 5 hsi 7 hsi
Kencana bali
Cisadane 100%
140-5 100%
102-92-1-1 80%
Gambar 2 Serangan blas pada hari ke-0, ke-5 dan ke-7 setelah infeksi.
0.5 cm 0.5 cm 0.5 cm
0.5 cm 0.5 cm
0.5 cm
0.5 cm 0.5 cm
0.5 cm
0.5 cm 0.5 cm 0.5 cm
bercak
bercak Apresorium
tabung kecambah
c
spora
10 µm spora
tabung kecambah
b
30 µmspora
20 µm
Hasil penapisan I atau uji di rumah kaca terhadap 48 galur padi menunjukkan bahwa terdapat 15 galur padi yang resisten terhadap serangan isolat Pyricularia oryzae ras 001 (Tabel 2). Tiga belas galur tanaman padi resisten dipilih dari kelima belas galur tersebut berdasarkan nilai tingkat ketahanan ≥ 80 % kemudian diverifikasi kembali untuk memastikan tingkat ketahanannya.
Hasil verifikasi atau penapisan II menunjukkan bahwa terdapat 13 galur padi
yang memiliki tingkat ketahanan ≥ 80%
berdasarkan munculnya bercak (lesio) pada tanaman uji saat hari ke-7 setelah infeksi (Tabel 3). Tiga belas galur yang memiliki
tingkat ketahanan ≥ 80% setelah verifikasi
kembali yaitu IPB140-F-2-1, IPB115-F-4-2-2, IPB113-F-2-2, IPB107-F-7-3, IPB140-F-5, IPB149-F-2, IPB149-F-8, Martapura, IPB149-F-4, IPB149-F-5, IPB113-F-1, IPB102-F-92-1-1, dan IPB107-F-5-1. Ketiga
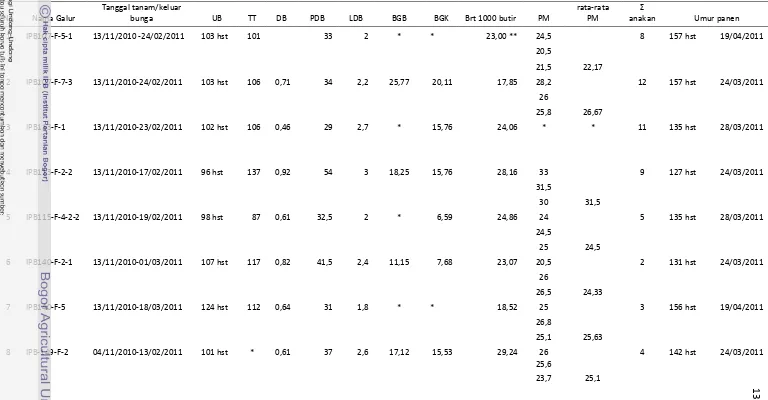
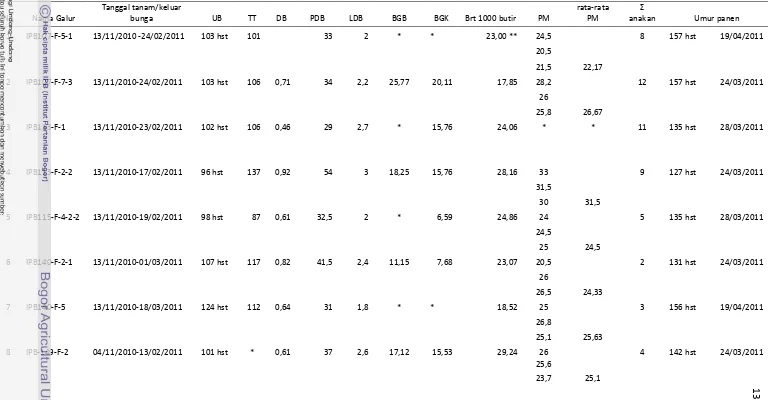
belas galur tersebut kemudian diambil dan menjadi sampel isolasi DNA. Selain itu, pengamatan terhadap morfologi galur padi tahan juga dilakukan dengan mengamati satu rumpun padi hingga tahap panen kemudian dilakukan pengambilan gambar morfologi tanaman padi (Lampiran 2 dan 3).
Identifikasi Keberadaan Gen Pi-ta
Tiga belas galur tahan dan dua galur rentan diambil kemudian DNA tanaman diisolasi. Hasil isolasi DNA dikuantifikasi dan dilakukan pengujian kemurnian DNA dengan spektrofotometer pada panjang gelombang 260 dan 280. Rasio OD260/OD280 yang diperoleh berkisar antara 1.44 hingga 2.22. Amplifikasi PCR dilakukan dengan menggunakan 2 primer yang didesain untuk menguji keberadaan gen Pi-ta. Sekuen DNA gen Pi-ta diketahui memiliki dua daerah ekson yang mengapit satu daerah intron.
Tabel 2 Hasil penapisan I 48 galur padi terhadap isolat P. oryzae ras 001
Tetua Nama Galur TK (%) Tetua Nama Galur TK (%)
IPB6-d-10s-1-1 IPB97-F-13-1-1 77 Fatmawati x IPB116-F-24-2 63
x Fatmawati IPB97-F-15-1-1 57 Pinjan IPB116-F-42-2-1 57
IPB97-F-20-2-1 44 IPB116-F-44-1 48
IPB97-F-31-1-1 63 IPB116-F-46-1 40
IPB97-F-44-2-1 47 IPB116-F-46-2 57
Fatmawati x IPB102-F-91-2-1 36 IPB116-F-46-2-PG 57
IPB6-d-10s-1-1 IPB102-F-92-1-1 83 IPB116-F-50-1 53
Fatmawati x IPB107-F-5-1 80 Fatmawati x IPB117-F-17-4 79
Siam Sapat IPB107-F-7-3 100 Pulu Mandoti
IPB107-F-8-3-3 40 (Fatmawati x IPB140-F-1-1 79
IPB107-F-16-5-1 27 IPB26-d-14J-1-1-2) IPB140-F-2-1 97
IPB107-F-16E-1 70 x Sintanur IPB140-F-3 57
IPB107-F-16E-6 32 IPB140-F-4 80
IPB107-F-18-4-2 35 IPB140-F-5 100
Pare Bau x IPB113-F-1 93 IPB140-F-7 17
Fatmawati IPB113-F-2-2 90
Fatmawati x IPB115-F-3-2 75 Sintanur x IPB149-F-1 60
Lambau IPB115-F-4-2-2 97 Lambau IPB149-F-2 90
IPB115-F-6-1 16 IPB149-F-3 57
IPB15-F-11 32 IPB149-F-4 100
IPB115-F-16-2 40 IPB149-F-5 90
Fatmawati x IPB116-F-1-1-2 67 IPB149-F-7 82
Pinjan IPB116-F-3-1 48 IPB149-F-8 100
IPB116-F-3-2 43 Martapura 90
IPB116-F-4-9-3 56 Margasari 77
Keterangan: TK= Tingkat Ketahanan
Tabel 3 Hasil penapisan II galur padi tahan dari hasil penapisan I terhadap isolat P. oryzae ras 001
No Nama Galur TK (%) No Nama Galur TK (%) No Nama Galur TK (%)
1 IPB140-F-2-1 100 6 IPB149-F-2 85 11 IPB113-F-1 100
2 IPB115-F-4-2-2 95 7 IPB149-F-8 90 12 IPB102-F-92-1-1 90
3 IPB113-F-2-2 95 8 Martapura 100 13 IPB107-F-5-1 95
4 IPB107-F-7-3 95 9 IPB149-F-4 100
5 IPB140-F-5 95 10 IPB149-F-5 85
Pi-ta diduga menyandikan protein sitoplasmik dengan lokasi sentral NBS dan daerah pemotongan tinggi LRR (yang selanjutnya disebut LRD) pada daerah terminal carboxyl yang cocok mengenali gen avirulence AVR-Pita, sehingga dapat menggerakkan resisten ras spesifik. Substitusi asam amino tunggal, serine (Ser) ke alananin (Ala) pada posisi 918, pada LRD protein Pita mendemonstrasikan untuk menetapkan secara langsung interaksi dengan AVR-Pita dan spesifik resisten untuk patogen blas yaitu Magnaporthe oryzae (Bryan et al. 2000).
Hasil elektroforesis dari amplifikasi DNA dengan menggunakan primer Pi-ta ekson 1 dan Pi-ta ekson 2 menunjukkan adanya pita yang terbentuk dengan ukuran di antara 400-500 bp. Pita yang terbentuk berukuran sesuai dengan target yang diinginkan, hal itu menandakan bahwa gen Pi-ta yang menjadi target telah berhasil teramplifikasi.
PEMBAHASAN
Penyakit Blas Daun pada Padi
Galur padi yang digunakan merupakan galur hasil persilangan dari varietas dan galur tetua terpilih. Varietas Sintanur, Pare bau, Lambau, dan Pinjan termasuk dalam padi aromatik yang banyak ditanam di Sulawesi Selatan, sedangkan varietas Fatmawati memiliki keunggulan karena tingkat kerontokan gabah rendah hingga mengurangi kehilangan hasil pada saat panen (Masniawati 2005). Galur IPB6-d-10s-1-1 memiliki keunggulan karena bulir padi yang dimiliki terisi penuh dari pangkal hingga ujung malai (Dr. Hajrial 20 Januari 2010, komunikasi pribadi). Jika dilihat dari segi ketahanan terhadap penyakit, varietas Sintanur dan Fatmawati memiliki ketahanan terhadap Hawar Bakteri Daun (HBD) strain III.
Sedangkan, informasi mengenai ketahanan akan blas pada varietas tetua belum diketahui, sehingga dilakukan pengujian tahan blas.
Penyakit blas disebabkan oleh cendawan Pyricularia oryzae, cendawan ini termasuk dalam kelompok Ascomycetes dan ditemukan di alam dalam bentuk aseksualnya saja. Sedangkan bentuk seksualnya (teleomorf) Magnaporthe grisea (Hebert) Barr hanya dihasilkan dengan pengkulturan di laboratorium (Agrios 1997). Pyricularia oryzae mempunyai konidiofor bersekat-sekat jarang bercabang, berwarna kelabu, membentuk konidium pada ujungnya, konidium berbentuk bulat telur dengan ujung runcing, jika masak bersekat 2, dengan ukuran 20-22 x 10-12 µm.
Gejala blas daun diamati pada daun berupa bercak berbentuk belah ketupat berujung runcing. Bentuk warna dan ukuran lesio atau bercak bervariasi tergantung pada ketahanan varietas, umur tanaman dan lingkungan. Bercak pada varietas yang tidak tahan (rentan) pada kondisi yang lembab memperlihatkan pinggiran coklat dengan sedikit berwarna kuning (halo), sedangkan pada varietas yang tahan bercak hanya berupa bintik coklat sebesar jarum atau lebih (Ou 1985). Selain itu, dilihat dari waktu kemunculan bercak, bercak pada varietas yang peka lebih cepat timbul, dibandingkan dengan varietas yang resisten.
Kemampuan infeksi cendawan dipengaruhi haplotype suatu cendawan. Keragaman haplotip P. oryzae antara lain dipengaruhi oleh faktor lingkungan seperti suhu, kelembaban, baik pada lokasi yang sama maupun berbeda (Reflinur 2005). Suhu yang digunakan pada penelitian ini berkisar antara 26oC-30oC, dengan kelembapan berkisar antara 75 sampai 90%. Hal ini menunjukkan kondisi lingkungan yang Gambar 3 Hasil elektroforesis amplifikasi DNA gen Pi-ta (a) ekson 1 (b) ekson 2 (M) DNA ladder 1 kb (1)
IPB140-F-2-1 (2) IPB115-F-4-2-2 (3) IPB113-F-2-2 (4) IPB107-F-7-3 (5) IPB140-F-5 (6) IPB149-F-2 (7) IPB149-F-8 (8) Martapura (9) IPB149-F-4 (10) IPB149-F-5 (11) IPB113-F-1 (A) Asahan (Kb) Kencana bali* (Ni) Nipon bare (12) IPB102-F-92-1-1 (13) IPB107-F-5-1 (14) IPB107-F-18-4-2* (15) IPB107-F-16E-6* (-) kontrol negatif/ ddH2O. keterangan: *rentan (susceptible).
a.
418 bp M 1 2 3 4 5 6 7 8 9 10 11 A Kb Ni 12 13 14 15 (-)
448 bp b.
mendukung bagi perkembangan penyakit. blas. Spora dihasilkan dan dilepaskan pada kondisi kelembaban yang relatif tinggi, dan tidak ada spora yang dihasilkan pada kondisi kelembaban di bawah 89% (Bonman 1992). Sporulasi berlangsung secara optimum pada suhu 28°C dengan kelembaban 95% dan dalam kondisi gelap selama 15 jam (Kato 1976), sedangkan temperatur optimum untuk perkecambahan spora, pembentukan lesio dan sporulasi adalah 77-82°C (32-35°C) (Scardaci et al. 1997).
Hasil percobaan menunjukkan bahwa gejala penyakit blas pada tanaman rentan mulai muncul pada hari ke-3 dan ke-4 setelah inokulasi. Hal ini sesuai dengan Leung dan Shi (1994) yang menyatakan kelembaban yang tinggi pada tanaman yang rentan akan menyebabkan cendawan P. oryzae menghasilkan konidia dalam waktu 3-4 hari. Sementara, menurut Ou (1985) periode laten penyakit blas di daerah tropis adalah empat sampai lima hari setelah inokulasi. Bercak secara cepat berkembang menjadi lebih besar pada suhu 32°C dan sesudahnya perkembangan bercak akan menurun.
Reaksi tanaman padi terhadap penyakit blas dibagi menjadi tiga yaitu tahan (completely resistant), moderat (partially resistant) dan peka (susceptible). Kencana Bali dan Cisadane digunakan sebagai varietas peka dan tahan mengacu pada Santoso (2005) yang membuktikan bahwa varietas Cisadane merupakan varietas tahan blas, dan varietas Kencana Bali merupakan varietas yang peka dengan intensitas serangan tertinggi di Indonesia (Mogi et al. 1991; Nugraha 2005).
Faktor-faktor yang mempengaruhi ketahanan atau kerentanan suatu varietas padi terhadap cendawan P. oryzae adalah adanya gen ketahanan pada tanaman inang, tingkat virulensi cendawan P. oryzae dan lingkungan (Ou 1985). Varietas Cisadane dan tiga belas galur tahan memperlihatkan gejala adanya blas dengan tingkat serangan yang rendah dan sangat rendah. Hal tersebut diduga karena galur tahan tersebut memiliki kemampuan untuk membatasi penetrasi apresorium blas dan mematikan patogen blas. Deposisi senyawa silikat yang berada dalam jaringan epidermis daun varietas tahan dapat melindungi invasi hifa cendawan secara mekanis (Takahashi 1997; Kim et al. 2002). Selain itu, varietas padi yang tahan juga cenderung menghambat pembentukan spora blas dengan memproduksi fitoaleksin tertentu sebagai akibat interaksi antar patogen
dan tanaman padi (Dillon et al. 1997; Rodrigues et al. 2004). Hal ini dapat pula disebabkan adanya reaksi hipersensitif yang cepat dari tanaman inang sehingga patogen tidak berkembang.
Osburn (1996) menyatakan bahwa sistem pertahanan tanaman terhadap serangan pathogen terdiri dari pertahanan pasif dan pertahanan aktif. Mekanisme pertahanan pasif diantaranya adalah kultikula yang berlilin dan senyawa antimikroba yang bertujuan mencegah terjadinya kolonisasi jaringan oleh cendawan. Sedangkan, aktifnya mekanisme seluler yang terinduksi karena elisitor yang dikeluarkan oleh pathogen merupakan sistem pertahanan aktif, diantaranya adalah terjadi akumulasi metabolit sekunder antimikroba dan ekspresi protein pathogenesis related (Kim et al. 2000).
Hasil pengamatan menunjukkan bahwa kebanyakan daun padi yang terserang adalah daun bagian atas atau daun yang masih muda. Menurut Ou (1985) kerentanan daun padi terhadap infeksi patogen berhubungan dengan kandungan silikat pada dinding sel epidermis daun padi. Sel-sel daun muda mempunyai kandungan silikat yang masih rendah sehingga apresorium dapat menembus dinding sel epidermis daun padi muda.
Isolat yang digunakan dalam penelitian ini adalah isolat P. oryzae ras 001, yang berasal dari koleksi Balai Penelitian Tanaman Padi Muara Bogor. Utami et al. (2000), Amir dan Nasution (2001) dan Utami et al. (2005) menyatakan bahwa isolat 001 adalah isolat yang selalu dijumpai pada setiap musim tanam artinya isolat ini memiliki penyebaran yang luas dan mampu bertahan lama di lapangan. Oleh sebab itu isolat ini selalu dipakai sebagai dasar dalam analisis rata-rata generasi. Ras 001 mempunyai tingkat patogenesitas yang paling rendah virulensinya dibandingkan ras lain (Sari 2008).
Identifikasi Keberadaan Gen Pi-ta
dimana terdapat dua bentuk interaksi, yaitu reaksi inkompatibel dan kompatibel. Interaksi inkompatibel adalah interaksi antara gen resisten (R gene) pada tanaman inang dengan gen avirulen (Avr) pada patogen yang selanjutnya menyebabkan terbentuknya reaksi hipersensitif (HR) pada tanaman inang (Agrios 1997). Sedangkan interaksi kompatibel adalah interaksi antara tanaman inang yang rentan dengan patogen yang virulen hingga menyebabkan timbulnya penyakit (Kurnianingsih 2008). Galur padi resisten dengan tingkat ketahanan 100 % yaitu IPB107-F-7-3, IPB140-F-5, IPB149-F-4 dan IPB1IPB149-F-49-F-8 tidak memperlihatkan gejala blas sampai akhir pengamatan (9 hsi), hal ini mengindikasikan adanya reaksi hipersensitif pada tanaman padi tersebut. Oleh karenanya dapat dikatakan pada keempat galur tersebut terdapat interaksi inkompatibel yaitu interaksi antara R gene pada padi terhadap gen Avirulen (Avr) pada Pyricularia oryzae ras 001.
Perbedaan respon ketahanan dari 48 galur padi yang digunakan diduga disebabkan karena perbedaan genotipe dari varietas tersebut, yaitu adanya perbedaan gen resistensi (gen Pi) yang dimiliki oleh masing-masing galur. Galur yang memiliki ketahanan hingga 100% menunjukkan bahwa tidak ada satupun tanaman yang diinfeksi yang memperlihatkan gejala penyakit blas. Pemilihan 13 galur resisten (tahan) dikarenakan dugaan bahwa galur padi dengan
tingkat ketahanan ≥ 80% berpotensi
membawa gen ketahanan Pi. Gen Pi-ta diduga menyandikan protein sitoplasmik dengan lokasi sentral NBS dan daerah pemotongan tinggi LRR pada daerah terminal karboksil yang cocok mengenali gen avirulen (AVR-Pita), sehingga dapat menggerakkan resisten ras spesifik (Bryan et al. 2000).
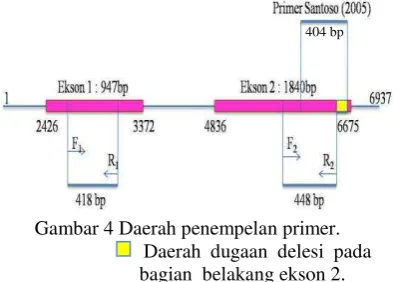
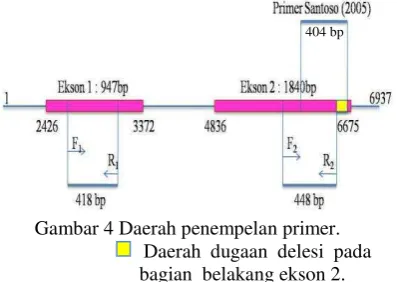
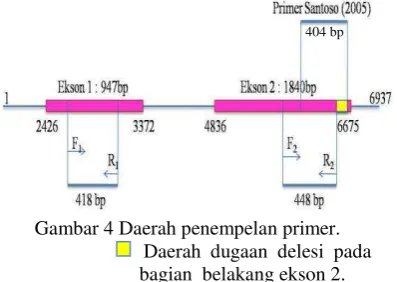
Hasil elektroforesis dari hasil PCR menunjukkan adanya gen Pi-ta yang teramplifikasi baik dari DNA tanaman yang tahan (resistence) maupun tanaman rentan (susceptible) seperti varietas Kencana Bali, galur IPB107-F-18-4-2 dan IPB107-F-16E-6 (Gambar 3). Hasil ini berbeda dengan Santoso (2005), hal ini di duga karena perbedaan primer yang digunakan. Primer yang digunakan pada penelitian ini mengamplifikasi daerah gen Pi-ta pada daerah ekson 1 di urutan basa ke- 2772-3189 (F1 menempel pada urutan basa ke-
2771-2790; R1 menempel pada urutan basa ke-
3169 – 3188) dan daerah ekson 2 di urutan basa ke- 6089-6536 (F2 menempel pada
urutan basa ke- 6089-6108; R2 menempel
pada urutan basa ke- 6517-6536), sedangkan primer Santoso (2005) mengamplifikasi daerah gen Pi-ta di urutan basa ke- 6257-6660. Primer Santoso (2005) memiliki primer forward yang menempel pada urutan basa ke-6257–6276 dan primer reverse yang menempel pada urutan basa ke- 6640 – 6660 dengan hasil amplifikasi sebesar 404 bp (Gambar 4).
Gambar 4 Daerah penempelan primer.
Daerah dugaan delesi pada bagian belakang ekson 2. Sehingga, diduga adanya kemungkinan delesi pada daerah ekson 2 di bagian belakang pada Kencana Bali, hingga primer Santoso tidak dapat mengamplifikasi gen Pi-ta dari tanaman Kencana Bali. Sementara, menurut Bustamam et al. (2004) Kencana Bali diduga mempunyai gen ketahanan terhadap penyakit blas namun isolat blas yang tidak kompatibel (avirulen) untuk varietas ini belum diketahui. Hasil ini menunjukkan kemungkinan gen Pi-ta tidak spesifik berperan dalam mekanisme pertahanan tanaman pada galur-galur padi ini, sehingga diduga ada gen lain yang berperan dalam proses ketahanan tanaman padi terhadap serangan P. oryzae ras 001.
SIMPULAN
Tiga belas galur padi tahan blas didapatkan dari hasil penapisan 48 galur padi harapan yang diuji terhadap ras 001. Kemunculan gen Pi-ta dijumpai baik pada galur padi yang tahan maupun peka terhadap P.oryzae ras 001, sehingga disimpulkan gen Pi-ta tidak bertanggung jawab atas ketahanan terhadap P.oryzae ras 001.
SARAN
Perlu adanya pengujian dengan desain primer yang lain untuk kelanjutan identifikasi gen Pi-ta guna mengetahui kemungkinan adanya delesi yang terdapat pada sekuens atau urutan basa bagian belakang ekson 2 di gen Pi-ta tanaman Kencana Bali.
DAFTAR PUSTAKA
Agrios GN. 1997. Plant Pathology. Academic Press. A Harcourt Science and Technology Company. hlm 286. Amir M, Nasution A, Santoso. 2000.
Inventarisasi ras P. grisea didaerah Sukabumi Jawa Barat musim tanam 1995-1998. Prosiding Kongres Nasional XV. PFI, Purwokerto. hal.148-151. Amir M, Nasution A. 2001. Identification of
Major Genes for Blast Resistance. Current Status and Future Direction. Bogor: Central Research Institute for Food Crops, Agency for Agriculture Research and Development.
[BPS] Badan Pusat Statistik. 2010. Informasi Data Luas Panen, Produksi Tanaman Padi Seluruh Provinsi Jakarta.
Bonman JM. 1992. Durable resistance to rice blast disease-environmental influences. Euphytica 63: 115-123.
Bryan GT, Wu KS, Farrall L, Jia Y, Hershey HP, McAdams SA, Faulk KN, DonaldsonGK, Tarchini R, and Valent B. 2000. A single amino acid differences distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi-ta. The Plant Cell 12: 2033-2045.
Bustamam M, Reflinur, Agisimanto D, Suyono. 2004.Variasi genetik padi tahan blas berdasarkan sidik jari DNA dengan markah gen analog resisten. J Biotek Pertanian 9: 56-61.
Chen D. 1993. Population structure of Pyricularia grisea (Cooke) Sacc. in two screening site quantitive characterization of mayor and minor resistance genes [thesis doctor]. Los Banos: University of Philippines. Dillon VM, Overton J, Grayer RJ, Harborne
JB. 1997. Differences in phytoalexin response among rice cultivars of different resistance to blast. Phytochem 44: 599-603.
Flor HH. 1971. Curent status of the gene-for-gene concept. Annu Rev Phytopathol 9: 275-296.
Kato H. 1976. Some topics in a disease cycle of rice blast and climatic factors. Proceeding of the symposium on climate and rice. Los Banos, Philippines. p.417-425
Kim S, Ahn IP, Park CH, Park SG, Park SY, Jwa NS, and Lee YH. 2000. Molecular characterization of the cDNA encoding
an acidic isoform of PR-1 Protein in Rice. Mol Cells 11: 115-121.
Kim SG, Kim KW, Park EW, Choi D. 2002. Silicon-induced cell wall fortification of rice leaves: A possible celluler mechanism of enhanced host resistance to blast. Phytopathology 92: 1095-1103. Kurnianingsih R. 2008. Ekspresi gen PR1 dan PBZ1 yang terlibat dalam sistem toleransi tanaman padi terhadap penyakit blas (isolat 173) [tesis]. Bogor: Pascasarjana, Institut Pertanian Bogor. Leung H, Shi Z. 1994. Genetic regulation of
sporulation in the rice blast fungus. Di dalam : Zeigler RS, Leong SA, Teng PS, editor. Rice blast disease. Manila Philippines : CAB International-IRRI. hlm 65-68.
Masniawati. 2005. Karakterisasi molekuler dan analisis stabilitas sifat aromatik plasma nutfah padi aromatik Sulawesi Selatan. [laporan penelitian]. Makasar : Pascasarjana, Universitas Hasanudin. Mogi S, Wibowo BS. 1991. Establishment of
the Differential Variety Series for Pathogenic Race Identification of Rice Blast Fungus and the Distribution of Race Based on the New Differential in Indonesia. Jatisari-Indonesia : Rice Disease Study Group Karawang. 30 hlm.
Naqvi Ni, Chatto BB. 1996. Development of sequences characterized amplified region (SCAR) based indirect selection method for a dominant blast resistences in rice. Genome 39:26-30.
Nugraha MFI. 2005. Identifikasi genotipe padi yang memiliki gen-gen ketahanan terhadap penyakit blas [tesis]. Bogor : Pascasarjana, Institut Pertanian Bogor. 54 hlm.
Ou SH. 1985. Rice Disease. Ed ke-2. England Commonwealth Mycological Institut, CAB. Hal. 101.
Osbourn AE. 1996. Preformed antimicrobial compounds and plant defense against fungal attack. Plant Cell 8: 1821-1831. Qu S, Liu G, Zhou B, Bellizzi M, Zeng L,
Dai L, Han B, and Wang GL. 2006. The broad spectrum blast resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice. Genetics 172: 1901-1914.
Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian Pusat Penelitian dan Pengembangan Tanaman Pangan, editor. Prosiding Seminar Hasil Penelitian Rintisan dan Bioteknologi Tanaman; Bogor, 26-27 Desember 2001. Bogor: BPP Press. Hlm 52-62.
Reflinur. 2005. Keragaman genetik cendawan Pyricularia grisea berdasarkan primer spesifik gen virulensi dan interaksinya terhadap gen ketahanan padi [tesis]. Bogor : Pascasarjana, Institut Pertanian Bogor. 42 hlm.
Rodrigues FA, McNally DJ, Datnoff LE, Jones JB. 2004. Silicon enhances the accumulation of diterpenoid phytoalexins in rice: a potential mechanism for blast resitance. Phytopath 94: 177-183.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning a laboratory manual (2nd ed.). USA : Cold Spring Harbor Laboratory Press. 1567 hlm.
Santoso. 2005. Analisis ketahanan 28 genotipe padi terhadap penyakit blas daun dan hubungannya dengan keberadaan gen Pi-b dan Pi-ta1 [Tesis]. Bogor : Pascasarjana, Institut Pertanian Bogor.
Sari L. 2008. Ekspresi gen penyandi pathogenesis related protein (PR1&PBZ1) pada padi indica yang terlibat dalam sistem toleransi terhadap penyakit blas isolat 001 [tesis]. Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor.
Scardaci SC, Webster RK, Greer CA, Hill JE, Williams JF, Mutters RG, Brandon DM, McKenzie KS, and Oster JJ. 1997. Rice blast: a new disease in California. Agronomy Fact Sheet Series. Davis: University of California.
Semangun H. 1991. Penyakit-penyakit Tanaman Pangan di Indonesia. Yogyakarta: Gadjah Mada University Press.
Suharsono U, Fujisawa Y, Kawasaki T, Iwasaki Y,Satoh H, and Shimamoto K.
2002. The heterotrimeric G protein α
subunit acts upstream of the small GTPase Rac in disease resistance of
rice. Proc Natl Acad Sci USA 99(20): 13307-13312.
Suwarno, Lubis E, Soenarjo E. 2001. Breeding of upland rice in Indonesia. Di dalam: Kardin MK, Prasaja I, Syam M, editor. Upland Rice Research in Indonesia. Bogor: Central Research Institute for Food Crops, Agency for Agricultural Research and Development. hlm. 1-6.
Takahashi E. 1997. Uptake mode and physiological functions of silica. Di dalam: Tanane M, Yuzo M, Fumio K, Hikoyuki Y, editor. Science of Rice Plant Physiology Volume 2. Tokyo: Food and Agriculture Policy Research Center. hlm: 420-433.
Utami DW. 2000. Evaluasi sifat ketahanan terhadap tiga ras uji penyakit blas (Pyricularia grisea) pada spesies padi liar Oryza rufipogon dan populasi tanaman BC2F3 turunannya [tesis].
Bogor: Pascasarjana, Institut Pertanian Bogor.
Utami DW, Amir M, Moeljopawiro S. 2000. Analisis RFLP kelompok ras dan haplotipe isolat blas dengan DNA pelacak MGR586. Biotek Pertanian 5: 28-33.
Utami DW, Moeljopawiro S, Aswidinnoor H, Setiawan A, dan Hanarida I. 2005. Gen Pengendali Sifat Ketahanan Penyakit Blas (Pyricularia grisea Sacc.) pada Spesies Padi Liar Oryza rufipogon Griff. dan Padi Budi Daya IR64. AgroBiogen 1(1):1-6.
Wang ZX, Yano M, Yamanouchi U, Iwamoto M, Monna L, Hayasaka H, Katayose Y, and Sasaki T. 1999. The Pib gene for rice blast resistance belongs to the nucleotide binding and leucine-rich repeat class of plant disease resistance genes. Plant J 19:55-64. Zhou B, Qu S, Liu G, Dolan M, Sakai H, Lu
Lampiran 1 Skala penyakit blas berdasarkan sistem evaluasi standar IRRI (1996)
Keterangan : 0-2 = Resisten (R), 3 = Moderat Resisten (MR), 4-6 = Moderat Sensitif (MS), 7-9=Sensitif (S)
Skala Gejala
0 Tidak ada bercak
1 Bercak kecil berukuran sebesar ujung jarum/lebih besar dan berwarna coklat, tidak ada pusat sporulasi
2 Bercak abu-abu berbentuk bundar agak lonjong diameter 1-2 mm,tepi warna coklat, pada daun bawah
3 Sama dengan skala 2, pada daun atas
4 Bercak khas blas ukuran 3 mm/lebih panjang, luas daun terinfeksi < 4%
5 Bercak khas blas, luas daun terinfeksi 4-10%
6 Bercak khas blas, luas daun terinfeksi 11-25%
7 Bercak khas blas, luas daun terinfeksi 26-50%
8 Bercak khas blas, luas daun terinfeksi 51-75%, banyak daun yang mati
Lampiran 2 Data morfologi dan panen galur padi tahan
Tabel 1 Karakterisasi morfologi galur padi harapan yang tahan
No Nama Galur
Tanggal tanam/keluar
bunga UB TT DB PDB LDB BGB BGK Brt 1000 butir PM
rata-rata PM
Σ
anakan Umur panen
1 IPB107-F-5-1 13/11/2010 -24/02/2011 103 hst 101 33 2 * * 23,00 ** 24,5 8 157 hst 19/04/2011
20,5
21,5 22,17
2 IPB107-F-7-3 13/11/2010-24/02/2011 103 hst 106 0,71 34 2,2 25,77 20,11 17,85 28,2 12 157 hst 24/03/2011
26
25,8 26,67
3 IPB113-F-1 13/11/2010-23/02/2011 102 hst 106 0,46 29 2,7 * 15,76 24,06 * * 11 135 hst 28/03/2011
4 IPB113-F-2-2 13/11/2010-17/02/2011 96 hst 137 0,92 54 3 18,25 15,76 28,16 33 9 127 hst 24/03/2011
31,5
30 31,5
5 IPB115-F-4-2-2 13/11/2010-19/02/2011 98 hst 87 0,61 32,5 2 * 6,59 24,86 24 5 135 hst 28/03/2011
24,5
25 24,5
6 IPB140-F-2-1 13/11/2010-01/03/2011 107 hst 117 0,82 41,5 2,4 11,15 7,68 23,07 20,5 2 131 hst 24/03/2011
26
26,5 24,33
7 IPB140-F-5 13/11/2010-18/03/2011 124 hst 112 0,64 31 1,8 * * 18,52 25 3 156 hst 19/04/2011
26,8
25,1 25,63
8 IPB-149-F-2 04/11/2010-13/02/2011 101 hst * 0,61 37 2,6 17,12 15,53 29,24 26 4 142 hst 24/03/2011
25,6
23,7 25,1
9 IPB149-F-4 13/11/2010-01/03/2011 107 hst 121 0,81 38 3,2 19,47 16,44 29,66 31 3 131 hst 24/03/2011
30
27 29,33
10 IPB149-F-5 13/11/2010-3 Maret 2011 110 hst 90 0,98 22,2 2,3 * * 22,4 24,2 2 157 hst 19/04/2011
20,7 22,45
11 IPB149-F-8 4/11/2010-12/02/2011 101 hst 104 0,61 27,7 2,4 37,47 33,72 29,58 25,6 12 141 hst 24/03/2011
26,5
28 26,7
Keterangan : UB = Umur Bunga; TT = Tinggi Tanaman; DB= Diameter Batang; PDB = Panjang Daun Bendera; LDB = Lebar Daun Bendera; BGB = Berat Gabah Basah; BGK = Berat Gabah Kering; PM = Panjang Malai; * = tidak diamati; ** = dari tetua.
Umur Bunga = Tanggal keluar bunga – tanggal tanam
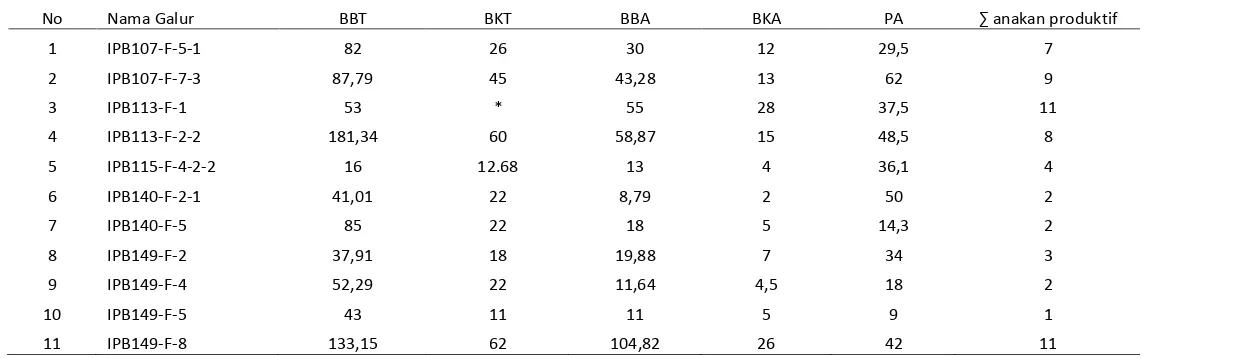
Tabel 2 Pengamatan bobot dan jumlah anakan dari 18 galur yang tahan
No Nama Galur BBT BKT BBA BKA PA ∑ anakan produktif
1 IPB107-F-5-1 82 26 30 12 29,5 7
2 IPB107-F-7-3 87,79 45 43,28 13 62 9
3 IPB113-F-1 53 * 55 28 37,5 11
4 IPB113-F-2-2 181,34 60 58,87 15 48,5 8
5 IPB115-F-4-2-2 16 12.68 13 4 36,1 4
6 IPB140-F-2-1 41,01 22 8,79 2 50 2
7 IPB140-F-5 85 22 18 5 14,3 2
8 IPB149-F-2 37,91 18 19,88 7 34 3
9 IPB149-F-4 52,29 22 11,64 4,5 18 2
10 IPB149-F-5 43 11 11 5 9 1
11 IPB149-F-8 133,15 62 104,82 26 42 11
Jika dilihat dari banyaknya bernas dan panjang malai, maka galur yang mempunyai kualitas bagus yaitu IPB149-F-2, IPB149-F-4, IPB107-F-7-3, IPB140-F-2-1, dan IPB149-F-8.
ABSTRAK
YULITA FITRIANNA. Uji Ketahanan 48 Galur Padi terhadap Penyakit Blast (Pyricularia oryzae) Ras 001. Dibimbing oleh UTUT WIDYASTUTI dan HAJRIAL ASWIDINNOOR.
Penyakit blas yang disebabkan oleh cendawan Pyricularia oryzae Cav. merupakan salah satu penyakit utama yang menyerang padi di Indonesia. Penelitian ini bertujuan untuk menguji tingkat ketahanan galur-galur padi terhadap serangan penyakit blas dan mengidentifikasi keberadaan gen ketahanan Pi-ta pada galur padi yang tahan. Penelitian dilakukan dengan menggunakan padi varietas Kencana bali (peka) dan Cisadane (tahan) sebagai kontrol serta 48 galur padi hasil seleksi Program Pemuliaan di Departemen Agronomi dan Hortikultura IPB dengan metode injeksi menggunakan P. oryzae ras 001. Padi berumur 16 hari setelah tanam diinokulasikan dengan 1 ml suspensi spora (104-105 spora/ml) isolat P. Oryzae. Tanaman diinkubasi pada kondisi gelap lalu dipindah ke ruang terang dan dilakukan pengamatan hingga 7-9 hari setelah infeksi. Pengujian tahan blas dilakukan 2 tahap, yaitu penapisan I dilakukan sebanyak 3 ulangan dan penapisan II dilakukan sebanyak 2 ulangan untuk masing-masing galur terpilih dari penapisan I, dengan 10 tanaman pada tiap ulangan. Pengamatan hingga penapisan II menunjukkan adanya 13 galur padi tahan terhadap P. oryzae ras 001 yaitu IPB140-F-2-1, IPB115-F-4-2-2, IPB113-F-2-2, IPB107-F-7-3, IPB140-F-5, IPB149-F-2, IPB149-F-8, Martapura, IPB149-F-4, IPB149-F-5, IPB113-F-1, IPB102-F-92-1-1, IPB107-F-5-1 dengan persentase ketahanan ≥ 80 %. Dua pasang primer spesifik untuk gen Pi-ta didesain dari sekuen DNA dengan nomor aksesi AF207842 untuk mengamplifikasi fragmen DNA gen Pi-ta dengan metode PCR. Hasil identifikasi gen ketahanan Pi-ta menunjukkan bahwa 13 galur tahan maupun 2 galur rentan (18-4-2 dan IPB107-F-16E-6) mempunyai gen Pi-ta, bahkan pada Kencana Bali. Percobaan sebelumnya, menunjukkan bahwa Kencana Bali tidak memiliki gen ketahanan Pi-ta. Diduga bahwa varietas Kencana Bali mengalami delesi di daerah bagian belakang ekson 2. Hasil identifikasi gen Pi-ta juga mengindikasikan gen Pi-ta tidak spesifik berperan dalam mekanisme pertahanan tanaman pada galur-galur padi ini, sehingga diduga ada gen lain yang berperan dalam proses ketahanan tanaman padi terhadap serangan P. oryzae ras 001.
Kata kunci: Padi (Oryza sativa), blas, Pyricularia oryzae, resisten
ABSTRACT
YULITA FITRIANNA. Resistance Test toward of 48 Lines Rice to Blast Disease (Pyricularia oryzae) Race 001. Supervised by UTUT WIDYASTUTI and HAJRIAL ASWIDINNOOR.
Blast disease, caused by Pyricularia oryzae Cav., is one of the major fungal diseases infected rice (Oryza sativa L.) in Indonesia. The aim of this research was to examine the resistance of rice from the infection of P. Oryzae race 001 and to detect the existance of Pi-ta gene in resistant lines of rice. Resistance evaluation to blast disease (Pyricularia oryzae) on Cisadane (resistant), Kencana Bali (susceptible) as control, and 48 lines of rice was carried out in the green house by using injection inoculated method with P. oryzae race 001. Rice seedling were planted on sterilize soil, and sixteen days old of rice were soaked in the fungal spore suspension (105 spores/ ml) in the dark for 24 hours, and moved to the light. Rice disease was observered on 7-9th days after infection. There were 2 step screening of blast-resistant, which screening I performed a total of three replicates and screening II performed from selected plant in screening I a total of two replicates for each line of rice, with 10 plants in each replication. The result of second screening at green house showed that 13 lines resistant to P. oryzae 001 were IPB140-F-2-1, IPB115-F-4-2-2, IPB113-F-2-2, IPB107-F-7-3, IPB140-F-5, IPB149-F-2, IPB149-F-8, Martapura, IPB149-F-4, IPB149-F-5, IPB113-F-1, IPB102-F-92-1-1, IPB107-F-5-1 with the percentage of resistance ≥ 80 %. Two pairs of DNA primers specific to the dominant indica Pi-ta gene were designed from DNA sequence in gene bank with accession number AF207842 to amplify the Pi-ta DNA fragments by polymerase chain reaction (PCR) method. The results revealed that all of 13 resistant and 2 susceptible lines (IPB107-F-18-4-2 and IPB107-F-16E-6) of rice carrying Pi-ta genes, even in Kencana Bali. The previous experiment shown that Kencana Bali does not have Pi-ta resistance gene. It is expected that Kencana Bali has deletion of Pi-ta resistance gene in the end part of exon 2. Further more, those results also indicated that Pi-ta gene did not confer resistance to race 001 of Pyricularia oryzae. Resistance to P. oryzae race 001 might be controlled by different resistant gene.
PENDAHULUAN
Latar Belakang
Tanaman padi (Oryza sativa L.) di Indonesia banyak terserang oleh bermacam-macam penyakit (Semangun 1991). Salah satu penyakit yang sangat merugikan petani adalah penyakit blas. Penyakit blas yang disebabkan oleh cendawan Pyricularia oryzae Cav. masih merupakan masalah utama pada padi gogo (Rais et al. 2001). Penyakit blas sebelumnya diketahui hanya menyerang padi gogo (Semangun 1991), tetapi saat ini penyebaran penyakit blas sudah mulai menyerang pertanaman padi sawah (Amir et al. 2000). Serangan penyakit blas di Indonesia diperkirakan mencapai 19.629 hektar dari total luas lahan pertanaman padi Indonesia sebesar 12.833.578 hektar pada tahun 2009 (BPS 2010). Kehilangan hasil pada varietas yang peka seperti Bical dapat mencapai 50-90% (Rais et al. 2001).
Secara umum P. oryzae menyerang tanaman padi pada bagian daun, buku, leher, malai, cabang malai, bulir padi, kulit gabah (Chen 1993; Rais et al. 2001), dan collar daun (Scardaci et al. 1997). Gejala serangan penyakit blas mempunyai bentuk khas, gejala daun yang terserang oleh blas daun (leaf blast) berupa bercak berbentuk belah ketupat sampai elips dengan kedua ujungnya runcing. Bagian tengah bercak berwarna abu-abu atau putih keabu-abuan, pada bagian tepi berwarna coklat sampai coklat kemerahan dengan ukuran panjang bercak 1-1.5 cm dan lebar 0.3-0.35 cm (Ou 1985; Scardaci et al. 1997).
Bentuk, warna, dan ukuran bercak dapat beragam bergantung pada ketahanan varietas, umur tanaman, dan umur lesio. Bercak tidak akan berkembang dan tetap seperti titik kecil pada varietas yang tahan dan bercak akan berkembang sampai beberapa milimeter berbentuk bulat atau elips dengan tepi berwarna coklat pada varietas dengan reaksi sedang (Ou 1985). Blas daun kadang-kadang menyebabkan kematian keseluruhan pada tanaman yang masih muda sampai stadia pembentukan anakan atau tillering (Scardaci et al. 1997).
Menurut Amir et al. (2000) ras 001, 003, 033, dan 173 merupakan ras yang ada pada setiap musim tanam, artinya isolat ini memiliki penyebaran yang luas. Isolat ras 001 mempunyai tingkat patogenesitas yang paling rendah virulensinya, menyebar luas dan mampu bertahan lama di lapangan (Sari 2008; Utami 2000). Adapun penentuan ras baru dari patogen blas di Indonesia dilakukan dengan
menggunakan varietas diferensial yaitu Asahan, Cisokan, IR64, Krueng Aceh, Cisadane, Cisanggang, dan Kencana Bali (Mogi et al. 1991).
Studi genetika untuk mendapatkan gen resisten terhadap penyakit blas telah banyak dilakukan. Pada tanaman padi telah diidentifikasi sekitar 40 gen mayor yang mengendalikan ketahanan terhadap penyakit blas dan 5 gen resisten, Pi-b, Pi-ta, Piz-5(Pi-2), Piz-t, dan Pi9 telah berhasil diisolasi (Wang et al. 1999; Qu et al. 2006; Zhou et al. 2006). Menurut Ou (1985), gen-gen ketahanan tersebut (Pi) akan berinteraksi secara spesifik dengan ras-ras tertentu dari cendawan Pyricularia grisea.
Menurut Suwarno et al. (2001) sumber gen Pi banyak terdapat pada gene pool padi Indonesia baik yang berasal dari varietas budidaya maupun spesies padi liar. Dua puluh delapan genotip padi indica yang terdapat di Indonesia telah dianalisis oleh Santoso (2005) dan ditemukan bahwa banyak padi indica yang mempunyai gen ketahanan baik Pi-b maupun Pi-ta. Gen Pi-b dan Pi-ta merupakan 2 gen mayor resisten blas hasil introgresi dari kultivar padi indica. Kedua gen resisten tersebut mempunyai struktur gen yang mirip, yaitu NBS (nucleotide binding site) dan LRR (leucine rich repeat) dan di duga menyandikan protein sitoplasmik yang bertanggung jawab merespon gen avirulen AVR-Pita, memicu resisten ras spesifik (Wang et al. 1999; Bryan et al. 2000; Sari 2008).
Tujuan Penelitian
Penelitian ini bertujuan untuk menguji tingkat ketahanan galur-galur padi terhadap serangan penyakit Blas (Pyricularia oryzae) ras 001 dan mengidentifikasi keberadaan gen ketahanan Pi-ta pada galur padi yang tahan.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Februari 2010 – Maret 2011, bertempat di laboratorium BIORIN dan Rumah Kaca Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB), Institut Pertanian Bogor.
Bahan dan Alat
Bahan tanaman yang digunakan yaitu 48 galur padi hasil seleksi Program Pemuliaan di Departemen Agronomi dan Hortikultura IPB, varietas peka (Kencana Bali) dan varietas tahan (Cisadane) sebagai kontrol, dan isolat cendawan P. oryzae ras 001. Bahan kimia yang digunakan yaitu media oatmeal, Potato Dextrose Agar (setiap liter mengandung 39 g PDA dan supaya lebih padat ditambah 5 g agar), antibiotik kloramfenikol, aquades, larutan Tween 0.025% (v/v), nitrogen cair, buffer ekstrak 2% (w/v) (dengan komposisi 2% CTAB (w/v), 0.1 M Tris HCl pH 8.0, 1.5
M NaCl, 0.02 M EDTA), 2% β-mercapto etanol, kloroform, isoamil alkohol, phenol, Na acetat 3.2 M, alkohol absolut, RNAse 10 µg/µl, aquabides, gel agarosa konsentrasi 1% (b/v), larutan TAE 1X, loading dye, dan ethidium bromida 0.5 mg/L.
Alat yang digunakan yaitu autoklaf, laminar, mikroskop, hemasitometer, magnetik stirrer, mikropipet berbagai ukuran, sentrifuse Jouan BR-4i, shaker, cawan petri, jarum ose, mesin PCR, dan alat elektroforesis.
Metode penelitian
Uji Ketahanan Tanaman Padi
Penyiapan Tanaman Padi. Benih padi dicuci kemudian dikecambahkan dalam cawan petri yang berisi kertas merang selama kurang lebih empat hari lalu ditanam ke media tanah. Masing-masing galur ditanam sebanyak 10 benih dan dibuat sebanyak 3x ulangan.
Produksi Spora. Potongan miselium isolat cendawan P. oryzae ras 001 asal padi diinokulasikan pada media oatmeal (30 g oat/l, 20 g agar/l, 5 g sukrosa/l), dan diinkubasi selama 10-14 hari pada suhu ruang, lalu kultur dimaserasi atau dicuci secara aseptik untuk menghilangkan hifa aerial. Kultur disinari n-UV (300-400 nm) terus menerus selama
kurang lebih enam hari untuk menginduksi pembentukan spora. Sebanyak 1-3 ml air steril yang mengandung 0.025% (v/v) Tween 20 disiramkan pada permukaan koloni cendawan. Permukaan koloni cendawan diusap-usap dengan bantuan kaca objek steril untuk melepaskan spora dari konidiofor. Spora-spora dari isolat yang sama dipanen dan dikumpulkan kemudian dilakukan pengukuran kerapatan spora dengan hemasitometer hingga diperoleh suspensi spora 104-105 spora/ml (Chao & Ellingboe 1997).
Uji Ketahanan terhadap Infeksi. Benih padi yang telah berkecambah ditanam pada bak berukuran 25x35x15 cm yang diberi tray berisi campuran tanah dan pupuk dengan perbandingan 10 kg tanah, 2.5 gr urea, 1.5 gr TSP, dan 1.2 gr KCl. Tanah yang digunakan adalah tanah yang telah disterilkan. Masing-masing galur ditanam sebanyak 10 benih dan dibuat sebanyak 3x ulangan. Selanjutnya, padi berumur 16 hari setelah tanam (memiliki ±5 daun) diinokulasikan dengan 1 ml suspensi spora (104-105 spora/ml) isolat P. Oryzae melalui penyuntikan di bagian pelepah daun kelima. Sebagai kontrol negatif, padi disuntik menggunakan akuades (tidak diinfeksi). Selanjutnya semua tanaman diinkubasi pada kondisi gelap dan lembab selama 24 jam, kemudian dipindahkan ke ruang terang selama enam hari pada suhu 25-30˚C dan kelembaban 70-90 %. Pengamatan kemampuan infeksi terhadap inang berupa kemunculan bercak pada daun dilakukan setiap hari selama kurang lebih 7-9 hari setelah infeksi. Tingkat ketahanan diukur dengan menggunakan tabel skala penyakit yang ditentukan oleh IRRI (Lampiran 1). Uji ketahanan terhadap infeksi dilakukan sebanyak 2 tahap penapisan (Screening). Penapisan I dilakukan sebanyak 3 ulangan untuk masing-masing galur dengan 10 tanaman pada setiap ulangan. Setelah didapatkan galur padi dengan nilai persentase ketahanan terhadap blas ≥ 80% dilakukan tahap penapisan II yang dilakukan sebanyak 2 ulangan untuk masing-masing galur dengan 10 tanaman pada setiap ulangan. Persentase akhir ketahanan merupakan rata-rata persentase ketahanan dari kedua ulangan. Persentase
ketahanan ≥ 80% digolongkan menjadi tanaman resisten (tahan).
Identifikasi Keberadaan Gen Ketahanan Pi-ta pada Galur Padi Tahan Blas
– 0.2 g lalu ditambahkan 600 µL buffer ekstrak (dengan komposisi 2% Cetyl Trimethyl Ammonium Bromide [CTAB] (w/v), 0.1 M Tris HCl pH 8.0, 1.5 M NaCl, 0.02 M Ethylene Diamine Tetra acetic Acid [EDTA] ), dan mercapto etanol 0,2%. Campuran dikocok, diinkubasi selama 30 menit pada suhu 650C. Campuran dalam tabung eppendorf didinginkan di es selama 5 menit, kemudian ditambahkan 600 µL kloroform : isoamil alkohol (24:1). Tabung eppendorf kemudian disentrifuse pada 10000 rpm (g = ± 9.8 m/s2) dengan suhu 40C, selama 10 menit. Supernatan bagian atas (mengandung DNA) diambil dan dipindahkan pada tabung ependorf lain kemudian ditambahkan PCI (Phenol, Chloroform, dan Isoamil alkohol) dingin dengan perbandingan (25:24:1) sebanyak 500 µL, disentrifuse kembali pada 10000 rpm (g = ± 9.8 m/s2) dengan suhu 40C selama 5 menit. Supernatan diambil kemudian ditambahkan 2M NaOAC pH 5,2 (0.1x volume supernatan) dan EtOH 100% sebanyak 2x volume supernatan. Presipitasi dilakukan selama 30 menit dalam freezer. Pelet diambil dan dibilas dengan 500 µL etanol 70% lalu disentrifuse 5 menit dengan kecepatan 10000 rpm (g = ± 9.8 m/s2). Cairan dibuang dan pelet dikeringkan dengan vakum. Pelet dilarutkan ke dalam 20-50 µL aquades steril, diberi RNAse (10 µg/µl) sebanyak 0.2x volume aquades steril selama 30 menit pada suhu 370C. Tahap terakhir yaitu inaktif RNAse pada suhu 700C, selama 10 menit.
Uji Kualitas dan Kuantitas DNA. Uji kualitas dan kuantitas DNA dilakukan menurut metode Sambrook et al. (1989) dengan beberapa modifikasi. DNA padi yang berhasil diisolasi dielektroforesis pada gel agarosa dengan konsentrasi 1% (b/v) dalam buffer penyangga 1x TAE. Perbandingan DNA dengan loading dye adalah 5:2 µL. Elektroforesis dilakukan selama kurang lebih 30 menit pada tegangan 100 volt. Selanjutnya gel agarose hasil elektroforesis dimasukkan ke dalam larutan ethidium bromida (konsentrasi larutan 0.5 mg/L) selama 5 menit. Panjang pita DNA diamati dengan melihat pita DNA pada UV transiluminator.
Identifikasi Gen Pi-ta. Amplifikasi DNA guna mengidentifikasi gen ketahanan Pi-ta dilakukan dengan bantuan Polymerase Chain Reaction (PCR). Primer yang digunakan adalah Pi-ta ekson 1 dan Pi-ta ekson 2. Primer spesifik ini didesain dari sekuen DNA pada database dengan nomor kode aksesi AF207842. Primer spesifik Pi-ta ekson 1 digunakan untuk amplifikasi gen Pi-ta di
ekson 1 dengan ukuran produk PCR sebesar 418 bp. Sedangkan, primer spesifik Pi-ta ekson 2 digunakan untuk amplifikasi gen Pi-ta di ekson 2 dengan ukuran produk PCR sebesar 448 bp.
Tabel 1 Primer spesifik gen Pi-ta Primer spesifik Pi-ta ekson 1
forward 5’ACTGCTGGTGCCAAGAAGAT3’
reverse 5’GGCCATGCAGACGATAGAAT3’
Primer spesifik Pi-ta ekson 2
forward 5’CCCAGGATGACCTTGACACT3’
reverse 5’TGTGCCAAATCTTCATCCAA3’
Reaksi PCR mengikuti metode yang dilakukan Naqvi dan Chattoo (1996). PCR dilakukan dengan total reaksi 10µl mengandung 100 ng DNA genomik cetakan (1 µl), dNTPmix (dATP, dCTP, dGTP, dTTP) 0.2 mM (1 µl), masing-masing primer (forward dan reverse) 1 pmol (masing-masing sebanyak 0.4 µl), enzim taq DNA polymerase 1 unit (0.2 µl) dalam larutan buffer 1X (1 µl) dan ditambahkan air bebas ion hingga mencapai volume 10 µl. Amplifikasi PCR dilakukan sebanyak 30 siklus. Progra