PENGARUH CENDAWAN ENDOFIT TERHADAP BIOLOGI
DAN PERTUMBUHAN POPULASI
Polyphagotarsonemus latus
Banks (ACARI: TARSONEMIDAE) PADA TANAMAN CABAI
ELIN TASLIAH
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Pengaruh Cendawan Endofit terhadap Biologi dan Pertumbuhan Populasi Polyphagotarsonemus latus Banks (Acari: Tarsonemidae) pada Tanaman Cabai adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau kutipan dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2014
Elin Tasliah A34100021
ABSTRAK
ELIN TASLIAH. Pengaruh Cendawan Endofit terhadap Biologi dan Pertumbuhan Populasi Polyphagotarsonemus latus Banks(Acari: Tarsonemidae) pada Tanaman Cabai. Dibimbing oleh SUGENG SANTOSO dan WIDODO.
Tungau Polyphagotarsonemus latus merupakan salah satu hama penting pada tanaman cabai. Populasi P. latus yang tinggi dapat menyebabkan kematian tanaman cabai. Salah satu alternatif pengendalian yang ramah lingkungan adalah dengan memanfaatkan cendawan endofit. Penelitian ini bertujuan menguji pengaruh cendawan endofit terhadap biologi dan pertumbuhan populasi P. latus pada tanaman cabai. Benih cabai direndam pada suspensi miselium selama 12 jam kemudian ditanam. Pada 3 minggu setelah sebar benih, tanaman cabai kemudian disiram dengan suspensi miselium. Perbanyakan tungau kuning dilakukan pada tanaman cabai di laboratorium. Lima imago betina berumur seragam dari hasil perbanyakan diinfestasikan ke setiap tanaman pada satu minggu setelah inokulasi cendawan endofit terakhir. Pengamatan pertumbuhan populasi P. latus dilakukan setiap minggu selama 10 minggu. Di laboratorium, 50 telur tungau kuning berumur seragam dipelihara secara satu per satu pada daun cabai dan diamati setiap 6 jam sampai menjadi imago. Imago yang terbentuk kemudian dipasang-pasangkankan dan diamati setiap hari sampai imago mati. Siklus hidup, keperidian, nisbah kelamin, dan lama hidup imago diamati. Hasil penelitian ini menunjukkan bahwa perlakuan cendawan endofit dapat meningkatkan viabilitas benih. Perlakuan beberapa jenis cendawan endofit dapat memperpanjang masa praoviposisi, menekan pertumbuhan populasi, menurunkan keperidian dan meningkatkan jumlah individu jantan.
ABSTRACT
ELIN TASLIAH. The Effects of Endophytic Fungi on Biology and Population Growth of Broad Mite, Polyphagotarsonemus latus Banks (Acari: Tarsonemidae), on Pepper. Supervised by SUGENG SANTOSO and WIDODO.
Broad mite Polyphagotarsonemus latus is one of the most important pests of pepper. High population of P. latus on pepper may cause death on plants. One of the alternative environmental-friendly control methods is the use of endophytic fungi. This study aimed to know effect of the endophytic fungi on biology and population growth of broad mite on pepper. The pepper seeds were soaked in miselium suspension for 12 hours and then planted. Three weeks after seeds planting, peppers were sprayed with miselium suspension. Broad mite collected from the field were reared on pepper in the laboratory. The same age-five female adults were infested on each pepper seedling a week after last inoculation of endophytic fungi. The population growth of broad mite were observed each week for 10 weeks. In the laboratory, 50 broad mite eggs were reared on pepper leaves and observed every 6 hours until adult emergence. P. latus then paired and observed everyday until all adults dead. Life cycle, fecundity, sex ratio and adult longevity were observed. The result showed that treatment of endhophytic fungi could increase pepper seeds viability. Several kinds of endophytic fungi also suppressed population growth, elongated praoviposition period, reduced mite fecundity and increase the number of male.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
PENGARUH CENDAWAN ENDOFIT TERHADAP BIOLOGI
DAN PERTUMBUHAN POPULASI
Polyphagotarsonemus latus
Banks
(ACARI: TARSONEMIDAE) PADA TANAMAN CABAI
ELIN TASLIAH
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Skripsi : Pengaruh Cendawan Endofit terhadap Biologi dan Pertumbuhan Populasi Polyphagotarsonemus latus Banks(Acari:
Tarsonemidae) pada Tanaman Cabai Nama : Elin Tasliah
NIM : A34100021
Disetujui oleh
Dr. Ir. Sugeng Santoso, MAgr. Dr. Ir. Widodo, MS.
Pembimbing I Pembimbing II
Diketahui oleh
Dr. Ir. Abdjad Asih Nawangsih, MSi. Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT, yang atas segala karunia-Nya sehingga penulis dapat menyelesaikan laporan tugas akhir yang berjudul “Pengaruh Cendawan Endofit terhadap Biologi dan Pertumbuhan Populasi Polyphagotarsonemus latus Banks(Acari: Tarsonemidae) pada Tanaman Cabai”. Penelitian dilaksanakan dari Oktober 2013 sampai Maret 2014. Penelitian ini didanai oleh Dirjen DIKTI melalui beasiswa Bidik Misi.
Ucapan terima kasih penulis sampaikan kepada
1. Dr. Ir. Sugeng Santoso, MAgr. dan Dr. Ir. Widodo, MS. selaku dosen pembimbing yang senantiasa memberikan bimbingan, masukan, pengetahuan, saran dan arahan kepada penulis.
2. Prof. Dr. Ir. Meity S. Sinaga, MSc. selaku dosen penguji tamu yang telah memberikan masukan kepada penulis.
3. Dr. Ir. Tri Asmira Damayanti, MAgr. selaku dosen pembimbing akademik yang telah memberikan saran dan dukungan selama penulis mengikuti perkuliahan.
4. Dirjen DIKTI yang telah memberikan dukungan dana selama perkuliahan melalui Beasiswa Bidik Misi.
5. Bapak, ibu, kakak, adik, serta seluruh keluarga, atas doa dan kasih sayang. 6. Teman-teman Departemen Proteksi Tanaman angkatan 47 yang telah
mendukung terlaksananya laporan tugas akhir penulis. Serta pihak lain yang turut membantu dalam penyusunan laporan tugas akhir ini.
Semoga laporan tugas akhir ini dapat bermanfaat bagi penulis dan pembaca.
Bogor, Juli 2014
DAFTAR ISI
Tempat dan Waktu Penelitian 3
Bahan 3
Persiapan 3
Pemeliharaan P. latus 3
Penyediaan Isolat Cendawan Endofit 3
Pengaruh Cendawan Endofit terhadap Pertumbuhan Populasi P. latus di
Rumah Kaca 4
Perlakuan Benih Cabai dengan Cendawan Endofit 4
Penanaman Cabai ke dalam Polibag 4
Infestasi P. latus 4
Pengamatan 4
Pertumbuhan Populasi P. latus 4
Biologi P. latus 4
Analisis Data 5
HASIL DAN PEMBAHASAN 6
Pertumbuhan Tanaman Cabai yang Diinokulasi Cendawan Endofit 6
Pertumbuhan Populasi P. latus 7
Siklus Hidup P. latus 10
Biologi P. latus 13
SIMPULAN DAN SARAN 16
1 Pengaruh cendawan endofit terhadap lama perkembangan pradewasa
Polyphagotarsonemus latus 12
2 Pengaruh cendawan endofit terhadap sifat biologi imago
Polyphagotarsonemus latus 14
DAFTAR GAMBAR
1 Penempelan daun bergejala serangan P. latus 3
2 Perkecambahan benih cabai yang diinokulasi cendawan endofit 7 3 Pengaruh cendawan endofit terhadap perkembangan populasi
Polyphagotarsonemus latus 9 4 Tunas baru yang muncul pada tanaman cabai setelah serangan P. latus 9 5 Pengaruh cendawan endofit terhadap pertumbuhan tunas baru tanaman cabai
kontrol (a) dan diinokulasi cendawan endofit II7-1 (b) pada 8 MSI 10 6 Fase perkembangan Polyphagotarsonemus latus (a) telur (b) larva
(c) nimfa (d) imago betina (e) imago jantan (f) kopulasi 11 7 Pengaruh cendawan endofit terhadap keperidian P. latus 15
DAFTAR LAMPIRAN
1 Kompensasi kerusakan Polyphagotarsonemus latus pada tanaman cabai
yang diinokulasi cendawan endofit 20
2 Pengaruh cendawan endofit terhadap perkembangan populasi
Latar Belakang
Tanaman cabai merah (Capsicum annum L.) merupakan salah satu komoditas hortikultura andalan di Indonesia. Menurut BPS (2013), tahun 2012 produktivitas cabai besar segar dengan tangkai sebanyak 7.93 ton/ha meningkat dibandingkan dengan tahun 2011 sebanyak 7.34 ton/ha. Sampai saat ini budidaya cabai masih menghadapai berbagai permasalahan sehingga produktivitas cabai besar masih dibawah potensi hasil 12 ton/ha (Purwati et al. 2000). Rendahnya produksi cabai antara lain disebabkan oleh serangan hama dan patogen tanaman.
Polyphagotarsonemus latus Banks yang dikenal tungau teh kuning merupakan salah satu hama penting pada tanaman cabai. P. latus dapat menyerang pada semua fase pertumbuhan tanaman cabai (Setiawati et al. 2005) . Gejala serangan tungau kuning muncul 8-10 hari setelah terserang. Pucuk tanaman seperti terbakar pada 4-5 hari setelah gejala muncul. Serangan berat mengakibatkan pucuk tanaman mati, buah cabai kaku, permukaan buah kasar dan malformasi. Siklus hidup P. latus yang pendek dengan keperidian yang tinggi mengakibatkan populasi P. latus bisa meningkat dengan cepat sehingga mempercepat perkembangan tingkat kerusakan yang ditimbulkannya. Untuk mengurangi kerugian yang ditimbulkan oleh tungau kuning tersebut, perlu dilakukan beberapa pengendalian.
Pengendalian tungau secara terpadu pada pertanaman dapat dilakukan dengan menggunakan berbagai metode, diantaranya pengendalian secara kimia, kultur teknis, tanaman resisten, regulasi, pengamatan hama dan musuh alami dan pengendalian secara biologi (Hoy 2011).
Pengendalian yang selama ini banyak dilakukan oleh petani adalah dengan insektisida atau akarisida. Pengendalian dengan penggunaan pestisida sintetik belum optimal dalam menurunkan kerusakan akibat P. latus. Menurut Zhang (2003), pengendalian menggunakan pestisida non sistemik akan sulit dilakukan karena tungau kuning memiliki kebiasaan berlindung diantara lipatan daun yang mengeriting sehingga tungau kuning sulit terpapar oleh cairan semprot pestisida.
Pengendalian hayati merupakan salah satu alternatif pengendalian yang ramah lingkungan dan dapat menjaga keseimbangan ekosistem di alam. Pengendalian secara biologi dapat dilakukan dengan menggunakan musuh alami seperti predator, parasitoid dan agens antagonis (Hoy 2011). Cendawan endofit merupakan salah satu agens antagonis yang berpotensi untuk mengendalikan tungau. Mekanisme penghambatan cendawan endofit dapat terjadi secara langsung dengan mekanisme antagonis dan secara tidak langsung dengan mekanisme ketahanan terinduksi. Perlindungan tanaman dengan ketahanan terinduksi didasarkan pada rangsangan mekanisme ketahanan oleh adanya perubahan metabolik yang memungkinkan tanaman untuk lebih mengefektifkan ketahanannya (Agrios 1997).
tanaman inangnya dari serangan hama dan penyakit, dan hasil dari fotosintesis dapat digunakan oleh cendawan untuk mempertahankan kelangsungan hidupnya (Gao et al. 2010).
Cendawan endofit menginfeksi tumbuhan pada jaringan tertentu dan mampu menghasilkan mikotoksin, enzim serta antibiotik sehingga asosiasi cendawan endofit dengan tumbuhan inang mampu melindungi tumbuhan inang dari beberapa patogen virulen, kondisi ekstrim maupun herbivora (Clay 1988 dalam Istikorini 2008). Mekanisme cendawan endofit dalam melindungi tanaman terhadap serangga ataupun patogen meliputi (1) penghambatan pertumbuhan patogen secara langsung; (2) penghambatan tidak langsung melalui induksi ketahanan tanaman dalam pembentukan metabolit sekunder; (3) perangsangan pertumbuhan tanaman; kolonisasi jaringan tanaman sehingga patogen sulit penetrasi; dan (4) Hiperparasit (Gao et al. 2010).
Penggunaan cendawan endofit dalam meningkatkan pertumbuhan dan mengendalikan serangga sudah banyak diteliti. Cendawan endofit Beauveria bassiana mempengaruhi perilaku makan dari Ostrinia nubilalis (Bing dan Lewis 1991). Cendawan endofit Nigrospora sp. pada tanaman cabai dapat menekan pertumbuhan populasi, menurunkan kepiridian dan memperpanjang siklus hidup Aphis gossypii (Hermawati 2007).
Penelitian tentang potensi cendawan endofit pada cabai sebagai agens pengendali hayati tungau belum dilakukan di Indonesia, sehingga sangat penting untuk dilakukan.
Tujuan Penelitian
Penelitian ini bertujuan menguji pengaruh cendawan endofit pada tanaman cabai terhadap biologi dan pertumbuhan populasi P. latus.
Manfaat Penelitian
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di Rumah Kaca Cikabayan, Laboratorium Bionomi dan Ekologi Serangga, Laboratorium Taksonomi Serangga dan lahan pertanaman di belakang Laboratorium Pendidikan I, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor (IPB), dari Oktober 2013 sampai Maret 2014.
Bahan
Cendawan endofit yang digunakan merupakan hasil isolasi dari tanaman cabai dan sudah dibiakan di Laboratorium Klinik Tanaman, Departemen Proteksi Tanaman, Institut Pertanian Bogor. Tungau yang digunakan berasal dari pertanaman cabai di Bogor. Benih cabai yang digunakan benih cabai besar merah varietas Landung. Pupuk yang digunakan pupuk urea, SP-36 dan KCl.
Persiapan
Pemeliharaan P. latus
P. latus dari lapangan diperbanyak pada tanaman cabai. Daun yang menunjukkan gejala serangan P. latus ditempelkan pada permukaan bawah daun tanaman cabai. Setelah jumlah P. latus memadai, kemudian P. latus dipelihara di laboratorium untuk mendapatkan tungau yang berumur seragam.
Gambar 1 Penempelan daun bergejala serangan P. latus
Penyediaan Isolat Cendawan Endofit
Rumah Kaca
Perlakuan Benih Cabai dengan Cendawan Endofit
Benih cabai disterilisasi permukaan dengan NaOCl 1% selama 2 menit kemudian dibilas dengan akuades steril. Benih kemudian direndam dalam suspensi cendawan endofit selama 12 jam, dan disemai dalam polibag. Inokulasi kedua dilakukan dengan cara penyiraman suspensi miselium 1 ml di sekitar perakaran cabai pada 3 minggu setelah sebar (MSS) benih.
Penanaman Cabai ke dalam Polibag
Benih cabai ditanam pada polibag 30 cm x 30 cm dengan menggunakan media tanam berupa tanah dan kompos dengan perbandingan 1:1 (v/v). Masing-masing polibag diletakkan diatas piring plastik. Benih cabai sebanyak 3 buah ditanam sedalam 0.5 cm. Pupuk terdiri dari urea (5gr/tanaman) dan SP-36 (20 gr/tanaman) diberikan pada saat sebar benih. Pada 6 MSS, diberikan pupuk urea (5 gr/tanaman) dan KCl (5 gr/tanaman). Perkecambahan benih diamati pada 3 MSS. Pada setiap polibag disisakan satu tanaman cabai sebagai objek penelitian.
Infestasi P. latus
Infestasi tungau kuning dilakukan pada 7 hari setelah inokulasi cendawan endofit yang kedua. Setiap tanaman cabai diinfestasi dengan 5 imago betina P. latus yang berumur seragam. P. latus diinfestasikan pada daun ke-3 dari pucuk tanaman.
Pengamatan
Pertumbuhan Populasi P. latus
Pengamatan dilakukan seminggu sekali selama 10 minggu. Pengamatan dimulai 2 minggu setelah infestasi P. latus . Pada setiap tanaman dipetik satu daun ke-3 dari pucuk secara acak. Pengamatan dilakukan pada daun tersebut karena menurut Fasulo (2013), P. latus dapat ditemukan pada daun muda, sedangkan menurut Kalshoven (1981), pada tanaman teh, P. latus banyak ditemukan di pucuk dan diantara rambut pada permukaan bawah daun. Populasi P. latus pada setiap daun diamati menggunakan mikroskop stereo. Setelah diamati, P. latus direinfestasi ke tanaman perlakuan.
Biologi P. latus
P. latus yang berasal dari tanaman cabai perlakuan digunakan sebagai objek penelitian. P. latus dari masing-masing perlakuan kemudian dibawa ke laboratorium.
masing-masing perlakuan menggunakan 25 individu P. latus.
Analisis Data
Rancangan percobaan yang dilakukan dalam pengujian adalah rancangan acak lengkap (RAL) dengan 1 faktor, 6 taraf (kontrol, cendawan endofit I2-2’, II7-1,
II9-1, IV2, dan IV5-2) dan 10 ulangan. Pengolahan data pertumbuhan populasi,
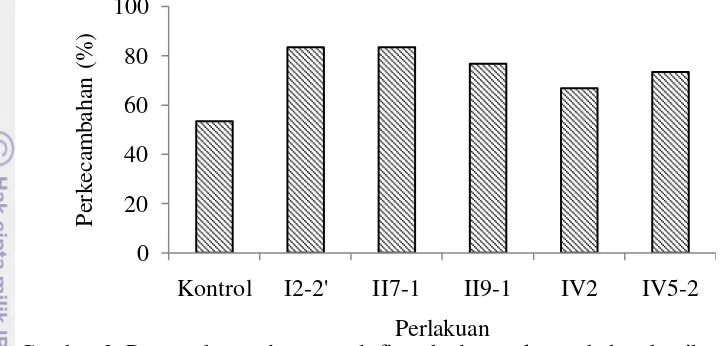
Pengaruh Cendawan Endofit terhadap Perkecambahan Benih Cabai Perendaman benih cabai menggunakan cendawan endofit mampu meningkatkan persentase perkecambahan benih. Benih cabai yang diinokulasi cendawan endofit menunjukkan persentase perkecambahan lebih tinggi dibandingkan dengan kontrol. Persentase perkecambahan benih cabai yang diinokulasikan cendawan endofit I2-2’ dan II7-1 sama yaitu sebesar 83.33%,
sedangkan pada cendawan endofit II9-1, IV2 dan IV5-2 masing-masing sebesar
76.67%, 66.67%, dan 73.33% (Gambar 2).
Inokulasi cendawan endofit pada benih dapat meningkatkan persentase perkecambahan. Menurut Dearnaley dan Brocque (2006), kolonisasi cendawan endofit memiliki peran penting pada saat perkecambahan. Pada saat berkecambah, benih hanya memiliki sedikit kandungan nutrisi esensial. Pada saat benih berkecambah, hifa cendawan endofit mengolonisasi jaringan tanaman dan membentuk struktur yang disebut pelotons. Struktur tersebut digunakan cendawan endofit untuk melakukan pertukaran nutrisi, sedangkan benih mendapatkan suplai gula dan substansi anorganik (nitrogen dan fosfor) yang berguna bagi pertumbuhan tanaman muda tersebut. Selain itu, beberapa cendawan endofit memproduksi fitohormon seperti etilen, auksin dan sitokinin (Bacon dan Hinton 2002; Tan dan Zon 2001).
Penelitian Xin et al. (2009) menunjukan bahwa beberapa strains cendawan endofit mampu memproduksi hormon indole-3-acetic acid (IAA) setelah diinkubasi oleh triptofan. Cendawan endofit Williopsis saturnus asal tanaman jagung mampu menghasilkan hormon perangsang pertumbuhan tanaman, yaitu IAA dan indole-3-pyruvic acid (IPYA) (Nassar et al. 2005). Sementara itu penelitian Kavroulakis et al. (2007) menunjukan bahwa cendawan endofit memiliki jalur metabolisme untuk membentuk etilen dan pembentukan hormon etilen tersebut dijadikan salah satu cara kerja bagi cendawan endofit dalam menginisiasi ketahanan tanaman inang terhadap cendawan Fusarium Solani. Dengan adanya hormon perangsang pertumbuhan tersebut maka persentase perkecambahan benih cabai dengan perlakuan cendawan endofit lebih tinggi dibandingkan dengan kontrol.
Persentase perkecambahan tertinggi yaitu lebih dari 80% ditunjukkan oleh tanaman cabai dengan perlakuan perendaman cendawan endofit I2-2’ dan II7.
Melalui perendaman, cendawan endofit ditransmisikan melalui biji tanaman inang, kemudian hifa cendawan tumbuh berkembang dalam ovul (Clay 1993 dalam Istikorini 2008). Persentase perkecambahan lebih dari 80% menunjukkan persentase perkecambahan yang tinggi sehingga benih memiliki viabilitas dan mutu fisiologis yang baik. Tinggi rendahnya viabilitas benih dapat diukur dengan daya kecambah benih dan bobot kering kecambah normal. Mutu fisiologis benih adalah tinggi rendahnya daya hidup atau viabilitas benih tercermin dari nilai daya berkecambah, kecepatan tumbuh, keserempakan tumbuh (Widajati et al. 2013).
0
Kontrol I2-2' II7-1 II9-1 IV2 IV5-2
P
sehingga tanaman lebih tahan dibandingkan dengan tanaman yang tidak terinfeksi cendawan endofit (Clay 1988 dalam Istikorini 2008).
Gambar 2 Pengaruh cwndawan endofit terhadap perkecambahan benih cabai
Pertumbuhan Populasi P. latus
Pertumbuhan populasi P. latus dipengaruhi oleh beberapa faktor lingkungan seperti iklim, sistem budidaya dan keberadaan musuh alami.
Perlakuan cendawan endofit pada tanaman cabai mampu mempengaruhi pertumbuhan populasi P. latus. Secara umum, peningkatan toleransi tanaman terhadap serangan serangga herbivora disebabkan oleh produksi senyawa alkaloid oleh cendawan endofit (Faeth dan Saari 2012). Pada kondisi normal, tanaman juga dapat memproduksi berbagai senyawa alkaloid seperti glucosinolates, phenolic glycosides, dan terpenes yang berpotensi meningkatkan ketahanan tanaman terhadap serangga herbivora tanpa bantuan dari cendawan endofit, akan tetapi tanaman memiki keterbatasan karena selain untuk pertahanan, tanaman juga harus mengalokasikan sumber daya tersebut untuk pertumbuhan dan reproduksi. Selain itu, keberadaan cendawan endofit juga memiliki peran dalam memperlambat mekanisme resistensi serangga herbivora terhadap metabolit sekunder yang diproduksi tanaman. Sifat resistensi dari serangga herbivora biasanya bersifat spesifik, sehingga dengan adanya metabolit yang dihasilkan cendawan endofit, mekanisme resistensi serangga hama akan sedikit dihambat.
Beberapa jenis senyawa alkaloid yang dihasilkan oleh cendawan endofit diketahui bersifat racun terhadap serangga. Menurut Tan dan Zon (2001), senyawa alkaloid lolines, agroklavin dan elimoklavin yang dihasilkan cendawan endofit Neotyphodium bersifat racun syaraf terhadap serangga mamalia dan herbivora. Senyawa 2α-Hydroxydimeninol dan pestalotiopsins yang dihasilkan cendawan endofit Pestalotiopsis spp. bersifat toksik terhadap larva ulat pucuk Choristoneura fumiferana. Kemampuan cendawan endofit menghasilkan senyawa alkaloid yang bersifat racun terhadap beberapa serangga hama dapat menurunkan populasi hama tersebut.
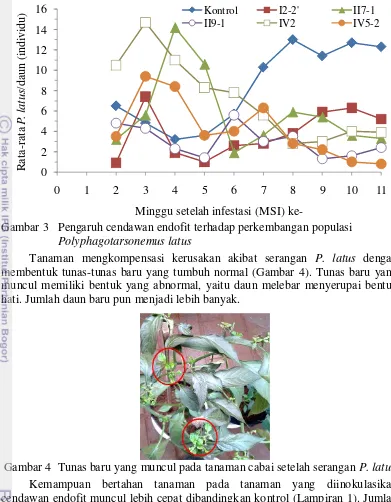
minggu berikutnya populasi P. latus pada cabai dengan perlakuan cendawan endofit menurun, sedangkan pada tanaman kontrol meningkat (Gambar 3). Hal tersebut disebabkan tingkat infeksi cendawan endofit pada jaringan tanaman meningkat secara konstan seiring dengan meningkatnya umur jaringan tanaman (Stone et al. 2004).
Populasi P. latus pada tanaman cabai yang diinokulasikan cendawan endofit II7-1 berbeda nyata dengan kontrol pada 4 MSI dengan populasi yang lebih
tinggi dibandingkan dengan kontrol (Gambar 3). Pada 8 sampai 11 MSI perkembangan populasi P. latus pada tanaman yang diinokulasi cendawan endofit II9-1 dan cendawan endofit IV2 lebih rendah dan berbeda nyata dengan kontrol.
Pada tanaman yang dinokulasikan cendawan endofit II7-1 dan cendawan endofit
IV5-2, populasi P. latus berbeda nyata dengan kontrol pada 9-11 MSI. Pada 11
MSI, pertumbuhan populasi P. latus pada seluruh perlakuan cendawan endofit menunjukan perbedaan yang nyata dan lebih rendah dari pada kontrol. Masing-masing perlakuan cendawan endofit menunjukan pengaruh yang tidak berbeda nyata terhadap pertumbuhan populasi P. latus pada 0 sampai 3 MSI, begitupun pada 5 sampai 11 MSI. Pada 4 MSI, perlakuan cendawan endofit II7-1 memberikan
pengaruh yang berbeda nyata dengan perlakuan cendawan I2-2’ dan II9-1 terhadap
pertumbuhan P. latus.
Efektivitas dari cendawan endofit terhadap hama bervariasi dan tergantung pada genotif dari cendawan dan tanaman inangnya serta faktor lingkungan. Kemampuan cendawan endofit dalam menolak keberadaan serangga, menginduksi penurunan bobot tubuh, penurunan laju pertumbuhan dan perkembangan populasi serangga, dan meningkatkan tingkat kematian serangga berkorelasi positif dengan produksi senyawa alkaloid yang bersifat racun. Akan tetapi, cara kerja cendawan endofit tergantung pada kemampuan cendawan endofit tersebut dalam menciptakan lingkungan yang tidak mendukung bagi pertumbuhan serangga (Azevedo et al. 2000).
Semua cendawan endofit yang diinokulasikan mampu menghambat pertumbuhan P. latus, walaupun memerlukan waktu yang lama. Penghambatan tersebut bisa terjadi pada siklus hidup, lama hidup maupun keperidian. Kolonisasi cendawan pada tanaman cabai menyebabkan terinduksinya produksi senyawa metabolit sekunder yang bersifat antagonis terhadap arthropoda herbivor (Faeth 2002). Kolonisasi hifa endofit menyebabkan jaringan tanaman yang menjadi sumber nutrisi bagi serangga herbivora ataupun arthropoda hama lain terkontaminasi oleh metabolit sekunder yang dihasilkan cendawan endofit tersebut. Akan tetapi, tingkat persentase metabolit yang dikonsumsi masing-masing jenis artropoda herbivor sulit untuk ditentukan (Stone et al. 2004).
Gambar 3 Pengaruh cendawan endofit terhadap perkembangan populasi Polyphagotarsonemus latus
Tanaman mengkompensasi kerusakan akibat serangan P. latus dengan membentuk tunas-tunas baru yang tumbuh normal (Gambar 4). Tunas baru yang muncul memiliki bentuk yang abnormal, yaitu daun melebar menyerupai bentuk hati. Jumlah daun baru pun menjadi lebih banyak.
Gambar 4 Tunas baru yang muncul pada tanaman cabai setelah serangan P. latus Kemampuan bertahan tanaman pada tanaman yang diinokulasikan cendawan endofit muncul lebih cepat dibandingkan kontrol (Lampiran 1). Jumlah daun baru yang muncul pada tanaman yang diinokulasikan cendawan endofit terlihat lebih banyak dari pada kontrol. Tingkat infeksi cendawan endofit meningkat seiring pertumbuhan tunas-tunas baru (Stone et al 2004). Pada 8 MSI, tunas-tunas baru pada tanaman yang diinokulasi cendawan endofit II7-1 tumbuh
normal dan tanaman terlihat lebih sehat dibandingkan kontrol (Gambar 5). Dengan adanya simbiosis mutualisme cendawan endofit tersebut dengan jaringan tanaman, maka pertumbuhan populasi P. latus terhambat, gejala kerusakan berkurang dan tanaman menjadi lebih sehat dibandingkan kontrol.
(a) (b)
Gambar 5 Pengaruh cendawan endofit terhadap pertumbuhan tunas baru tanaman cabai kontrol (a) dan diinokulasi cendawan endofit II7-1 (b) pada 8 MSI
Siklus Hidup P. latus
Pertumbuhan populasi P. latus dipengaruhi oleh siklus hidup dan reproduksinya. Siklus hidup P. latus meliputi telur, larva, nimfa dan imago. Telur berukuran rata-rata 0.06 mm diletakkan satu per satu pada permukaan bawah daun (Gambar 6a). Telur berbentuk oval dan memiliki bagian dasar yang pipih untuk menempel pada permukaan daun. Sebagian besar permukaan telur transparan kecuali pada permukaan atas memiliki garis longitudinal berwarna putih (Zhang 2003).
Larva berukuran rata-rata 0.1mm dan memiliki 3 pasang tungkai (Gambar 6b). Larva berwarna putih pucat ketika pertama menetas dan segera berubah menjadi transparan. Larva aktif bergerak dalam mencari makan. Larva menyelesaikan tahap perkembangannya tanpa mengalami perubahan yang berarti (Krantz 1978). Larva yang telah menyelesaikan stadianya berkembang menjadi stadia yang berdiapause disebut nimfa. Nimfa berukuran rata-rata 0.16 mm dan merupakan stadia yang inaktif (Gambar 6c).
daun yang baru (Gambar 6f). Imago jantan akan segera kopulasi dengan nimfa betina yang telah menyelesaikan stadianya. Imago betina P. latus dapat menyebar dengan menempel pada tibia dan tarsus dari beberapa seranggga seperti kutu daun (Myzus persicae) dan kutu kebul (Bemisia tabaci) (Hoy 2011).
(a) (b) (c)
(d) (e) (f)
Gambar 6 Fase perkembangan Polyphagotarsonemus latus (a) telur (b) larva (c) nimfa (d) imago betina (e) imago jantan (f) kopulasi
Lama siklus hidup P. latus adalah lama perkembangan telur hingga terbentuk imago. Pada tanaman cabai, lama perkembangan stadia telur, larva dan nimfa pada P. latus jantan berkisar antara 2.67-2.71, 2.42-3.0 dan 2.75-3.0 hari, sedangkan pada P. latus betina berkisar antara 2.5-2.55, 1.89-2.2, dan 2.74-3.13 hari pada suhu 25 ºC dan kelembaban 75% (Montasser et al. 2011).
Lama stadia telur jantan dan betina pada daun cabai dengan perlakuan cendawan endofit tidak berbeda nyata dengan kontrol. Pada daun dengan perlakuan cendawan endofit II9-1 dan IV2, lama stadia telur jantan dan betina lebih
lama dari pada kontrol (Tabel 1).
Lama stadia larva pada daun cabai dengan perlakuan cendawan endofit tidak berbeda nyata dengan kontrol (Tabel 1). Waktu yang dibutuhkan untuk meyelesaikan stadia larva pada daun cabai dengan perlakuan cendawan endofit lebih lama dibandingkan dengan kontrol, kecuali pada cendawan endofit II9-1.
Larva betina memiliki waktu lebih lama dibandingkan larva jantan untuk menyelesaikan fase perkembangannya. Selain pada daun dengan perlakuan cendawan endofit II9-1, lama larva jantan pada daun dengan perlakuan cendawan
endofit IV5-2 lebih cepat dibandingkan dengan kontrol.
Lama perkembangan nimfa jantan pada daun dengan perlakuan cendawan endofit tidak berbeda nyata dengan kontrol (Tabel 1). Waktu yang dibutuhkan
0.06 mm
0.1 mm
0.16 mm
0.2 mm
Jantan Betina Jantan Betina Jantan Betina Jantan Betina
Kontrol 2.17a 2.50a 2.32a 2.50a 1.57ab 1.58b 6.29a 6.25b
I2-2’ 2.17a 2.00a 2.41a 2.75a 1.66ab 1.56b 6.23a 6.51ab
kecuali pada cendawan endofit IV5-2.
Lama perkembangan nimfa betina pada daun dengan perlakuan cendawan endofit II7-1 lebih lama dan berbeda nyata kontrol (Tabel 1). Waktu yang
dibutuhkan untuk menyelesaikan stadia tersebut yaitu 1.90 hari sehingga siklus hidup P. latus pada perlakuan tersebut lebih lama. Selain pada cendawan endofit II7-1, lama perkembangan nimfa betina pada daun cabai dengan perlakuan
cendawan endofit II9-1, IV2 dan IV5-2 lebih lama dibandingkan dengan
kontrol,yaitu 1.65 hari, 1.66 hari dan 1.76 sehingga siklus hidup P. latus pada daun tersebut lebih lama. Lama stadia nimfa betina pada masing-masing perlakuan cendawan endofit tidak berbeda nyata, kecuali perlakuan cendawan endofit II7-1 dengan I2-2’, sedangkan lama stadia nimfa jantan, hanya perlakuan
cendawan endofit II7-1 yang berbeda nyata dengan perlakuan cendawan endofit
IV5-2. Total siklus hidup P. latus jantan pada daun cabai dengan perlakuan
cendawan endofit tidak berbeda nyata dengan kontrol, sedangkan siklus hidup P. latus betina pada perlakuan cendawan endofit II7-1 II9-1 dan IV2 berbeda nyata
dengan kontrol. Akan tetapi, masing-masing perlakuan cendawan endofit tidak memberikan pengaruh yang berbeda nyata terhadap siklus hidup P. latus.Dengan adanya siklus hidup yang lebih lama maka pertumbuhan populasi P. latus berjalan lebih lambat. Menurut Hermawati (2007), proses metabolisme A. gossypii terganggu dengan adanya metabolit sekunder dari cendawan endofit dalam tubuhnya, sehingga hormon yang berperan dalam pergantian stadia pun terganggu dan akhirnya waktu yang dibutuhkan lebih lama.
Lama perkembangan nimfa betina pada daun cabai dengan perlakuan cendawan endofit I2-2’ lebih cepat dibandingkan dengan perlakuan cendawan yang
lain dan tidak berbeda nyata dengan kontrol sehingga waktu yang dibutuhkan untuk menyelesaikan siklus hidup lebih cepat (Tabel 1). Perlakuan cendawan tersebut tidak mempengaruhi siklus hidup P. latus, sehingga pertumbuhan populasinya pun tidak terlalu terhambat.
Tabel 1 Pengaruh cendawan endofit terhadap lama perkembangan pradewasa Polyphagotarsonemus latus
a
disebabkan suhu yang lebih tinggi di laboratorium, yaitu rata-rata 26.8 ºC. Menurut Zhang (2003), semakin tinggi suhu di rumah kaca, maka lama stadia tungau lebih cepat.
Biologi P. latus
Setiap makhluk hidup memiliki kemampuan untuk berkembangbiak untuk melestarikan keturunannya. Kemampuan berkembangbiak antara lain dipengaruhi oleh keperidian, perbandingan nisbah kelamin dan lama hidup (Krantz 1978).
Perkawinan imago P. latus pada masing-masing perlakuan menghasilkan populasi imago betina lebih banyak dari pada imago jantan (Tabel 2). Imago betina yang tidak dibuahi meletakkan telur semuanya jantan, sedangkan imago betina yang dibuahi meletakkan telur betina dan jantan dengan perbandingan 4:1 (Fasulo 2013). Pada daun dengan perlakuan cendawan endofit, jumlah imago betina lebih rendah dibandingkan dengan kontrol, sehingga keturunan yang dihasilkan pada masa berikutnya lebih rendah dan populasinya pun lebih rendah. Cendawan endofit asal padi Fusarium moniliforme mampu mempengaruhi nisbah kelamin dari Meloidogyne graminicola dengan jantan yang lebih banyak dibandingkan dengan betina sehingga pertumbuhan populasi dapat berjalan lebih lambat (Huong 2010 dalam Yulianti 2013).
Imago jantan berperan membawa nimfa betina untuk berpindah ke daun lain. Setelah nimfa betina berkembang menjadi imago, imago jantan segera mengawini imago betina tersebut dan kemudian imago jantan akan segera mati. Lama hidup imago jantan pada daun dengan perlakuan cendawan endofit II7-1
berbeda nyata dan lebih lama dibandingkan dengan kontrol (Tabel 2). Hal tersebut menjadi indikator terhambatnya waktu perkawinan imago jantan dengan imago betina. Terhambatnya waktu perkawinan tersebut disebabkan oleh stadia nimfa betina yang lebih lama sehingga imago jantan tidak dapat segera mengawini P. latus betina yang masih berada dalam stadia nimfa. Waktu kopulasi imago yang terhambat mengakibatkan waktu regenerasi P. latus terhambat, sehingga pertumbuhan populasinya pun menjadi lebih lambat.
Lama hidup imago betina pada daun dengan perlakuan cendawan endofit tidak berbeda nyata dan lebih lama dibandingkan dengan kontrol. Lama hidup imago betina tergantung dari masa praoviposisi, masa oviposisi, dan masa pascaoviposisi. Imago betina melewati masa sebelum peletakkan telur pertama yang disebut praoviposisi. Masa praoviposisi P. latus pada perlakuan cendawan endofit II7-1, II9-1 dan IV5-2 lebih lama dan berbeda nyata dengan kontrol, akan
tetapi masing-masing perlakuan cendawan endofit tidak memberikan pengaruh yang berbeda nyata (Tabel 2). Masa praoviposisi yang lebih lama akan menunda waktu untuk meletakkan telur harian (oviposisi) sehingga lama hidup imago betina lebih lama.
Kontrol I2-2’ II7-1 II9-1 IV2 IV5-2 Masa praovipos isi (hari) 0.99b 1.13ab 1.26a 1.16a 1.10ab 1.17a Masa ovipos isi (hari) 7.45a 7.87a 7.78a 7.92a 7.90a 8.00a Masa pascaoviposisi (hari) 0.38a 0.25a 0.22a 0.15a 0.17a 0.13a Keperidian (telur) 40.57a 36.03ab 34.32b 33.57b 34.73b 33.50b Lama hidup imago (hari)
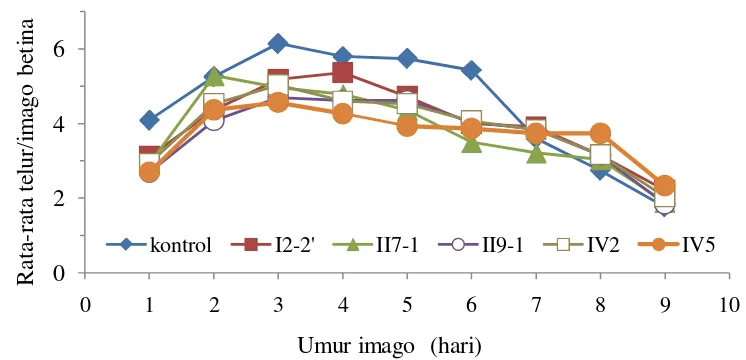
Imago betina P. latus meletakkan 30 sampai 76 telur di permukaan bawah daun (Montasser et al. 2011). Keperidian imago betina pada perlakuan cendawan endofit lebih rendah dan berbeda nyata dengan kontrol, kecuali pada perlakuan cendawan endofit I2-2’. Keperidian imago betina pada masing-masing perlakuan
cendawan endofit menunjukan pengaruh yang tidak berbeda nyata. Jumlah telur yang diletakan pada cendawan endofit II7-1, II9-1, IV2 dan IV5-2 yaitu 34.32 hari,
33.57 hari, 34.72 hari dan 33.5 hari. Hal tersebut menunjukkan semua cendawan endofit yang diaplikasikan mampu menghambat keperidian P. latus. Dengan keperidian yang lebih rendah walaupun masa oviposisi lebih lama maka keturunan yang dihasilkan berkurang dan pertumbuhan populasi pada tanaman dengan perlakuan cendawan endofit tersebut lebih lambat dibandingkan dengan kontrol. Menurut Yulianti (2013), salah satu pengaruh dari cendawan endofit mengurangi kerusakan dari serangan seranggga herbivora melalui penurunan keperidian, penurunan laju pertumbuhan dan perkembangan, sehingga populasi serangga menurun. Penelitian Jaber (2010) menunjukkan bahwa keperidian Aphis fabae merupakan parameter biologi yang terpengaruh oleh cendawan endofit secara signifikan.
Tabel 2 Pengaruh cendawan endofit terhadap sifat biologi imago Polyphagotarsonemus latus
a
Angka dalam lajur yang sama diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan uji Duncan pada taraf nyata 5%
Hal tersebut ditunjukkan oleh turunnya jumlah telur yang dihasilkan nematoda pada akar tanaman baik jumlah massa telur maupun jumlah telur per massa telur yang diletakkan. Dengan jumlah telur per hari yang diletakkan imago betina pada daun yang diinokulasikan cendawan endofit lebih sedikit maka generasi P. latus berkurang dan populasinya pun menjadi lebih rendah dibandingkan dengan kontrol.
Jumlah telur yang diletakkan imago betina mulai menurun pada hari ke-8 dan hari ke-9. Pada hari ke-10, imago betina masuk pada tahap pascaoviposisi dan sudah berhenti bertelur.
Gambar 7 Pengaruh cendawan endofit terhadap keperidian P. latus 0
SIMPULAN DAN SARAN
Simpulan
Perlakuan cendawan endofit pada benih cabai dapat meningkatkan viabilitas benih. Beberapa jenis cendawan endofit dapat memperpanjang masa praoviposisi, menekan pertumbuhan populasi, menurunkan keperidian dan meningkatkan proporsi individu jantan P. latus.
Saran
Agrios GN. 1997. Plant Pathology.Tokyo (JP): Pres Tokyo.
Azevedo JL, Jr WC, Pereira JO, Araújo WL. 2000. Endophytic microorganisms: a review on insect control and recent advances on tropical plants. EJB. 3:1. Bacon CW, Hinton DM. 2002. Endopytic and biological control potential of
Bacillus mojavensis and related species. Biolog Con. 23:274-284.
Bing LA, Lewis LC. 1991. Suppression of Ostrinia nubilalis (Hübner) (Lepidoptera: Pyralidae) by endophytic Beauveria bassiana (Balsamo) Vuilemin. Environ Entomol. 20:1207-1211.
[BPS] Badan Pusat Statistik. 2013. Produksi Cabai Besar, Cabai Rawit, dan Bawang Merah. Jakarta (ID): BPS.
Dearnaley JDW, Brocque AFL. 2006. Endophytic fungi associated with Australian orchids. In press Australasian Mycologist.
Fasulo TR. 2013. Broad mite, Polyphagotarsonemus latus (Banks) (Arachnida: Acari: Tarsonemidae). EENY-183. [Internet] [diunduh 2013 Sep 15]. Tersedia pada: http://edis.ifas.ufl.edu/pdffiles/IN/IN34000.pdf?.
Faeth SH. 2002. Are endophytic fungi defensive plant mutualists?. Oikos. 98(1): 25-36.
Faeth SH, Saari S. 2012. Fungal grass endophytes and arthropod communities: lessons from plant defence theory and multitrophic Interactions. Fungal Ecol. 5:364-371.
Gao F, Dai C, Liu X. 2010. Mechanisms of fungal endophytes in plant protection against pathogens. Afr J Microbiol Research. 4(13):1346-1351.
Hermawati H. 2007. Pengaruh cendawan endofit terhadap biologi dan pertumbuhan populasi Aphis gossypii Glov. (Homoptera: Aphididae) pada tanaman cabai [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Hoy MA. 2011. Agricultural Acarology: Introduction to Integrated Mite Management. New York (US): CRC Press.
Istikorini Y. 2008. Potensi cendawan endofit untuk mengendalikan penyakit antraknosa pada cabai (Capsicum annum L.) [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Jaber LR. 2010. The effects of the root endophytic fungus Acremonium strictum on plant-herbivore interactions. [disertasi]. Göttingen (DE): University Göttingen.
Jeppson LR. 1975. Mites Injurious to Economis Plants. California (USA): University of California Press.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Laan PA van der, penerjemah. Jakarta (ID): Ichtiar Baru-van Hoeve. Terjemahan dari: De Plagen vande Culturgewassen in Indonesie.
endophyte-free tall fescue. Plant Disease. 757-761.
Krantz GW. 1978. A Manual of Acarology Second Edition. Corvallis (US): Oregon State University Book Store.
Montasser AA, Taha AM, Hanafy, ARI, Hassan GM. Biology and control of the broad mite Polyphagotarsonemus latus (Banks, 1904) (Acari: Tarsonemidae). IJESE. [Internet]. [diunduh 2013 Sep 15]; 1(1):26-34. Tersedia pada: %20the %20 broad...?.
Nassar AH, El-Tarabily KA, Sivasithamparam K. 2005. Promotion of plant growth by an auxin-producing isolate of the yeast Williopsis saturnus endophytic in maize (Zea mays L.) roots. Biol Fertil Soils. 42:97-108.
Purwati E, Jaya B, Duriat AS. 2000. Penampilan beberapa varietas cabai dan uji resistensi terhadap penyakit virus kerupuk. J Hort. 10(2):88-94.
Setiawati W, Udiarto BK, Muharam A. 2005. Pengenalan dan Pengendalian Hama-hama Penting pada Tanaman Cabai Merah. Bandung (ID): Balitsa, Balitbang Pertanian.
Stone JK, Polishook JD, White JF. 2004. Endophytic fungi. Di dalam: Mueller GM, Bills GF, Foster MS. Biodiversity of Fungi. California (USA): Elsevier Academic Press.
Tan RX, Zon WX. 2001. Endophytes: a rich source of functional metabolites. Nat Prod Rep. 18:448–459.
Widajati E, Murniati E, Palupi ER, Kartika T, Suhartanto MR, Qadir A. 2013. Dasar Ilmu dan Teknologi Benih. Bogor (ID): IPB Press.
Xin G, Glawe D, Doty SL. 2009. Characterization of three endophytic, indole-3-acetic acid producing yeasts occurring in Populus trees. Myco Research. 1:973-980.
Yulianti T. 2013. Pemanfaatan Endofit sebagai agensia pengendali hayati hama dan penyakit tanaman. Buletin Tanaman Tembakau, Serat & Minyak Industri. 5(1):40-49.
Tabel 1 Kompensasi kerusakan Polyphagotarsonemus latus pada tanaman cabai yang diinokulasi cendawan endofit Minggu setelah
infestasi ke-
Perlakuan
Kontrol I2-2’ II7-1 II9-1 IV2 IV5-2
4
6
Polyphagotarsonemus latus per pada tanaman cabai
Perlakuan Jumlah P. latus pada minggu setelah infestasi (MSI)
ke-a
0 2 3 4 5 6 7 8 9 10 11
Kontrol 5a 6.5a 4.8a 3.2b 3.6a 5.7a 10.3a 13.0a 11.4a 12.7a 12.3a I2-2’ 5a 0.9a 7.4a 1.9b 1.0a 2.6a 2.8a 3.8ab 5.9ab 6.3ab 5.2b II7-1 5a 3.2a 5.6a 14.2a 10.6a 1.9a 3.6a 5.9ab 5.4b 4.0b 3.3b II9-1 5a 4.8a 4.3a 2.3b 1.4a 5.6a 3.0a 3.5b 1.3b 3.6b 2.4b IV2 5a 10.5a 14.7a 11.0ab 8.3a 7.8a 5.6a 2.8b 3.0b 1.6b 3.9b IV5-2 5a 3.5a 9.4a 8.4ab 3.6a 4.0a 6.3a 5.8ab 2.2b 1.0b 0.8b
a
RIWAYAT HIDUP
Penulis dilahirkan di Tasikmalaya pada tanggal 26 Juli 1992 dari ayah Tasdik dan ibu Aminah. Penulis adalah putri ketiga dari empat bersaudara. Tahun 2010 penulis lulus dari SMA Negeri 5 Tasikmalaya dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB dan diterima di Departemen Proteksi Tanaman, Fakultas Pertanian. Penulis merupakan penerima Beasiswa Bidik Misi.
Selama mengikuti perkuliahan, penulis menjadi asisten praktikum Virologi Tumbuhan pada tahun ajaran 2011/2012, asisten praktikum Ilmu Penyakit Tumbuhan Dasar tahun ajaran 2012/2013 dan asisten praktikum Ilmu Penyakit Tumbuhan Dasar tahun ajaran 2013/2014. Penulis pernah aktif sebagai staf Departemen Sumber Daya Manusia UKM Lises Gentra Kaheman periode 2012, bendahara Himpunan Mahasiswa Tasikmalaya (HIMALAYA), ketua Divisi Kewirausahaan HIMASITA IPB. Penulis juga pernah mengikuti kepanitiaan sebagai staff divisi Dana Usaha TPB Cup 2011 dan Masa Perkenalan Mahasiswa Baru (MPKMB) IPB, ketua pelaksana Pemilihan Mahasiswa Berprestasi Departemen Proteksi Tanaman 2012, sekretaris umum pada Festival Seni Budaya (FSB) IPB, dan ketua divisi Konsumsi pada Plant Protection Even (NPV) 2012.