m E R I S A S I
BIOLOGI, SEROLOGI DAN
ANALISIS
SIDIK
JAR1
DNA VIRUS
PENYEBAB PENYAKIT
DAUN
KF,RITING KUNING CABAI
Oleh:
Sri Sulandari
SEKOLAH
PASCASARJANA
SRI SULANDARI. Karaherisasi Biologi, Serologi
dan
Analisis Sidik Jari DNA Virus Penyebab Penyakit Daun Keriting Kuning Cabai.Dibimbing
oleh RUSMlLAH SUSENO,SRI
HENDRASTUTI HIDAYAT, -0HARJOSUDARMO,
dan
SOEMARTONO SOSROMARSONO.Sejak tahun
2000
tejadi peningkatan kejadim penyakit &un keriting kuning cabaidi
Indonesia sehinga menirnbulkan kerugian besarpada p
e m cabai. Penyebab penyakit tersebut sdalah geminivirus.Penelitian dilakukan uutuk mengetahui (1) intensitas dan kejadian penyalut di lapangan; (2) mengidentifikasi virus penyebab penyalut melalui kajian sifat-sifat biologi, serologi
dm
p l a sidik jari DNA virus. Metode penelitian meliputi: (1) s w e i di lapangan;(2)
kajian
kisaran inang dan hubungan virus penyehab penydatd e n p
seraflgga vektornyadi
rumah kaca, (3)pembutan
antibodipolildonal
drrn
kajian serologinya; (4) diferensiasi isoht geminivims melalui polasidik
jari
DNA b e r d a w h po/ymerase chain retactton-
restriction fiagmenr length polymorphism (PCR-RFLP), ( 5 )analisis hubungan
kekerahatan mtarisolat
geminivhs melalui pembandingan runutan susunan DNAHasiI pengamatan
di
hpmgmntenunjukkan
bshwaepidemi
penyakitdam
keriting kuning cabai telabterJadi
di tiga Propinsi diIndonesia,
yaitu Daerah Istimewa Yogyakarta,Jawa
Tengahdan
Jawa Barat sej& tahun 2000. Tingkat kejadian penyakit path cabai rawit ( Capictm: fiurescem) mencapai 100% sedangkan pada cabai besar(C.
annuurn) terdapt secara sporadis yaitu sekitar 10-
35%, kecuali pada kultivar cabai besarTM
999 dengan tingkat kejadian penyakit mencapai 70-
100%. Semua kultivar abi yang diuji rentan terhadap virus penyebab penyakit dam keritingkuning.
Gemhivirus tersebut mempunyai kisaran hang yang cukup luas me1 iputi familiSolanaceae
(cabai, tomat, terong, tembalrau dan ceplukan), Legurn inosac (kedelai, ksrcang pnjmg, kacang hijau, orok-orok), dan Compositae (bunga matahari, babadotan, dan Hyptis sp. ). Tanaman dari famili Cucurbitmeae, Malvaceae, Chenopodiaceaeclan
Ammthaceae tahan terhadap infeksi penyebab penyakit tersebut.Kutukebul tembakau (Bemisio tahci Genn.),
merumhi
vektor v i m penyebab penyakit daun keritingkuning
cabai yang efektif. ~ a t u ekor kutukebul yang viruli ferus sudah clapat menularkan geminiviruske
tanaman cabai. Geminivirus isolat cabai ditularkan oleh kutukebul tembakau secara persisten, tetapi tidak diturunkanke
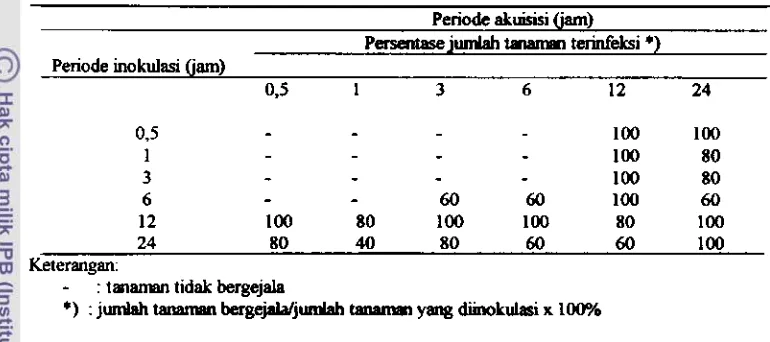
pnerasi berikutnya ( non tramovarial transmission). Periode akuisisi dan inokulasi yang optimal untuk penularan geminivirus adalah 3 - 6 jam, memerlukan periode laten di &lamtubuh
vektor minimal 9jam,
clan pacia p e r c o b ini, p e r i d retensinya sampi serangga mati. P e n u l m virusakan
lebih efektif apabila periode akuisisidan
inokulasioyafernakin
lama serta s m a bbanyak
jurnlah kutukebulnya. Kutukebul tembakaubetina lebih
efektif menularkangeminivirus dibandingkan yang jantan.
rnengkilkan virus
rnurni sekihr
220 ugdari
250 g bahmsew.
Berdasarkan analisis elektroforesis secara SDS-PAGE,
ukuran
protein selubung geminivim sekitar 29k
Da.Antiserum
yangdihasilkoln
&patdigunalcan
untuk
mendeteksi geminivirus
dari sampel
tanaman
saldt menggunakanmetode
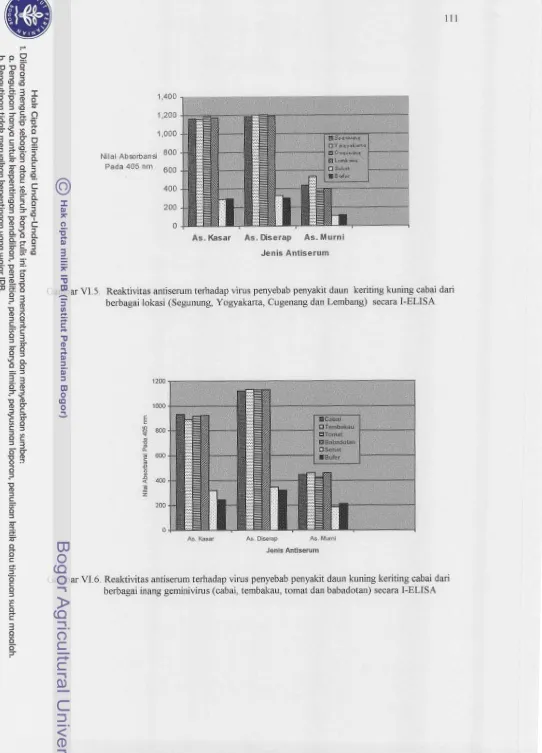
I-EZ.ISA ataupun DIBA Reaktivitas antiserum tersebut sama terhdap antigen isolat mbai dari berbagail o h i
(Segunung, Yogydmta, Cugamg
dm
Lcmbang)
mupun antigenc
h
i
h h g a i inang (cabai,tembakau,
tomatdm
babadotan). Antiserum j u g &pat digunakanuntuk mendeteksi virus penyebat, penyakit daun keriting Mng p d a serangga vektornya. Menggumlm
satu
ekor serangga sudah
&patdidetebi keberadem
geminivirus &lamtub&
B. tabacid
e
w
baik.
Teknik serologi I-ELISAdan
DIBA sangat ptensial untuk digunakaashagti
alat deteksigeminivim.
Idmtifikasi penyebab penyakit daun keritingkuning cabi
jwdap&
dilakukan
meldui analisis western-bZottPenyebab penyakit daun ken'ting
kunrng
cabai
d a h h gerninivirus ymg &pat terdeteksi dengan baikmenggunakan sampel
DNA
dari
jaringan ttinaman sakit maupun serangga vektornyamelalui
t e W PCR menggudmempat
pasang primer, pALlv 1978 & pALlc 715, pALlv 1978 & pALlc 4% pUPvl & pllPv2,dm
pAv 494 & PAC 1048.Geminivirus jugs &pat
diidwtifikasi
melalui
isobiDNA
p d a jarin* tanaman sakit denganmeagguaakan metode gumidin
U l i nDua
m e nDNA
berukumn 2600 bpdrur
1600 bp yang diperolehmeucerminh konformasi
DNA
(bentuk sirkuler dan Iinier) @virus.Hasil pernotongan dengan enzim restriksi (BamHI, EcoRI, Hindm, clan
Pd)
SRI SLJUNDARI.
Bidogical
Chm&zhtion, Serologicoll Assayand
DNA
Finger Printing Analysisof Pepper
yellow l@ml v i m . Supervised by RUSMILAHSUSENO, SRI HENORASIVTI HIDAYAT,
-0
HARJOSUDARMO,AND
SOEMARTONO SOSROMARSONO.High
incidence of
pepper
yellow leaf curlvirus was
observed
in Indonesia
since
2000.
The
disease causes serious yieldreduction
inmany
fields.
Cieminvirusis
the
causal agent of thedisease
The aims of
this
research
is (1) to confirm the disease incidenceand
severityin
thefield;
(2) to identifythe
Pepper yellow la$ csrrl v i m by biological characterization, serological assayarid
DNA fhgx printinganalysis.
To
acbievethe
obyectiveof
the researchthe
mekdology
chosen involvedfield
survey
and identification of thecausal
agentthro*
(1)host
range and vim-vector relationships study; (2) antiserum productionand
serological assay of thevirus;
( 3 ) PCR-RFLP- basedDNA
finger printing to shtdy thegeminivirus
diversity; (4)DNA
sequencing analyse similarity amongthe
&minivim
Field observation s b w d that
the
epi-cof
pepptr @ow leaf cur1 diseaseO C C
in
~three provincesof
lndomsia, i-e. : Special provinceof
YO@CN@ Central Java,and
WestJava
since the year2000.
Thedisease
incidence
found
on
Capsamjwrescem wasup to 100percentbut
on
C anmnnrritwasintherangeof lot035 per cent. Asan
exception, thedisease
incidence
onTM
999 cultivarof
C. annuum was in the rangeof
70-
100 per cat.AU
of
the pepper cultiyarstested
were s q b l e to the Pepper yellow leaf curl vW.The
Pepper
p l h l@cwIVW
has
an
irdemaediate ht range incldmg plants belong to the family of Solamume @pp, tomato,eggplant,tobacco,
and
ground cherry, Leguminosae (soyban, long k a q green bean and CrotaIaria sp.), and Compositae (sun flower, Ag- conyzoih,d
and@& sp,). Thespecies
belongingto the families of Cucurbitsceae, MaImceae, Chqmbaase,
and Amamthaceae
were resistant to infectionof
the c8usal agentof the
disease.The tobacco whitefly (Bemisia tabaci Gam) was definitely prwen to
k
an
effective vector of the causal agent of pepper yellow leaf curl disease. It was found that a single tobacco whitefly could@amnit
the virus to pepper plant.The
insect vtxtmcould b m m i t the g e m i n i d
in
a pers~stancemanner,
but it is not tmnmvariallytransmitted Acquisition
and inoculation
feeding
period totransmit
the virus was identified to be optlmum inthe
rangeof
3-
6 hours.The
virus needs at least 9 horn in the vector to complete a latent p e r i d The retention perid of the virus is until the insect he. The transmission of the virus will be more effectiveif
the acquisition mi inoculationf*
perid is longerand
thenumber
oftobruxo
whitefly is bigher.The female
of
tobcm whitefly is more effectivein
lnmmithngthe
virusc o m ~
tothe male.
mated
(P.
flor&iw) yieldedabout
220ug
ofpure
gemhivirus.
Etas&on
tbe
SDS-
PAGE analysis, molecular weightof
the-virus
watprotein
was about 29kDa
I-ELISA
and DIBA methods viere able to &tect thevirus
in
m p l e s . . Thereolctivity
of
antisenrm was
found similar
amongpepper
isolateshum
different
l m o m
(Segunun&Yo-
Cugemg
and h n b g )and those from
different hosts @epper, tab, tomatoand
Agemharr ampoi&) Antiserum codd also beused
for detectionof
P e p r p d I o w ZexfmrI vims j6romits
vector. A A e insect vector issufficient
for the detection of geminivirus pro^.^ I-ELISAand
DIBA method arev e r y u s e f u l a s t o o l s t o ~ t h e ~ ~ T h e m e t h o d s a r e v e r y e a v y t o b e ~ e d
out ,
fast
, andneed
only a minimum costfor
operation. Gemhiviruscould also
be
identified by western blott analysis.Pepper yefh~w leqf nal gemirrivims
could
be
deteckdby
using templates ofDNA
frominfected-
a s w d a s vectorsbyusing ~ ~ P C R m t h i with fourpairs
of primer, i.e : pALlv 2978 & pALlc 715, pALlc 1978 &pAEl
Ic 4%, pUPvl& pUPc2,and
p A v
494 &pAc
1048.Geminivirus
could
dm
be
Qetectedby
isolating the DNAof
the
infectedplant
using guanidine W i n e methods.
Two
spesific hqpmts,2600
bp a d
1600bp,
wasobserved
as thecirmrlar
and linear
of
DNA
(DNA confmmalion) of the the gerniuivirw.Dige&on using four
dction
enqmes (&zmH& EwRl, H i d &and
PHI)showed
that
the
DNA
of
pninivjrus kstdvary
in
theDNA
finger printing Although it stillneed
somemore &Cation,
the
PepprpIIow ZeqfwI geminhimis the gemhivirus
which
has amomprbte
gcmm
andmore
thanone
strain maybe
present in IndonesiaSURAT PERNYATAAN
Saya
menyatakan
dengan
sebenar-benarnya bahwa
segals p e r n y a a
dalarndisertasi saya yang be judul: -RISASI BIOLOGI, SEROLOGI
DAN
ANALISIS SIDIK JAIUDNA
VIRUS PENYEBAB PENY- DAUN KEFkITING KUNING CABAI, merupakan gagasan atau hasil penelitian disertasisaya sendiri, dengan pembibingm para Komisi
Pembimbing
kecwli yang den@ jelas ditunjukkan rujukamya Diserhsiini
belum pemah diajukan untuk memperolehgelar pada program sejenis
di
perguruan tinggi lainSernua data
dan
informasi yang digunakan telah dinyatakan secara jelas dan da pat di periksa kebenarannya.Bogor, April 2004
O\L%
LeKARAKTERlSASl
BIOLOGI,
SEROLOGl
DAN ANALlSlS SlDlK JAR1
DNA VIRUS PENYEBAB PENYAKIT DAUN
KERlTlNG
KUNlNG
CABAl
Oleh:
Sri
Sulandari
Disertasi
sebagai satah satu
syarat
untuk mempemleh gelar DoMorpada Progtam Studi Entornoiogi dan Fkpatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANlAN
BOGOR
Judul Disertasi
Nama NRP
Program Studi
: Karakterisasi Bidogi, Serologi dan
Anatiiis
Sidik
Jari DNA Virus Penyebab Penyakit Oaun Keriting Kuning Cabai:
Sri
Sulandari: 9850551 FtT
: Entomdogi dan Fiopatobgi
Men yetujui , I. Komisi Pembimbing
Prof,
Ir, Rusmi
tah Suseno. M. Sc.. P h.
D. KetuaIr. Sri Hendrastuti Hidavat. M,Sc,Ph.D
.
JumantoHar@sudarmo
AnssoW Anggoh
L 7 q m
Angg ota
Mengetahui,
2. Ketua Program Studi Entomologi
dan
FitopablqiRIWAYAT
E D U P
Penulis
dilahirkan
di
Yogyakarta pada tanggal 7 Mei 1958dari
ibu Suharminahdan
Bpk Suyatman Wiroatmodjo,dan
merupakananak
kelima
dari
sembilan
bersaudara.
Penulismenikah
dengan Dr.
Ir. Sudaryono, M.Eng. dandikaruniai sepasang putra-putri, yaitu Ninggar Seganten ( 12 th) dan Lintang Sagoro
(9 thl-
Pendidikan
dasar
sampai menengab diselesaikan di Yogyakarta,clan
pa& tahun1977
diterima sew mahasismdi
Fakultas Pertanian UGM, Yogydmta Tahun 1982 penulis rnempmleh gelar Sarjana Pertanian (Ir.),
sedangkan
gelar Sa jana Utama(SU)
diperoleh pada tahun 1986di
Jurusan Fitopatologi, Program Pascasarjana UGM.
Mulai tahun 1998 penulis mengikuti Program Doktor (S3) di Program Studi Entomologidan
Fitopatologi, Program PascawjamPB.
PMIKATA
Puji syukur
ke
hadapan AllahSWT
atasrakhmat
dm
kmuunia-Nya sehinga penelitian yang berjudul: Karakterisasi Biologi, Serologi dm Analisis Sidik JariDNA
Virus Penyebab Penyakit Keriting Kuning Cabai telah dapat terselesaikan.Penulis menyampaikan rasa terimakasih yang tak terhingga kepada Prof.
Ir.
Rusmilah Suseno,M.Sc.,
Ph.D.,Ir.
Sri Hen- Hidayat, M.Sc., Ph.D., Dr,Jurnanto Haqosudarmo dan Prof. Ir. Soemarton0 Sosromarsono, M-Sc., Ph.D , atas
segala kesabaran, bimbingan, pengkayaan wawasm, kn'tik, serta dukungan moil yang sangat besar peranannya dalam
terselesaikannya
penulisan disertasi ini.Peran para penguji yang telah memberikan banyak masukan sangat bermanfaat &lam diserfasi ini. Untuk itu penulis mengucapkan banyak terimakasih kepada
Dr.
Ir. Gede Suastika,MSc.
dm
Dr.
Lr. Ati Srie Wt,APU.Ucapan terimakasih juga disampaikan
kepada
Rektor dan Direktur Program Pascasajana IPB atas kesempatan yang diberikan kepdapwulis
untuk rnengiw program doktordi
IPB; Rektor Universitas GadjahMa&
ban D e b Fak. Pertanian UGM yang telah memberi ijin untuk mengikuti program d o h rdi
IPB; Kepda Balai Penelitian Bioteknologi Pertanim Palitbio) Bogor yang tehh mengijinkan penulis menggumkan fasilitas rumah kaca.Ucapan terimakasih juga penulis
mpaikan
kep& Departemen Pendidiksn Nasional melalui BPPS Dikti yang telah memberikan dukungandam
untuk meng kuti program doktor, Toray Foundation yang telahmem
biayai sebagian danauntuk penelitian, Ketua Laboratoriurn Virolog Twnbuhan IPB
dan
UGM
yang telah mengij inkan petlul is menggunakan fasilitas, peralatan dan bahan penelitian.Saya ucapkan banyak terimakasih
kepada
Prof.Dr.
Ir.
YB.
S m d i y o n o atas segala bantuannya, dan Prof.Dr.
Ir. Haryono Semangun ksertasemw
reRaMekan di minat studi Fitopatologi, Fak. Pert&UGM
atas dorongan dan dm reshmySL;Kepada mas Wawan, mbak
Ida,
mbak Woro, pak Edi, pak Sodiq,bu
Parmidi
laboratorium Virologi Tumbuhan, baik sewaktudi
kampus Baranangsiang maupun di Darmaga yang selalumembuat
ceriadan
semarak
suasana laboratorium sehingga menumbuhkan sernangat kembalidan
menghi langkan rasa putus asa saat penulis mengalami beberapa kali kegagalan&lam
percobaan. Atas ke jasama dan keakraban yang terbina, saya ucapkan ban yak terimakasi h.Rasa hormat dan bangga penulis sarnpaikan
kepada
kedua orang tua tercinta,i bu Suharminah W iroatmdjo (Alrn) yang telah mencurahkan kasi h sayang, bimbingan dan doanya serta keteladanannya dalarn mengasihi sesama, serta kepada yang tercinta Bapak Suyatman
W
iroatmodjo (Alrn) yang seIa1 u mendorong untuk rnenempuh pendidikan tertinggi dan selalumemberi
yang terbaik bagi putra-putrinya serta meneladani arti sebuah kepercayaan. Untuk semua itu penulis ucapkan banyak terimakasih. Semoga AllahSWT
memberi ampunan dan tempat yang terbaik di sisi- Nya sesuai amal ibadahnya. Amien.Kepada yang tercinta ibu mertua, ibu Subarti Suhartono
penulis
ucapkan terimakasih atas doa clankasih
saymgnya Kepada semuakakak-kakak
, adik-adik , ipar-ipar clan segenap keluarga besar bpWibu Wiroatrnodjo dan bpldibu Suhartono, atas doa dan kasih sayangnya, penulis ucapkan banyak terimakasih.Yang tersayang kedua anakky Ninggar Seganten dan Lintang Sagoro saya sam pai kan penghargaan yang tinggi atas doa, kasih sayang dan segenap pengorbanan
yang telah diberikan selama ini, dan karena tawa dan tangismu membuat jarak antara Yogakarta
-
Bogor terasa sangat dekat. Kepada yang tersayang suamiku Dr. Ir.Sudaryono, M.Eng., saya ucapkan terimakasih yang
tak
terhingga atas doa, kasih sayang dan kesetiaan, dorongan dan semangatdi
saat-saat sulit serta waktu untukNinggar dan Lintang diantara kesibukannya yang sangat padat.
Semoga hasil penelitian ini bermanfaat untuk kepentingan m a t manusia clan ilmu pengetahuan.
Y
ogyakarta-Bogor, April 2004xii
DAFTAR IS1
DAFTAR
TABEL
DAFTAR GAMI3ki-R
I1 TINJAUAN
PUSTAKA
Cabai (C 'apsicum spp.) Geminivirus
Identifikasi Virus
Keragaman Gerninivirus
Daftar Pustaka
I11
RESPONSBEBERAPA
KULTlVAR CABAI, TANAMAN LAIN DAN GULMA TERHADAP VIRUS PENYEBABPENYAKIT DAUN KElUTING KLJNING CABAI 24
Abstrak Abstract
Pendahuluan
Bahan dan Metode
Hasil Penelitian
dan
Pembahasan KesimpulanDaftar Pustaka
IV
KAJIAN PENCTLARAN VIRUS PENYEBAB PENYAKITDAUN KERITING
K W G
CABAI DENGANSERANGGA VEKTOR Bemisia tabaci
Genn.
(Hemiptera: Aleyrodidae)Abstrak Abstract Pendahuluan Tujuan Penelitian Bahan
dan
MetodeHasil Penelitian dan Pembahasan Kesimpulan
Daftar Pustaka
V PEMURNLPITU' VRUS PENYEBAB PENYAKIT
DAUN
KERITNG KUNMG CABAI
Abstrak Abstract Pendahuluan Tujuan Penelitian Bahan dm Metode
Hasil Penelitian clan Pembahassn Kesimpulan
xiv
VI
PEMBUATAN ANTISERUM DAN KkTIAN SEROLOGI VIRUS PENYEBAB PENYAKIT DAUN KERJTINGKUNING CABAI
Abstrak
Abstract Pendahuluan Tuj uan Peneli tian Bahan dan Metode
Hasil Penelitian dm Pembahasan Kesimpulan
Daftar Pustaka
VII DETEKSI DAN KAJIAN KERAGAMAN VIRUS
PENYEBAB PENYAKIT DAUN KElUTING KUNING CABAI MELALUI ANALISIS
SIDIX JAM DNA
Abstrak Abstract Pendahuluan Tujuan Penel I tian Bahan dan Metode
Hasil Penelitian dan Pembahasan Kesimpulan
Daftar Pustaka
IX
KESIMPULAN DAN SARANDAFTAR TABEL
Tabel 111.2
Tabel 111.3
Tabel IV. 1
Geminivirus yang telah dilaprkan menyerang
...
cabai (Capsicum spp)..
Macam dan fungsi gen kelompok geminivirus.. Hubungan geminivirus dengan serangga
...
vektornya, B. tabaciLuas serangan clan keparahan gejala geminivim pada berbagai jenis cabai di lapangan (tahun 200 1 ... -2003).
[image:16.611.88.523.67.784.2]Respons beberapa kultivar cabai terhadap infeksi geminivirus isolat cabai Segunung.
...
Hasil penularan geminivirus isolat cabai Segunung...
ke
berbagai jenis tanaman&n
tumbuhan liar.. Pengaruh j urnlah serangga terhadap efektivi taspenularan dan masa inkubasi geminivims isolat cabai setelah periode makan akuisisi selama 48
jam
dan periode inokulasi selama 24 jam.. ...Pengamh berbagai periode makan akuisisi
B. tubuci (10 ekor setiap tanaman) terhadap efektivitas penularan penyakit setelah 24 jam
...
periode inokulasi..
Pengaruh berbagai periode inokulasi R. fubctci (10
ekor setiap tanaman) terhadap efektivitas penularan penyakit setelah 48 jam periode akuisisi. ... Hasil pengujian periode laten geminivirus dalam ... tubuh B. tabaci..
DAFTAR
GAMBAR
Gambar I. 1
Gambar 1.2
Gambar 1.3 Gambar II. 1
Gambar fl.2
Gambar 111.1
Gambar 111. 2
Gambar 111.3
Garnbar 111.4
Gambar 111.5
Gambar 111.6
Garn bar 111.7
Gejala penydat daun keriting kuning pad. pertanaman cabai rawit
di
lapangan.....
Intensitas serangan penyaht dam keriting kuning pa& cabai rawitdm cabai
besar di...
lapangan.
Skema dan susunan genom geminivirus mono-
...
partit..Skema
dm
s u s u m genom gemhivirus biparht...
Gejala penyakit dam keriting kuning pada cabai
...
besar (A) dan cabai rawit (B) di lapangan. Variasi gejala penyakit keriting d u n kuning
...
cabai rawit..Perkembangan gejala penyakit daun keriting kuning pa& cabai rawit kultivar Cakra (hari setelah inokulasi)
...
Hasil penularan virus penyebab pen
yak]
t daun keriting kuning pada berbagai kultivar cabai...
Hasil inokulasi geminivints isolat cabai pada berbagai AJicotium spp....
Hasil inokulasi gerninivirus isolat cabai ke...
berbagai tanaman pertanian..
Hasil inokulasi geminivirus isolat cabai
ke
be- tanaman indikator, gulma dan...
Gambar
TV.
1Gam bar
W .
2Morfologi imago B. tabaci
dan
puprium-n
ya...
Gejda pada tanaman cabai (C. fiwctarcens) kultivar
C
h
yang diinokulasi geminivirus...
menggunakan B. fabaci
Garnbar
V.
1 Zona virus murni setelah ultrasenrifugasi...
dengan
Cs2$04..
Nilai absorbansi virus murni pada A 260 nrn =
1,83..
...
GambarV.2
Gambar V . 3 Hasil analisis protein selubung geminivirus isolat c a b i secara
secam
SDS-PAGE.....
Gambar VI. 1Gambar VI.2
Absorbansi antiserum tidak diserap..
...
Absorbansi antiserum y mg diserap mengym- kan sap perasan P. ~70ridam sehat
...
Abwrbansi antiserum setelah di presi pitasi rnenggunakansul
fat amonium...
Gambar VI.3
Gambar VI.4 Gambar VI.5
...
Titer antiserum..
Reaktivitas ant iserurn terhadap virus penyebab penyakit daun keriting kuning cabai dari berbagai lohsi (Segunung, Yogyakarta,
Cugenang dan Lembang) secara I-ELISA..
...
11 1Reak ivitas antiserum terhadap virus penyebab penyakit daun kuning keriting cabai dari berbagai hang (cabai, tembakau, tomat
dan
babadotan) secara I-EL1 SA ... Gam barVI.
6Gambar VI. 7 Hasil penguj ian I-ELISA dengan pgarnatan visual rnenggunakan antiserum yang tidak diserap pada geminivirus yang berasal dari
...
lokasi dan inang be*.
Hasil penguj
ian
I-ELISA dengan pengamatan visual rnenggunakan antiserum yang diserap. pa& geminivirus yang berasal dari lokasi clan...
Gambar VI.9
Gambar W. 10
Gambar Vi. 1 1
Garnbar VI. 12
Gambar VI. 13
Gambar VII. 1
Gambar VI1.2
Gambar VII.3
Gambar V11.4
Gambar V11.5
Gambar VII.6
Gambar W. 7
Gambar VII.8
Reaktivitas antiserum terhadap B. tabaci
...
virulifems secara I-ELISA
Reaktivitas antiserum terhadap virus penyebab penyakit daun keriting
kuning
dari berbagai lokasi (Segunung, Yogyakarta, Cugenang dan...
Lembang) secara DIBA..Reaktivitas antiserum terhadap vim penyebab pen yakit daun keri ting kuning dari berbagai inang (cabai, tembakay tomat clan babadotan)
...
secara DIBA..
Reaktivitas antiserum terhadap eksbak B. tabaci v i r u l i f m secara DIBA..
...
Hasil analisis secara western blotting protein selubung geminivirus....
Arah transkripsi pasangan primer yang digunakan untuk amplifikasi fmgmen DNA-A
dan
DNA-B...
N. benthumiuna yang diinokulasi virus penyebab penyakit
daun
keriting kuning ( 1 0...
hari setelah inokulasi)
Fragrnen
DNA
hasil amp1 ifi
kasi menggunakan primer pAv 494 &PAC 1048 (A) dan primer pALlv 1978 & pARlc 496 (B)......
Fragmen DNA hasi1 amplifikasi menggunakan primer pUv 1 & pUc2.. ...
Ampl
ifi
fi
kasi
fragmen DNA-A dan DNA-B nlenggunakan be- primer....
Pola pernotongan pita DNA geminivirus oteh...
berbagai enzim restriksi..
Fragmen DNA geminivirus hasil PCR dari
...
serangga vektor
Gambar W.9 Dendrogram hasil PCR-RFLP geminivirus isolat
cabai
dan
tomi....
152 GambarVII.
10
Runutan susunan DNA has11 PCR isolatSegunung-2 menggunakan pasangan primer pALv 1 978 (plus sense) dm pARc 7 15 (minus
.sense) ... 153
Garnbar VII. 1 1 Runutan susunan
DNA
hasil PCRisolat
Y
ogyakarta menggunakan pasangan primer p ALv 1 978 (plus sense) clan pARc 7 15 (minusI. PENDAHULUAN
Latar Belakslng
Tanaman cabai (Capsicum spp.) merupakan salah satu jenis tanaman
sayuran yang prospeknya sangat bai k untuk di kembangkan sebagai tanaman utama karena mempunyai nilai ekonomis tinggi. Buah cabai bermanfaat antara
lain sebagm penyedap masakan, penambah selera makan
dm
mengandung berbagai vitamin. Kandmgan vitamin A dan C pada buah cabai sangat tinggi. Pada setiap 100 gram buah cabai segar mengandung vitamin A sebesar 470 TUdan 18 mg vitamin C pada cabai merah atau wbai besar, mhngkm pada cabai
rawit mengandung 11.050
IU
vitamin A clan 70 mg vitamin C (Dir. Gizi. Depkes RI. 1 989). Peranan cabai dalam rumah tangga setam dengan beras yang sudah menjadi kebutuhan pokok sehari-hmi dan posisinya tidak dapat dgantikan dengan komoditas lain. Rata-rata konsumsi cabai di Indonesia seJritar 2,4 kg/oran9, tahun(VOS 1 994).
Dari tahun ke tahun, luas pertanaman cabai di Indonesia meningkat
pesat sejalan dengan perkernbangan agroindusbi. Pada tahun 2000 Iuas
pertanaman cabai mencapai 174.708 ha atau sekitar 20,39?h dari total areal
pertanaman sayuran (Ditjen Bina Produksi Hortikultura 200 1 ). Walaupun luas areal pertanaman cabai smakin meningkat , akan tetapi sering terjadi fluhasi harga maupun produksinya. Faktcw yang menyebabkan produksi turun dm h g a
menyerang cabai
dm
sangat merugikan. Wah satuvirus
penyebab penyalut cabai di Indonesia adalah geminivirus (Hidayat et al. 1999),Mdai tahun 2000 sampai sekmng terjadi epidemi penyalut daun keriting kuning pada pertanaman cabai di pulau Jawa (Gambar I. 1). Epidemi penyalut terjadi apabila teqsdi peningkatan populasi ttinaman sakit Marn suatu areal dalam waktu yang singkat dm sangat merughn (Agrios 2000). Epidemi
penyakit daun keriting kuning cabtii terjadi di Indonesia berdasarkan pada
kejadian p e n w t clan intensitasnya di lapangan sem- meaingkat dm menjadi masalah besar k m m menimbulkan baayak kerugian pada pertmaman cabai di
berbagai lokasi.
b i l p e n p a t a n di beberap senha produksi cabai di
DIY,
Jawa Tengah dan Jawa Barat menmjukkan, bahwa kejadian penyakit pa& cabai rawit mencapai 70 - 100% dan tersebar lms; sedangkan pada cabai besar terdapat sebaran secara sporadik dengan kejadian peny-alut berkisar antara 10 - 35%.Namun, kejadian penyakit pada cabai besar ku1tivar TM 999 di daerah Kopeng (Jateng) dm Kulon Progo
(DIY)
pada t&un 2001-
2002, dan di Sieman padatahun 2003 rnencapai 70
-
IGO% (Gambar 1.2). Gejala penyakit yang miripdengan penyaki t daun keribng kuning cabai yang banyak ditemukan akhir-akhir
Berdasarkan gejalmya yang mirip dengan -&an
geminivinrs,
maka identi-i d i l W a n meMui pengujim sifht-sifat suatuvirus,
pituki-
h g , cara penularan men- m g g a vektm, pernumian virus dan kajian smlogi serta andisis sidik jari DNA.Alur
penelitian disajikan pada Gambar L3.Tnjuan Penelltian
Penelitian ini bertujuan untuk mengehhui status penyalat di lapangan dm identifikasi virus penyebab penyakit rnelalui pengujisn sifat-sifat biologi, mlogi dan pola sidik jari
DNA
m b a b penyakit daun keriting kuuing cahai.Penelitian dilakukan melalui s w e i di lapanm di rumah kaca dm labmtorium. Penelitian di lapangan untuk men@ahui kejadian penyalut dotn k e p d a n gejala. Penelitian di rumah
kaca
dan laboraton'um yaog dilakukan terdiri atas:1. Kajian biologi meliputi: a) kisarm inang dan b) hubmgan virus penyebab penyakit dengm serangga vektornya
2. Pembuatan a n t i b d p o l i k l d
dm kajian
serdoginya.3. Difwensiasi geminivirus melalui pola idik jari DNA berdasarkan poIymemre chain reaction - resrricrion fragment length polymorphism ( P C R - R E . .
4. Analisis hubungan kekerabatan antar isulat geminivirus meldui pembandingan runutan susunan DNA
Hijmttsis
1. Penyebab penyakit daun keriting kuning cabai adalah geminivirus, clan mempunyai kismn h gyang luas serta ditularkm oleb m g g a vektor. 2.
Di
antara jenis tamman yang digunakan pada pengujim kisaran inangakan
3. Anfibodi pdiklmal terhdap vinrr penyeQab penyakit dron keriting Luning yang diproduksi dapat d i wsewalat deteksi ysng cepi dan alnuat
4. Analisis P C R - W ,
dam
menlmjukkan keragaman antar isolat geminivirusyang menyerang cabai.
5. Melahi perunutan sunman DNA basil PCR geminiviruP isdrt cabai di Indonesia, hubmgan kekerab- dekat d e n w Tmato yellow Ieqfcurl v i m dari
benua
lama.'Jabar
westemblott
s8tu isolat y~ bsttmda pol8
piIa DNAn ya dg isolal
Segunung I
Perunutan smunen DNA
w
Pemnutan susunan DNA
\
Analisis
~ ~ u n a " O N *
..,*....
*.,....*..
hubunganI
dari Gene Bank kekerabatan
IL
TINJAUAN
PUSTAKA
C a w (Capsicum spp)
Di Indonesia cabai rnempakm tanaman dataran rendah maupun tin@ dan menrpakan kamoditas pilihan untuk suatu usaha tani. Berdasarkan data statistik diketahui bahwa luas
d
tsnmsn cabai dmi tahun 1989 sampai 2000 selalu menductuki posisi tertinggi di antara tanamansayuran
yang diusabkau di Indonesia @It. Bigram. Ditjen. TPH 1983, 2001). Buah cabai banyak dimanfaadcan untuk berbagai keperluan misalnya sebagai penyedap clan penambah selmmaan.
Di
Indonesia terdapat dua jenis cabai yang banjrclk dihmm yaitu abai rawit (Capsicum frtrlescensL.)
dan cabai besar (Capsicum a n m m L.), serta masing-masing mempunyai kultivar yang banyak. Produksi cabai di Indonesia tidak stabil dan mengdarni fluktuasi barga yang sangat tajam. Salah satu faktor penyebab tmnnya produksi c&ai adalah serangan patogen. Terdapat banyak patogen yang menyerang pertanaman cabai di l a p g a n , antarszlain jamur, Wteri, nematodrt dan virus. Di antam patogen-patogen tersebut yang sering ditemukan pada pertanman cabai adalab virus, dm patogen ini sering menimbui kan kerugian besar.
Virus yang menyerang cabai antara lain Cucumber mosaic virus
(CMV),
Tobacco mosaic virus (TMV), Potato v i mY
(PVY), Tobacco etch virus (TEV),Tobacco rattle v i m (TRV), Tobacco ringspi v i m (TRSV) ,Potato v i m X (PVX)
,Alfca
m u i c v i m(AMV),
Chili vein mottle v i m (CVMV), virusgemhivirus (Hadayat et al. 1999, Sulandari et a/. 2001). Geminivirw p d a cabai di Indonesia masih belum banyak &&ti, s a h & n di luau negeri pnelitian tentang penyakit
ini
sudah dildmkm sews intensif.GeminIn'nis
AFtI
e h o m iGemhivirus rnerupakan salah wtu kelompok vim yang banyak menimbulkan kerusakan pada berhqy tanamsn yang dibudidayakan di dmah
tropik m a u p subtropik (Polston & Andeman 1997,
Brown
1987). Selorin menyerang babajpi tanaman-an,
geminivirus jugt dapt mmghfkksi berbagai gulma (Rojas el d. 1993, Roye et ul. 1997, Mati et al. 2002).Pada tahun
1480 dilaporkanbshwa
gemhivirus m q g p a m produksi tamat di Florida, Karibia, Mebiko, Amerika Teagah, VenemeIa, Brasilia dan Jepang (Polston & Anderson 1 997, 1 999, Idris & Brown 1 998, Kato ei a). 1998).Total kerugian yang ditimbulkan &bat m g a u gemhivirus pada tomat
di
Republik Dominika pads tahun 1989 - 1995 &tar 50 juta S
US
(cit Poistan & Anderson 1977),dm di
Florida pada tahun 1990 - 1991 total kerugimya rnencapai 140 juta $US
( c i ~ Polston el 01. 1W3). Salah satu jenisnya, yaituCassava mosaic v i m &pat rnenimbulkan banyak kerugian pada ketela pohon di Kenya (Bmk 1983). Geminivirus j u p &laporkan menyerang k a m g buncis di Amerika Serikat dan menimbulkm banyak kaugian (H~dayat et
d.
1-31.dilaporkan menyerang tanman cstbai dan h y a k menimbdkan kerugian di
Am& Serikat, M W o , Cuba, Guatemala dan Thailand. Berbagai geminivirus
yang menyerang cabai dapat dilihat @a Tabel 11. 1.
Tam 11.1. Germnr\-irus yang tebh d i l a p o h menyefang cahi (Capinun spp.)
virus
Pepper husteco v i m (PHV)
Serrano goiden
masaic virus
(Sew)
T e r n p w r geminivims (TPV)
Peppr jalaptw
vim (PN)
SimIoa r m f o laaf
curl virus (STLCV)
Tomato yeIIow leaf curl nrus (TYLCV)
-daun nulggdlm&
penebah tdang
dauq kerupuk, d m menguning dan kerdil -dam tnengmmg dan rnenggulung ke at=
Negara
Meksiko
Meksiko dm
Arnenka Senkat Texas. Costa Rica Mekslko dan Amerika Senkat Thailand, Cuba Meksiko Sumber Torres-Patcheco e: al.
19%.
Brown & Podos 1990.
Stenger el al. 1990;
L o t W er al. 2000
Tom-Pacheco ei 01. 1996.
Idris & Brown 1998.
Chiernsodat & Kittipakom 1 997; Samretwanich
er a1.2000; Quinones er al.
2002
Torres-Pdew er al. 1996.
Hidqat ef al. 19W
Morfologi dan t a b o m i gemiaivirus
Gerni ni virus merupakan kelompok virus yang menipunyai genom DNA
M v i r u s termad ke
ddam
h i l i -viridae. E l d a m h struktrrr genom, jenis serangp vektmyadm
jenis tansrman inangap familiini dibedakan m e n j d e m p t genus yaitu: Mmtrevirur. CWOY~F~LS, B e g ~ m ~ ~ i l ~ r s dan Toponrvim (Van Regemnortel 2000, Fauquet sf d. 2000, cif Hull 2002).
Genus Mustminu, anggota jenisnya dicirikour den- tipe Maize streak
virus, mempunyai gemm monopartit yang bendman sekitar 2,6
-
2,skb,
menyerang tanaman monokotil clan ditularkm oleh wereng dam. Genus Curtminu anggota jenisnya dicirikntn den- tip Beet curly top v i m . Pada Currwirrrs, struktur genom dan jenis m g g a vektornya m a d e n p genus Mmtmvims, akau tetapi hstnya menyerang tanaman dikotil. Gaus B e g o m w i ~ ~ ~ atau sebelumnya d i k e d sebagai subkelompokiII
anggotmya dicirikan dengan tipe Bean gd&n mosaic v i m , dengan s&&w genom bipartii(be-
monoparti t), inangnya adalah tanman dikotil, vektornya Bemisia t a h i Gem.T o p m n i l u ~ merupakan genus baru h i 1 paisahan
dari
Curtovirt~~. Genus ini mempunyai struktur genom yang mirip dengm Cwtovinrr &an tetapi ditulaba oleh treehppr yaitu Micnridis malle Ifera.Begornovim sebagian besar genomnya terdin atas dua mdekul (bipartit) yang masing-masing berukuran sekitar 2,78 kb dan terpisah menjadi dua, DNA-
A dan DNA-B. Selain itu ada yang hanya terdiri atas satu molekul saja atau
monopartit yang berukuran
sekitar
2,7-
2,8 kb. Geminiviruis dengan genombiparht banyak ditemukan di benus baru (Amerika), sdmgkan yang monowt bmyak terdapat it dmia lama yaitu meliputi Eurasia, Ahka, dm Australia ( cif Hull 2002). Geminivinrs dengan mom rnmrn-t pada u m q a h y a
Geminivirus p g manopartit dengan yang bipwtit keduanya mempunyai uhran, fungsi dan mganisasi gm yang hampir saraa khususnya DNA-A. Adapun maam gen dan fungsinya dapat d i l h t p d a Tabel 11. 2.
Gen penymdi protein selubung virus (coat protein) merupakm daerah
genom yang mempunyai runutan susunsn DNA den- demjat kesamw yang tinggi (conserved) mtar anggota geminivirus d d m satu p u s {Rojas ei d. 1993,
W p t t & Brown 1%). Intergenic region
(IR)
sltau common region(CR)
,
selain mempunyai demjat kwamaan mutan asam nukleat ysng tinggi juga mempunyai derajat kesamaan yang rendah (drversed) antar anggota genus g d v i r u s( M e w s 1992, Padidam et al. 1995 cit,
Brcwn
1997, Van RegemnmteI el al.Tabd 11.2. Macm dm fimgsi gen Lsbompok g - d
Jenisgen pada gerrinivirus Fungsinya
No Monopartit B p a m
1 V l (CP) AV1 (CP) p n h r & n p r o h d u h g vim,
pdaran rml8lui semgga vektor dan
pergerakan v i w di dalm inaagnya
2 C1 (Rep} ACl (Rep) repWcasivirus
3 C2 (TrAP) AC2 (TrAP) p m b e n t w Tramucfiwring prorein
P A P I
4 C3 (Ren) AC3 @en} parlbeatukm protein replimlion embmw
(.En)
5 C4 MP (BV 1 clan ekspresi geJala
BVl)
6 MpIV2) NSP(BVI) dm p e r g e virus mmmm yang
MP (BCI) t&d;si Sumber : Van Regenmortel er at. 2000, Haul 2002.
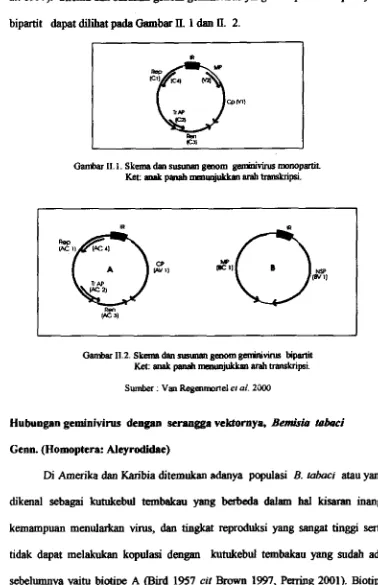
a&. 1997). Skema dan sammm p a n gemhivirus yrtog monapht m a p ymg bipartit &pat dilihat pada Gambar II. 1 dan
II.
2.Gambar Ii. l . Skeara dm susunan genom geminivitus mmopmht.
Ket: anak panah ma* arah -psi.
Gambar 11.2. Skenra dan srrnman geaom g m h i ~ i m bipartit
W: a d panah nmunpkkm arah m k r i p s i .
Sumber : Van ECegenmonel el af. 2XK3
Hubungan geminivirus dengan sersngga vclrtornya, Be&& robaci
Genn. (Hornoptera: Aleyrddae)
Di
Amerika dan Karibia ditemukrtn adanya populasi B. iabaci atau yangp g menyebutnya sebagai @es B. argt!nt$dii. Selain biotipe
B,
tadspat
tujuh kelompok biotipe B. t-
,
dan biotipe I3 t d u t sangat potensial dalam menularkan geminivinzs porda berbagai tamman budidaya (Paring 2001).Sikh hidup kutukebul tern- di daerah t q i k jmda kundisi optimum
sekitar 3 minggu. kmgga M h a dewma mampu W u r sebanyak 200 butir. Perkembangan dari telur mpai dewasa melewd empat fa& yaitu telur, nim& (tip instar), pupa clan imago (Motmd 1993). M e n W Badri (1983), sdclus hidup kutukebultembakau pPrda
~~
plasuhu28-30Csekitar 17hari.Masa inkubasi telur &ah (5,78
+
021) hari, instar pertams lmanya (3,14 2024)h i ,
nimh instarkedua
(3212
0,16 ) han,instar
ketiga (3,14 1 q16) hari,danpupa nrtrrlrrh(2,Sl ?0,21)bari. Tubuhkutukebd d e w a s a h a r n a kuning dengan serbuk putih @absdlan
dan
sppnp.Semngga
tersebut sangntpolifag, imgnya sangat banyak yaitu b glebih rnencapai 300 spesies dari
63 fmiii.
Peranan kutukebul ternbhu sebagai vektor geminivirus yang
menyebabkan penyakit sudah banyak
~~
(Tabel 11. 3):Pada
umumnya hubungan virus dengan vektomya h i f i t persisten &an tetapi tidak diturunkan ke generasi berikutnya melalui telurnya ( m n tmnrovarial trummission). , Walaupun dernikian a& pula geminivirus yang dapat diturunkan ke generasibmhtnya, m i h y a TYLCV dari berms
lama (Cmmek
et al. 2001) clanT
W-No Vuud inangnya Periode Periode Periode Sumber
akuisisi laten (jam) retensi
1 +as l e a f w l virus
pada d m
2 Texaspepper
geminivirsls pada cab J
3 Tomato pI1w leof ctirl
virtls pada t o m i
4 Pepper yellow leaf cur2
virus pada calm
5 Texas pepper
gminivirrrs pada adxu
24 9 Idris & Brown
1998.
0,25 - 0.5 2 1 - 2 4 1 0 - 2 0 NaWlla&Maxwell
1993.
1 10
-
Smetwaruchet al. 2000. 10 Stmger el al.
1 m. 6 Tobacco Ieafcurl virus O,5 S m p i Noodawati
pads t d u mati etn1.2002
Keterangan:
-
: tidak ada dataldentifikasi Virus
Deteksi agens penyebab penyakit pada tanaman, benih ataupun bahan vegetatif selain untuk mengetahui sifat biologinya juga merupakan langkah awal yang sangat diperlukan dalarn strategi pengelolaan penyakit. Banyak cara yang
dapat dilakukan untuk deteksi dan identifikasi virus. Identifikasi virus antara lain
dapat dilakukan melalui identifikasi secm bi ologi ataupun nonbiologi serta
meialui analisis komponen patogennya (Brown & Ogle 1 997). Identi
fi
kasi secarabiologi a n m lain dilakukan dengan penularan ke tanaman indikator, penularan
m e n g g u n h serangga vektor dan kajian kisaran inang. Untuk mengetahui
adanya komponen spesiiik dari suatu virus clapat dilakukan dengan identi fikasi
secara non biologi an tara lain dengan pengamatan menggunakan mi kroskop elehon, uji irndogi atau serologi, hibridisasi asam nukleat rnenggmdcan
Identiiibi gemhivhs scmra biohgi
Identiaasi vinzs s e a m bid& d q a t ctilakukaa melalui uji kisrtran inmg dan atau melalui uji hubungan virus dan smmgga vektomya.
Cara
tersebuttelab ddakukan untuk mendeteksi
dan
rnengidenti- Bean IhuM mosoicgernirrivim (BPMV) (Morales et d. 1990), Texas p p r gemjnivirur (Stenper et al. 1990), Simlw tomato leqf curl geminivirus (STLCV) (Idris & Brown 1598).
IdenliAkasi geminivlrPs mtlalni uji m l o g i
Serodiagnosis merupakan cara deteksi virus den= m d t k a n reaksi antam antigen dan antibaii (Trrrrmce 1992). Cam ini rnempuyai banyak keuntungan antara lain
cepat,
tqmtdm
dapat d i mmtuk karakterisasi virus serta menptahui hubungau k e k m h m suatu virus ( Van R e g m u d 1992).Serodiagnosis telah banyak digunakm mtuk mengidentifikasi berbagai virus yang menyemag tanaman maupun mdahri serangga vektaanya mars kualitatif ataupun h t j t a t i f . Salab satu bhm dasar yang digunakau untuk pengujian secara serologi adalah tersedianya antibdi. Sebagai laagkah awal mtuk membuat a n t i M adalah dengan melakukan isolasi virus dari inangnya dengan
cam pemurnian virus. Metode pernumian virus sangat bervariasi tergxrntung dari sifat dm jenis virusnya (Matthews 1992). Bebetap anggda geminivirus yang
telah Mail dimurnikan misalnya TPV, dan Bean gd&n masaic geminivims (BGMV) menggunakan gradien sul fat sesium (Stager ef ai. 1990; Modes er
d.
1990). Tomato goI&n yeIlaw mosaic v i m dirn w hleqfcwl v i m (SLCV) rnenggmakm gradien &osa 1W% (Hamilton et al. 1981, Cohen ef
d.
dim&
secam
m v e m PEG yang d i l m ~ den- graden sukrosa (Honda erd.
1983). Tridowati (1989) b g a n cam pemutaian sebagian (pamal puri$cation berhasil memurnikan virus kenrpuk yang mmyemg temtdauVirus yang telah diinumikan sebelum d i w
untuk
imunisasi pada hewan percobam perlu dikarakterisasi mtuk melihat tingkat kemurniannya. Kmlderisasi clapat dil* dengan meagmati bentuk danukuran
p d e l menggunakan mihodcop elehrm, nilai a b s m h s ' menggunakan spekmfotamem (Van Regemnod 1981), atau melibat h a t molehd protein selubung virus seam elhfbresis (Sanbrmk el d. 1989). Virus mumi padaumumnya menrpakan salah sstu antigm yang sanpt baik mtuk menj@dkm antibodi pnda hewanpembmi.
Antibodi menrpakan imunoglobulin yang dihilkan oleh sel limfosit
B
pada hewan percobaan sebagai tanggapan adsoya rangsangan dmi swtu antigen (Memaugh ei 01. 1990). B d m a h a jlrmlah epitop pengimbas kekebalan , antibodi dibedakan menjadi poliklmal dan mmddonal.Antibodi
poliklonal dibentuk dari sejumlah klon lim6bsitB
yrmg dirangsang deh banyak epitop daFiantigen tertentu sehingga rnempunyai variabilitasnya sangat tmggi, khususnya &tam ha1 titer, klas antibodi dan spesifitamp. Hal ysng sebaliknya terdapst p d a antibodi monoklonal. Antibodi poliklonal lebih mudah pembuatamya dan biayanya lebih murah dibmcbg antibodi rnmddoplal. Spesifibmya a n t i M poliklonal akan tiaggi apbila antigen yang digunakan b e n d dari
v h s
mumi. Antibodi polikiod banyak digunakanmtuk
identilibxi suatu v h s WunBanyak uji serologi yang berkembang pesat dan has dewam ini baik untuk
deteksi maupun karakterisasi suatu virus. M d e serologi kmebut antara lain uji presipitasi dalam tabung, agglutinasi kloroplas,
difusi
p d a dalam agar, dtrn uji flokulasi lateks (Ball ef al. 1990). Untuk melihat kekerahtaa suatu virus banyak digunakan pengujim difusi p d a dalam agar yituden=
menmati pitapresipitat yang terbentuk
.
Metode presipitasi clan agglutinasi klwoplas walaupun sangat mudah dilstkukan akan tetapi h e n a terlalu b y a k menggunakan antibodi saatini
jarang digunakaaDi
antara carst serodiagnosisyang paling banyak digunakan saat ini adalab uji w elinked immmorber?t
assay (ELISA) meliputi &Ie anif- sarodwich ELJSA (DAS-ELISA) (Clatck & Adams 1977) dao i d m c t
ELIS4
(I-ELISA) (Koenig 1981). Selain ELlSA menggunakan lempeng m i k d t e rdari
plystyrene juga dapat digrmakan membran nitroselulosa ymg dikenal dengan teknik dor immunubiding a s s q (DIBA). Teknik ini banyak digunakan lebih mudah dm hernat karena r e a pyang d i g u n h lebih sedikit (Bmttari & Gocdwin 1985).
Identi fikasi geminivirus seco~a serologi masih jarang dilakukan dan masib
terbatas hanya pada beberapa geminivirus saja karena adanya beberapa kendala. Kendala tersebut antara lain konsentmsi virus dalam tumbuhan inang sangat
rendah, w i n g teqadi reaksi silang dan kesulitan dalam penyediaan antibodi ( crt Rojas et al. 1993, Van Regumortel2000).
Identifikasi geminivirus melalui a d b b sidik jari DNA
Dewasa hi untuk kmkterisasi maupun detelcsi geminivirus tumbuhao banyak dikernbaagkan teknik molekder. Teknik FCR akhk-akhir ini berkembang
&tin memberikan hasil p g akurat, cepat dan sangat peka. Teknik PCR b p
memerlukan jumlah sampel sedikit,
dan
sarnpel &pat bentpa bahan sew, sudah dikeringkan atau beku (Rojas et al. 1993, Wyatt & Brown 1996). Teknik PCRclapat mengatmi kendda pada penguj ian gemhivirus sema serologi.
Pengujian dengan
teknik
PCR memerlukan sepmang primer yangsfKsia
yang akan menginduksi pembentukau dan perban* rangha asam nukleat
atau mtai DNA tertentu dengan btmtuan enzim Tag plymerase ddm mesin PCR atau tlaermaycIer. PPemilihan primer pug tepat sangat menentukan kebdasilan identiflkasi suatu virus. Pada geminivirus , primer &pat dipilih pa& bagian common mgion atau intergenic region, gen yang menpdikan protein selubung virus, maupm protein yang berasosiasi dm- proses replikasi virus (Rojas et a!. 1993, Wyatt & Brown 19%, Pdstoll& Anderson 1W). Untuk keperluan taksonomi geminivims banyak digunakan primer dari gen penyandi protein selubung yaitu AVl ORF DNA-A karma daerah ini mempunyai derajat kesamaan yang sangat tin& (conserve4 antar anggota genus geminivirus (Wyatt & Brown 1946, Idris & Brown 1 998). Penyusunan pnmer tersebut misalcya Av
494 & Ac 1048 yang akan mengamplifikasi sebagan gen protein selubung virus berukuran sekitar 550 bp. Berdasarkan ukuran fragmen DNA tersebut dapat hgunakan untuk rnengidenti fikasi keberadaan geminivirus pada berbagai
tanaman wan,tanaman hias dan gulma (Wyatt & Brown 1996). Penggunaan primer yang disusun dari daerah ALl dan ARl yaitu PAL1 v 1978 dan PAR 1 c
tanaman, juga dam rnenmuikebedam gemhivirus dm l o h i virusnya di dab tubuh m g g a vektomya (Mehta et ai. 1%).
Identi-I gemhivirus melalui analisis sidik jari DNA dapat juga dengan cara mengisolasi DNA replicm'ive fom secara langmng rnenggmkan metode guanidin alkalin, dm ha1
ini
telah b h i l dilakukan untuk WPsear &aMv i m (Bendahmane et 01. 1995).
Kemgaman geminhim
Begmovirus merupakan sal& satu genus geminivirus ymg mempunyai
anggota yang -&at banyak.
Di
lapangan &bat pendaran geminivirus oleh=raw@ v e m a ,
w
inag Y m g ataupun Y a w .-
kekmMmnya, dapat m t M oleh sstu jenis ganinivirus sema tunggal, bersamaan dengan jenis geminivirus lain ataupun oleh strain lain Oleh karena adanya infeksi geminivirus seam bersamaan pada inang yang m a maka memungkinkan timbdnya 6 baru ~ h i n g g s teajadi keragaman geminivirus.
Adanya strain geminivirus dapat didetehi melalui pengujiau kisaran inang, uji
serologi maupun den= analisis sidik jari DNA
Mdalui uji kiwan hang adanya strain geminivirus &pat diketahui oleh adany a perbedaan respons pa& tanaman, misalnya p d a buncis y ang terinfeksi TYLCV-Is dapat menirnbulkm bean crumple disease sedangkan TYEV-Sar tidak dapat menimbulkan gejala peuyaktt tersebut (Naws€astillo et al. 1W). Menggunakan antibodi monoklwal dapat diketahui adanya
strain-
strain chi(Rojas et al. 1993). Meldui tekoik t d u t juga dam ctiketahui kera- geminivirus ymg menyerang mbai di W m i a (Hidayat et d. 1999).
Untuk melihat kmgaman sustu imlat virus mu timbulnya suatu strain
virus &pat juga dilakukan d i s i s berdawkm mutan suz~man asam nukleahya, mi* P e p r jdopno v i m merupakan strain Pepper hnusleco
v i m (PHV), dan Chino cdel t m t o v i m merupakau strain Tomato Ieqfcnmple
v i m ( T o m ) (Tofm-Pacheco el al. 1%). Menggunakan cam ini adanya
berbaM strain gemhivirus juga d i t e m h
p d a
Tomdo yellow leqfcw3 v i m (TYLCV) (NakMa & Maxwell 1983, Navas-Castilio el 1999, Polston & Mc Govern 1999, Chatcbawankanphmicb & Maxwell 2002X Texas p e p r v i w (TPV) (W eld.
2000), dan ACMV (Bock 1983, cir. Brows 1997).DAFTAR PUSTAKA
Attathorn, S., Chiemsombat P, Sutabutra
T,
Pongpani tanond R 1990.Characterizatmn of nucleic acid of T m t o pIlow let# clrrl v i m . Kaselsart J flat. Sci St&) (24) : 1 -5.
Badri. I
B.
1983. IrdentrJicofion ofthe A l e y r d i h e on sqbemjkrn two location In West Java md some biormomics of Bemisia tabaci Gem merniptem: A l e y m d h ) on t h e sgybem varieties. Biotrop. SEAMEO, RegionalCenter for Tropical Bioloa Indonesia. 62 p.
Ball, E M, Hampton RO, Boer
SH
de, SchaadNW.
1990. PolycIonalan ti bodies. In: Serological methods. Hmpton, RO, Bat1
EM,
Boer SH de (4s). APS Press Minnewta 389 p.Banttari
EE,
G d w i nPH.
1985. Detmon of Potato virus S, X andY
byenyme-linked i m m m sorlmt assay on nitrocellulose membranes (Dot ELISA). Ann PhytopathoI Soc J a p 69: 202 -205.
Bendahmane M, Schalk
W ,
GronenbmB.
1995. Identification andc h a r a c t a m of Wheai &a# v i m h m France using rapid method for gerninivirus
DNA
preparation. Pbtopxthd 85: 1449-
1455.Bosco
D, Mason G, AccottoGP.
2001. Investigations on lmmmrial trans-mission of TYLCV-Sar by Bemisia ? h i (Hemiptera: Aieymhdae). Europem Khitejly 5)lnp (Sicilia, I*), 27 th Feb - 3 rd March 2001.
Brown
JF,
OgleHI.
1997. P h i Pathogem a d Plmrt Disemes. Australia: Rockvale Publ.Brown JK. 1997. The Biology ond MolecuIar Epidemidagy of Geminivims Subgroup Ill. dalam: Stacey G, Keen
NT,
editar. Plant-Miaobe Interactions Vol 2.
New
York: Interuatianal Thomson Publishing. Hlm125 - 195.
&own JK, Poulos
BT.
1990. S e m pI&n mosaic v i m : A new whitefly transmitted geminivirus of pepper and tomato in U.S.
Plmt Dis 74:720.Chatchawankmphmich 0, Maxwell DP. 2002. Tomato ledcurl k a m a t h v i m fiom Bangalore, hdia, appars to be a rekombinmt begornovirus. P h y f ~ p ~ h d 92 : 637
-
645.Chiemsombat P, Kittipakm K
.
1997. Conhnaticm of potentially impportant pepper viruses. Proceeding of A W E T-II Final Workshop, Bangkok, Thailand 1 - 6 September 19%. AVRDC.Clarck MF, Adams AN. 1977. Characteristics of mimoplate methods of enzyme linked i m m u n e t assay fadetection
of
p l a t v i m J Gen v i r d 34: 475 - 483.Cohen S, D u f h
JE,
hrsenRC, Liu HY
,
Flwk RG