KELIMPAHAN DAN POTENSI CENDAWAN ENDOFIT
UNTUK MENEKAN PENYAKIT KUNING PADA TANAMAN
CABAI (
Capsicum annuum
L
.
)
DAMAYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Kelimpahan dan
Potensi Cendawan Endofit untuk Menekan Penyakit Kuning pada Tanaman
Cabai (Capsicum annuum L.) adalah benar karya saya dengan arahan dari
komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2013
Damayanti
RINGKASAN
DAMAYANTI. Kelimpahan dan Potensi Cendawan Endofit untuk Menekan Penyakit Kuning pada Tanaman Cabai (Capsicum annuum L.) . Dibimbing oleh SRI HENDRASTUTI HIDAYAT dan SURYO WIYONO.
Penyakit kuning yang disebabkan oleh Pepper yellow leaf curl begomovirus (PepYLCV), telah menjadi ancaman utama di daerah penanaman cabai, terutama di Jawa dan Sumatera sejak awal tahun 2000. Gejala penyakit di lapangan sangat mudah dikenali yaitu daun berwarna kuning cerah dan keriting, tanaman menjadi kerdil. Kejadian penyakit di lapangan dilaporkan mencapai 20% sampai 100% dan upaya pengendalian penyakit yang telah dilakukan belum berhasil menekan penyakit secara nyata.
Penelitian dilakukan dengan tujuan untuk mempelajari kelimpahan cendawan endofit dari daerah endemis penyakit kuning cabai di Yogyakarta dan mengevaluasi potensinya dalam menekan penyakit kuning cabai. Penelitian terdiri dari tiga kegiatan yaitu (1) Isolasi cendawan endofit yang diambil dari tanaman cabai di daerah endemis di Sleman, Yogyakarta, sampel terdiri dari daun yang bergejala yaitu daun keriting dan kuning dan daun yang tidak bergejala; (2) Uji patogenisitas menggunakan benih cabai TM 888; dan (3) Uji keefektivan cendawan endofit terpilih untuk menekan penyakit kuning dilakukan di rumah kaca.
Hasil penelitian menunjukkan bahwa cendawan endofit yang diperoleh dari tanaman cabai didaerah endemis penyakit daun keriting kuning cabai di Yogyakarta memiliki kelimpahan dan keragaman yang tinggi. Tanaman yang tidak menunjukkan gejala infeksi virus memiliki keragaman dan kelimpahan cendawan endofit yang lebih tinggi dibandingkan pada tanaman yang menunjukkan gejala. Spesies cendawan endofit yang diidentifikasi dari tanaman tidak bergejala berjumlah 19, sedangkan yang berasal dari tanaman yang bergejala berjumlah 13. Komunitas spesies cendawan endofit diantara keduanya berbeda, hal tersebut didukung dengan rendahnya indeks sorensen, SC: 0,5625 yang artinya kemiripan komunitas spesies cendawan endofit diantara keduanya hanya 56,25%.
Berdasarkan uji patogenisitas menggunakan 17 isolat terpilih, cendawan endofit dapat dibedakan menjadi 2 kelompok, yaitu patogenik (13 isolat) dan non- patogenik (4 isolat). Dengan demikian cendawan endofit yang diisolasi sebagian besar berpotensi sebagai patogen. Diantara 4 spesies cendawan endofit non patogenik, Cercospora nicotianae, Pleosporaceae, Guignardia mangifera dan Dothideomycete sp. diketahui berpotensi sebagai plant growth promoting fungi (PGPF) dan 2 spesies yaitu C. nicotianae dan Pleosporaceae berpotensi sebagai agens biokontrol terhadap penyakit kuning.
Cendawan endofit hasil isolasi yang berpotensi sebagai agens biokontrol diperoleh dari frekuensi kelimpahan rendah. C nicotianae berdasarkan isolasi bersifat singleton dan berpotensi sebagai agens biokontrol terhadap penyakit kuning.
SUMMARY
DAMAYANTI. Abundance and Potency of Endophytic Fungi to Suppress Yellow Leaf Curl Disease in Chilli Pepper (Capsicum annuum L.). Supervised by SRI HENDRASTUTI HIDAYAT and SURYO WIYONO.
Incidence of yellow leaf curl diseases caused by Pepper yellow leaf curl begomovirus (PepYLCV) has become a major problem in chilli pepper growing area, especially in Java and Sumatra since early 2000. Disease incidence in the field may reach 20 % to 100% and the disease was obviously recognized by its unique symptoms involving bright yellow mosaic, mottle, curling, and stunting. A research was conducted to explore the potency of endophytic fungi as an alternative strategy to suppress PepYLC disease.
Research activities consisted of 3 parts, i.e. (1) Isolation of endophytic fungi collected from chilli pepper plants at endemic area in Sleman, Yogyakarta; (2) Pathogenicity assay using chilli pepper seeds, and (3) Efficacy of selected endophytic fungi to suppress disease incidence.
The result showed that diversity of endophytic fungi isolated from unsymptomatic plants were significantly higher than those from infected plants. There were 19 isolates of species identified from unsymptomatic plants; whereas 13 isolates from infected plants. Based on pathogenicity assay using 17 selected isolates, the endophytic fungi could be differentiated into 2 groups, i.e. pathogenic (13 isolates) and non-pathogenic (4 isolates). Further screening of non-pathogenic isolates showed that 4 isolates might have a potential as plant growth promoting fungi (PGPF) and 2 isolates, identified as Cercospora nicotianae and Pleosporaceae, might be recommended as bio-control agent.
Endophytic fungi isolated in this study having potency as biocontrol agent was taken from low abundance population C. nicotianae was singleton and has potency as biocontrol agent for PepYLC disease.
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KELIMPAHAN DAN POTENSI CENDAWAN ENDOFIT
UNTUK MENEKAN PENYAKIT KUNING PADA TANAMAN
CABAI (
Capsicum annuum
L
.
)
DAMAYANTI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains
pada
Program Studi Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Kelimpahan dan Potensi Cendawan Endofit untuk Menekan Penyakit Kuning pada Tanaman Cabai (Capsicum annuum L.)
Nama : Damayanti
NIM : A352100011
Disetujui Komisi Pembimbing
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc. Dr. Ir. Suryo Wiyono, M.Sc.Agr.
Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc. Dr Ir . Dahrul Syah, MSc.Agr.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas karuniaNya sehingga penulis dapat menyelesaikan penelitian berjudul “Kelimpahan dan Potensi Cendawan Endofit untuk Menekan Penyakit Kuning pada Tanaman Cabai (Capsicum annuum L.)”. Penelitian ini bertujuan untuk melakukan eksplorasi dan seleksi cendawan endofit sebagai langkah awal eksplorasi agens biokontrol yang berpotensi menekan perkembangan penyakit kuning cabai. Sumber dana penelitian diperoleh dari Hibah Tim Pascasarjana, Direktorat Pendidikan Tinggi, Kementrian Pendidikan Nasional atas nama Dr. Sri Hendrastuti Hidayat.
Ucapan terima kasih dan penghargaan yang setulusnya penulis sampaikan kepada Dr. Sri Hendrastuti Hidayat dan Dr. Suryo Wiyono sebagai komisi pembimbing, atas bimbingan, saran dan masukan selama penyelesaian penelitian ini; kepada Prof. Dr. Meity Suradji Sinaga selaku dosen penguji luar komisi yang telah memberikan saran, masukkan pada penulisan hasil penelitian ini; kepada Program Beasiswa IM HERE B2C. IPB atas dana pendidikan Pascasarjana.
Penulis menyampaikan rasa terima kasih dan penghargaan kepada Dr. Widodo dan Dr. Efi Toding Tondok atas saran dan masukkannya, kepada teman-teman Laboratorium Virologi Tumbuhan, Pak Emput, Pak Edi, mbak Ita, mbak Yuke, Tuti Susanti Legiastuti, SSi, teman-teman Fitopatologi angkatan
2010 dan seluruh rekan-rekan mahasiswa Program Studi Entomologi dan Fitopatologi.
Ungkapan terima kasih juga disampaikan kepada bapak dan ibu, serta seluruh keluarga, atas segala doa dan kasih sayangnya. Ucapan terima kasih dan penghargaan yang setulusnya penulis sampaikan kepada suami tercinta Muhammad Nur Amin MSi atas kesabaran dan ketabahannya dalam mendampingi, memberi motivasi dan inspirasi selama penulis menyelesaikan penelitian. Kepada semua pihak yang tidak bisa penulis sebutkan satu per satu yang telah membantu selama penelitian diucapkan terima kasih.
Semoga karya ilmiah ini bermanfaat bagi ilmu pengetahuan dan masyarakat.
Bogor, Juni 2013
DAFTAR ISI
Tujuan Penelitian Manfaat Penelitian 3
Ruang Lingkup 3
II. TINJAUAN PUSTAKA 5
Biologi dan Ekologi PepYLCV Penyebab Penyakit Kuning Cabai 5
Pengendalian Virus dengan Pemanfaatan Senyawa Antiviral 7
Pemanfaatan Cendawan Endofit sebagai Agens Biokontrol 8
III. BAHAN DAN METODE 11
Waktu dan Tempat 11
Isolasi Cendawan Endofit 11
Deteksi Virus Kuning Cabai 11
Analisis Keragaman Cendawan Endofit 12
Uji Patogenisitas Cendawan Endofit 12
Identifikasi Cendawan Endofit 13
Perbanyakan Serangga Vektor 13
Perbanyakan Sumber Inokulum PepYLCV 13
Penyediaan Isolat Cendawan Endofit 13
Pengujian Efektivitas Cendawan Endofit sebagai Pemicu 13
Pertumbuhan Tanaman Cabai
Pengujian Efektivitas Cendawan Endofit sebagai Agens 14
Biokontrol Penyakit Kuning Cabai
IV. HASIL DAN PEMBAHASAN 16
Hasil 16
Deteksi Virus Penyebab Penyakit Kuning 16
Kelimpahan Cendawan Endofit 17
Patogenisitas Cendawan Endofit 20
Identifikasi Cendawan Endofit Potensial 20
Pengaruh Cendawan Endofit terhadap Pertumbuhan Tanaman 22 Cabai
Pengaruh Cendawan Endofit terhadap Infeksi Virus Kuning Cabai 22
DAFTAR TABEL
1 Skoring keparahan gejala pada tanaman cabai yang terinfeksi 15
PepYLCV
2 Kelimpahan spesies cendawan endofit berbasis segmentasi dan 19
tanaman hasil isolasi dari daun tidak bergejala dan bergejala yang berasal dari daerah endemis penyakit kuning di desa Minggir, DIY
3 Pengaruh cendawan endofit terhadap daya kecambah benih, panjang 20 akar dan tajuk bibit cabai
4 Identifikasi cendawan endofit potensial berdasarkan karakter morfologi 21 dan molekuler
DAFTAR GAMBAR
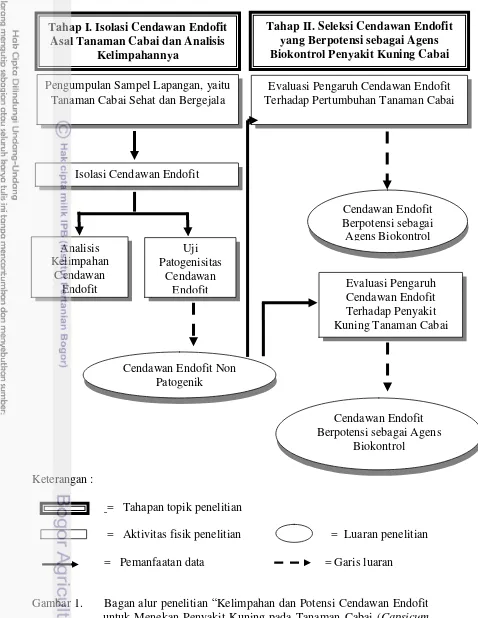
1 Bagan alur penelitian”kelimpahan dan potensi cendawan endofit untuk 4
menekan penyakit kuning pada tanaman cabai (Capsicum annuum L.)
2 Organisasi genom Begomovirus : (a) genom monopartit dan (b) genom 5
bipartit
3 Kondisi lahan di Kecamatan Minggir pada waktu pengambilan sampel 16
daun cabai. (a) lahan cabai tumpangsari dengan terung; (b) gejala penyakit kuning
4 Visualisasi fragmen DNA hasil amplifikasi dengan primer universal 17 Geminivirus pAL1v 1978/ pAR1c715. (A) sampel daun bergejala; (B) sampel daun tidak bergejala; (M) penanda DNA 1 kb; (P) kontrol positif (DNA Geminivirus); (N) kontrol negatif (tanpa DNA); nomor 1- 20 kode sampel daun
5 Kelimpahan cendawan endofit pada tanaman cabai yang diisolasi pada 18
tanaman bergejala dan tanaman tidak bergejala dari daerah endemis penyakit kuning
6 Pengaruh cendawan endofit terhadap tinggi tanaman (A) dan bobot 22 tajuk, bobot akar, volume akar pada tanaman cabai (B).
7 Pengaruh cendawan endofit terhadap kejadian penyakit (A), keparahan 23
penyakit (B) dan laju infeksi (C) penyakit kuning cabai
8 Pengaruh cendawan endofit terhadap tinggi tanaman (A) dan bobot 24 tajuk, bobot akar, volume akar (B) pada tanaman cabai yang
DAFTAR LAMPIRAN
1 Uji patogenesis cendawan endofit. A, Benih yang ditanamn dalam 36 media PDA tanpa dan dengan cendawan endofit; B, Screening cendawan patogenik dan non patogen (Gambar atas berpotensi non patogen dan gambar bawah berpotensi patogen); C, panjang bibit cabai (ujung kiri tanpa cendawan selainnya dengan cendawan endofit)
2 Identifikasi Isolat-isolat Non-patogenik 38
3 Identifikasi kutukebul Bemisia tabaci 39
4 Isolat PepYLCV dan gejala yang ditimbulkan. A, Sumber inokulum 40 virus dari lapang; B, Inokulum diperbanyak pada tanaman tomat varietas Ratna; C, Gejala yang ditimbulkan ketika PepYLCV ditularkan pada cabai TM 888
5 Kriteria skoring gejala pada waktu 1 bulan setelah inokulasi. A; Tanpa 41 gejala; B; gejala skor 1; C; gejala skor 2; D; gejala skor 3; E; skor gejala 4; F; skor gejala 5
I.
PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L.) merupakan salah satu komoditas hortikultura penting di Indonesia dan mempunyai nilai ekonomi yang cukup tinggi. Berdasarkan data statistik 2010 luas panen cabai mencapai 237, 520 ha dengan produktivitas 6.50 ton/ha untuk cabai besar dan 4.56 ton/ha untuk cabai rawit (Direktorat Jenderal Hortikultura 2012). Hasil tersebut lebih rendah dibandingkan dengan potensinya yang dapat mencapai 18 ton/ha. Banyak faktor pembatas yang dihadapi petani dalam sistem budidaya cabai salah satunya adalah gangguan penyakit yang disebabkan oleh virus. Tanaman cabai dilaporkan dapat terinfeksi oleh 35 jenis virus, dari jumlah tersebut lebih dari setengahnya ditularkan oleh vektor kutudaun dan sisanya ditularkan melalui vektor nematoda, trips, wereng, kutukebul, kumbang, cendawan maupun secara mekanis (Green dan Kim 1991).
Hidayat et al. (1999) melaporkan adanya penyakit kuning pada tanaman cabai di daerah Bogor yang disebabkan oleh Pepper yellow leaf curl begomovirus (PepYLCV), anggota kelompok Geminivirus. Penyakit kuning cabai umumnya menimbulkan gejala berupa mosaik kuning, belang-belang klorotik, daun mengerut, dan kerdil. Penularan di lapangan terjadi melalui serangga vektor Bemisia tabaci (Gennadius) (Hemiptera : Aleyrodidae) (Mason et al. 2000), dan PepYLCV dilaporkan memiliki tanaman inang alternatif yang luas (Shafiq et al. 2010).
Pada tahun 2000 sampai 2003 terjadi epidemi penyakit kuning di daerah Jawa Tengah (Sulandari et al. 2006). Kerusakan karena penyakit kuning cabai juga sudah dilaporkan terjadi di beberapa sentra produksi sayuran seperti di Sumatera Utara, Sumatera Barat, Lampung, Bengkulu, Kalimantan Barat, Kalimantan Selatan, Jawa Barat, Jawa Tengah, Daerah Istimewa Yogyakarta, Jawa Timur, dan Nusa Tenggara Barat, dengan intensitas penyakit berkisar antara 20% sampai 100% (Hikmat 2005).
Strategi pengelolaan penyakit kuning yang umum dilakukan adalah dengan mengendalikan serangga vektor, baik secara fisik maupun menggunakan insektisida kimia. Pengendalian fisik dapat dilakukan dengan menggunakan sungkup kedap serangga di tempat persemaian yang berfungsi menghalangi masuknya serangga vektor ke areal pesemaian. Penggunaan kultivar yang tahan ataupun toleran merupakan alternatif yang cukup baik dalam upaya pengendalian. Walaupun demikian, upaya untuk mendapatkan varietas tahan tidak mudah karena sumber ketahanan terhadap penyakit kuning cabai sangat terbatas. Pengendalian hayati yang sudah dilakukan menggunakan mikrob pemacu pertumbuhan tanaman (misalnya plant growth promoting rhizobactera) dan penggunaan ekstrak tanaman seperti bunga pukul empat (Mirabilis jalapa), bayam duri (Amaranthus spinosus) dan eceng gondok (Eichhornia crassipes) memberikan efek yang cukup baik dalam menekan perkembangan penyakit kuning cabai (Badan Litbang Pertanian 2011).
2
endofit di alam cukup tinggi. Pada tahun 2000 ada rata-rata 50 jenis cendawan endofit per tanaman yang ditemukan dan jumlahnya semakin meningkat ketika metode molekuler mulai digunakan untuk identifikasi (Zabalgogeazcoa 2008). Umumnya, cendawan mitosporik, Ascomycetes, dan beberapa cendawan steril tercatat sebagai endofit yang berhasil diisolasi dari semua tanaman inang. Isolasi yang dilakukan pada Deschampsia antarctica Desv. (Poaceae), yang merupakan tanaman vaskular dari benua Antartika dan hidup pada kondisi lingkungan ekstrim, berhasil memperoleh 26 isolat dari 273 segmen daun yang diambil. Diantaranya 91 individu tanaman teridentifikasi sebagai genus Alternaria dan Phaeosphaeria dengan kelimpahan tinggi (Rosa et al. 2009). Suryanaranan et al. (2000) melakukan isolasi cendawan endofit dari beberapa jaringan tanaman yang berbeda, dan melaporkan hasil sebagai berikut: pada jaringan batang Abutilon indicum diperoleh 14 spesies, dari Cucurbita maxima 2 spesies, dari Achyranthes aspera 11 spesies, dari Colotropis gigantea 6 spesies, dari Pongamia gabra 11 spesies, dari Phyllantus reticulantus 14 spesies, dan dari Zizyphus jujuba 11 spesies. Hasil tersebut menunjukkan adanya beberapa derajat spesifisitas antara inang dengan cendawan endofit.
Kajian cendawan endofit di bidang pertanian sudah banyak dilakukan, antara lain berkaitan dengan potensinya meningkatkan toleransi tanaman terhadap kekeringan, menghambat perkembangan serangga herbivora, cendawan patogen, virus dan nematoda (Rranzluebbers et al. 1999). Beberapa hasil penelitian menunjukkan potensi cendawan endofit dalam menekan populasi serangga vektor. Penelitian Lehtonen et al. (2006) menyatakan bahwa aplikasi cendawan endofit secara nyata mampu mengurangi frekuensi tanaman Meadow ryegrass (Lolium pratense) yang terinfeksi oleh Barley yellow dwarf virus (BYDV). Penurunan frekuensi tanaman yang terinfeksi BYDV berkorelasi positif terhadap penurunan
kutudaun vektor BYDV pada Meadow ryegrass. Dilaporkan lebih lanjut
cendawan endofit Acremonium coenophialum dapat mengendalikan
Rhopalosiphum padi L. (Hemiptera: Aphididae) yang merupakan vektor BYDV yang menginfeksi tanaman Festuca arundinacea.
Eksplorasi dan seleksi cendawan endofit merupakan langkah awal untuk menyediakan agens biokontrol yang berpotensi menekan penyakit kuning pada tanaman cabai. Penelitian dilakukan dengan tujuan untuk mempelajari kelimpahan cendawan endofit dari daerah endemis penyakit kuning cabai di Yogyakarta dan potensinya dalam menekan penyakit kuning cabai.
Perumusan Masalah
Begomovirus dilaporkan sebagai salah satu virus yang berpotensi menurunkan produktivitas tanaman baik di daerah tropis maupun sub tropis dan memiliki daerah penyebaran yang sangat luas. Kelompok Begomovirus yang menginfeksi tanaman cabai teridentifikasi sebagai PepYLCV. Penyebaran PepYLCV sangat cepat karena ditularkan melalui serangga vektor B. tabaci. Di Indonesia, PepYLCV sudah tersebar luas dengan intensitas serangan berkisar antara 20% sampai 100%, bahkan dilaporkan bahwa 1.4 ha tanaman cabai besar mengalami puso di Sumatera Barat.
aplikasi ekstrak tanaman. Pengendalian menggunakan insektisida dirasa kurang efektif dan apabila digunakan secara intensif dapat mengganggu keseimbangan lingkungan. Aplikasi insektisida tidak efektif untuk serangga vektor PepYLCV karena satu serangga viruliferous sudah mampu menularkan virus. Selain itu vektor Begomovirus ini memiliki kisaran inang yang banyak, pergerakannya cepat, dan kemampuannya menjadi resisten terhadap insektisida sangat cepat. Penggunaan varietas tahan merupakan alternatif pengendalian yang cukup menjanjikan, akan tetapi untuk mendapatkan varietas tahan tidaklah mudah. Diperlukan kajian secara mendalam untuk mendapatkan teknik pengendalian Begomovirus tersebut, salah satunya mengkaji peranan cendawan endofit.
Kajian tentang cendawan endofit sudah banyak dilakukan di sektor pertanian, farmasi bahkan di sektor industri. Beberapa studi cendawan endofit di bidang pertanian yang sudah dilakukan dalam kaitannya sebagai agens biokontrol yaitu pemanfaatan cendawan endofit dalam mengendalikan nematoda, serangga hama, dan cendawan patogen tumbuhan, sedangkan terhadap virus tumbuhan di Indonesia tergolong sebagai hal yang relatif baru. Dalam penelitian ini dilakukan analisis kelimpahan cendawan endofit pada tanaman cabai di daerah endemis penyakit kuning serta seleksi cendawan endofit yang bermanfaat sebagai agens biokontrol terhadap penyakit tersebut.
Tujuan Penelitian
1. Mengetahui keragaman dan kelimpahan cendawan endofit pada tanaman cabai yang tidak bergejala dan yang menunjukkan gejala penyakit kuning cabai.
2. Melakukan eksplorasi cendawan endofit non patogenik yang berpotensi menekan perkembangan penyakit kuning cabai.
3. Melakukan pengujian untuk mencari cendawan endofit yang potensial sebagai agens biokontrol untuk penyakit kuning cabai.
Manfaat Penelitian
Penelitian ini akan menghasilkan agens biokontrol yang dapat dimanfaatkan dalam upaya pengendalian penyakit kuning cabai melalui strategi pengendalian berbasis biointensif.
Ruang Lingkup
4
II. TINJAUAN PUSTAKA
Biologi dan Ekologi PepYLCV Penyebab Penyakit Kuning Cabai Penyebab penyakit kuning yang menginfeksi tanaman cabai termasuk dalam famili Geminiviridae, genus Begomovirus. Berdasarkan struktur genom, serangga vektor, dan jenis tanaman inangnya famili Geminiviridae dibedakan
menjadi empat genus yaitu Mastrevirus, Curtovirus, Begomovirus, dan
Topocuvirus (Hull 2002). Mastrevirus adalah anggota Geminivirus yang menginfeksi tanaman monokotil, ditularkan oleh vektor wereng daun, dan memiliki genom monopartit. Curtovirus adalah anggota Geminivirus yang menginfeksi tanaman dikotil, ditularkan oleh vektor wereng daun, dan memiliki genom monopartit. Topocuvirus adalah anggota Geminivirus yang menginfeksi tanaman dikotil, ditularkan oleh wereng pohon, dan memiliki genom mirip Curtovirus karena genus ini merupakan bagian dari genus Curtovirus. Begomovirus adalah anggota Geminivirus yang menginfeksi tanaman dikotil, ditularkan oleh vektor kutukebul, dan memiliki genom bipartit atau monopartit.
Begomovirus yang memiliki genom bipartit terdiri dari dua molekul DNA utas tunggal berbentuk sirkuler yang berbeda yaitu DNA A dan DNA B masing-masing berukuran 2.7-2.8 kb yang terkapsidasi dalam partikel kembar (geminate). Begomovirus dengan genom monopartit semua gennya terletak pada satu molekul DNA utas tunggal berbentuk sirkuler yang berukuran 2.8 kb (Gambar 2).
Gambar 2. Organisasi genom Begomovirus : (a) Genom monopartit dan (b) Genom bipartit (Sumber : Hull 2002)
Pada umumnya Begomovirus yang berasal dari bumi belahan barat memiliki genom bipartit dengan komponen DNA A dan DNA B. Molekul DNA A mempunyai peranan menyandi gen yang bertanggung jawab terhadap replikasi virus, regulasi ekspresi gen, dan enkapsidasi partikel, sedangkan molekul DNA B mempunyai peranan menyandi protein yang berperan dalam pergerakan virus, membedakan kisaran inang dan perkembangan gejala penyakit. Begomovirus yang menginfeksi tanaman cabai memiliki genom monopartit, mengandung DNA
A dan β-segmen (Sinha et al. 2011).
Inang Begomovirus meliputi tanaman pangan penting termasuk
6
Ageratum conyzoides merupakan gulma yang umum menjadi inang Begomovirus pada daerah tropis dan subtropis.
Begomovirus memiliki daerah penyebaran yang sangat luas terutama di daerah tropis dan sub tropis. Begomovirus mempunyai anggota yang paling banyak dibandingkan dengan genus lainnya dari famili Geminiviridae. Di Indonesia Begomovirus pertama kali diketahui menginfeksi tanaman tembakau di Bojonegoro, Jawa Timur yang menyebabkan kerusakan sebesar 30% (Trisusilowati 1990 dalam Meliansyah R 2010). Hidayat et al. (1999) melaporkan adanya penyakit kuning yang disebabkan oleh Begomovirus pada tanaman cabai di wilayah Bogor yang kemudian dinamakan sebagai Pepper yellow leaf curl Indonesia virus (PepYLCIV). PepYLCV juga sudah dilaporkan di USA, Nigeria, Pakistan, Bangladesh. Di India PepYLCV telah terdeteksi dan menjadi sebuah ancaman karena dapat menyebabkan kehilangan hasil mencapai 100% (Sinha et al 2011).
Infeksi PepYLCV pada umumnya menyebabkan daun menjadi lebih kecil dibandingkan dengan tanaman normal, warna daun menjadi kekuningan, beberapa menunjukkan mosaik kuning yang dimulai dari pangkal daun yang kemudian menyebar disertai pelekukan tepi daun kearah atas, dan tanaman menjadi kerdil (Rusli et al. 1999). Gejala yang timbul karena infeksi Begomovirus pada tanaman cabai dipengaruhi oleh strain virus, kultivar, lingkungan, dan umur tanaman saat terinfeksi. Berdasarkan penelitian yang dilakukan Sulandari et al. (2006) infeksi pada cabai rawit menyebabkan semua helaian daun menguning, sedangkan pada cabai besar daun dibagian pucuk menguning sedangkan bagian bawah masih tetap hijau. Tanaman cabai akan kehilangan produktivitasnya apabila terinfeksi pada saat tanaman masih muda karena akan menjadi kerdil sehingga tidak berbuah.
Di Indonesia, PepYLCV sudah tersebar luas. Pada tahun 2001 kejadian penyakit dan luas serangan pada cabai rawit sekitar 70 % sampai100% yang diamati di daerah Yogyakarta (Bantul, Sleman, Kulon Progo dan Gunung Kidul), Jawa Tengah (Klaten, Kopeng dan Muntilan), dan Jawa Barat (Segunung, Cugenang, Lembang, dan Megamendung). Infeksi pada cabai merah jenis keriting menyebabkan kejadian penyakit terjadi secara sporadis sekitar 10 % sampai 35% dan pada pengamatan lapangan tahun 2002 di Kulon Progo dan awal tahun 2003 di Sleman dan Kopeng meningkat mencapai 70 % sampai100% dengan gejala yang sangat parah yaitu daun menguning dan tanaman kerdil (Sulandari et al. 2006). Daryanto (2005) dalam Haryadi (2006) melaporkan bahwa 1.4 ha tanaman cabai besar mengalami puso di Sumatera Barat. Di Lampung, intensitas serangan penyakit berkisar antara 20 % sampai100%.
menggunakan vektor lebih tinggi dibandingkan dengan penyambungan. Satu ekor B. tabaci viruliferous mampu menularkan virus dan menyebabkan infeksi.
Epidemi penyakit terjadi karena adanya inang yang rentan, patogen yang virulen dan lingkungan yang mendukung. Pada kasus PepYLCV vektor sangat mempengaruhi terjadinya epidemi karena satu ekor B. tabaci viruliferous mampu menularkan virus dan menyebabkan infeksi. Ketahanan varietas tanaman juga memiliki pengaruh nyata terhadap epidemi, peningkatan kejadian penyakit yang cukup tinggi di daerah Kulon Progo, Kopeng dan Sleman terjadi sejak petani menanam cabai varietas TM 999 yang rentan terhadap PepYLCV (Sulandari et al.
2006). Perkembangan penyakit yang disebabkan oleh PepYLCV meningkat
secara nyata seiring berjalannya waktu. Pengendalian penyakit tanaman yang disebabkan oleh virus termasuk penyakit kuning cabai sulit dilakukan. Secara umum pengendalian yang dilakukan untuk penyakit yang disebabkan oleh virus adalah penggunaan bibit sehat, perlakuan benih, pengendalian vektor virus, secara kultur teknis, pencegahan dengan menggunakan strain avirulen, aplikasi senyawa antiviral dan penggunaan kultivar tahan (Su dan Chen 1991). Selama ini yang dilakukan untuk mengendalikan penyakit kuning adalah dengan mengendalikan serangga vektornya. Pengendalian vektor dilakukan secara kimiawi menggunakan insektisida dan secara fisik dengan mengeradikasi sumber inokulum, menggunakan sungkup kedap serangga pada waktu persemaian dan menanam tanaman pinggir (border crop) sehingga menghalangi masuknya serangga vektor ke areal pertanaman. Penggunaan kultivar tahan merupakan alternatif yang cukup baik dalam pengendalian virus. Ganefianti et al. (2008) mendapatkan satu genotipe cabai tahan (genotipe IPB12) dari 27 genotipe yang diuji terhadap penyakit kuning cabai. Hal tersebut menunjukkan bahwa untuk mendapatkan varietas tahan tidak mudah.
Pengendalian Virus dengan Pemanfaatan Senyawa Antiviral
Pemanfaatan senyawa antiviral atau inhibitor pada virus tanaman dapat dibedakan menjadi dua kategori yaitu: inhibitor terhadap multiplikasi dan inhibitor terhadap infeksi virus. Aplikasi subtansi inhibitor sebelum atau bersamaan pada saat inokulasi virus dapat menghambat proses infeksi. Subtansi antiviral yang sudah dipelajari berasal dari ekstrak tanaman, proteinaceous dan subtansi dari mikroorganisme. Subtansi dari mikroorganisme yang sudah dipelajari adalah polisakarida kelompok glucan dan turunannya. Rouhier et al. (1995) melaporkan bahwa dinding sel Phytophtora infestans, yang mengandung polisakarida, ketika dicampur dengan Potato virus x dapat menekan kejadian penyakit pada tembakau. Disimpulkan bahwa glucan mempunyai aktivitas sebagai antiviral pada beberapa virus.
8
Potato virus Y pada tembakau. Pengujian lain yang dilakukan menggunakan kultur filtrat dari Trichotesium roseum menunjukkan penghambatan yang disebabkan oleh polisakarida termasuk D-galaktosa yang merupakan komponen gula. Inhibitor trichothecin dinyatakan efektif jika disemprotkan di daun pada waktu tidak lebih dari dua hari sebelum inokulasi atau tidak lebih dari satu hari setelah inokulasi. Efisiensi dari inhibitor tergantung dari spesies inang dan virus yang diinokulasikan. Antiviral yang lainnya seperti RNA ragi dengan konsentrasi tinggi (5 mg/ml) diaplikasikan pada daun atau dicampur pada inokulum dapat mengurangi jumlah lesio lokal yang disebabkan oleh TMV pada N. glutinosa.
Berdasarkan uraian di atas, eksplorasi antiviral dari mikroorganisme termasuk cendawan endofit perlu dikaji lebih mendalam dalam upaya pengendalian penyakit yang disebabkan oleh virus tanaman.
Pemanfaatan Cendawan Endofit Sebagai Agens Biokontrol
Endofit didefinisikan sebagai semua organisme yang menghuni organ tanaman, yang dalam beberapa waktu dalam hidupnya dapat mengolonisasi bagian dalam jaringan tanaman tanpa menimbulkan kerusakan nyata bagi inangnya (Petrini 1991). Cendawan endofit telah dilaporkan terdeteksi pada ratusan spesies tanaman, yang pada awalnya terdeteksi sebagai simbion pada tanaman rumput (Zabalgogeazcoa 2008). Saat ini cendawan endofit banyak dikaji pada tanaman komersial seperti gandum, pisang, kedelai, dan tomat (Vega et al. 2008).
Berdasarkan teori evolusinya, cendawan endofit berasal dari cendawan patogen yang berkembang menjadi patogen minor dan kemudian menjadi simbion yang bersifat mutualisme. Strategi hidup simbion ini berevolusi dari saprofit fakultatif, parasit, exploitive, dan mutualisme. Lebih lanjut semua interaksi cendawan endofit berperan dalam menyediakan nutrisi dan penyangga dari tekanan lingkungan eksternal dan kompetisi mikrob (Schulz dan Boyle 2005). Sifat mutualisme cendawan endofit pada tanaman mempunyai peranan yang kompleks. Beberapa studi menyatakan bahwa cendawan endofit mempunyai peranan penting sebagai sumber produk alami untuk farmasi dan pertanian (Suryanarayanan et al. 2009).
deteksi yang sangat sensitif untuk mendeteksi keberadaannya (Koulman et al. 2007).
Dewasa ini cendawan endofit secara nyata dapat meningkatkan nilai hasil pertanian dan menekan gangguan hama dan penyakit (Koulman et al. 2007). Cendawan endofit dari daun sudah dilaporkan dapat berperan sebagai agens biokontrol terhadap panyakit pada tanaman kakao di daerah tropis. Beberapa strain dari Trichoderma, seperti T. harzianum, T. stromaticum, dan T. asperellum sudah dilaporkan mempunyai efek antagonis pada beberapa penyakit yang disebabkan oleh cendawan (Gazis et al. 2010). Terdapat perbedaan mekanisme endofit dalam melindungi tanaman, tetapi produksi antibiotik dan metabolit seluler yang bersifat toksik merupakan mekanisme utama endofit dalam proteksi tanaman (Chaves et al. 2009).
Peranan cendawan endofit di bidang pertanian yang sudah dikaji antara lain sebagai proses adaptasi tanaman seperti peningkatan pertumbuhan, toleransi terhadap cekaman kekeringan, anti serangga herbivora, dan pertahanan terhadap penyakit tanaman. Mekanisme cendawan endofit dalam proses adaptasi tanaman terhadap cekaman kekeringan dengan menurunkan proses fotosintesis, ketahanan stomata terhadap kekeringan dan terjadinya proliferasi pada bagian akar (Rranzluebbers et al. 1999). Selain itu cendawan endofit mampu menginduksi atau mensintesis senyawa pemicu pertumbuhan tanaman seperti auksin, sitokinin, dan fiksasi nitrogen (Hanoda et al. 2010). Mekanisme penghambatan cendawan endofit terhadap patogen dapat secara langsung dengan mekanisme parasitisme, antibiosis, kompetisi nutrisi penambahan pertumbuhan tanaman dan mekanisme ketahanan terinduksi (Gaziz et al. 2010).
Pada mekanisme penghambatan patogen, cendawan endofit menghasilkan metabolit fungsional yang termasuk dalam group terpenoids, steroids, xanthones, chinones, phenol, isocoumarins, benzopyranones, tetralones, cytochalasines dan enniatines. Metabolit fungsional yang dihasilkan mempunyai peranan sebagai anti bakteri, antiviral, anti cendawan dan anti kanker (Suryanarayanan et al. 2009). Hasil penelitian Sumarah et al. (2010) menunjukkan tiga strain dari 150 ekstrak metabolit seluler dari cendawan endofit yang diisolasi pada tanaman Picea rubens bersifat toksik terhadap serangga Choristoneura fumiferana. Metabolit fungsional yang dihasilkan oleh cendawan endofit yang mempuyai peranan sebagai insektisida diantaranya : Indol derivativ-lolitrem, Pyrrozilidin-lolin, heptalidic acid (dihasilkan oleh cendawan Phyllosticta). Metabolit yang berfungsi sebagai fungisida yaitu : chokol, phynilpropanoid, 6-isoprenylindole-3-carboxylic acid, dan yang berfungsi sebagai bakterisida yaitu: subglutinol yang dihasilkan oleh F. subglutinon sedangkan phenol dan phenolic mempunyai peranan yang cukup luas dapat sebagai anti cendawan, anti bakteri dan antiviral. Beberapa cendawan endofit yang telah dilaporkan berpotensi menekan cendawan patogen
yaitu Trichoderma, Fusarium, Penicillium, Phoma dan Acremonium.
Trichoderma dan Fusarium non patogen dapat menekan patogen F. oxysporum secara in vitro (Istikorini 2008).
10
frekuensi tanaman yang terinfeksi oleh BYDV berkorelasi positif terhadap
penurunan kutudaun pada Meadow ryegrass yang merupakan vektor BYDV.
Endofit Acremonium coenophialum dilaporkan dapat mengendalikan
III. BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan mulai bulan April 2011 sampai Desember 2012. Pengambilan sampel daun cabai dilakukan di Kecamatan Minggir, Sleman, Daerah Istimewa Yogyakarta (DIY). Isolasi cendawan dilakukan di Laboratorium Mikologi Tumbuhan Departemen Proteksi Tanaman, Fakultas Pertanian, IPB, dan kegiatan deteksi virus dilakukan di Laboratorium Virologi Tumbuhan Departemen Proteksi Tanaman, Fakultas Pertanian, IPB. Kegiatan identifikasi serangga vektor dilakukan di Laboratorium Taksonomi Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, IPB dan kegiatan pengujian efektivitas cendawan endofit dilakukan di rumah kaca Cikabayan, Fakultas Pertanian, IPB.
Isolasi Cendawan Endofit
Isolasi cendawan endofit dilakukan menggunakan sampel daun cabai yang diambil di daerah endemis penyakit kuning cabai di Kabupaten Sleman, DIY. Sampel terdiri dari daun yang bergejala, yaitu daun keriting dan kuning, dan daun yang tidak bergejala.
Metode isolasi cendawan endofit mengikuti metode Photita et al. (2001) dengan modifikasi. Sampel daun tanaman cabai dicuci bersih kemudian diberi perlakuan sterilisasi permukaan. Perlakuan sterilisasi dilakukan secara bertahap, pertama-tama merendam daun dalam alkohol 96% selama 30 detik, dilanjutkan dalam larutan NaOCl 1% selama 1 menit, alkohol 70% selama 1 menit, alkohol 70% selama 30 detik, kemudian dibilas dua kali dengan air steril serta dikeringkan pada kertas steril. Setelah tahap sterilisasi daun dipotong-potong menjadi segmen kecil dan ditanam pada cawan petri yang berisi media potato dextrose agar (PDA) 25%. Pada setiap cawan petri ditanam 4 segmen daun, jumlah cawan petri masing-masing 20 untuk sampel daun bergejala dan tidak bergejala. Miselium yang tumbuh pada potongan segmen daun kemudian dimurnikan pada media PDA 100% dan isolat disimpan dalam media PDA 100% pada agar miring sampai dilakukan pengujian.
Deteksi Virus Kuning Cabai
Konfirmasi infeksi virus pada sampel daun dari lapangan dilakukan dengan metode polymerase chain reaction (PCR) menggunakan primer universal
Geminivirus yaitu PAL1v 1978 (5’GCATCTGCAGGCCCACATYGTCTTYCC
NGT3’) dan PAR1c 715 (5’GATTTCTGCAGTTDATRTTYTCRTCCATCC
A3’) (Rojas et al. 1993).
12
diambil, diletakkan pada suhu ruang selama 2 menit kemudian ditambahkan 500 µl campuran kloroform:isoamil alkohol (CI) dengan perbandingan 24:1. Larutan tersebut dicampur dengan vorteks selama 5 menit kemudian disentrifugasi selama 15 menit pada kecepatan 12,000 rpm. Supernatan yang terbentuk kemudian diambil secara hati-hati dan dipindahkan ke tabung eppendorf baru dan ditambahkan 1/10 sodium asetat (CH3CooNa 3 M pH 5,2), 2,5 x volume etanol
absolut dicampur dengan baik untuk presipitasi DNA, kemudian diinkubasi pada -20 °C semalam. Suspensi tersebut kemudian disentrifugasi selama 10 menit pada kecepatan 12,000 rpm. Supernatan dibuang dan endapan dicuci dengan etanol 80% kemudian disentrifugasi selama 5 menit pada kecepatan 12,000 rpm. Supernatan dibuang, endapan dikeringkan di laminar flow atau mesin vacum. Setelah kering endapan DNA dilarutkan kembali dengan 100 µl bufer TE dan disimpan pada suhu -20 °C sampai dilakukan PCR.
Komponen reaksi PCR terdiri dari : H2O (18,8 µ)l; bufer 10x+Mg2+
(2,5µl); d NTP 10 mM (0,5 µl); primer PAL1v 1978 dan primer PAR1c 715 (masing-masing 1 µl); Dream taq DNA polymerase (0,2 µl) dan DNA template (1 µl), sehingga total volume satu reaksi 25 µl. Program amplifikasi pada mesin PCR terdiri atas pradenaturasi pada suhu 94 °C selama 4 menit, denaturasi 94°C selama 1 menit, penempelan primer (anneling) pada suhu 55 °C selama 1 menit dan extension 72 °C selama 5 menit. Siklus diulang sebanyak 35 kali. Hasil amplifikasi PCR divisualisasi menggunakan gel agarosa (1%) dan dielektroforesis pada tegangan 100 V selama 30 menit. Hasil elektroforesis dapat dilihat menggunakan sinar ultraviolet pada transiluminator.
Analisis Keragaman Cendawan Endofit
Analisis keragaman dan kelimpahan cendawan endofit dihitung berdasarkan frekuensi kolonisasi (FK) dan koefisien kemiripan (Cs). Frekuensi kolonisasi cendawan endofit dapat dihitung mengikuti metode Hata et al. (1995) dalam Suryanarayan et al. (2001), dengan rumus:
FK = (NcoI / Nt) x 100%
dengan Ncol adalah jumlah segmen yang dikolonisasi cendawan, Nt adalah jumlah segmen yang diamati.
Koefisien kemiripan dapat dihitung berdasarkan indeks Sorensen (Cs) (Maguran 1988), dengan rumus:
Cs = 2j/(a+b)
dengan j adalah jumlah spesies cendawan endofit pada kedua jaringan yang diujikan, a adalah jumlah spesies cendawan yang mengolonisasi jaringan a dan b adalah jumlah spesies cendawan yang mengolonisasi jaringan b.
Uji Patogenisitas Cendawan Endofit
cawan petri ditanam 20 biji cabai dan diulang tiga kali untuk masing-masing isolat cendawan endofit. Setelah dua minggu diamati persentase daya kecambah, panjang tajuk, dan panjang akar.
Identifikasi Cendawan Endofit
Berdasarkan hasil uji patogenisitas, isolat cendawan yang memiliki potensi sebagai agens biokontrol diidentifikasi berdasarkan warna koloni dan morfologi secara mikroskopik serta dibandingkan dengan kunci identifikasi menurut Watanabe (2002) (Lampiran 2).
Perbanyakan Serangga Vektor
Serangga vektor yang digunakan untuk penularan virus adalah kutukebul Bemisia tabaci yang telah dikonfirmasi melalui metode identifikasi menurut Watson (2007) (Lampiran 3). Perbanyakan serangga vektor dilakukan pada tanaman kapas dalam kurungan kedap serangga.
Perbanyakan Sumber Inokulum PepYLCV
Virus daun keriting kuning cabai (PepYLCV) yang digunakan dalam penelitian ini adalah isolat yang berasal dari tanaman cabai di Desa Minggir, Sleman, DIY yaitu pada lokasi yang sama dengan asal sampel cendawan endofit. Isolat tersebut diperbanyak pada tanaman tomat varietas Ratna melalui penularan dengan serangga vektor (Aidawati 2006) dan dua bulan setelah inokulasi tanaman digunakan sebagai sumber inokulum (Lampiran 4).
Penyediaan Isolat Cendawan Endofit
Cendawan endofit yang digunakan merupakan hasil seleksi berdasarkan hasil uji patogenisitas sebelumnya, yaitu Curvularia sp., C. nicotianae, menggunakan air steril dan divakum, kemudian miselia dihancurkan menggunakan blender hingga menjadi segmen-segmen kecil, selanjutnya dihitung konsentrasinya menggunakan haemositometer. Untuk pengujian efikasi, cendawan endofit diencerkan dari larutan stok hingga mencapai konsentrasi 105miselia/ ml dan ditambahkan Tween dengan konsentrasi 0,001% sebagai perekat.
Pengujian Efektivitas Cendawan Endofit sebagai Pemicu Pertumbuhan Tanaman Cabai
14
pupuk kandang steril (3:1). Satu minggu setelah pindah tanam, tanaman disemprot dengan suspensi cendawan endofit pada konsentrasi 105 miselia/ml. Sebagai kontrol adalah tanaman tanpa aplikasi suspensi cendawan endofit.
Rancangan percobaan yang digunakan adalah rancangan acak lengkap dengan sepuluh ulangan, setiap ulangan terdiri dari 1 tanaman contoh. Data diolah dengan analisis sidik ragam dan dilanjutkan dengan uji selang berganda Duncan (DMRT) pada taraf nyata 5% menggunakan SAS versi 6.12. Pengamatan dilakukan terhadap peubah agronomis seperti tinggi tanaman, bobot tajuk, bobot akar dan volume akar. Tinggi tanaman dihitung dari pangkal batang yang dekat dengan tanah sampai ke titik tumbuh, bobot tajuk ditimbang dengan memotong bagian pangkal batang sampai ke titik tumbuh, bobot akar ditimbang dari pangkal batang sampai ujung akar dan volume akar dihitung menggunakan gelas ukur.
Pengujian Efektivitas Cendawan Endofit sebagai Agens Biokontrol Penyakit Kuning Cabai
Cendawan endofit dan benih cabai yang digunakan sama dengan yang digunakan dalam efikasi terhadap pertumbuhan tanaman cabai di atas. Demikian pula penyiapan benih cabai sampai dengan pindah tanam dan aplikasi cendawan endofit dengan penyemprotan dilakukan seperti diuraikan pada uji efikasi cendawan endofit terhadap pertumbuhan tanaman di atas. Tanaman cabai berumur 4 minggu setelah semai siap untuk diinokulasi dengan isolat virus kuning melalui serangga vektor. Penularan dilakukan dengan 10 ekor serangga viruliferous setiap tanaman, periode makan akuisisi 24 jam dan periode makan inokulasi 48 jam (Aidawati 2006). Sebagai kontrol adalah tanaman tanpa aplikasi suspensi cendawan endofit.
Rancangan percobaan yang digunakan adalah rancangan acak kelompok dengan tiga ulangan, setiap ulangan terdiri dari 25 tanaman contoh. Data diolah dengan analisis sidik ragam dan dilanjutkan dengan uji selang berganda Duncan (DMRT) pada taraf nyata 5% menggunakan SAS versi 6.12. Pengamatan dilakukan terhadap peubah agronomis seperti tinggi tanaman, bobot tajuk, bobot akar dan volume akar dan peubah penyakit meliputi masa inkubasi, kejadian penyakit, laju infeksi dan keparahan penyakit. Kejadian penyakit dihitung mengikuti rumus:
KP = (n/N) x 100%
dengan n adalah jumlah tanaman bergejala dan N adalah jumlah seluruh tanaman yang diamati.
Laju infeksi dihitung berdasarkan rumus:
r=2,3/t2-t1[log10(1/1-x2)-log10(1/1-x1)],
Tabel 1. Skoring keparahan gejala pada tanaman cabai yang terinfeksi PepYLCV
indeks Gejala
0 Tidak menunjukkan gejala
1 Daun pucuk menunjukkan warna kuning pada pangkal daun
2 Daun pucuk menguning seluruhnya
3 Sekitar 50% daun menguning
4 Daun keriting dan hampir 100% daun menguning
5 Tanaman kerdil
Keparahan penyakit dihitung berdasarkan rumus Townsend dan Heuberger (1943) dalam Agrios (2005):
Kp = (ni x zi) /(NZ) x 100% ]
16
IV HASIL DAN PEMBAHASAN
Hasil
Deteksi Virus Penyebab Penyakit Kuning
Pengambilan sampel dilakukan pada satu luasan lahan yang menunjukkan gejala penyakit kuning cabai. Sampel daun cabai berasal dari varietas TM 999 yang telah berumur 3 bulan. Pola tanam pada lahan cabai yang diamati adalah pola tanam tumpang sari antara tanaman cabai dengan tanaman terung, dengan sedikit aplikasi pestisida kimia dan tidak disiangi sehingga gulma tumbuh subur pada lahan tersebut (Gambar 3a). Cara budidaya dilakukan dengan bedengan dan ditutupi mulsa plastik. Sampel daun tanaman menunjukkan gejala menguning dibagian pucuk sedangkan bagian bawah masih tetap hijau, terdapat juga gejala mosaik dan daun mengecil (Gambar 3b).
Gambar 3. Kondisi lahan di Kecamatan Minggir pada waktu pengambilan sampel daun cabai. (a) lahan cabai tumpangsari dengan terung; (b) gejala
penyakit kuning.
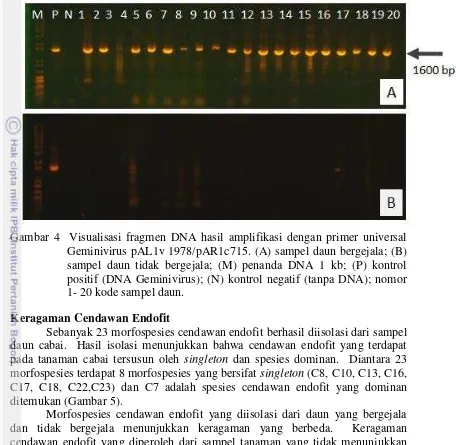
Amplifikasi fragmen DNA menggunakan primer universal Geminivirus PAL1v 1978 dan PAR1c 715 berhasil mendapatkan pita DNA berukuran 1600 bp dari semua sampel daun yang bergejala dan satu daun yang tidak bergejala (Gambar 4). Hasil tersebut membuktikan adanya infeksi pada semua daun bergejala dan satu daun tidak bergejala. Fragmen DNA hasil amplifikasi tersebut digunakan sebagai bahan untuk perunutan DNA (sequensing) dan dilanjutkan dengan analisis genetik berdasarkan rujukan di GenBank. Berdasarkan analisis sekuen DNA diperoleh 100% kemiripan dengan Pepper yellow leaf curl Indonesia virus (AB267838.1) (Lampiran 6).
Gambar 4 Visualisasi fragmen DNA hasil amplifikasi dengan primer universal Geminivirus pAL1v 1978/pAR1c715. (A) sampel daun bergejala; (B) sampel daun tidak bergejala; (M) penanda DNA 1 kb; (P) kontrol positif (DNA Geminivirus); (N) kontrol negatif (tanpa DNA); nomor 1- 20 kode sampel daun.
Keragaman Cendawan Endofit
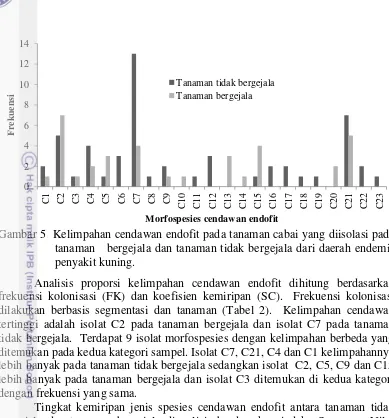
Sebanyak 23 morfospesies cendawan endofit berhasil diisolasi dari sampel daun cabai. Hasil isolasi menunjukkan bahwa cendawan endofit yang terdapat pada tanaman cabai tersusun oleh singleton dan spesies dominan. Diantara 23 morfospesies terdapat 8 morfospesies yang bersifat singleton (C8, C10, C13, C16, C17, C18, C22,C23) dan C7 adalah spesies cendawan endofit yang dominan ditemukan (Gambar 5).
18
Gambar 5 Kelimpahan cendawan endofit pada tanaman cabai yang diisolasi pada tanaman bergejala dan tanaman tidak bergejala dari daerah endemis penyakit kuning.
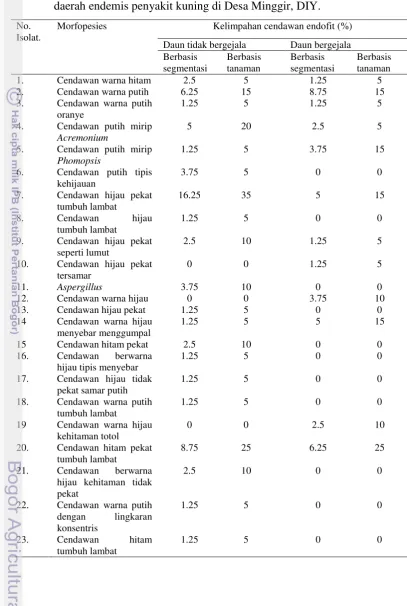
Analisis proporsi kelimpahan cendawan endofit dihitung berdasarkan frekuensi kolonisasi (FK) dan koefisien kemiripan (SC). Frekuensi kolonisasi dilakukan berbasis segmentasi dan tanaman (Tabel 2). Kelimpahan cendawan tertinggi adalah isolat C2 pada tanaman bergejala dan isolat C7 pada tanaman tidak bergejala. Terdapat 9 isolat morfospesies dengan kelimpahan berbeda yang ditemukan pada kedua kategori sampel. Isolat C7, C21, C4 dan C1 kelimpahannya lebih banyak pada tanaman tidak bergejala sedangkan isolat C2, C5, C9 dan C15 lebih banyak pada tanaman bergejala dan isolat C3 ditemukan di kedua kategori dengan frekuensi yang sama.
Tingkat kemiripan jenis spesies cendawan endofit antara tanaman tidak bergejala dan tanaman bergejala dianalisis berdasarkan indeks Sorensen. Nilai indeks Sorensen tergolong rendah (SC= 0,5625) yang artinya kemiripan komunitas spesies cendawan endofit yang ditemukan dari tanaman bergejala berbeda dengan yang ditemukan pada tanaman tidak bergejala.
Tabel 2. Kelimpahan spesies cendawan endofit berbasis segmentasi dan tanaman hasil isolasi dari daun tidak bergejala dan bergejala yang berasal dari daerah endemis penyakit kuning di Desa Minggir, DIY.
No. Isolat.
Morfopesies Kelimpahan cendawan endofit (%)
20
Patogenisitas Cendawan Endofit
Berdasarkan hasil isolasi, sebanyak 17 dari 23 isolat morfospesies berhasil ditumbuhkan pada media PDA 100%. Setelah uji patogenisitas, isolat-isolat tersebut dapat dibedakan menjadi dua kategori yaitu isolat berpotensi patogenik (13 isolat) dan berpotensi non patogenik (4 isolat) (Tabel 3). Isolat-isolat yang berpotensi patogenik ditandai dengan pengaruh negatifnya terhadap pengamatan daya kecambah benih, pertumbuhan akar dan batang. Isolat non patogenik berasal dari sampel daun tidak bergejala (isolat C1, C23, C8) dan dari sampel daun bergejala (isolat C21).
Tabel 3. Pengaruh Cendawan Endofit terhadap Daya Kecambah Benih, Panjang Akar dan Tajuk Bibit Cabai
Tabel 4. Identifikasi cendawan endofit potensial berdasarkan karakter morfologi
membentuk hifa aerial, terdapat jari-jari radial pada koloni, media tidak berubah warna,
membentuk hifa aerial, terdapat jari-jari radial pada koloni, media tidak berubah warna,
aerial kasar, miselium tumbuh keatas, media tumbuh berubah
menjadi hitam, tekstur
permukaan kasar, karakter tepi koloni rata.
Curvularia sp Tidak dilakukan analisis molekuler
C21 Warna koloni hitam,
pertumbuhan lambat, tidak
membentuk hifa aerial, media tidak berubah warna, tekstur permukaan kasar, karakter tepi koloni rata.
Hifa steril Dothideomycete sp
22
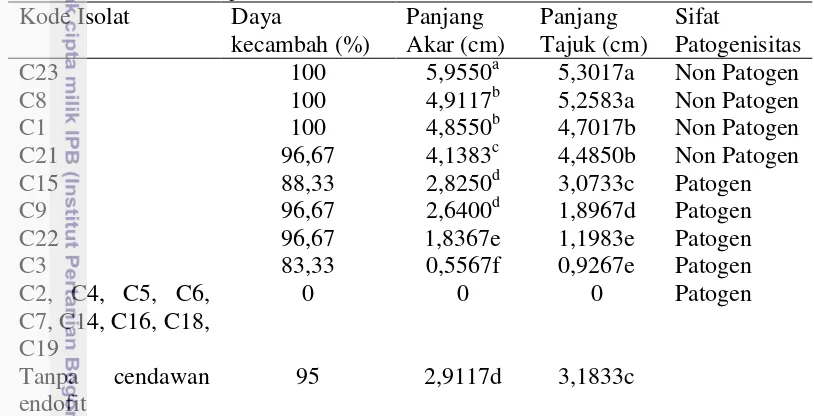
Pengaruh Cendawan Endofit terhadap Pertumbuhan Tanaman Cabai
Lima cendawan endofit yang diuji memperlihatkan potensinya dalam meningkatkan tinggi tanaman terutama pada minggu keempat setelah tanam dengan G. mangiferae dan Pleosporaceae menunjukkan potensi tertinggi sebagai agens pertumbuhan tanaman (Gambar 6A). Lebih lanjut perlakuan G. mangiferae menyebabkan bobot tajuk lebih tinggi dibandingkan dengan perlakuan lainnya, sedangkan C. nicotianae memiliki potensi menginduksi pertumbuhan di perakaran (Gambar 6B).
Gambar 6. Pengaruh cendawan endofit terhadap tinggi tanaman (A) dan bobot tajuk, bobot akar, volume akar pada tanaman cabai (B).
Pengaruh Cendawan Endofit terhadap Infeksi Virus Kuning Cabai
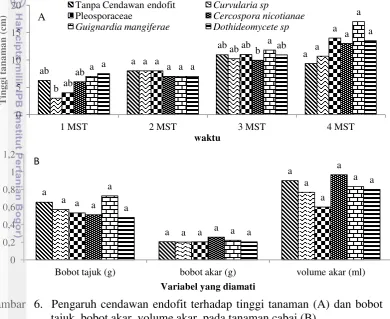
Diantara 5 cendawan endofit yang diuji, C. nicotiana, Pleosporaceae dan Dothideomycete sp. memperlihatkan potensinya dalam menekan kejadian penyakit terutama pada minggu pertama setelah inokulasi PepYLCV. Potensi Pleosporaceae dan C. nicotianae bahkan terlihat sampai 4 minggu setelah inokulasi dimana keduanya menyebabkan kejadian penyakit lebih rendah dibandingkan dengan perlakuan tanpa cendawan endofit (kontrol) (Gambar 7A). Lebih lanjut kedua cendawan tersebut menyebabkan keparahan penyakit yang lebih rendah dibandingkan dengan perlakuan tanpa cendawan endofit (Gambar 7B) dan Pleosporaceae memiliki laju infeksi yang lebih rendah dibandingkan kontrol (Gambar 7C). Kemampuan cendawan endofit menekan kejadian penyakit, keparahan penyakit dan laju infeksi virus daun keriting kuning cabai menunjukkan potensinya sebagai agens biokontrol.
ab a
Tanpa Cendawan endofit Curvularia sp
Gambar 7. Pengaruh cendawan endofit terhadap kejadian penyakit (A), keparahan penyakit (B) dan laju infeksi (C) penyakit kuning cabai
Selain kejadian penyakit dan intensitas penyakit, parameter keberhasilan aplikasi cendawan endofit terhadap PepYLCV diukur pula berdasarkan indikator pertumbuhan tanaman yang meliputi tinggi tanaman, bobot tajuk, bobot akar dan volume akar. Berdasarkan pengamatan tinggi tanaman, cendawan C. nicotianae dan G. mangifera memberikan respon positif terhadap pertumbuhan dibandingkan dengan cendawan yang lain maupun kontrol (Gambar 8). Dengan demikian G. mangiferae memiliki potensi yang tinggi sebagai agens penginduksi pertumbuhan baik pada kondisi tanaman terinfeksi virus maupun tanaman sehat.
a a
24
Gambar 8. Pengaruh cendawan endofit terhadap tinggi tanaman (A) dan bobot tajuk, bobot akar, volume akar (B) pada tanaman cabai yang diinokulasi PepYLCV.
Pembahasan
Penyakit kuning cabai dilaporkan sebagai penyakit utama di beberapa daerah penanaman cabai di Jawa dan Sumatera (Hikmat 2005, Sulandari et al. 2006). Hidayat et al. (1999) melaporkan bahwa isolat Begomovirus yang menginfeksi tanaman cabai di Cugenang, Jawa Barat memiliki kesamaan berdasarkan pola enzim restriksi dengan isolat Baranangsiang, Jawa Barat, tetapi berbeda dengan isolat Segunung, Jawa Barat. Sulandari et al. (2006) melaporkan bahwa strain Begomovirus yang menginfeksi tanaman cabai di Yogyakarta, Lembang, dan Cugenang memiliki kesamaan berdasarkan pola enzim restriksi, tetapi terdapat perbedaan dengan isolat asal Segunung. Lebih lanjut diketahui bahwa isolat Segunung yang dilaporkan Sulandari et al. (2006) berbeda dengan
Tanpa Cendawan endofit Curvularia sp
Pleosporaceae Cercospora nicotianae
Guignardia mangiferae Dothideomycete sp B
isolat Hidayat et al. (1999). Keragaman gejala dan strain Begomovirus yang menginfeksi tanaman cabai juga dilaporkan di Sumatera Barat oleh Trisno et al. (2009).
Hasil deteksi menggunakan primer universal Geminivirus menunjukkan bahwa seluruh sampel daun yang menunjukkan gejala dan satu daun tidak bergejala positif terinfeksi PepYLCV. Infeksi PepYLCV yang terdeteksi dari sampel daun tidak bergejala menunjukkan sifat infeksi virus dan respon tanaman inang. Matthews (1992) menguraikan gejala yang muncul pada tanaman yang terinfeksi virus dipengaruhi oleh konsentrasi virus, faktor lingkungan dan genetik tanaman. Tanaman yang terinfeksi pada awal masa pertumbuhan cenderung mengalami kerusakan lebih besar dibandingkan dengan tanaman yang terinfeksi setelah fase generatif.
Salah satu tujuan akhir dari penelitian ini adalah untuk mendapatkan kandidat agens biokontrol untuk penyakit kuning cabai. Mikrob yang mempunyai peranan dalam perlindungan tanaman pada umumnya diisolasi dari tanaman yang sehat diantara populasi tanaman yang terserang oleh patogen. Tanaman yang sehat diantara tanaman sakit diduga memiliki mekanisme pertahanan terhadap penyakit sehingga memiliki peluang besar untuk mendapatkan kandidat agens biokontrol. Peluang untuk memperoleh kandidat agens biokontrol juga dipengaruhi oleh cara budidaya tanaman. Rendahnya penggunaan pestisida kimia akan memberikan pengaruh positif terhadap mikrob yang bermanfaat terhadap tanaman sehingga peluang memperoleh agens biokontrol akan meningkat. Saikkonen et al. (2000) menyatakan cendawan endofit lebih banyak ditemukan pada rumput liar yang tumbuh secara alami dibandingkan pada rumput yang dibudidayakan. Oleh karena itu eksplorasi cendawan endofit untuk mencari kandidat agens biokontrol terhadap penyakit kuning cabai dilakukan pada lahan pertanaman cabai dengan sedikit aplikasi pestisida kimia dan tidak disiangi sehingga gulma tumbuh subur.
26
cendawan endofit yang tersedia di Genbank masih pada taraf taksa kelas cendawan bahkan masih berupa kode-kode isolat.
Konsep keragaman merupakan hal yang sederhana dan jelas, dimana terkandung dua faktor yang menjadi bahan pertimbangan utama yaitu kekayaan jenis (species richness) yang menunjukkan jumlah spesies dan kenetralan (evenness) yang kadang kala dikenal sebagai equitability, yang menunjukkan bagaimana spesies yang sama melimpah (Maguran 1988). Kekayaan dan kelimpahan spesies cendawan endofit sangat ditentukan oleh metode isolasi, letak geografis, asal tanaman dan bagian jaringan yang diisolasi (Hyde dan Soytong 2008). Berdasarkan variasi geoklimat dilaporkan bahwa jenis endofit lebih banyak diperoleh pada daerah tropis dibandingkan daerah zona iklim sedang atau dingin; sedangkan berdasarkan umur tanaman diketahui bahwa tanaman yang tua memiliki lebih banyak endofit dibandingkan tanaman muda. Penelitian Arnold dan Lutzoni (2007) menunjukkan bahwa kejadian infeksi endofit menurun secara linear dari daerah tropis ke arktik.
Hasil uji patogenisitas pada biji cabai menunjukkan bahwa isolat yang diperoleh lebih banyak yang berpotensi sebagai patogen. Patogen yang melakukan infeksi laten juga merupakan mikrob endofit karena berdasarkan teori evolusinya, endofit berasal dari patogen berkembang menjadi patogen minor dan kemudian menjadi simbion yang bersifat mutualisme (Schulz dan Boyle 2005). Namun pada umumnya istilah endofit lebih digunakan pada mikrob yang bermanfaat.
Kajian tentang pengaruh suatu mikroorganisme terhadap pertumbuhan tanaman telah dilaporkan pada berbagai jenis tanaman, baik pada kondisi rumah kaca dalam media organik maupun di lapangan. Mikroorganisme yang berperan
meningkatkan pertumbuhan tanaman disebut sebagai PGPR (plant growth
promoting rhizobacteria) dengan bakteri rizosfer sebagai agens penginduksi atau
PGPF (plant growth promoting fungi) dengan cendawan sebagai agens
penginduksi. Mekanisme peningkatkan pertumbuhan tanaman oleh
mikroorganisme tersebut dapat berupa proteksi terhadap infeksi akar oleh patogen minor dan mayor, meningkatkan ketersediaan nutrisi ke tanaman inang, menurunkan tingkat etilen pada inang atau peningkatan produksi senyawa stimulasi, seperti zat pengatur tumbuh (Gravel et al. 2007). Senyawa yang umum dihasilkan oleh mikroorganisme seperti asam indol-3-asetat (IAA), siderophores dan 1–aminocyclopropane-1-karboksilat (ACC) deaminase, menstimulasi jalur metabolik tertentu seperti fiksasi nitrogen dan fosfat serapan, mempengaruhi mobilitas logam dan ketersediaannya untuk tanaman (Chen 2010).
Selain PGPR dan PGPF terdapat organisme menguntungkan lainnya yang disebut plant growth promoting endophytes (PGPE) (Chen 2010). Kajian peranan endofit dibidang pertanian sudah banyak dilakukan. Bakteri endofit dapat meningkatkan ketersediaan fosfat untuk Solanum nigrum L dan meningkatkan biomasa tanaman (Chen et al. 2010). Lebih lanjut Zabalgogeazcoa et al.
(2006) menyatakan bahwa konsentrasi Fosfor mempunyai hubungan yang jelas dengan keberadaan endofit. Tanaman yang berasosiasi dengan endofit memiliki P yang lebih besar dibandingkan tanpa endofit. Pada tanaman F. rubra tidak ada efek nyata antara perlakuan endofit dengan produksi biomasa, tinggi tanaman, atau jumlah batang yang diamati. Satu dari dua genotipe N. coenophialum menyebabkan biomasa tanaman yang lebih tinggi ketika diaplikasikan pada kultivar F. arundinacea. Efek endofit terkait dengan faktor lingkungan, pada tanaman Fescue peningkatan konsentrasi P pada tanaman terinfeksi hanya terjadi
bila P tersedia dalam tanah rendah (Malinowsky et al. 1998b dalam
Zabalgogeazcoa et al. 2006).
Pada penelitian ini terlihat bahwa empat isolat cendawan endofit memberikan respon pertumbuhan positif dibandingkan dengan kontrol (tanpa aplikasi cendawan endofit), bahkan G. mangiferae masih tetap menginduksi pertumbuhan ketika terdapat infeksi PepYLCV. Kemampuan cendawan endofit dalam meningkatkan pertumbuhan tanaman ini diduga adanya metabolit seluler yang dihasilkan cendawan endofit yang berperan sebagai induksi pertumbuhan tanaman.
Selain sebagai agens pemicu pertumbuhan tanaman cendawan endofit dapat berperan sebagai agens biokontrol (Gaziz et al. 2010). Cendawan endofit telah banyak dimanfaatkan untuk mengendalikan cendawan (Gazis et al. 2010), nematoda (Chaves et al. 2009), dan insekta (Rranzluebbers et al. 1999). Kajian endofit sebagai agens biokontrol terhadap virus belum banyak dipublikasikan secara ilmiah. Beberapa penelitian sudah dilakukan namun tidak dititik beratkan pada virus melainkan pada serangga vektornya. Latch et al. (1985) dalam Smales (1996) menemukan tanaman Fescue (Festuca arundinacea Schreber) yang
terinfeksi cendawan endofit Acremonium coenophialum terhindar dari
Rhopalosiphum padi L. (Hemiptera: Aphididae) yang merupakan vektor Barley yellow dwarf virus (BYDV) dan berpengaruh terhadap penurunan kejadian BYDV. Hasil penelitian Lehtonen et al. (2006) menunjukkan bahwa pada tanaman Meadow ryegrass (Lolium pratense) yang terinfeksi oleh BYDV, aplikasi cendawan endofit secara nyata mampu mengurangi frekuensi tanaman yang terinfeksi. Penurunan frekuensi tanaman yang terinfeksi oleh BYDV berkorelasi positif terhadap penurunan populasi kutudaun pada M. ryegrass.
28
Dilaporkan sebelumnya, bahwa cendawan endofit adalah salah satu agens penginduksi ketahanan tanaman. Mekanisme induksi ketahanan tanaman yang sudah diketahui dapat dibedakan menjadi dua yaitu Induced systemic resistance (ISR) dan Systemic Acquired Resistace (SAR). ISR adalah induksi ketahanan pada tanaman oleh mikrob non patogenik yang melibatkan signal asam jasmonik dan etilen sebagai senyawa penginduksi, tidak melibatkan ekspresi gen PR (pathogenesis related genes) tetapi gen lain yang belum diketahui. SAR adalah induksi ketahanan tanaman yang dipicu oleh infeksi mikrob patogenik, aplikasi bahan kimia yang melibatkan signal asam salisilat dan ekspresi gen PR (Bakker et al. 2003).
Dilaporkan oleh Gao et al. (2010) bahwa Fusarium solani yang diisolasi dari jaringan akar tomat merupakan agens ISR terhadap Septoria lycopersici yaitu dengan mekanisme menstimulasi gen PR5 dan PR7 yang terekspresi di akar. Lebih lanjut Colletotrichum magna mutan non patogenik dapat melindungi tanaman Citrullus lanatus dan Cucumis sativus dari patogen C. orbiculare dan F oxysporum dengan menunjukkan aktifitas peroxidase dan pada level yang tinggi. Gao et al. (2010) menyatakan bahwa mekanisme cendawan endofit adalah ISR yang melibatkan signal gen PR.
V. SIMPULAN DAN SARAN
Simpulan
Sebanyak 23 morfospesies cendawan endofit berhasil diperoleh dari sampel daun cabai yang menunjukkan gejala dan yang tidak menunjukkan gejala. Jumlah cendawan endofit dari tanaman yang tidak bergejala cenderung lebih tinggi (19 morfospesies) dibandingkan dari tanaman yang bergejala (13 morfospesies). Berdasarkan analisis koofisien kemiripan dengan indek Sorensen terdapat perbedaan komunitas spesies endofit diantara keduanya dengan nilai indeks Sorensen 0,5625. Berdasarkan uji patogenisitas, cendawan endofit yang diisolasi lebih banyak yang berpotensi sebagai patogen (13 isolat) dibandingkan yang bersifat non patogenik (4 isolat). Diantara 4 isolat yang bersifat non patogenik, 3 isolat diperoleh dari sampel daun tidak bergejala. Identifikasi yang dilakukan menunjukkan bahwa 3 isolat merupakan hifa steril dan 1 isolat adalah Curvularia sp.
Beberapa isolat cendawan endofit yang diisolasi dari pertanaman cabai di daerah endemis penyakit kuning cabai memiliki potensi sebagai agens pemicu pertumbuhan dan agens biokontrol terhadap penyakit kuning. Isolat-isolat C. nicotianae, Pleosporaceae, Guignardia mangifera dan Dothideomycete sp. memiliki potensi sebagai agens pemicu pertumbuhan tanaman dan 2 isolat diantaranya (C. nicotianae dan Pleosporaceae) dapat menjadi kandidat agens biokontrol untuk penyakit kuning karena mampu menekan kejadian penyakit, laju infeksi dan keparahan penyakit.
Saran
30
DAFTAR PUSTAKA
Agrios GN. 2005. Plant Pathology. Ed 5. New York (US): Academic Pr.
Aidawati N. 2006. Keanekaragaman Begomovirus pada tomat dan serangga vektornya, Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae), serta pengujian ketahanan genotipe tomat terhadap strain Begomovirus. [disertasi]. Bogor (ID) : Fakultas Pertanian, Institut Pertanian Bogor.
Arnold AE and Lutzoni F. 2007. Diversity and host range of foliar fungal endophytes: are tropical leaves biodiversity hotspots?. Ecology. 88(3):541-549.
Arteca RN. 1996. Plant growth substances: Principles and Applications. Chapman and Hall. New York. Pp 332.
Badan Litbang Pertanian. 2011. Cara menghindar dari serangan virus cabai. Sinar Tani : Agroinovasi. Edisi 2-8 Pebruari 2011 No.3390 Tahun XLI
Chen et al. 2010. Application of plant growth promoting endophytes (PGPE) isolated from solanum nigrum L. for phytoextraction of Cd-polluted soils. Applied Soil Ecology 461:383-389.
Chave NP, Pocosangre LE, Elango F, Rosales FE, Sikora R. 2009. Combining
endophytic fungi and bacteria for the biocontrol of Radopholus
similis.Scientia Horticulturae. 122:472-478.
Direktorat Jenderal Hortikultura. 2010. Perkembangan luas panen hortikultur tahun 2000-2009. [terhubung berkala]. http://www.deptan.go.id.[Desember 2010]
Doyle JJ, Doyle JL. 1990. Isolation of Plant DNA from fresh tissue. Focus. 12:13-15.
FE Vega, F Posada, MC Aime, M Rava-Ripoll, F Infante, SA Rehner. 2008. Entomopathogenic fungal endophytes. Biological Control 46: 72-82. Doi. 10. 1016/j.biokontrol.2008.01.008.
Ganefianti DW, Sujiprihati S, Hidayat SH, Syukur M. 2008. Metode penularan
dan uji ketahanan genotipe cabai (Capsicum spp.) terhadap
Begomovirus. J Akta Agros . 11(2):162-169.