KELAPA SAWIT (
Elaeis guineensis
)
DEVI LIA WATI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ii
ABSTRAK
DEVI LIA WATI. Aktivitas Terbang Harian dan Mencari Polen Trigona laeviceps Smith di Perkebunan Karet (Hevea braziliensis) dan Kelapa Sawit (Elaeis guineensis). Dibimbing oleh
RIKA RAFFIUDIN dan TRI ATMOWIDI.
Konversi hutan menjadi lahan perkebunan berpengaruh pada kestabilan ekosistem yang meliputi jaring makanan dan faktor lingkungan. Perilaku Trigona atau stingless bee dapat digunakan sebagai model untuk memahami konversi lahan tersebut. Penelitian ini bertujuan untuk mempelajari aktivitas terbang harian Trigona laeviceps dan aktivitas mencari polen di perkebunan kelapa sawit dan karet. Pengamatan aktivitas terbang harian dan aktivitas mencari polen T. laeviceps dilakukan selama 26 hari di kedua perkebunan. Pengamatan dilakukan dari pukul 05.00-18.10 dan dikelompokkan menjadi tiga kelompok yaitu pengamatan pagi, siang, sore. Korelasi antara lingkungan (suhu, kelembaban, intensitas cahaya) dengan aktivitas T. laeviceps yang diamati dianalisis dengan Principle Component Analysis dan Korelasi Pearson. Hasil penelitian ini menunjukkan aktivitas T. laeviceps mencari polen di perkebunan kelapa sawit terjadi mulai pukul 08.00 dengan puncak aktivitas mencari polen pukul 12.00-12.10 sedangkan di perkebunan karet terjadi mulai pukul 07.30 dengan puncak aktivitas mencari polen pukul 12.30-14.10. Aktivitas terbang harian dan mencari polen T. laeviceps pada setiap koloni, lebih tinggi pada siang hari (11.00-14.10) dibandingkan aktivitas pada pagi maupun sore hari. Intensitas cahaya dan suhu berkorelasi positif terhadap aktivitas terbang harian dan mencari polen T. laeviceps, sedangkan kelembaban berkorelasi negatif. Korelasi Pearson yang signifikan terjadi antara faktor lingkungan (suhu, kelembaban dan intensitas cahaya) dengan aktivitas terbang harian dan mencari polen T.laeviceps, sedangkan korelasi tidak signifikan terdapat antara suhu dengan aktivitas mencari polen di perkebunan sawit.
Kata kunci: Perkebunan karet, Perkebunan kelapa sawit, Trigona (Tetragonula) laeviceps, aktivitas terbang harian, aktivitas terbang mencari polen
ABSTRACT
DEVI LIA WATI. Daily Flight Activity and Pollen Collection of Trigona laeviceps Smith in Rubber (Hevea braziliensis) and Oil Palm (Elaeis guineensis) Plantations. Supervised by RIKA RAFFIUDIN and TRI ATMOWIDI.
Conversion of forest to plantations affect the stability of ecosystems i.e. food webs and environmental factors. Stingless bee Trigona can be used as a model to understand the land conversion effect. This research were aimed to study daily flight activity of Trigona laeviceps and forage for pollen at oil palm and rubber plantations. Observation of daily flight activity and pollen collection activity of T. laeviceps were conducted in 26 days at oil palm and rubber plantations. The behaviour observations were taken from 05:00 to 18:10 and the observations were clustered into three groups, i.e. morning, noon and afternoon groups. The correlation of environmental effects (temperature, humidity, light intensity) activity to T. laeviceps behaviour were analysed by using Principle Component Analysis and Pearson correlation. The results of this study showed that pollen collection activity of T. laeviceps in oil palm plantations were commenced from 08.00 having peak activity at 12:00 to 12:10 while from 07.30 in pollen collection (peak activity at 12:30 to 14:10) for T. laeviceps in rubber plantation. Daily flight activity and pollen collecting T. laeviceps on each colony, was higher during noon (11:00 to 14:10) compared to morning and afternoon activities. Significant Pearson correlations were observed between environmental factors (temperature, humidity and light intensity) with daily flying activities and pollen collection. However, non-significant correlation occured between temperature and pollen collection activity in oil palm plantation.
AKTIVITAS TERBANG HARIAN DAN MENCARI POLEN
Trigona
laeviceps
Smith DI PERKEBUNAN KARET (
Hevea braziliensis
) DAN
KELAPA SAWIT (
Elaeis guineensis
)
DEVI LIA WATI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
iv
Judul Skripsi : Aktivitas Terbang Harian dan Mencari Polen Trigona laeviceps Smith di Perkebunan karet (Hevea braziliensis) dan Kelapa Sawit (Elaeis guineensis) Nama : Devi Lia Wati
NRP : G34080061
Disetujui:
Pembimbing I Pembimbing II
Dr. Ir. Rika Raffiudin, M.Si NIP. 19670617 199203 2 001
Dr. Tri Atmowidi, M.Si NIP. 19670827 199303 1 003
Diketahui, Ketua Departemen Biologi
Dr. Ir. Ence Darmo Jaya Supena, M.Si NIP. 19641002 198903 1 002
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala rahmat dan karunia-Nya, sehingga karya ilmiah yang berjudul “Aktivitas Terbang Harian dan Mencari Polen Trigona laeviceps Smith di Perkebunan Karet (Hevea braziliensis) dan Kelapa Sawit (Elaeis guineensis)” dapat diselesaikan. Penelitian ini dilaksanakan mulai bulan April sampai September 2012, di perkebunan kelapa sawit Perseroan Terbatas Perkebunan Nusantara (PTPN) VIII, Afdeling (AFD) IV Toge, Kebun Cikasungka, Kecamatan Cigudeg dan di perkebunan karet Balai Penelitian Bioteknologi Perkebunan Indonesia (BPBPI), Desa Ciaruteun ilir, Kecamatan Cibungbulang. Identifikasi Trigona dilakukan di bagian Ekologi dan Sistematika Hewan, Departemen Biologi, FMIPA, Institut Pertanian Bogor.
Terimakasih penulis ucapkan kepada Dr. Ir. Rika Raffiudin, M.Si dan Dr. Tri Atmowidi, M.Si sebagai pembimbing dan Dr. Ir. Muhadiono, M.Sc. sebagai penguji atas segala bimbingan, arahan, dan ilmu yang diberikan. Ucapan terimakasih tak terhingga penulis sampaikan kepada keluarga tercinta Bapak (Ruspendi), ibu (Sri), adik (Dena) yang telah memberikan doa, kasih sayang, semangat serta dukungan berupa moril maupun materil, suami tercinta Fery Juliari Saputra atas kasih sayang, kesabaran dan dukungan kepada penulis selama menyelesaikan penelitian ini, serta kepada putra tersayang Azka Nur Alamsyah yang telah memberikan semangat kepada penulis selama proses penelitian sampai selesai. Seluruh staf PTPN, staf BPBPI, Keluarga bapak Juhro, ibu Etih dan mba Lastri yang telah membantu kelancaran penelitian dilapangan. Ucapan terima kasih penulis sampaikan kepada seluruh keluarga Zoologi dan teman-teman Biologi angkatan 45 atas dukungan dan semangat yang telah diberikan selama proses penelitian.
Penulis berharap semoga karya tulis ini dapat bermanfaat dalam perkembangan ilmu pengetahuan baik bagi penulis maupun pembaca.
Bogor, Desember 2012
vi
RIWAYAT HIDUP
Penulis dilahirkan di Ciamis provinsi Jawa Barat pada tanggal 7 Juni 1990 dari pasangan Bapak Ruspendi dan ibu Sri Mulyawati. Penulis merupakan anak pertama dari dua bersaudara.
Penulis lulus dari SD Negeri Karang Kendal pada tahun 2002, lalu SMP Negeri 1 Baregbeg pada tahun 2005. Penulis melanjutkan pendidikan di SMA Negeri 2 Ciamis dan lulus tahun 2008. Kemudian pada tahun yang sama penulis diterima di jurusan Biologi Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB (USMI).
Penulis aktif sebagai anggota Gentra Kaheman IPB pada tahun 2009, anggota Bina Desa BEM KM IPB pada tahun 2010-2011 pada divisi PRF (Public Relation and Fund Rising). Penulis juga aktif dalam beberapa kepanitiaan kegiatan kampus seperti Biologi Interaktif, MPD Biologi, dan pernah mengikuti pagelaran seni sunda di Gunung Walat Sukabumi. Penulis telah melakukan studi lapang di Taman Wisata dan Cagar Alam Pangandaran pada tahun 2010 mengenai “Lumut Epifil di Cagar Alam Pangandaran, Kab. Ciamis, Jawa Barat”. Pada tahun 2011, penulis melakukan praktik lapangan di Pusat Penelitian dan Pengembangan Hasil Hutan (P3HH) mengenai
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... viii
PENDAHULUAN... 1
Latar Belakang ... 1
Tujuan ... 1
BAHAN DAN METODE ... 1
Waktu dan Tempat ... 1
Koleksi dan Identifikasi T. laeviceps ... 2
Pemeliharaan Koloni T. laeviceps ... 2
Pengamatan Aktivitas Terbang Harian T. laeviceps ... 3
Pengamatan Karakter Lingkungan Sekitar ... 3
Pengamatan Struktur Sarang T. laeviceps ... 3
Analisis Data ... 3
HASIL ... 3
Identifikasi Trigona ... 3
Aktivitas Terbang Harian T. laeviceps di Perkebunan Sawit (SW) ... 4
Aktivitas Mencari Polen T. laeviceps di Perkebunan Sawit (SW) ... 4
Aktivitas Terbang Harian T. laeviceps di Perkebunan Karet (KR) ... 4
Aktivitas Mencari Polen T. laeviceps di Perkebunan Karet (KR) ... 6
Perbandingan Aktivitas Terbang Harian T. laeviceps di SW dengan KR ... 6
Perbandingan Aktivitas Mencari Polen T. laeviceps di SW dengan KR ... 8
Persentase Aktivitas Mencari Polen T. laeviceps di SW dan KR... 8
Hubungan Aktivitas Terbang T. laeviceps dengan Faktor Lingkungan ... 12
Pengamatan Struktur Sarang T.laeviceps ... 12
PEMBAHASAN ... 13
SIMPULAN ... 14
SARAN ... 15
DAFTAR PUSTAKA ... 15
viii
DAFTAR TABEL
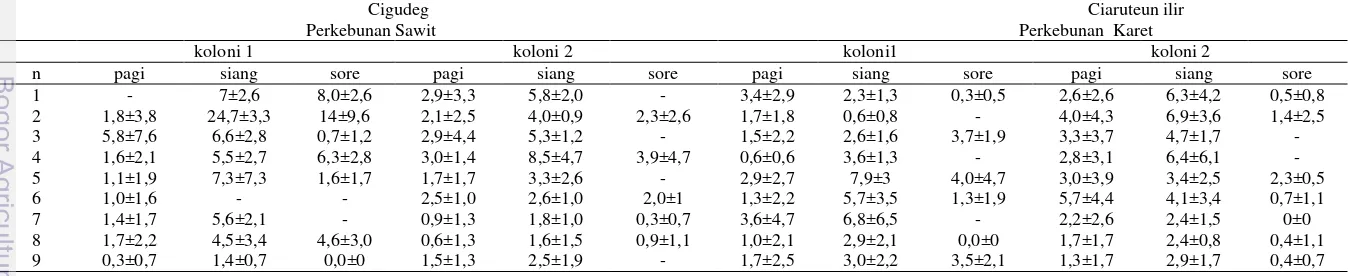
1 Jumlah Trigona laeviceps kembali ke sarang pada pagi hari (05:00-10:40), siang hari
(11:00-14:40) dan sore hari (15:00-18:10) ... 9
2 Jumlah Trigona laeviceps yang kembali ke sarang membawa polen pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10) ... 9
3 Rata-rata suhu udara (oC) di sekitar sarang pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10) ... 10
4 Rata-rata kelembaban udara (%) di sekitar sarang pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10) ... 10
5 Rata-rata intensitas cahaya (lux) di sekitar sarang pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10) ... 11
6 Nilai korelasi antara komponen lingkungan dengan jumlah T. laeviceps masuk sarang dan jumlah T. laeviceps bawa polen di perkebunan sawit ... 13
7 Nilai korelasi antara komponen lingkungan dengan jumlah T. laeviceps masuk sarang dan jumlah T. laeviceps bawa polen di perkebunan karet ... 13
DAFTAR GAMBAR
1 Lebah ratu Trigona laeviceps dan lebah pekerja diantara sel anakan...
12 Pemeliharaan koloni T. laeviceps ... 2
3 Struktur morfologi Trigona laeviceps ... 4
4 Identifikasi Trigona (Tetragonula) laeviceps ... 5
5 Aktivitas terbang harian dan mencari polen T. laeviceps tiap koloni di SW ... 6
6 Aktivitas terbang harian dan mencari polen T. laeviceps tiap koloni di KR ... 7
7 Perbandingan jumlah T. laeviceps masuk sarang di perkebunan sawit dengan perkebunan karet ... 7
8 Perbandingan jumlah T. laeviceps mencari polen di perkebunan sawit dengan perkebunan karet ... 8
9 Persentase jumlah aktivitas mencari polen (hitam) terhadap aktivitas terbang harian (putih) ... 8
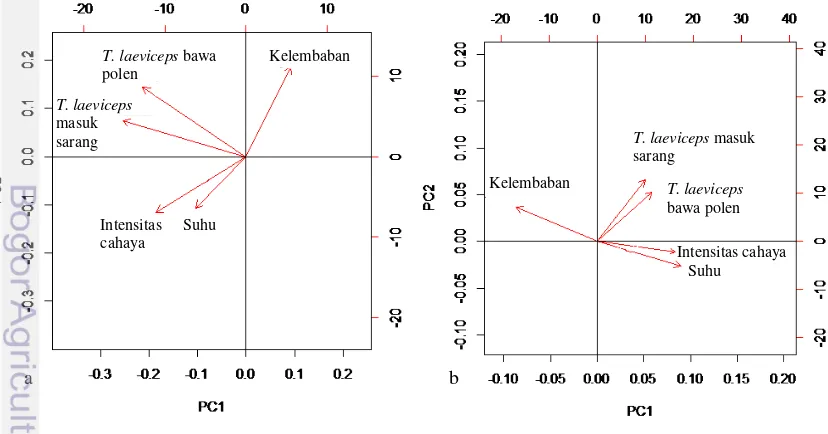
10 Hasil analisis Principle Component Analysis (PCA) hubungan aktivitas harian T. laeviceps dengan faktor lingkungan ... 12
11 Susunan sarang pada koloni 1 T. laeviceps di perkebunan sawit ... 13
DAFTAR LAMPIRAN
1 Peta lokasi perkebunan sawit di kecamatan Cigudeg, Kabupaten Bogor. ... 182 Peta lokasi perkebuna karet di kecamatan Cibungbulang, Kabupaten Bogor ... 18
PENDAHULUAN Latar Belakang
Keanekaragaman hayati di Indonesia saat ini mengalami penurunan akibat adanya pembukaan lahan hutan yang dikonversi menjadi lahan perkebunan (Sunderline & Resosudarmo 1996). Konversi lahan yang dilakukan akan berpengaruh pada kestabilan ekosistem baru (Shahabuddin 2003). Pendekatan yang dapat digunakan untuk memahami dampak perubahan lingkungan adalah melalui identifikasi jenis dan perkembangan serangga. Serangga merupakan bioindikator yang responsif untuk menilai perubahan yang terjadi dalam suatu habitat (Speight et al. 1999). Serangga yang digunakan adalah lebah madu Trigona (Hymenoptera: Apidae, Meliponini) (Michener 1974).
Lebah Trigona merupakan serangga sosial tingkat tinggi yang hidup dalam suatu koloni dan termasuk golongan stingless bee yaitu kelompok lebah yang tidak bersengat. Karakter utama serangga sosial tingkat tinggi antara lain terdapat pembagian tugas yang jelas pada masing-masing kasta dan adanya komunikasi diantara anggota koloni mengenai letak pakan. Koloni Trigona terdiri atas kasta reproduktif (ratu, jantan) dan non-reproduktif (pekerja) (Michener 1974) (Gambar 1). Trigona tersebar di daerah tropis dan sub tropis di Amerika Selatan, setengah bagian Afrika Selatan dan Asia Tenggara. Koloni Trigona terdiri atas 300-80.000 individu lebah (Free 1982).
Trigona termasuk serangga yang mengalami metamorfosis sempurna. Setiap individu mempunyai siklus hidup mulai dari telur, larva, pupa dan imago. Telur Trigona memiliki struktur yang lunak, berukuran kecil memanjang. Larva berwarna keputih-putihan dan memakan makanan yang tersedia di dalam sel. Kulit pupa relatif lunak, anggota badannya sudah membentuk lipatan dan memiliki sayap (Michener 1974). Menurut Lisboa et al. (2005), terdapat perubahan warna mata pada pupa Trigona yaitu putih, merah muda, dan hitam. Setelah beberapa hari pupa akan berganti kulit dan berubah menjadi imago (Michener 1974).
Trigona umumnya membuat sarang di lubang atau cabang pohon (Michener 2000). Sarang Trigona dibuat dengan mencampur lilin dan resin propolis dari tanaman. Sarang tersusun atas brood cells (sel pemeliharaan telur, larva, pupa), sel polen dan sel madu (Michener 1974).
Trigona adalah pencari pakan yang agresif (O’Toole & Raw 1991). Pakan Trigona berupa polen sebagai sumber protein dan nektar sebagai sumber karbohidrat. Trigona menyimpan polen di tungkai belakang dalam keranjang khusus yang disebut corbicula (Shuel 1992). Trigona pada sarang alami di Bogor, mulai melakukan aktivitas terbang harian pukul 06.00 sampai pukul 18.00 atau selama 12 jam. Trigona banyak membawa polen pada pagi hari (07.00-10.00) dan sore hari (14.00-17.00. puncak aktivitas membawa polen terjadi pukul 08.06-08.30 dan pukul 14.36-15.40 (Nelli 2003).
Gambar 1. Lebah ratu Trigona laeviceps dan lebah pekerja diantara sel anakan
Tujuan
Penelitian ini bertujuan untuk mempelajari aktivitas terbang harian dan mencari polen Trigona laeviceps di perkebunan karet dan kelapa sawit.
BAHAN DAN METODE Waktu dan Tempat
Penelitian dilaksanakan di perkebunan sawit dari tanggal 23 April - 23 Mei 2012 dan di perkebunan karet dari tanggal 28 Mei - 24 Juni 2012. Penelitian dilakukan di perkebunan kelapa sawit Perseroan Terbatas Perkebunan Nusantara (PTPN) VIII, Afdeling (AFD) IV Toge, Kebun Cikasungka, Kecamatan Cigudeg (6°32'5.07" LS, 106°33'11.49" BT) dengan ketinggian 544 m dpl, Kabupaten Bogor (lampiran 1) dan di perkebunan karet Balai Penelitian Bioteknologi Perkebunan Indonesia, Desa Ciaruteun Ilir, Kecamatan Cibungbulang (6°37'31.17" LS, 106°40' 18.59" BT) dengan ketinggian 365 m dpl, Kabupaten Bogor (lampiran 2). Identifikasi T. laeviceps dilakukan di bagian Ekologi dan Sistematika Hewan, Departemen Biologi, FMIPA, Institut Pertanian Bogor.
2
Koleksi dan Identifikasi T. laeviceps T. laeviceps dikoleksi dari koloni yang terdapat di desa Bantarsari, kecamatan Rancabungur (6° 31' 27.01"LS & 106° 41' 39.84"BT) dengan ketinggian 141 m dpl, kabupaten Bogor (Gambar 2a). T. laeviceps ditangkap menggunakan jaring serangga dan diawetkan dalam larutan alkohol 70%. T. laeviceps dibedah dengan melepaskan sayap, tungkai, abdomen dan kepala. Identifikasi T. laeviceps dilakukan untuk mengetahui spesies dari genus Trigona yang dikoleksi. T. laeviceps diidentifikasi menggunakan kunci determinasi Borror et al. (1981) sampai tingkat famili dan Sakagami et al. (1990) pada tingkat spesies.
Pemeliharaan Koloni T. laeviceps
Ukuran koloni T. laeviceps sesuai dengan ukuran koloni yang diperoleh dari sarang alami (Gambar 2). Koloni T. laeviceps dipindahkan dari sarang alami yang berupa bambu ke kotak kayu berukuran 40,2 cm x 20,5 cm x 19,7 cm. Dinding bagian atas kotak dilapisi dengan kaca untuk memudahkan pengamatan (Gambar 2c). Sebelum pengamatan dimulai, koloni T. laeviceps diadaptasi selama 8 hari dengan tujuan agar T. laeviceps beradaptasi dengan kondisi lingkungan di dalam kotak yang baru. Koloni diletakkan pada ketinggian 1 m dari permukaan tanah (Gambar 2d). Penyangga dari kotak diolesi dengan vaselin atau oli supaya koloni T. laeviceps terhindar dari serangan predator, seperti semut dan cicak.
a b
c d
Pengamatan Aktivitas Terbang Harian T. laeviceps
Pengamatan perilaku aktivitas terbang harian T. laeviceps dilakukan pada koloni 1 dan koloni 2 yang diamati antara pukul 05.00 sampai pukul 18.00 WIB. Masing-masing diamati 13 hari selama 26 hari menggunakan metode ad libitum (Martin & Bateson 1993). Tiap pengamatan dilakukan selama 10 menit dengan interval 20 menit. Jumlah T. laeviceps yang masuk dikelompokan menjadi tiga kelompok yaitu pertama kelompok pagi (05:00-10:40), kedua kelompok siang (11:00-14:40) dan ketiga kelompok sore (15:00-18:10), setiap kelompok diamati 3 sampai 8 kali pengamatan tergantung dengan kondisi cuaca. Jumlah T. laeviceps yang masuk ke sarang dan yang masuk membawa polen dihitung menggunakan counter (modifikasi Pierrot & Schlindwein 2003).
Aktivitas T. laeviceps selama pengamatan direkam dengan handycam (Sony Digital HDD DCRSR80). Penggunaan bantuan sinar infra merah dari handycam dilakukan jika cahaya tidak cukup untuk pengamatan. Pengamatan dilakukan pada jarak 1 meter dari lubang sarang.
Pengamatan Karakter Lingkungan Sekitar
Faktor-faktor lingkungan seperti suhu, kelembaban dan intensitas cahaya diukur tiap kisaran satu jam. Pengamatan karakter lingkungan dilakukan menggunakan termometer untuk mengukur suhu, hygrometer untuk mengukur kelembaban dan luksmeter untuk mengukur intensitas cahaya,
Pengamatan Struktur Sarang T. laeviceps Pengamatan komponen struktur sarang dilakukan dalam masing-masing lokasi antar koloni 1 dan koloni 2 dan antar lokasi. Parameter uji t adalah jumlah T. laeviceps masuk sarang dan jumlah T. laeviceps membawa polen. Kedua parameter tersebut dibandingkan untuk masing-masing lokasi dan antar lokasi. Uji t independent yaitu masing-masing variabel berdiri sendiri tidak tergantung satu dengan lainnya (Walpole 1992).
Probabilitas uji Korelasi Pearson dilakukan pada setiap data yang merupakan hubungan antara aktivitas terbang harian T. laeviceps dengan faktor lingkungan (suhu, kelembaban, intensitas cahaya). Terdapat beberapa kriteria koefisien korelasi mengenai kekuatan hubungan linear antara dua variabel diantaranya 0 : Tidak ada korelasi antara dua variable; >0 – 0,25: Korelasi sangat lemah; >0,25 – 0,5: Korelasi cukup; >0,5 – 0,75: Korelasi kuat; >0,75 – 0,99: Korelasi sangat kuat; 1: Korelasi sempurna (Walpole 1992). Data juga dianalisis dengan metode Principle Component Analysis (PCA) menggunakan software R yang dapat diakses pada www.r-project.org. Parameter untuk korelasi Pearson (Walpole 1992) dan R adalah suhu, kelembaban, intensitas cahaya, jumlah T. laeviceps masuk sarang dan jumlah T. laeviceps bawa polen.
HASIL Identifikasi Trigona
4
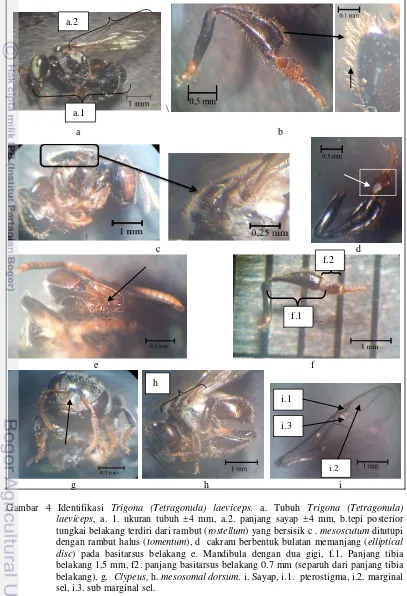
Gambar 3 Struktur morfologi Trigona laeviceps. a. flagella, b. pedicel, c. scape, d. mata, e. gena. f. mesoscutum,g. tegula, h. propodeum, i. mesoscutellum, j. sayap depan, k. metasoma, l. metasomal terga, m. tibia belakang, n.basitarsus belakang, o. tungkai belakang, p.tungkai tengah, q. tungkai depan r. mandibula, s. clypeus.
Aktivitas Terbang Harian T. laeviceps di Perkebunan Sawit
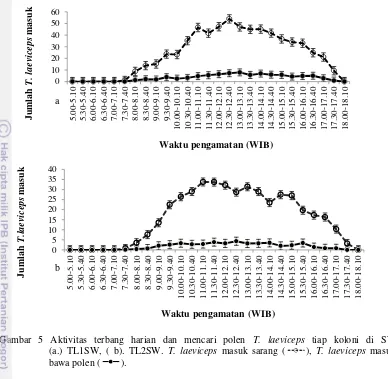
Semua koloni T. laeviceps di perkebunan sawit (SW) mulai melakukan aktivitas terbang pukul 07.30 pagi (suhu 25oC, kelembaban 85%, intensitas cahaya 140 lux) mempunyai puncak aktivitas terbang pada pukul 11.00-12.40 dengan jumlah T. laeviceps yang masuk sarang 47 individu/10 menit (Gambar 5a). Aktivitas terbang harian TL1SW lebih tinggi daripada koloni 2 T. laeviceps di perkebunan sawit (TL2SW). Koloni TL2SW mempunyai puncak aktivitas terbang harian pada pukul 11.00-13.10 dengan jumlah T. laeviceps masuk sarang 32 individu/10 menit (Gambar 5b).
Berdasarkan uji t-test tidak berpasangan aktivitas terbang harian koloni 1 dan koloni 2 menunjukan nilai t=1,759, nilai p=0,084, karena nilai p=0.084 > alpha=0.05 maka aktivitas terbang harian T. laeviceps pada koloni 1 tidak berbeda nyata dengan aktivitas terbang harian koloni 2. Pada koloni 1 dan 2, jumlah T. laeviceps yang kembali ke sarang lebih tinggi pada siang hari (pukul 11:00-14:40 WIB) dengan jumlah rata-rata masing-masing 45,4 individu (suhu 34oC, kelembaban 61,1%, intensitas cahaya 429 lux) dan 30,5 individu (Tabel 1) (suhu 31,7
T. laeviceps yang mencari polen terlihat membawa polen pada tungkai belakang, sedangkan T. laeviceps tanpa polen diduga membawa nektar, air atau resin. Koloni TL1SW mulai mencari polen pada pukul 08.00 dan berhenti pada pukul 17.40 (Gambar 5a). sedangkan koloni TL2SW mulai mencari polen pada pukul 08.30 berhenti pada pukul 17.10 (Gambar 6b). TL1SW mencari polen 30 menit lebih awal dan berhenti 30 menit lebih akhir daripada TL2SW. Puncak aktivitas mencari polen dan jumlah individu yang membawa polen tiap 10 menit pada TL1SW dan TL2SW berturut-turut adalah pukul 12.30-13.10 (8 individu) dan 11.30-12.40 (4 individu) (Gambar 5 a, b). Puncak aktivitas mencari polen pada TL1SW terjadi pada suhu 34oC, kelembaban 61,1%, intensitas cahaya 429 lux, sedangkan pada TL2SW terjadi pada suhu 31,7
o
C, kelembaban 63%, dan intensitas cahaya 409 lux (Tabel 3, 4, 5).
Berdasarkan uji t-test tidak berpasangan aktivitas mencari polen koloni 1 dan koloni 2 menunjukan nilai t=2,644, nilai p=0,01, karena nilai p=0,01 < alpha=0.05 maka aktivitas mencari polen T. laeviceps pada koloni 1 berbeda nyata dengan aktivitas terbang harian koloni 2.Jumlah T. laeviceps yang kembali ke
Berdasarkan uji t-test tidak berpasangan aktivitas terbang harian TL1KR dan TL2KR menunjukan nilai t=2,369, nilai p=0,02, karena nilai p=0,02 < alpha=0.05 maka aktivitas terbang harian T. laeviceps pada TL1KR berbeda nyata dengan aktivitas terbang harian
TL2KR. Jumlah T. laeviceps yang kembali ke sarang pada TL1KR dan TL2KR paling banyak terjadi siang hari (pukul 11:00-14:40 WIB), dengan jumlah rata-rata masing-masing 38,7 individu dan 32,4 individu (Tabel1).
\
a b
c d
e f
g h i
Gambar 4 Identifikasi Trigona (Tetragonula) laeviceps. a. Tubuh Trigona (Tetragonula) laeviceps, a. 1. ukuran tubuh ±4 mm, a.2. panjang sayap ±4 mm, b.tepi posterior tungkai belakang terdiri dari rambut (rostellum) yang bersisik c . mesoscutum ditutupi dengan rambut halus (tomentum), d cakram berbentuk bulatan memanjang (elliptical disc) pada basitarsus belakang e. Mandibula dengan dua gigi, f.1. Panjang tibia belakang 1,5 mm, f2. panjang basitarsus belakang 0.7 mm (separuh dari panjang tibia belakang), g. Clypeus, h. mesosomal dorsum. i. Sayap, i.1. pterostigma, i.2. marginal sel, i.3. sub marginal sel.
f.1 f.2
h
i.1
i.3
i.2 a.1
6
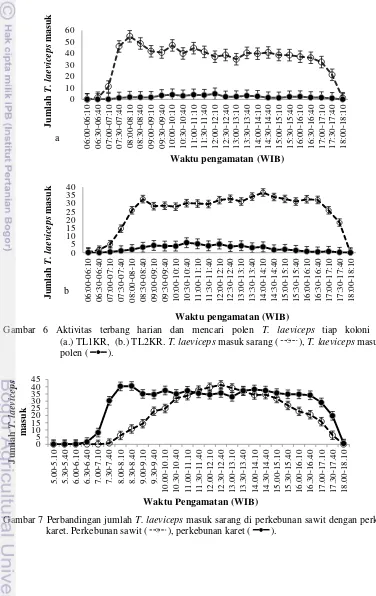
Setiap koloni T. laeviceps di KR mulai melakukan aktivitas mencari polen pukul 07.30 (suhu 25oC, kelembaban 88% dan intensitas cahaya 90 lux) dan berhenti pada waktu yang berbeda. TL1KR berhenti mencari polen pada pukul 17.40 sedangkan TL2KR berhenti pada pukul 17.10 (Gambar 6 a, b). Puncak aktivitas mencari polen dan jumlah individu yang membawa polen tiap 10 menit TL1KR dan TL2KR berturut-turut adalah pukul 12.00-12.10 (5 individu) (Gambar 6a) dan 10.30-10.40 (6 individu) (Gambar 6b). Puncak aktivitas mencari polen pada TL1KR terjadi pada suhu 30oC, kelembaban 68%, dan intensitas cahaya 317 lux sedangkan pada TL2KR terjadi pada suhu 31oC, kelembaban 66% dan intensitas cahaya 357 lux.
Berdasarkan uji t-test tidak berpasangan aktivitas mencari polen TL1KR dan TL2KR menunjukan nilai t=-0,918, nilai p=0,363, karena nilai p=0,36 > alpha=0.05 maka aktivitas mencari polen T. laeviceps pada
TL1KR tidak berbeda nyata dengan aktivitas terbang mencari polen TL2KR. Jumlah T. laeviceps yang kembali ke sarang membawa polen pada TL1KR dan TL2KR paling banyak terjadi siang hari (pukul 11:00-14:40 WIB) dengan jumlah rata-rata masing-masing 3,3 individu (suhu 31oC, kelembaban 67,5%, intensitas cahaya 287 lux) dan 3,9 individu (Tabel 2) (suhu 32,1oC, kelembaban 62,9%, intensitas cahaya 332 lux) (Tabel 3, 4, 5).
Perbandingan Aktivitas Terbang Harian
T. laeviceps di SW dengan KR
SW berlangsung selama 10,5 jam, sedangkan di KR berlangsung selama 11,5 jam.
Puncak aktivitas terbang harian T. laeviceps di KR lebih pagi yaitu pukul 08.00-08.40 (suhu 28oC, kelembaban 80%, maupun di SW yaitu 40 individu (Gambar 7).
Berdasarkan uji t-test tidak berpasangan aktivitas terbang harian SW dan KR menunjukan nilai t=-2,134, nilai p=0.03, karena nilai p=0.03 < alpha=0.05 maka hal ini menunjukan bahwa aktivitas terbang T. laeviceps di SW berbeda nyata dengan aktivitas terbang harian di KR.
Gambar 6 Aktivitas terbang harian dan mencari polen T. laeviceps tiap koloni di KR (a.) TL1KR, (b.) TL2KR. T. laeviceps masuk sarang ( ), T. laeviceps masuk bawa
polen ( ).
Gambar 7 Perbandingan jumlah T. laeviceps masuk sarang di perkebunan sawit dengan perkebunan karet. Perkebunan sawit ( ), perkebunan karet ( ).
8
Perbandingan Aktivitas Mencari Polen T. laeviceps di SW dengan KR
Koloni T. laeviceps di KR mulai mencari polen lebih pagi (pukul 07.00 dengan suhu 25oC, kelembaban 86%, intensitas cahaya 89 lux) daripada di SW (pukul 08.00 dengan suhu 25oC, kelembaban 85%, intensitas cahaya 66 lux) (Gambar 8). Semua koloni T. laeviceps di SW dan di KR berhenti pada waktu yang sama yaitu pukul 17.40.
Berdasarkan uji t-test tidak berpasangan aktivitas mencari polen di SW dan KR menunjukan nilai t=0,854, nilai p=0,39, karena nilai p=0,39 > alpha=0.05
maka hal ini menunjukan bahwa aktivitas mencari polen T. laeviceps di SW tidak berbeda nyata dengan aktivitas mencari polen di KR.
Persentase Aktivitas Mencari Polen T. laeviceps di SW dan KR
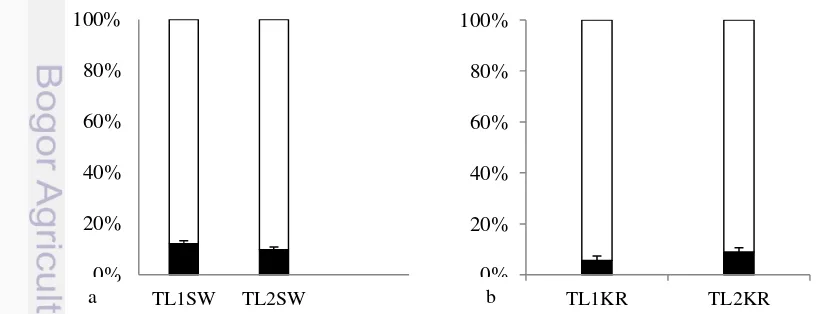
Persentase aktivitas mencari polen di SW bervariasi. Koloni TL2SW memiliki persentase lebih rendah dari TL1SW. Persentase aktivitas mencari polen TL1SW dan TL2SW masing-masing 12,1% dan 9,7% (Gambar 10a). Persentase aktivitas mencari polen di KR lebih rendah daripada di SW. Persentase aktivitas mencari polen TL1KR dan TL2KR masing-masing 5,7% dan 9% (Gambar 10b). Pada grafik berwarna putih T. laeviceps diduga membawa nektar, resin, air, atau material penyusun sarang lain seperti lumpur atau kotoran hewan.
Gambar 8 Perbandingan jumlah T. laeviceps mencari polen di perkebunan sawit dengan perkebunan karet. Perkebunan sawit ( ), perkebunan karet ( ).
Gambar 9 Persentase jumlah aktivitas mencari polen (hitam) terhadap aktivitas terbang harian (putih). (a.) Perkebunan Sawit, (b.) Perkebunan Karet.
Tabel 1 Jumlah Trigona laeviceps kembali ke sarang pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10), (n= pengamatan, x= rata-rata, sd= standar deviasi, dan - = tidak ada data karena hujan)
Tabel 2 Jumlah Trigona laeviceps yang kembali ke sarang membawa polen pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10), (n= pengamatan, x= rata-rata, sd= standar deviasi, dan - = tidak ada data karena hujan)
Cigudeg Ciaruteun ilir
Perkebunan Sawit Perkebunan Karet
koloni 1 koloni 2 koloni1 koloni 2
n pagi siang sore pagi siang sore pagi siang sore pagi siang sore 1 - 7±2,6 8,0±2,6 2,9±3,3 5,8±2,0 - 3,4±2,9 2,3±1,3 0,3±0,5 2,6±2,6 6,3±4,2 0,5±0,8 2 1,8±3,8 24,7±3,3 14±9,6 2,1±2,5 4,0±0,9 2,3±2,6 1,7±1,8 0,6±0,8 - 4,0±4,3 6,9±3,6 1,4±2,5 3 5,8±7,6 6,6±2,8 0,7±1,2 2,9±4,4 5,3±1,2 - 1,5±2,2 2,6±1,6 3,7±1,9 3,3±3,7 4,7±1,7 - 4 1,6±2,1 5,5±2,7 6,3±2,8 3,0±1,4 8,5±4,7 3,9±4,7 0,6±0,6 3,6±1,3 - 2,8±3,1 6,4±6,1 - 5 1,1±1,9 7,3±7,3 1,6±1,7 1,7±1,7 3,3±2,6 - 2,9±2,7 7,9±3 4,0±4,7 3,0±3,9 3,4±2,5 2,3±0,5 6 1,0±1,6 - - 2,5±1,0 2,6±1,0 2,0±1 1,3±2,2 5,7±3,5 1,3±1,9 5,7±4,4 4,1±3,4 0,7±1,1 7 1,4±1,7 5,6±2,1 - 0,9±1,3 1,8±1,0 0,3±0,7 3,6±4,7 6,8±6,5 - 2,2±2,6 2,4±1,5 0±0 8 1,7±2,2 4,5±3,4 4,6±3,0 0,6±1,3 1,6±1,5 0,9±1,1 1,0±2,1 2,9±2,1 0,0±0 1,7±1,7 2,4±0,8 0,4±1,1 9 0,3±0,7 1,4±0,7 0,0±0 1,5±1,3 2,5±1,9 - 1,7±2,5 3,0±2,2 3,5±2,1 1,3±1,7 2,9±1,7 0,4±0,7
Cigudeg Ciaruteun ilir
Perkebunan Sawit Perkebunan Karet
koloni 1 koloni 2 koloni 1 koloni 2
10
10 0,8±1,1 3,8±3,1 0,7±1,1 2,4±2.1 3,0±1,5 0,1±0,3 1,9±1,5 1,3±1,3 1,3±1,3 1,2±1,3 2,9±1,5 1,6±0,9 11 0,1±0,3 1,7±0,9 - 0,3±0,7 1,0±1 - 0,9±1,5 2,7±1,2 0,0±0 1,0±1,1 2,1±1,7 2,3±2,1 12 0,0±0 - - 0,4±0,7 1,1±0,9 - 0,7±0,8 2,1±1,6 0,7±0,9 1,7±1,8 2,1±1,6 0,3±0,4 13 0,1±1,1 1,8±1,2 0,8±1,1 0,0±0 2,8±1,2 0,6±0,7 1,2±1,7 2,0±1 0,4±0,7 2,9±2,6 4,9±2,3 0,1±0,3 x±sd 1,3±1,5 6,4±6,5 4±4,6 1,6±1,1 3,3±2,1 1,4±1,4 1,7±1 3,3±2,1 1,5±1,6 2,6±1,3 3,9±1,7 1±0,8
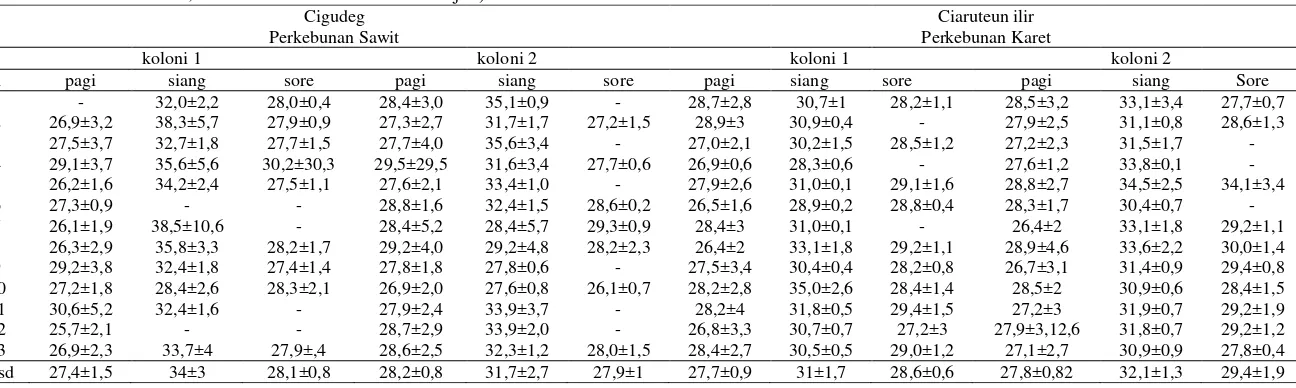
Tabel 3 Rata-rata suhu udara (oC) di sekitar sarang pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10), (n= pengamatan, x= rata-rata, sd= standar deviasi, dan - = tidak ada data karena hujan)
Tabel 4 Rata-rata kelembaban udara (%) di sekitar sarang pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10), (n= pengamatan, x= rata-rata, sd= standar deviasi, dan - = tidak ada data karena hujan).
Cigudeg Ciaruteun ilir
Perkebunan Sawit Perkebunan Karet
koloni 1 koloni 2 koloni 1 koloni 2
n pagi siang sore pagi Siang sore pagi siang sore pagi siang Sore 1 - 68,5±0,7 84,0±1,7 80,8±6,7 56,0±2,6 - 69,4±10,8 65,3±5,7 80,7±5,2 77,6±9,4 58,5±8,8 80,0±2,6 2 83,0±7,6 45,0±13,3 78,5±11,1 74,0±5,9 63,3±3,5 79,7±6,6 75,8±11,5 66,0±2,8 - 76,0±11,6 61,5±2,5 77,0±5
Cigudeg Ciaruteun ilir
Perkebunan Sawit Perkebunan Karet
koloni 1 koloni 2 koloni 1 koloni 2
3 78,7±11,3 61,5±8,7 77,0±11,5 76,2±13 55,5±17 - 79,8±6 69,8±8,7 83,0±2 82,6±7,3 82,6±7,9 - 4 74,7±14,4 47,8±16,1 67,5±0,7 73,0±73 71,3±3,4 83,0±4,9 80,0±4 80,5±2,1 - 86,2±3,2 65,0±2 - 5 83,8±1,7 83,8±4,5 76,0±7,8 83,8±6,8 56,5±3,5 - 80,0±8,2 68,3±0,9 79,7±9,5 77,2±11,5 55,5±6 69,0±69 6 82,8±2,8 - - 74,0±3,6 66,8±4,4 81,5±2,1 87,0±4,3 81,3±1,7 83,3±3 84,4±4,3 74,0±4,9 - 7 87,4±5,0 70,0±2,8 - 77,8±17 58,0±10,6 76,3±9,7 79,0±13,3 67,0±4,5 - 71,6±11,8 57,8±5,4 72,0±7,2 8 83,2±8,2 47,5±15,8 75,5±12,2 76,5±13,3 50,3±11 74,3±8,1 83,6±7,1 60,5±4,5 75,7±6,8 74,8±15,2 59,8±9,7 72,0±7,5 9 72,5±14,5 59,8±4,1 84,3±5,1 81,7±4,7 73,0±1,4 - 78,4±9 72,5±1,7 79,7±0,7 77,0±13,2 62,0±4,7 73,7±5,8 10 83,0±4,2 75,3±10,3 90,0±4 84,4±4,9 85,0±1,8 91,3±2,5 77,4±11,4 52,5±5,8 81,0±6,5 71,2±10,3 61,0±3,4 77,7±6,6 11 78,8±11,6 61,3±2,8 - 80,3±8,9 63,7±9,8 - 69,2±15,7 55,5±10,4 63,0±8,6 70,6±11,5 55,5±4,2 73,7±13,2 12 83,0±1,4 - - 77,0±7,5 57,0±5,3 - 81,2±13,2 68,5±7 77,7±10,5 77,0±9,8 62,3±4,2 76,7±3,7 13 72,3±4,8 51,5±7,9 71,3±5,0 76,5±7,8 62,3±4,9 81,0±6,5 83,2±9,3 70,5±9,1 78,0±2,6 78,2±10,1 62,8±4,1 84,3±4 x±sd 80,3±4,9 61,1±12,5 78,2±7 78,1±3,7 63±9,3 81±5,5 78,8±5 67,5±8,3 78,2±5,8 77,3±4,8 62,9±7,6 75,6±4,5
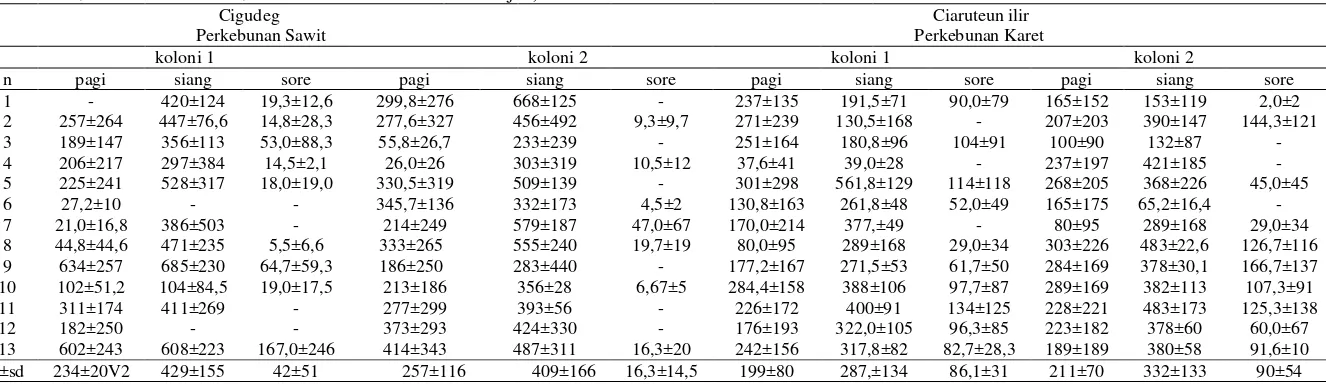
Tabel 5 Rata-rata intensitas cahaya (lux) di sekitar sarang pada pagi hari (05:00-10:40), siang hari (11:00-14:40) dan sore hari (15:00-18:10), (n= pengamatan, x= rata-rata, sd= standar deviasi, dan - = tidak ada data karena hujan).
Cigudeg Ciaruteun ilir Perkebunan Sawit Perkebunan Karet
koloni 1 koloni 2 koloni 1 koloni 2
n pagi siang sore pagi siang sore pagi siang sore pagi siang sore 1 - 420±124 19,3±12,6 299,8±276 668±125 - 237±135 191,5±71 90,0±79 165±152 153±119 2,0±2 2 257±264 447±76,6 14,8±28,3 277,6±327 456±492 9,3±9,7 271±239 130,5±168 - 207±203 390±147 144,3±121 3 189±147 356±113 53,0±88,3 55,8±26,7 233±239 - 251±164 180,8±96 104±91 100±90 132±87 - 4 206±217 297±384 14,5±2,1 26,0±26 303±319 10,5±12 37,6±41 39,0±28 - 237±197 421±185 - 5 225±241 528±317 18,0±19,0 330,5±319 509±139 - 301±298 561,8±129 114±118 268±205 368±226 45,0±45 6 27,2±10 - - 345,7±136 332±173 4,5±2 130,8±163 261,8±48 52,0±49 165±175 65,2±16,4 - 7 21,0±16,8 386±503 - 214±249 579±187 47,0±67 170,0±214 377,±49 - 80±95 289±168 29,0±34 8 44,8±44,6 471±235 5,5±6,6 333±265 555±240 19,7±19 80,0±95 289±168 29,0±34 303±226 483±22,6 126,7±116 9 634±257 685±230 64,7±59,3 186±250 283±440 - 177,2±167 271,5±53 61,7±50 284±169 378±30,1 166,7±137 10 102±51,2 104±84,5 19,0±17,5 213±186 356±28 6,67±5 284,4±158 388±106 97,7±87 289±169 382±113 107,3±91 11 311±174 411±269 - 277±299 393±56 - 226±172 400±91 134±125 228±221 483±173 125,3±138 12 182±250 - - 373±293 424±330 - 176±193 322,0±105 96,3±85 223±182 378±60 60,0±67 13 602±243 608±223 167,0±246 414±343 487±311 16,3±20 242±156 317,8±82 82,7±28,3 189±189 380±58 91,6±10 x±sd 234±20V2 429±155 42±51 257±116 409±166 16,3±14,5 199±80 287,±134 86,1±31 211±70 332±133 90±54
12
Hubungan Aktivitas Terbang T. laeviceps
dengan Faktor Lingkungan
Berdasarkan analisis Principle Component Analysis (PCA) yang digunakan untuk menganalisis hubungan aktivitas harian dan faktor lingkungan (Gambar 10) menunjukan bahwa intensitas cahaya di SW lebih berpengaruh dibandingkan suhu dan kelembaban. Intensitas cahaya dan suhu berkorelasi positif terhadap aktivitas terbang harian di SW, dengan nilai korelasi Pearson dan probabilitas masing-masing r=0,43 (kekuatan korelasi cukup), p=2.2e-16 dan r=0,17 (kekuatan korelasi sangat lemah), p=0.0001 (Tabel 1), artinya jika intensitas cahaya naik maka suhu akan naik dan meningkatkan aktivitas harian T. laeviceps mencari polen. Namun apabila intensitas cahaya sangat tinggi, T. laeviceps akan berkurang aktivitas di luar sarang. Kelembaban udara di SW berkorelasi negatif terhadap aktivitas terbang dengan nilai korelasi r=-0,11 (kekuatan korelasi sangat lemah), p=0.0203 (Tabel 6). Pada saat intensitas cahaya naik maka suhu akan naik dan menyebabkan kelembaban udara menjadi rendah. Kelembaban udara yang tinggi mengakibatkan aktivitas terbang T. laeviceps menurun. Intensitas cahaya dan suhu di KR berkorelasi positif dengan aktivitas terbang harian T. laeviceps dengan nilai korelasi masing-masing r=0.30 (kekuatan korelasi cukup), p=1.554e-15 dan r=0.31 (kekuatan
korelasi cukup), p=8.26e-14 (Tabel 7). Hal ini menunjukan bahwa intensitas cahaya di KR lebih rendah daripada di SW karena semua koloni berada dibawah naungan pohon karet yang rindang sehingga cahaya yang datang terhalang oleh dedaunan karet. Kelembaban udara di KR berkorelasi negatif dengan nilai r=-0.26 (kekuatan korelasi cukup), p=1.457e-10 (Tabel 7).
Korelasi antara suhu dengan jumlah T. laeviceps yang membawa polen di SW memiliki nilai probabilitas lebih besar dari 0.05 sehingga hubungan kedua variabel tersebut tidak signifikan. Faktor lingkungan lain (suhu, kelembaban, intensitas cahaya) di SW dan KR memiliki nilai probabilitas lebih kecil dari 0.05 sehingga semua faktor lingkungan berhubungan secara signifikan dengan aktivitas terbang harian dan aktivitas mencari polen.
Tabel 6. Nilai korelasi antara komponen lingkungan dengan jumlah T. laeviceps masuk sarang dan jumlah T. laeviceps bawa polen di perkebunan sawit
Parameter
Jumlah Trigona masuk sarang Jumlah Trigona membawa Polen Korelasi
Pearson
Probabilitas Korelasi Pearson Probabilitas
Suhu 0.17 0.0001 0.09 0.0615
Intensitas cahaya 0.43 2.2e-16 0.14 0.0022
Kelembaban -0.11 0.0203 -0.09 0.0306
Tabel 7. Nilai korelasi antara komponen lingkungan dengan jumlah T. laeviceps masuk sarang dan jumlah T. laeviceps bawa polen di perkebunan karet
Parameter
Jumlah Trigona masuk sarang Jumlah Trigona bawa Polen Korelasi
Pearson
Probabilitas Korelasi Pearson Probabilitas
Suhu 0.32 1.554e-15 0.35 2.2e-16
Intensitas cahaya 0.31 8.26e-14 0.41 2.2e-16
kelembaban -0.26 1.457e-10 -0.29 1.516e-12
Gambar 11 Susunan sarang pada koloni 1 T. laeviceps di perkebunan sawit (data pada tanggal 6 Mei 2012).
PEMBAHASAN
Puncak aktivitas terbang T. laeviceps dan jumlah individu yang mencari polen bervariasi pada setiap koloni. Aktivitas Trigona mencari polen dipengaruhi oleh faktor luar dan faktor dalam koloni. Faktor luar yaitu ketersediaan pakan di sekitar sarang. Sumber polen yang ada di perkebunan sawit diantaranya bunga kelapa sawit (Elaeis guineensis) pada saat anthesis, putri malu (Mimosa pudica), babadotan (Ageratum conyzoides), dan rerumputan sekitar sarang. Sedangkan sumber polen yang ada di perkebunan karet yaitu, putri malu (Mimosa pudica), bunga pohon pinang (Areca catechu), dan rerumputan (data tidak dipublikasikan). Faktor lingkungan yang mempengaruhi aktivitas terbang dan mencari polen adalah suhu, angin, curah hujan, kelembaban dan intensitas cahaya (Michener 1974). Dibandingkan dengan lingkungan di perkebunan karet, kelembaban, intensitas
cahaya dan curah hujan di perkebunan sawit lebih tinggi. Faktor lingkungan seperti suhu, kelembaban, intensitas cahaya, ketinggian tempat dan curah hujan mempengaruhi aktivitas terbang harian Melipona bicolor bicolor yang termasuk stingless bee (Hilario et al. 2000).
14
pada pagi hari (07.00-10.00) dan sore hari (14.00-17.00) (Nelli (2003). Perbedaan
tersebut terjadi karena kemungkinan terdapat dua masa pemekaran bunga di
daerah tempat penelitian Trigona pada Nelli (2003).
Pada stingless bee terjadi komunikasi untuk memberikan informasi keberadaan sumber polen dengan komunikasi menggunakan sinyal suara (getaran sayap) (Aguilar & Briceno 2002). Hal ini sesuai dengan penelitian ini T. laeviceps mengumpulkan pakan dari tanaman di dekat sarang sekitar 1-4 meter (pengamatan pribadi), sehingga dapat menggunakan energi secara optimal, hal ini sesuai dengan pernyataan Ciar et al. (2009) pada Trigona biroi. Melipona costaricensis yang kembali ke sarang akan menggetarkan sayap diantara anggota koloni yang lain kemudian mengeluarkan polen yang dibawa M. costaricensis tersebut agar anggota koloni yang lain mengetahui jenis polen yang dibawa. Suara yang timbul akan menarik perhatian anggota koloni yang lain dan akan pergi meninggalkan sarang untuk mencari pakan yang telah diperlihatkan (Aguilar & Briceno 2002).
Sistem komunikasi merekrut ke sumber makanan pada stingless bee juga ada yang menggunakan penciuman (scent trail) seperti Melipona seminigra (Hrncir et al. 2004) , Trigona recursa dan Trigona hypogea menandai sumber makanannya menggunakan feromon jejak dari sarang ke letak sumber pakan yang berasal dari kelenjar mandibular yang dikenali oleh Trigona lain dalam satu koloni (Kerr 1967). Trigona yang menemukan sumber polen akan kembali ke sarang dan akan hinggap setiap beberapa meter pada daun, ranting, atau batu yang dapat membuat Trigona dapat mengingat rute tersebut. Setiap melakukan aktivitas tersebut Trigona akan menggigit permukaan daun, ranting, atau batu sambil mengeluarkan cairan dari kelenjar mandibular, cairan itu mempunyai bau yang sangat kuat dan terdiri dari komponen aromatik yang hanya dikenali oleh Trigona lain dalam satu koloni (Kerr 1967).
Aktivitas terbang serangga sangat dipengaruhi oleh proses termoregulasi (pengaturan suhu tubuh). Seperti pada Bombus sp. yang akan memulai aktivitas termasuk golongan serangga poikilotermik yang peka terhadap perubahan cuaca dan suhu tubuh dipengaruhi oleh perubahan lingkungan (Winston 1897).
Struktur susunan sarang dimulai dari pintu masuk (tunnel), tumpukan propolis yang digunakan untuk perlindungan sarang dan mensterilkan sarang, tempat penyimpanan makanan, dan yang paling dalam adalah sel perawatan telur, larva, dan pupa (brood cells) (Gambar 11). Hal ini sesuai dengan pernyataan Kerr (1967) bahwa tempat penyimpanan makanan terletak disebelah luar sel anakan. Sel anakan terletak paling dalam agar terlindungi dari berbagai gangguan dan dilindungi oleh pelepah lembut yang disebut involucrum. Sel anakan berbentuk vertikal dan membuka pada bagian atasnya. Tempat penyimpanan makanan berbentuk bulat, bulat telur atau kerucut (Roubik 1989). Sel anakan disangga oleh pilar dan bagian luarnya dilapisi oleh batumen. Pintu masuk sarang berupa tabung yang mengarah kedalam sepanjang dinding rongga sarang. Hal ini bertujuan untuk mengarahkan lebah pencari pakan menuju tempat penyimpanan makanan (Free 1982).
Percobaan eksperimen memindahkan T. laeviceps dari lingkungan alami yang heterogen ke lingkungan homogen untuk mempelajari perubahan aktivitas mencari polen terhadap perubahan lingkungan. Berdasarkan penelitian ini aktivitas mencari polen rendah di lingkungan homogen dari pada di lingkungan alami (Nelli 2003). Dengan demikian data hasil penelitian ini dapat digunakan sebagai data awal untuk menjadikan T.laeviceps sebagai bioindikator lingkungan seperti definisi dari McGeoch (1998) bahwa bioindikator adalah kelompok organisme yang sensitif terhadap perubahan lingkungan dan memperlihatkan gejala terpengaruh terhadap tekanan lingkungan yang terjadi. Selanjutnya Speight (1999) menyatakan bahwa faktor biotik berkaitan dengan faktor abiotik, dimana faktor biotik akan mengalami perubahan kehadiran, vitalitas dan respon sebagai akibat pengaruh kondisi lingkungan.
SIMPULAN
07.30 dengan puncak aktivitas pukul 12.30-14.10. Aktivitas terbang harian dan polen T. laeviceps pada setiap koloni, lebih tinggi pada siang hari (11.00-14.10).
Intensitas cahaya dan suhu berkorelasi positif terhadap aktivitas terbang harian dan aktivitas mencari polen T. laeviceps, sedangkan kelembaban berkorelasi negatif. Korelasi Pearson antara faktor lingkungan seperti suhu, kelembaban dan intensitas cahaya memiliki hubungan yang signifikan dengan aktivitas terbang harian dan mencari polen T.laeviceps, sedangkan untuk suhu dengan aktivitas mencari polen di perkebunan sawit tidak signifikan.
SARAN
Penelitian ini merupakan dasar sebagai model untuk penelitian penggunaan Trigona sebagai bioindikator perubahan lingkungan. Penelitian dilakukan di daerah perkebunan, sehingga perlu dilakukan lagi penelitian di daerah alami dengan jenis tanaman heterogen. Pengamatan anatomi sarang perlu dikembangkan untuk mengetahui pertambahan ukuran maupun volume pada komponen sarang T.laeviceps.
DAFTAR PUSTAKA
Aguilar I, Briceño D. (2002). Sounds in Melipona costaricensis (Apidae: Meliponini): effect of sugar concentration and nectar source distance. Apidologie 33: 375-388. Borror DJ, Johnson NF, Triphlehorn CA.
1981. An Introduction to the Study of Insects. New York: Saunders.
Ciar RR, Bonto LS, Bayer MHP. 2009. Foraging behaviour of Stingless bees (Trigona biroi Friese: Distance, Directionally and Height [tesis]. Pilipina: Fakultas Matematika dan Ilmu Pengetahuan Alam, University of the Philipines Losbanos.
Free JB. 1982. Bees and Mandkind. London: George Allen and Unwind.
Hilario SD, Imperatriz-Fonseca VL, Kleinert A de MP. 2000. Flight activity and colony strength in the stingless bee Melipona bicolor bicolor (Apidae, Meliponinae). Rev Brasil Biol. 60: 299-306.
Heinrich B. 1940. Bumblebee Economics. Cambridge: The Belknap of Harvard Univ Pr.
Hrncir M, Jarau S, Zucchi R., & Barth FG. (2004). On the origin and properties of scent marks deposited at the food source by a stingless bee, Melipona seminigra. Apidologie, 35: 3-13. Keller I, Fluri P, Imdorf A. 2005. Pollen
nutrition and colony development in honney bees-part II. Bee World. 86: 27-34.
Kerr WE.1967. The history of the introduction of African Bees to Brazil.South African Bee J. 39: 3-5 and Apiculture in Western Australia 2:53-55.
Lisboa LC, Serrao JE, Cruzlandim C, Campor RA. 2005. Effect of Larval Food Amount on Ovariole Development in Queens of Trigona spinipes (Hymenoptera, Apinae) [abstrak]. Anatomia, Histologia, Embriyologia 34: 179-184.
McGeoch M. 1998. The selection, testing and application of terrestrial insects as bioindicators. Biological Reviews, 73, 181 201.
Michener CD. 1974. The Social Behaviour of The Bees. Cambridge: The Belknap of Harvard Univ Pr.
Michener CD. 2000. The Bees of The World. Baltimore: The John Hopkins Univ Pr. Nelli. 2004. Waktu Pencarian Serbuk Sari
Lebah Pekerja Trigona
(Apidae:Hymenoptera) [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor. Melipona scutellaris Latreille (Apidae, Meliponini). Revista Brasileira de Zoologia. 20: 565-571. Roubik DW. 1989. Ecology and Natural
History of Tropical Bees. New York: Cambridge Univ. Press.
Sakagami SF, Ohgushi R, Roubik DW. 1990. Natural History of Social Wasps and Bees in Equatorial Sumatra. Sapporo: Hokkaido Univ. Press.
Shahabbudin. 2003. Pemanfaatan Serangga Sebagai Bioindikator Kesehatan Hutan. Pengantar Falsafah Sains. Bogor: Institut Pertanian Bogor. Shuel RW. 1992. The Production of Nectar
16
the honey bee. Illinois: Dadant & Hamilton, Ltd.
Speight MR, Hunter MD, Watt AD. 1999. Ecology of Insects, Concept and Aplications. USA: Blackwell Science. Sunderlin WD, Resosudarmo IAP. 1996.
Rates and Causes of Deforestation in Indonesia : Toward a Resolution of the Ambiguities. Occasional Paper No.9. CIFOR, Bogor.
Walpole RE. 1992. Pengantar Statistik. Ed ke-3. Sumantri B. Penerjemah Jakarta: PT. Gramedia Pustaka Utama. Terjemahan dari Introduction to statistics 3rd Edition.
18
Lampiran 1 Peta lokasi perkebunan sawit di Kecamatan Cigudeg, Kabupaten Bogor.
Lampiran 2 Peta lokasi perkebunan karet di Kecamatan Cibungbulang, Kabupaten Bogor.
Lampiran 3 kunci identifikasi Trigona laeviceps.
1. Bagian pinggir posterior dari tibia belakang terdiri dari rambut yang sederhana... 2
- Bagian pinggir posteror dari tibia belakang kebanyakan teriri dari rambut yang bersisik….. 7
7. Basitarsus belakang dibawah bagian basal tidak memiliki cakram (disc) berbentuk elips yang ditutupi bulu halus. Spesies yang lebih besar, tubuh dan sayap masing-masing lebih dari 8 mm. Keseluruhan atau hampir dominan ferrugious. Clypeus tanpa bulu yang keras, tegak lurus dan berwarna hitam………...……..T. (Homotrigona) fimbriata - Basitarsus belakang dibawah bagian basal memiliki cakram (disc) seperti yang disebutkan di atas (bulu halus terkadang ada)... 8
8. Mandibula dengan satu gigi yang lemah. Basitarsus belakang sekitar 2/3 kali lebarnya tibia belakang………...……….. T. (Heterorigona) 9 - Mandibula dengan dua gigi. Kecuali Geniotrigona, basitarsus belakangnya setengah kali lebarnya tibia belakang... 10
10. Bagian tengah propodeum ditutupi rambut yang seperti selimut. Gigi pada mandibulanya kuat. Spesies yag kuat dan berwarna hitam, tubuh dan sayapnya lebih dari 7 mm………...…... T. (Lepidotrigona) canifrons - Bagian tengah propodeum halus dan bersinar ... 11
11. Mesoscutellum memproyeksikan posterior yang melebihi lekukan propodeum secara jelas di bagian belakang. Jarak malar linear atau kebanyakan lebih pendek dari setengahnya lebar flagellomere ………....…………...………. T. (Tetragonula)17 - Mesoscutellum tidak memproyeksikan posterior yang melebihi lekuk propodeum. Jarak malar sama dengan lebar F2 atau lebih panjang ...12
17. Mandibula dengan gigi yang kuat seperti pada T. apicalis. Gena hampir sama lebar dengan mata, jarak ocellocipital hampir sama lebar dengan diameter ocellar. Ukuran tubuh 5 mm, dan ukuran sayap 6 mm ...reepeni - Mandibula dengan gigi yang lemah seperti pada T. moorei. Gena jelas lebih pedek dari mata. Jarak ocellocipital sekitar setengahnya jarak diameter ocellar. Spesies yang lebih kecil ...18
18. Predominan dari tubuhnya kemerahan atau coklat pucat ...19
- Setidaknya kepala kecuali clypeus dan dorsum berwarna kehitaman ... 2
20. Sayap depan bagian basal kira-kira lebih gelap dari bagian apikal ...21
- Sayap depan cukup seragam berwarna transparan atau hanya lebih gelap secara tipis ...3
23. Spesies yang sangat kecil, tubuh dan sayapnya masing-masing hingga 3,5 mm. Mesoscutalnya merupakan daerah yang halus, terutama bagian lateralnya (G3) berambut...……….…..fusobalteata - Lebih besar, mesoscutalnya merupakan daerah yang halus, terutama bagian lateralnya (G3) tidak berambut ... 24
24. Predominan tubuhnya berwarna hitam ... 2
20
25. Spesies yang lebih besar, dengan kaki belakang yang lebih panjang. Ukuran tubuh 4-4,5 mm, dan sayap 5-5.5 mm, tibia belakang lebih dari 2 mm ...…………geissleri Cockerell) - Lebih kecil, tibia belakang kurang dari 2 mm (perbedaan pada geissleri terlihat jelas pada jantan namun cukup metrik pada pekerja) ……….……… laeviceps s. lat
Dari hasil identifikasi diketahui bahwa spesies Trigona ini adalah Trigona (Tetragonula) laeviceps.