KARAKTERISTIK MORFOLOGI DAN PEMILIHAN JENIS PAKAN
OLEH KELELAWAR MEGACHIROPTERA DI HUTAN
PENDIDIKAN GUNUNG WALAT, KAB. SUKABUMI
AMALIA CHOIRUNNISA
DEPARTEMEN KONSERVASI SUMBERDAYA HUTAN DAN EKOWISATA FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Karakteristik Morfologi dan Pemilihan Jenis Pakan Oleh Kelelawar Megachiroptera di Hutan Pendidikan Gunung Walat, Kab. Sukabumi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2015
Amalia Choirunnisa
ABSTRAK
AMALIA CHOIRUNNISA. Karakteristik Morfologi dan Pemilihan Jenis Pakan oleh Kelelawar Megachiroptera di Hutan Pendidikan Gunung Walat, Kab. Sukabumi. Dibimbing oleh AGUS PRIYONO KARTONO dan IBNU MARYANTO.
Kelelawar megachiroptera merupakan kelelawar pemakan buah, nektar dan polen. Keberadaan satwa berfungsi sebagai agen penyebar biji dan penyerbuk bunga. Sehingga penting untuk mengetahui kemampuan adaptasi dan pakannya di alam. Penelitian ini dilaksanakan di HPGW mulai 14 juni-25 agustus 2014. Kelelawar ditangkap dengan menggunakan jaring kabut pada empat tipe tutupan lahan dengan metode purposive sampling. Terdapat
sembilan jenis kelelawar yang teridentifikasi dalam penelitian ini. Tumbuhan pakan yang teridentifikasi sebanyak 59 jenis dari 38 famili dan 13 tipe mahkota bunga yang diidentifikasi dari analisis. Fabaceae, Musaceae, dan Rubiaceae merupakan genus yang paling mempengaruhi perilaku makan. Hasil dari PCA dan regresi logistik biner menunnjukkan bahwa D3P2, D4P2, D4P1 dan D4P5 adalah peubah yang dapat dijadikan pembeda jenis pada setiap habitat. Hasil analisis CCA dengan Canoco menyatakan bahwa
caryophllaceous sangat dipengaruhi oleh ukuran C1M3, sedangakan bunga tipe appetallouse lebih fleksibel dengan ukuran tengkorak dan gigi.
Kata kunci: kelelawar Megachiroptera, morfometri, polen.
ABSTRACT
AMALIA CHOIRUNNISA. Morphological Characteristics and Diet Selection by Bats Megachiroptera In Gunung Walat University Forest, Sukabumi. Supervised by AGUS PRIYONO KARTONO and IBNU MARYANTO.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
pada
Departemen Konservasi Sumberdaya Hutan dan Ekowisata
AMALIA CHOIRUNNISA
DEPARTEMEN KONSERVASI SUMBERDAYA HUTAN DAN EKOWISATA FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR BOGOR
2015
KARAKTERISTIK MORFOLOGI DAN PEMILIHAN JENIS PAKAN
OLEH KELELAWAR MEGACHIROPTERA DI HUTAN
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia dan rahmat-Nya sehingga penyusunan karya ilmiah ini dapat diselesaikan. Penelitian ini memiliki tema kelelawar dengan judul Karakteristik Morfologi dan Pemilihan Jenis Pakan oleh Kelelawar Megachiroptera di Hutan Pendidikan Gunung Walat, Kab. Sukabumi. Pengumpulan data lapangan dilaksanakan pada bulan Mei sampai dengan bulan Agustus 2014 di Hutan Pendidikan Gunung Walat (HPGW).
Terima kasih penulis sampaikan kepada Dr Ir Agus Priyono Kartono, MSi dan Prof (RIS) Dr Ir Ibnu Maryanto, MSi selaku Pembimbing atas arahan dan saran yang telah diberikan. Penghargaan penulis sampaikan kepada Dr Ir Nandi Kosmaryandi, MScF selaku Direktur HPGW, Asisten Manajer Lapang, dan seluruh staf HPGW, serta keluarga Lab. Genetika Hutan, Lab. Mikoriza dan Lab. Entomologi Hutan, Dept. Silvikultur yang telah membantu selama penelitian berlangsung. Ungkapan terima kasih juga penulis sampaikan kepada ayah Wachid Hasyim, ibu Susilo Mardirini, kakak Nur M Arrozy dan adik Bintang Al-Bana atas dukungan dan kasih. Tak lupa penulis sampaikan terima kasih untuk partner penelitian (Kendy D. Prayogi dan Oktania Kusuma),
sahabat (Wida Agustina, Nuning Hamidah, Febriyanti Mutiara, Lyan Lavista dan Anugro P.), keluarga besar Fakultas Kehutanan IPB, Departemen KSHE, Nepenthes Raflessiana 47, dan HIMAKOVA serta semua pihak yang telah memberikan bantuan moral maupun material selama penelitian berlangsung.
Semoga karya ilmiah ini bermanfaat.
Bogor, Februari 2015
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR vii
DARTAR LAMPIRAN viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 1
Manfaat Penelitian 2
METODE 2
Lokasi dan Waktu 2
Alat dan Bahan 3
Metode Pengambilan Data 3
Metode Pengolahan Data 6
HASIL DAN PEMBAHASAN 8
Hasil 8
Pembahasan 17
SIMPULAN DAN SARAN 23
Simpulan 23
Saran 24
DAFTAR PUSTAKA 24
DAFTAR TABEL
1 Komposisi jenis kelelawar Megachiroptera di HPGW dan status
perlindungannya 9
2 Komposisi kelelawar Megachiroptera berdasarkan jenis kelamin
dan lokasi perjumpaan 9
3 Hasil uji data peubah morfometri sayap terhadap habitatnya
menggunakan analisis Regresi Logistik Biner 14 4 Perbandingan kerapatan tegakan pada tiap tutupan lahan 14 5 Matrik niche overlap pada jenis kelelawar Megachiroptera 17
DAFTAR GAMBAR
1 Peta lokasi penelitian dan plot pengamatan 2
2 Kegiatan melepas kelelawar dari jala kabut 3
3 Parameter pengukuran karakteristik luar 4
4 Parameter karateristik tengkorak dan gigi kelelawar menurut
Andersen (1912) diacu dalam Rahman dan Abdullah (2010) 5
5 Tipe-tipe mahkota bunga 7
6 Bunga Calliandra sp. (tipe mahkota bunga apetalouse/bunga
telanjang) (a); bunga Annona sp. (tipe mahkota bunga sepalloid) (b);
bunga Passiflora sp. (tipe mahkota bunga rosaceous-corona) (c) 7 7 Komposisi pakan kelelawar Megachiropthera berdasarkan famili
tumbuhan 10
8 Jumlah jenis tumbuhan pakan yang dimanfaatkan oleh kelelawar
Megachiroptera 10
9 Pengelompokan kelelawar Megachiroptera berdasarkan jenis sumber
pakan 11
10 Pengelompokan jenis kelamin kelelawar Megachiroptera
berdasarkan jenis sumber pakan 11
11 Komposisi pakan kelelawar Megachiroptera berdasarkan bentuk
mahkota bunga 12
12 Pengelompokan jenis kelelawar berdasarkan pemilihan bentuk
mahkota bunga 13
13 Pengelompokan jenis kelamin kelelawar berdasarkan pemilihan
bentuk mahkota bunga 13
14 ubungan karakteristik ukuran tengkorak dan gigi terhadap tipe
bentuk mahkota bunga pada axis 1 dengan axis 2 15 15 Hubungan karakteristik ukuran tengkorak dan gigi terhadap tipe
DAFTAR LAMPIRAN
1 Hasil identifikasi polen pada kelelawar Megachiroptera di HPGW 28 2 Jumlah pemanfaatan sumber pakan oleh kelelawar Megachiroptera 37 3 Ukuran morfologi sayap kelelawar Megachiroptera 40 4 Ukuran morfologi tengkotak (cranial) kelelawar Megachiroptera 42 5 Ukuran morfologi gigi (dental) kelelawar Megacriroptera 44 6 Hasil penghitungan analisis cluster variable jenis kelelawar
Megachiroptera berdasarkan kesamaan sumber pakan pakan
menggunakan software Minitab 16 45
7 Hasil penghitungan analisis cluster variable jenis kelamin kelelawar Megachiroptera berdasarkan kesamaan sumber pakan pakan
menggunakan software Minitab 16 46
8 Hasil penghitungan analisis cluster variable pengelompokan jenis kelelawar Megachiroptera berdasarkan kesamaan bentuk mahkota
bunga (corolla) menggunakan software Minitab 16 46 9 Hasil penghitungan analisis cluster variable pengelompokan jenis
kelamin kelelawar Megachiroptera berdasarkan kesamaan bentuk
mahkota bunga (corolla) menggunakan software Minitab 16 47 10 Tabel total variance explained metode PCA pada tampilan software
SPSS 47
11 Hasil uji Omnimbus dalam penghitungan metode regresi logistik
biner tiap tipe tutupan lahan menggunakan software SPSS 48 12 Hasil uji Hosmer and Lemeshow dan Negelkerke R Square dalam
metode regresi logistik biner pada tiap tipe tutupan lahan
menggunakan software SPSS 49
13 Tabel variabel in equation dalam metode regresi logistik biner pada tiap tipe tutupan lahan menggunakan software SPSS 50 14 Matrix niche overlap berdasarkan jenis kelamin kelelalar
PENDAHULUAN
Latar Belakang
Jumlah kelelawar didunia saat ini kurang lebih mencapai 1024 spesies (Tudge 2000) yang dibagi menjadi dua Sub-Ordo yaitu, Megachiroptera dan Mikrochiroptera (Miller 1907 diacu dalam Anderson dan Jones 1967; Young 1981). Pteropodidae merupakan satu-satunya famili dalam Sub-Ordo Megachiroptera dengan anggota jenis mencapai 246 spesies atau sama dengan 24% dari total spesies dalam Ordo Chiroptera (Emmons dan Feer 1997). Kelelawar Megachiroptera merupakan kelompok kelelawar yang memakan buah, nektar dan polen, yang berguna dalam membantu proses penyebaran biji dan berperan sebagai polinator bagi tumbuhan (Coldbert et al. 2001; Suyanto 2001;
Hikmann et al. 2007; Dumont et al. 2011; Fleming dan Kress 2011). Didukung
dengan kemampuan terbang (Mickleburgh et al. 1992; Emmons dan Feer 1997;
Tudge 2000; Suyanto 2001; Shepherd C dan Shepherd L 2012) dan distribusinya yang luas (Kingston et al. 2006; Dechmann 2006), kelelawar menjadi komponen
penting yang perlu diperhatikan dalam upaya konservasi tumbuhan pada suatu ekosistem hutan, khususnya hutan tropis (Maharadatunkamsi dan Maryati 2008).
Studi tentang morfologi penting dipelajari untuk mengetahui hubungan antara bentuk dan fungsi pada bagian tubuh individu yang mendukung kemampuan adaptasi suatu individu dengan lingkungannya (Galis 1996). Dumont
et al. (2011) menyatakan bahwa adaptasi secara morfologis merupakan salah satu
cara satwa untuk bertahan dari tekanan yang ada dalam lingkungan atau habitat salah satunya adalah dalam memperoleh pakan. Sebagai contoh, beberapa kelelawar yang berukuran kecil memakan nektar dengan cara hinggap (hovering)
sedangkan kelelawar yang berukuran lebih besar mendekati pakan dengan memanjat pada ranting menggunakan kuku di sayapnya (Voigt 2004).
Studi yang dilakukan Maryati (2008) menyatakan bahwa Cynopterus branchyotis memakan 16 jenis tumbuhan, Macroglossus sobrinus 20 jenis
tumbuhan dan Chinorax melanocephalus hanya 3 jenis tumbuhan dari 21 jenis
tumbuhan yang ditemukan di Taman Nasional Gunung Ciremai. Hal ini menunjukkan adanya perilaku pemilihan pakan pada kelelawar pemakan buah dan nektar. Selain itu, didalam studi tersebut juga dibuktikan adanya kompetisi yang tinggi, baik kompetisi intra maupun inter spesies yang menyebabkan terjadinya
overlap dalam memanfaatkan sumber daya pakan.
Berdasarkan hal tersebut maka perlu dilakukan studi tentang hubungan antara karakteristik morfologi kelelawar Megachiroptera dengan jenis sumber pakan dan kondisi habitatnya. Hasil penelitian ini diharapkan dapat digunakan untuk mengetahui kemampuan satwa untuk beradaptasi dengan lingkungannya.
Tujuan Penelitian
Penelitian mengenai karakteristik morfologi dan pemilihan jenis pakan kelelawar Megachiroptera di Hutan Pendidikan Gunung Walat bertujuan untuk : 1. Mengidentifikasi jenis sumber daya pakan yang dimanfaatkan kelelawar
2. Mengukur hubungan antara karakteristik morfometri kelelawar Megachiroptera dengan tipe habitat.
3. Mengukur hubungan antara karakteristik morfometri kelelawar Megachiroptera dengan pemilihan jenis pakan.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat menyediakan data dasar untuk selanjutnya dapat digunakan sebagai panduan pengamatan dan monitoring jangka panjang yang berguna bagi tujuan pendidikan di HPGW. Hasil penelitian ini dapat menjadi masukan bagi pengelolaan kawasan yang diharapkan dapat beriringan dengan upaya konservasi satwa liar khususnya kelelawar.
METODE
Lokasi dan Waktu
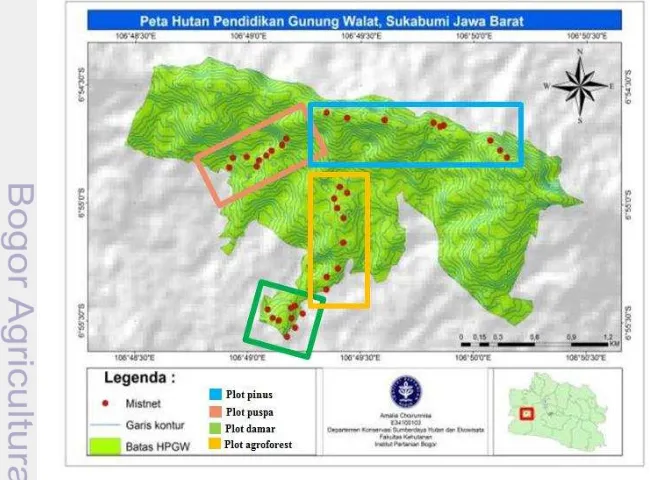
Pengambilan data dilakukan di kawasan Hutan Pendidikan Gunung Walat (HPGW) selama tiga bulan, yaitu pada bulan Mei-Agustus 2014. Secara Administrasi Pemerintahan, HPGW terletak dalam wilayah Kecamatan Cibadak Kabupaten Sukabumi, Jawa Barat. Secara Administrasi Kehutanan, kawasan ini masuk dalam wilayah BKPH Gede Barat, KPH Sukabumi Perum Perhutani Unit III Jawa Barat (Gambar 1). Kegiatan pengambilan data dilakukan pada empat area tutupan lahan berbeda yang ada dikawasan HPGW, yaitu tutupan lahan pinus, puspa damar dan lahan agroforest.
Alat dan Bahan
Alat yang digunakan untuk pengambilan spesimen adalah jaring kabut (mist net), headlamp, kantong blacu, sarung tangan dan penunjuk waktu. Kegiatan
pengukuran morfologi dan pencatatan bobot dilakukan dengan menggunakan Krisbow digital caliper KW06-351 dengan akurasi 0.05 mm dan Tanita digital scale KD-160 dengan ketelitian 0,5 gr. Alat lain yang digunakan adalah surgical blade nomor 10 dan 11, pinset, gunting anatomis, suntikan, dan sarung tangan
steril. Analisis polen dilakukan di laboraturium menggunakan mesin separator sentrifugal, mikroskop, tabung reaksi, object glass/kaca preparat, dan kuteks.
Alkohol 97% dan formalin 30% digunakan sebagai bahan untuk pengawetan spesimen. Selain itu, bahan lainnya berupa kapas berkloroform digunakan untuk membius spesimen. Alkohol 70% dan gliserol digunakan untuk analisis polen.
Identifikasi kelelawar dilakukan dengan menggunakan buku LIPI-Seri Panduan Lapangan Kelelawar Indonesia (Suyanto 2001) dan Panduan Lapang Mamalia di Kalimantan, Sabah, Serawak, dan Brunei Darusalam (Payne et al.
2000) dan Teknik Survey dan Identifikasi Jenis-Jenis Kelalawar Khas Agroforestry Sumatera (Prasetyo et al. 2011). Identifikasi polen sampai tingkat
famili dan genus dilakukan menurut kunci determinasi Erdmant (1952), Nayar (1999), Paldat (2005), dan Hesse et al. (2009).
Metode Pengambilan Data
Pengambilan data dilakukan pada empat plot pengamatan yaitu tutupan lahan agathis, tutupan lahan pinus, tutupan lahan puspa dan lahan agroforest di dalam kawasan HPGW. Setiap plot pengamatan dibagi menjadi tiga titik pengambilan data. Tiga mist net dipasang pada setiap titik dengan jarak 100 m dan
dilakukan pengulangan sebanyak tiga kali. Kelelawar Megachiroptera tidak memiliki kemampuan ekolokasi kecuali marga Rousettus yang menggunakan
getaran lewat ketukan gigi,sehingga penangkapan dengan jaring kabut merupakan cara yang paling baik (Payne et al. 2000) (Gambar 2).
Gambar 2 Kegiatan melepas kelelawar dari jala kabut
Jaring kabut yang digunakan berukuran 12 x 3 m dipasang dengan menggunakan bambu sepanjang ±5 m (Maharadatunkamsi dan Maryati 2008, Mohd-Azlan et al. 2003). Penempatan jaring kabut dilakukan dengan teknik purposive sampling sedangkan pengambilan sampel kelelawar dilakukan dengan
kemudian pada pukul 19.00 dan pukul 06.00 WIB dilakukan pengecekan dan pengambilan sampel kelelawar. Kelelawar ditangkap dan dimasukkan pada kantung blacu kemudian dipisahkan sesuai nomor jaring, lokasi penangkapan dan hari penangkapan.
Kelelawar yang ditangkap diidentifikasi jenis dan status perlindunganya berdasarkan status satwa yang dilindungi oleh PP No.7 Tahun 1999, kategori satwa langka oleh IUCN Red List dan status perdagangan oleh CITES. Penomoran
dilakukan bagi individu yang dijadikan sebagai spesimen sedangkan individu yang dilepas ditandai dengan kuteks berwarna kuning. Kelelawar yang dijadikan spesimen diukur dan diambil sample kotorannya. Sampel kotoran tersebut dianalisis untuk mencari polen yang tertelan oleh kelelawar. Polen yang telah teridentifikasi diasumsikan sebagai sumber pakan.
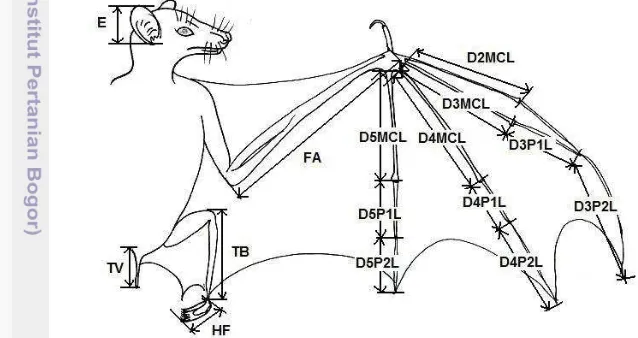
Rahman dan Abdullah (2010) menjelaskan bahwa terdapat 33 parameter karakteristik ukuran tubuh, tengkorak (cranial) dan gigi (dental) yang diperlukan
dalam analisi morfologi kelelawar. Terdapat 15 karakteristik luar tubuh kelelawar yang dilakukan pengukuran (Gambar 3), yaitu:
Keterangan :pajang telinga/Ear length (E), panjang betis/Tibia length (TB), kaki belakang/Hind foot length (HF), panjang ekor/Tail to ventral length (TVL), lengan bawah sayap/Fore arm (FA),
jari metakarpal kedua/Second digit metacarpal length (D2MCL), panjang jari metakarpal
ketiga/Third digit metacarpal length (D3MCL), jari metakarpal ketiga ruas phalanak
pertama/Third digit first phalanax length (D3P1L), jari metakarpal ketiga ruas phalanak
kedua/Third digit second phalanax length (D3P2L), jari metakarpal keempat /Fourth digit metacarpal length (D4MCL), jari metakarpal keempat ruas phalanak pertama/Fourth digit first phalanax length (D4P1L), jari metakarpal keempat ruas phalanak kedua/Fourth digit second phalanax length (D4P2L), jari metakarpal kelima/Fifth digit metacarpal length (D5MCL), jari metakarpal kelima ruas phalanak pertama/Fifth digit first phalanax length (D5P1L), jari
metakarpal kelima ruas phalanak kedua /Fifthdigit second phalanax length (D5P2L)
Gambar 3 Parameter pengukuran karakteristik luar
Bagian kepala kelawar dibedah untuk mendapatkan ukuran karakteristik tengkorak dan gigi. Kepala direndam menggunakan air basa (deterjen) kemudian
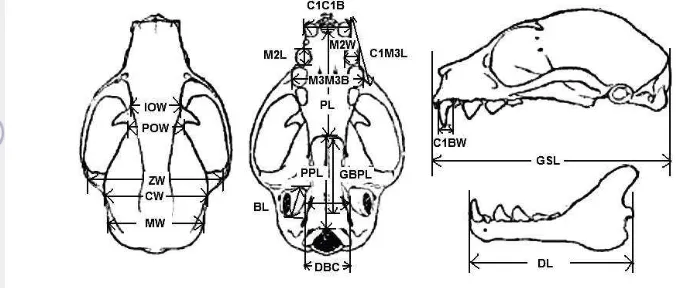
dan 6 peubah karakteristik dental dalam analisis morfometri kelelawar (Suyanto 2001; Nagorsen dan Peterson 1980 diacu dalam Rahman dan Abdullah 2010) (Gambar 4), antara lain :
Keterangan : Karakter Tengkorak= panjang tengkorak total/Greatest skull length (GSL), lebar interorbital/Interorbital width (IOW), lebar posorbital/Postorbital width (POW), lebar tempurung
otak/Carnial width (CW), lebar tulang pipi/Zygomatic width (ZW), lebar mastoid/Mastoid width
(MW), panjang langit-langit tengkorak/Post palatal length (PPL), panjang langit-langit
mulut/Palatal length (PL), jarak antara chochea/Distance between chochlea (DBL), Bulla length
(BL), panjangtulang basal/Greatest basial pit length (GBPL), panjang rahang/Dentary length (DL),
Karakteristik Gigi = ukuran taring/Canine tooth basal width (C1BW), jarak terluar antara gigi
taring/Breadth across both canine outside surface (C1C1B), jarak terluar antara kedua geraham
ketiga/Breadth across both third molar theeth outside surface (M3M3B), jarak terluar antara gigi
taring dan geraham ketiga/Canine molar length (C1M3L), panjang geraham kedua/Second molar tooth crown length (M2L), lebar geraham kedua/Second molar tooth crown width (M2W).
Gambar 4 Parameter karateristik tengkorak dan gigi kelelawar menurut Andersen (1912) diacu dalam Rahman dan Abdullah (2010)
Polen bunga diidentifikasi dari saluran pencernaan dengan mengambil sisa makanan di saluran pencernaan kelelawar. Hasil dari isi pencernaan kelelawar dicampur dengan alkohol 70% di dalam tube sentrufugal kemudian dilakukan
pemutaran hingga 2000 rpm selama 30 menit. Setelah itu, dilakukan penggantian cairan alkohol dengan alkohol yang baru. Proses ini diulang sebanyak tiga kali. Endapan yang dihasilkan dari proses diletakkan pada kaca preparat/object glass
sebanyak satu tetes kemudian dicampur gliserol satu tetes lalu ditutup dengan
cover glass dan direkatkan dengan kuteks bening. Menurut Yulianto (1992) diacu
dalam Soegiharto et al. (2010) gliserol ditambahkan sebagai bahan pengawet.
Obyek diamati dibawah mikroskop cahaya dengan bantuan kamera Optilab Advance. Hasil gambar dikalibrasi dengan bantuan software Image Raster
sehingga mendapatkan ukuran polen yang akurat. Hasil identifikasi jenis digunakan untuk mencari tipe bentuk mahkota bunga (corolla) melalui studi
Metode Pengolahan Data
Hubungan antar jenis kelelawar dengan pakan dan bentuk mahkota bunga
Hubungan antar jenis kelelawar dengan pakan dan bentuk mahkota bunga dianalisis dengan metode cluster variable yang diolah dengan menggunakan software Minitab 16. Pengelompokan dicari menggunakan rumus eudicleaen distance (Krebs 1989) yang kemudian digunakan untuk mengetahui tingkat
kesamaan. Hasil analisis menghasilkan dendrogram yang menunjukkan persentase kesamaan penggunaan sumber pakan baik berdasarkan jenis pakan maupun pemilihan bentuk mahkota bunga oleh kelelawar Megachiroptera.
Karakteristik morfologi kelelawar Megachiroptera
Karakteristik morfologi luar (15 peubah), tengkorak (12 peubah) dan gigi (6 peubah) diolah dengan statistik deskriptif meliputi nilai rataan, standar deviasi dan range. Data statistik deskriptif diolah menggunakan softwareSPSS 16. Hubungan antara ukuran morfologi dengan habitat dan mahkota bunga
Component Principal Analysis (PCA) digunakan untuk mereduksi peubah
yang tidak dapat dijadikan pembeda dalam suatu populasi data. Data yang direduksi adalah peubah yang memiliki ragam paling kecil. Peubah yang dikerjakan dengan PCA adalah ukuran morfologi sayap, tengkorak dan gigi. Pengolahan data dilakukan secara terpisah terhadap masing-masing kelompok peubah morfometri. Analisis PCA terhadap peubah morfometri luar digunakan untuk mengetahui peubah mana yang dapat digunakan sebagai pembeda jenis dalam suatu habitat.
Hasil dari PCA kemudian diolah dengan menggunakan Regresi Logistik Biner. Regresi logistik menjelaskan hubungan antara peubah dikotomik atau biner dengan variabel bebas yang berupa data berskala interval atau kategorik (Hosmer dan Lemeshow 1989). Hasil dari analisis Regresi Logistik menghasilkan peubah-peubah yang dapat dijadikan penciri jenis terhadap suatu habitat.
Hubungan antara peubah morfometri tengkorak dan gigi terhadap pemilihan bentuk mahkota bunga oleh kelelawar Megachiroptera diolah dengan metode
Multivariate Canonical Corespondence Analysis (CCA) menggunakan software
Canoco For Windows 4.5. Menurut Leps dan Smilauer (2003) metode CCA
dengan Canoco For Windows 4.5 digunakan untuk mengetahui hubungan antara
spesies dan lingkungannya (Leps dan Smilauer 2003). Dalam hal ini adalah. Hasil dari pengolahan data berupa gambar panah vektor dan titik yang menunjukkan hubungan antara peubah.
Menurut de Sauza dan Muscheta (1999) mahkota bunga dibagi menjadi dua kelompok berdasarkan penampakan kelopak yaitu, gamopetalouse (bunga dengan
kelopak yang saling bergabung) dan polypetalouse (bunga dengan kelopak yang
saling bebas). Polypetaouse dibedakan menjadi tiga jenis, yaitu rosaceouse
(bunga dengan lima atau lebih kelopak yang menumpuk ke arah luar),
caryophllaceous (bunga dengan lima kelopak atau lebih namun tidak saling
menumpuk) dan yang terakhir adalah papillionaceouse dimana terdapat lima
kelopak yang saling bebas namun memiliki ukuran kelopak yang berbeda. Kelopak posterior berukuran besar disebut vexillium dan dua lainnya mebentuk
Macam-macam jenis bunga menurut Swink dan Willhem (1994) ditampilkan pada Gambar 5.
Gambar 5 Tipe-tipe mahkota bunga
Bentuk bunga lain merupakan hasil modifikasi suatu tanaman. Menurut de Sauza dan Muscheta (1999) bentuk bunga modifikasi ini tidak masuk kedalam kategori. Tiga diantara bentuk modifikasi mahkota ini adalah bentuk apetalouse
dan rosaceous-corona, sepaloid. Apetalouse adalah tipe bunga yang tidak
memiliki mahkota bunga, bunga tipe ini biasanya terdiri dari benang-benang sari yang memanjang dari calyx. Sazima et al. (1999) menggambarkan tipe apetalouse
ini seperti bentuk sikat (Gambar 6).
(a) (b) (c
Gambar 6 Bunga Calliandra sp. (tipe mahkota bunga apetalouse) (a); bunga Annona sp. (tipe mahkota bunga sepalloid) (b); bunga Passiflora sp.
Modifikasi penebalan daging kelopak bunga adalah ciri bentuk sepaloid.
Famili tumbuhan yang memiliki bentuk mahkota bunga ini adalah Annonaceae. Bentuk modifikasi lain adalah rosaceous-corona pada bunga Passiflora sp. yang
mana merupakan gabungan antara rosaceous dan corona dengan modifikasi
sulur-sulur benang sari yang mendominasi.
Kerapatan tegakan
Analisis vegetasi digunakan untuk mengetahui kerapatan pohon pada setiap tanaman. Kerapatan pohon dicari dengan menggunakan petak lingkaran untuk tegakan homogen pada tutupan lahan pinus, puspa, dan damar serta jalur berpetak untuk lahan agroforest. Kerapatan tegakan diolah menggunakan rumus sebagai berikut:
⁄
Kerapatan pohon digunakan untuk mengetahui hubungannya dengan adaptasi morfologi kelelawar terhadap habitatnya. Selain itu jenis-jenis tumbuhan yang berada di sekitar plot pengamatan dicatat sebagai referensi tumbuhan yang diduga dimanfaatkan oleh kelelawar.
Niche overlap
Niche overlap digunakan untuk mengetahui hubungan antara jenis
kelelawar terhadap sumberdaya yang digunakan berdasarkan suku dan jenis tumbuhan yang ditemukan di dalam saluran pencernaan kelelawar. Persamaan yang digunakan adalah persamaan Simplified Morisita Index atau sering disebut Morisita-Horn Index (Krebs 1989).
∑ ∑
∑
Keterangan: CH= Indeks Morisita-Horn antara kelelawar jenis ke-j dan jenis ke-k, pij = proporsi tipe tutupan lahan yang digunakan kelelawar jenis ke-j (pij = n/N), pik= proporsi tipe tutupan lahan yang digunakan kelelawar jenis ke-k (pik = n/N), n= jumlah tipe tutupan lahan.
HASIL DAN PEMBAHASAN
Hasil
Komposis jenis kelelawar Megachiroptera
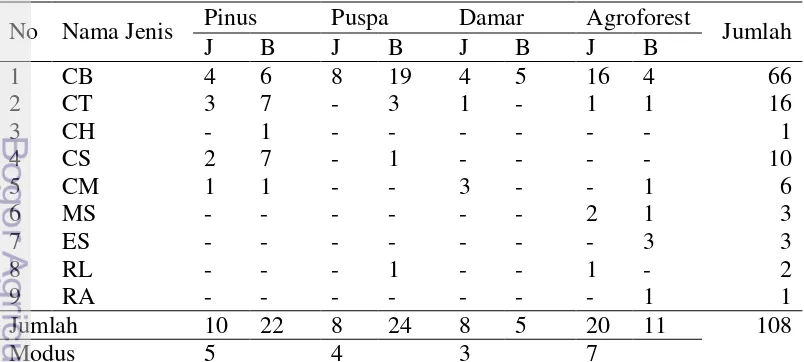
Tercatat sembilan jenis kelelawar Megachiroptera di HPGW kawasan antara lain: Cynopterus bracyothis, C. tithaecheilus, C. horsfieldii, C. sphinx, C. minutus, Macroglossus sobrinus, Eonycteris spelaea, Rousettus leschenaulti, dan R. amplexicaudatus (Tabel 1). Semua kelelawar yang ditemukan belum termasuk
kedalam daftar satwa yang dilindungi.
titthacheilus merupakan jenis yang paling umum ditemukan. C. bracyotis juga
merupakan jenis kelelawar yang paling banyak ditemukan dengna total jumlah tangkapan sebesar 66 ekor.
Tabel 1 Komposisi jenis kelelawar Megachiroptera di HPGW dan status perlindungannya
No Nama Jenis Status Konservasi
PP* IUCN CITES
1 Cynopterus brachyotis (Müller, 1838) - LC n.a.
2 Cynopterus titthaecheilus (Temminck, 1825) - LC n.a.
3 Cynopterus horsefieldi (Gray, 1843) - LC n.a.
4 Cynopterus sphinx (Vahl, 1797) - LC n.a.
5 Cynopterus minutus (Miller, 1906) - LC n.a.
6 Macroglossus sobrinus (K.Andersen, 1911) - LC n.a.
7 Eonycteris spelaea (Dobson, 1871) - LC n.a.
8 Rousettus leschenaulti (Desmarest, 1820) - LC n.a.
9 Rousettus amplexicaudatus (É.Geoffroy, 1810) - LC n.a.
Keterangan : LC= Least Concern, n.a = non appendix, PP*= PP No. 7 Tahun 1999
Pada plot agroforest didapatkan tujuh jenis kelelawar. Jumlah tersebut merupakan jumlah jenis tertangkap yang paling tinggi dari keempat tipe tutupan lahan yang diamati. Jenis kelelawar yang tertangkap di plot ini adalah C. bracyothis, C. tithaecheilus, C. minutus, Macroglossus sobrinus, Eonycteris spelaea, Rousettus leschenaulti, dan R. amplexicaudatus. Pada tutupan lahan
damar menunjukkan jumlah jenis tertangkap yang paling rendah. Tiga jenis kelelawar yang tertangkap di plot ini adalah C. brachyotis, C. titthacheilus dan C. minutus.
Komposisi pakan kelelawar Megachiroptera
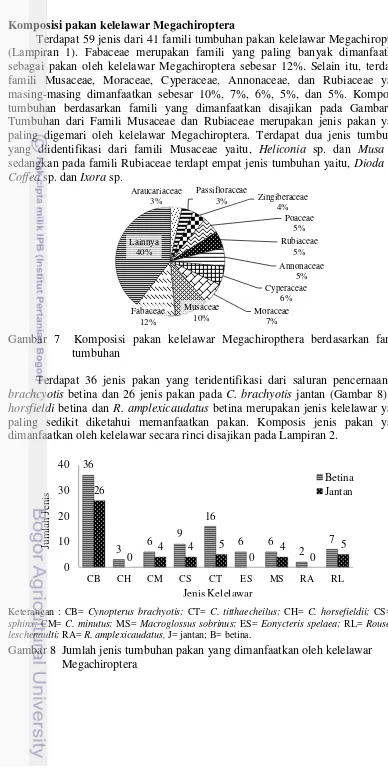
Terdapat 59 jenis dari 41 famili tumbuhan pakan kelelawar Megachiroptera (Lampiran 1). Fabaceae merupakan famili yang paling banyak dimanfaatkan sebagai pakan oleh kelelawar Megachiroptera sebesar 12%. Selain itu, terdapat famili Musaceae, Moraceae, Cyperaceae, Annonaceae, dan Rubiaceae yang masing-masing dimanfaatkan sebesar 10%, 7%, 6%, 5%, dan 5%. Komposisi tumbuhan berdasarkan famili yang dimanfaatkan disajikan pada Gambar 7. Tumbuhan dari Famili Musaceae dan Rubiaceae merupakan jenis pakan yang paling digemari oleh kelelawar Megachiroptera. Terdapat dua jenis tumbuhan yang diidentifikasi dari famili Musaceae yaitu, Heliconia sp. dan Musa sp.
sedangkan pada famili Rubiaceae terdapt empat jenis tumbuhan yaitu, Dioda sp., Coffea sp. dan Ixora sp.
Gambar 7 Komposisi pakan kelelawar Megachiropthera berdasarkan famili tumbuhan
Terdapat 36 jenis pakan yang teridentifikasi dari saluran pencernaan C. brachcyotis betina dan 26 jenis pakan pada C. brachyotis jantan (Gambar 8). C. horsfieldi betina dan R. amplexicaudatus betina merupakan jenis kelelawar yang
paling sedikit diketahui memanfaatkan pakan. Komposis jenis pakan yang dimanfaatkan oleh kelelawar secara rinci disajikan pada Lampiran 2.
Keterangan : CB= Cynopterus brachyotis; CT= C. titthaecheilus; CH= C. horsefieldii; CS= C. sphinx; CM= C. minutus; MS= Macroglossus sobrinus; ES= Eonycteris spelaea; RL= Rousettus leschenaulti; RA= R. amplexicaudatus, J= jantan; B= betina.
Pengelompokan jenis kelelawar berdasarkan jenis pakan
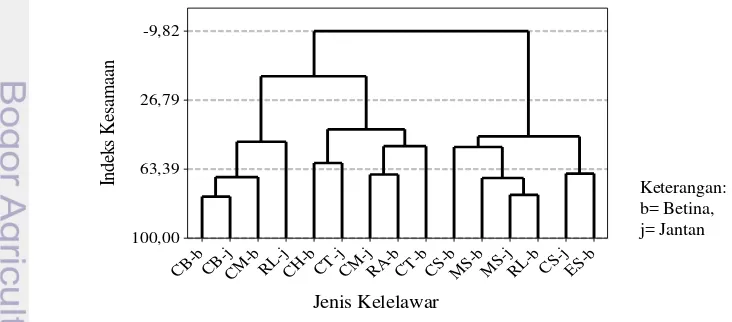
Terdapat tiga kelompok besar berdasarkan pemanfaatan jenis pakan oleh kelelawar Megachiroptera (Gambar 9). Kelompok pertama adalah C. brachyotis
dan C. minutus dengan indeks kesamaan mencapai 68.73%. Kelompok kedua
adalah gabungan dari pasangan C. sphinx dengan E. s spelaea (Ed=71.69%) dan M. sobrinus dengan R. leschenaulti yang (Ed=67.03%) pada tingkat kesamaan
sebesar 49.69%. Kelompok terakhir memiliki jarak terjauh dengan anggota C. horfieldii dan C. titthacheilus (Ed=56.20%) yang berasosiasi dengan R. amplexicaudatus dengan indeks kesamaan hanya sebesar 47.96%
Keterangan : CB= Cynopterus brachyotis; CT= Cynopterus titthaecheilus; CH= Cynopterus horsefieldii; CS= Cynopterus sphinx;CM= Cynopterus minutus; MS= Macroglossus sobrinus;
ES= Eonycteris spelaea; RL= Rousettus leschenaulti; RA= Rousettus amplexicaudatus
Gambar 9 Pengelompokan kelelawar Megachiroptera berdasarkan jenis sumber pakan
Didapatkan tiga kelompok besar kelelawar Megachiroptera berdasarkan pemanfaatan tumbuhan pakan terhadap jenis kelaminnya (Gambar 10). Kelompok pertama beranggotakan C. brachyotis betina dan jantan (Ed=77.83%) yang
berasosiasi dengan C. minutus betina (Ed=67.47%) dan saling berhubungan
dengan R. leschenautii jantan pada tingkat kesamaan mencapai 49.04%.
R. leschenautii betina dan M. sobrinus jantan (Ed=76.78%) dengan M. sobrinus betina (Ed=68.21%) berasosiasi dengan C. sphinx betina sebesar 51.46%. C. sphinx jantan dan E. spelaea betina berasosiasi dengan kesamaan pemanfatan
pakan sebesar 65.76%. Dua kelompok kecil ini membentuk kelompok kedua dengan indeks kesamaan mencapai 45.85%. Kelompok ketiga beranggotakan C. minutus jantan, R. amplexicaudatus betina dan C. ttthacheius betina (Ed=51.24%)
betina yang berasosiasi dengan C. horfieldii betina dan C. titthacheilus jantan
(Ed=60.33%) dengan indeks kesamaan sebesar 42.28%. Hasil perhitungan
analiasis cluster disajikan pada Lampiran 6 dan Lampiran 7
Pengelompokan jenis kelelawar berdasarkan pemilihan bentuk mahkota bunga
Bentuk mahkota bunga yang teridentifikasi sebanyak 13 tipe antara lain,
apetalouse (31%), tubular (13%), savelform (12%), grain (10%), sepalloid (9%), , rosaceous (5%), caryophllaceous (5%), funnelform (4%), rotate (3%), rosaceous -corona (3%), cruciform (2%), bilabiate (2%), dan ligulate (1%) (Gambar 11).
Tipe bunga apetalouse sebagian besar diidentifikasi dari famili Fabaceae dan tubular diidentifikasi dari famili Musaceae. Terdapat tiga jenis tumbuhan pakan
yang termasuk pada famili Fabaceae yaitu Inga sp., Calliandra sp. dan Acacia sp..
Pada studi ini Calliandra sp. merupakan jenis yang paling banyak dimanfaatkan
oleh kelelawar.
Keterangan : Apt= Apetalou, Lig= Ligulate, Bil= Bilabiate, Cru= Cruciform, Fun= Funnelform, Car= Caryophllaceous, Ros= Rosaceous, Rot= Rotate, Svl= Savelform, Tub= Tubular, R-c= Rosaceous-Corona, Sep = Sepalloid, Grn = Grain
Gambar 11 Komposisi pakan kelelawar Megachiroptera berdasarkan bentuk mahkota bunga
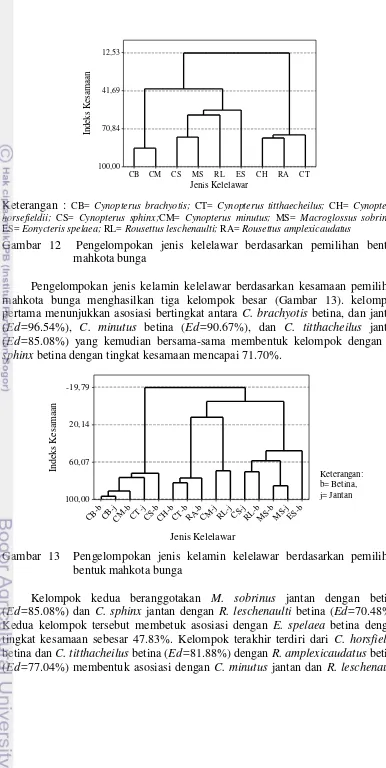
Berdasarkan kesamaan pemilihan bentuk mahkota bunga oleh kelelawar Megachiroptera dikelompokkan menjadi tiga besar (Gambar 12). Kelompok pertama adalah C. brachyotis dan C. minutus dengan kesamaan pemanfaatan
sumber daya pakan mencapai 85.61%. C. horsfieldii dan R. amplexicaudatus
(Ed=78.02%) membentuk kelompok kedua dengan C. titthacheilus dengan tingkat
kesamaan mencapai 75.63%. Kelompok ketiga beranggotakan C. sphix dan Macroglossus sobrinus (Ed=77.38%) yang berasosiasi dengan R. leschenaulti
(Ed=60.20%) kemudian membentuk asosiasi bertingkat dengan E. spelaea dengan
kesamaan pemanfaatan jensi pakan mencapai 56.62% (Lampiran 8).
Keterangan : CB= Cynopterus brachyotis; CT= Cynopterus titthaecheilus; CH= Cynopterus horsefieldii; CS= Cynopterus sphinx;CM= Cynopterus minutus; MS= Macroglossus sobrinus;
ES= Eonycteris spelaea; RL= Rousettus leschenaulti; RA= Rousettus amplexicaudatus
Gambar 12 Pengelompokan jenis kelelawar berdasarkan pemilihan bentuk mahkota bunga
Pengelompokan jenis kelamin kelelawar berdasarkan kesamaan pemilihan mahkota bunga menghasilkan tiga kelompok besar (Gambar 13). kelompok pertama menunjukkan asosiasi bertingkat antara C. brachyotis betina, dan jantan
(Ed=96.54%), C. minutus betina (Ed=90.67%), dan C. titthacheilus jantan
(Ed=85.08%) yang kemudian bersama-sama membentuk kelompok dengan C. sphinx betina dengan tingkat kesamaan mencapai 71.70%.
Gambar 13 Pengelompokan jenis kelamin kelelawar berdasarkan pemilihan bentuk mahkota bunga
Kelompok kedua beranggotakan M. sobrinus jantan dengan betina
(Ed=85.08%) dan C. sphinx jantan dengan R. leschenaulti betina (Ed=70.48%).
Kedua kelompok tersebut membetuk asosiasi dengan E. spelaea betina dengan
tingkat kesamaan sebesar 47.83%. Kelompok terakhir terdiri dari C. horsfieldii
betina dan C. titthacheilus betina (Ed=81.88%) dengan R. amplexicaudatus betina
jantan (Ed=69.37%) dengan tingkat kesamaan pemilihan bentuk mahkota bunga
sebesar 12.10% (Lampiran 9).
Hubungan antara karakteristik morfologi sayap kelelawar terhadap habitat
Komponen pertama menunjukkan bahwa peubah yang digunakan dapat mewakili data sebesar 91.18% (Lampiran 10). Karakteristik luar yang tersisa adalah peubah ukuran sayap. Pada tipe tutupan lahan damar, nilai maksimum
likelihood model reduksi dan model penuh tidak terjadi perubahan sehingga yang artinya tidak ada pengaruh antara peubah bebas secara simultan terhadap peubah tidak bebas (Lampiran 11). Uji Negelkerke menyatakan bahwa sebelas peubah pengukuran morfometri yang diolah sudah menjelaskan keragaman data pada peubah tipe tutupan lahan pinus, puspa, dan agroforest sebesar masing-masing 12%, 16% dan 16% (Tabel 3).
Tabel 3 Hasil uji data peubah morfometri sayap terhadap habitatnya
No Uji Kelayakan Tutupan Lahan
Pinus Puspa Damar Agroforest
1 Omnibus Tes 9.74 13.10 0.00 10.26
2 Negelkerkel Test (%) 0.12 0.16 0.00 0.16
3 Hosmer & Lemeshow Test
*Chi-Square hitung 4.19 3.26 5.48 6.15
perbedaan yang signifikan antara model dengan nilai observasi sehingga model sudah fit dengan data. Hasilnya didapatkan peubah D3P2 sebagai peubah morfometri yang dapat dijadikan penciri pada kelelawar yang ditemukan pada tutupan lahan pinus. Peubah D3P2, D4P2, dan D5P2 pada tutupan lahan puspa, dan D3P1 dan D3P2 pada tutupan lahan agroforestr. Peubah morfometri ini bersama-sama mempengaruhi keberadaan kelelawar pada tipe tutupan lahan yang ada. Tabel variable in equation disajikan pada Lampiran 13.
Kerapatan tegakan tertinggi terdapat pada tegakan pinus dan terendah terdapat pada tutupan lahan agroforest. Kerapatan tegakan disajikan pada Tabel 4.
Tabel 4 Perbandingan kerapatan tegakan pada tiap tutupan lahan Tutupan
Pinus 370 Pinus Kayu afrika, mahoni, puspa
Puspa 257 Puspa Damar, mahoni, kopi
Agathis 308 Damar Puspa, kayu afrika, kopi, meranti Agroforest 52 Puspa Damar, kayu afrika, mahoni, pinus,
Hubungan antara ukuran tengkorak dan gigi kelelawar terhadap tipe bentuk mahkota bunga
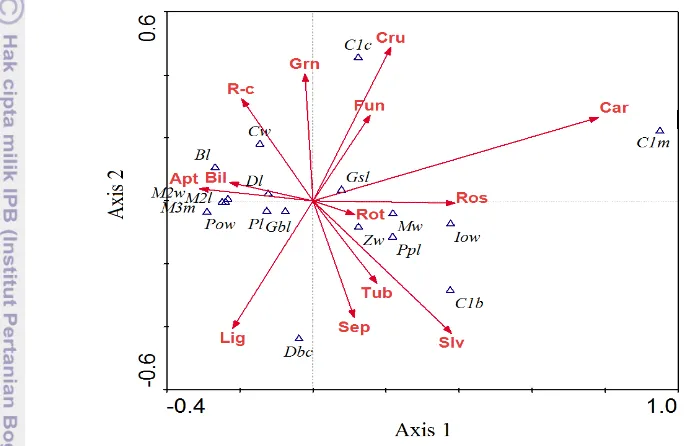
Hubungan antara ukuran tengkorak dan gigi dengan pemilihan bentuk mahkota bunga oleh kelelawar dijelaskan oleh axis 1, axis 2 dan axis 3 yang memiliki nilai total varians sebesar 74.9%. Hal tersebut menunjukkan bahwa model sudah dapat mewakili data sebesar 74.9%. Axis 1 memiliki nilai eigenvalue
sebesar 0.555, axis 2 sebesar 0.630 dan axis 3 sebesar 0.489. Hubungan antara axis 1 dan axis 2 disajikan pada Gambar 14
Keterangan : panjang tengkorak/greatest skull length (Gsl), lebar interorbital/interorbital width
(Iow), lebar postorbital/postorbital width (Pow), lebar carnial/carnial width (Cw), panjang
zigomatik/zygomatic length (Zl), lebar mastoid/mastoid width (MW), panjang post palatal/post palatal length (Ppl), panjang palatal/palatal length (Pl), jarak antara cochleae/distance beetwen cochleae (Dbl), panjang bulla/bulla length (Bl), panjang basial pit terbesar/greatest basial pit length (Gbp), panjang dentari/dentary length (Dl), canine tooth basal width (C1b), breadth across both canine outside surface (C1c), breadth across both third molar theeth outside surface (M3m), canine molar length/maxillary tooth length (C1m), second molar tooth crown length (M2l), second molar tooth crown width (M2w). Apt = apetalouse, Bil = bilate, Lig= ligulate, Sep= sepaloid,
Tub=tubular, Slv=salverform, Rot= rotate, Ros= rosaceous, Car= caryophyllaceous, Fun= funnelform, Cru= cruciform, Spr= spora/grain, R-c= rosaceous-corona.
Gambar 14 Hubungan karakteristik ukuran tengkorak dan gigi terhadap tipe bentuk mahkota bunga pada axis 1 dengan axis 2
Bentuk mahkota bunga caryophyllaceous sangat tergantung dengan ukuran
lebar jarak gigi taring ke geraham ketiga (C1m). Peubah jarak antara gigi taring (C1c) merupakan penciri kuat bagi kelelawar yang memanfaatkan tipe bunga
cruciform dan funnelform. Pemanfatan bentuk mahkota tubular, sepaloid, dan savelform berhubungan dengan ukuran gigi taring (C1b). Ukuran morfometri
tengkorak dan gigi terdapat pada Lampiran 4 dan Lampiran 5.
Ukuran lebar tengkorak (Cw) menjadi penciri bagi kelelawar yang memanfaatkan bentuk bunga rosaceous-corona. M2l, M2w, M3m adalaha peubah
morfometri geraham yang mana berkelompok pada satu titik mendekati panah vektor apetalouse dan bilate. Hal tersebut dapat diartikan bahwa ukuran geraham
bunga apetalouse, bilate dan ligulate. Gamabar axis 1 dan 3 menjelaskan
beberapa variabel yang tidak dapat dijelaskan pada gambar axis 1 dan 2 (Gambar 15).
Keterangan : panjang tengkorak/greatest skull length (Gsl), lebar interorbital/interorbital width
(Iow), lebar postorbital/postorbital width (Pow), lebar carnial/carnial width (Cw), panjang
zigomatik/zygomatic length (Zl), lebar mastoid/mastoid width (MW), panjang post palatal/post palatal length (Ppl), panjang palatal/palatal length (Pl), jarak antara cochleae/distance beetwen cochleae (Dbl), panjang bulla/bulla length (Bl), panjang basial pit terbesar/greatest basial pit length (Gbp), panjang dentari/dentary length (Dl), canine tooth basal width (C1b), breadth across both canine outside surface (C1c), breadth across both third molar theeth outside surface (M3m), canine molar length/maxillary tooth length (C1m), second molar tooth crown length (M2l), second molar tooth crown width (M2w). Apt = apetalouse, Bil = bilate, Lig= ligulate, Sep= sepaloid,
Tub=tubular, Slv=salverform, Rot= rotate, Ros= rosaceous, Car= caryophyllaceous, Fun= funnelform, Cru= cruciform, Spr= spora/grain, R-c= rosaceous-corona.
Gambar 15 Hubungan karakteristik ukuran tengkorak dan gigi terhadap tipe bentuk mahkota bunga pada axis 1 dengan axis 3
Gambar axis 1 dan 3 menjelaskan bahwa ukuran geraham, Pow dan Bl menjadi penciri bagi kelelawar yang memanfaatkan jenis ligulate dan grain
namun hubungannya lemah. Adapun C1m dan C1b berhubungan dengan pemanfaatan bentuk mahkota bunga sepaloid dan tubular, namun tidak sekuat
hubungan dengan keberadaan bentuk mahkota bunga caryophllaceous. C1c dan
Gsl merupakan penciri pemanfaatan bentuk mahkota bunga cruciform dan funnelform namun memiliki hubungan yang lemah digambarkan dengan jauhnya
jarak titik dan panah vektor. Selain itu, karakteristik Iow, C1m, dan Cw memiliki hubungan yang kuat terhadap pemanfaatan bentuk mahkota bunga rossaceous, caryophllaceous dan rosaceous-corona hal tersebut dapat disimpulkan dari hasil
yang pada kedua gambar. Begitu pula dengan ukuran geraham terhadap karakter
apetalouse, bilate dan ligulate.
adalah karakteristik tengkorak kelelawar yang tidak selektif terhadap bentuk mahkota bunga.
Niche overlap
Nilai niche overlap terbesar ditujukkan oleh kelelawar E. spelaea terhadap C. sphinx (CH= 0.45) dan kelelawar R. leschenaulti terhadap M. sobrinus (CH= 0.42) (Tabel 5). Selain itu, R. Leschenaulti memiliki nilai niche overlap terbesar
terhadap C. minutus (CH= 0.38) dan C. bracyotis (CH= 0,28). Pada hasil penelitian
C. brachyotis merupakan jenis kelelawar yang paling banyak memanfaatkan
pakan yang mana memiliki niche overlab terhadap semua jenis kelelawar. Jenis
ini menunjukkan besar pemanfaatan ruang yang tinggi terhadap kelelawar spesialisasi pemanakan buah seperti terhadap C. titthaecheilus, C. sphinx dan R. leschenaulti.
Tabel 5 Matrik niche overlap pada jenis kelelawar Megachiroptera
CB CH CM CS CT ES MS RA RL
Keterangan : CB= Cynopterus brachyotis; CT= Cynopterus titthaecheilus; CH= Cynopterus horsefieldii; CS= Cynopterus sphinx;CM= Cynopterus minutus; MS= Macroglossus sobrinus;
ES= Eonycteris spelaea; RL= Rousettus leschenaulti; RA= Rousettus amplexicaudatus
Persaingan yang terjadi antara individu sejenis terjadi pada individu jantan dan individu betina. C. brachyotis betina dan jantan menunjukkan nilai persaingan
tertinggi yakni sebesar 0.69. Demikian pada M. sobrinus jantan dan betina yang
memiliki nilai niche overlap sebesar 0.45. Matriks niche overlap berdasarkan
jantan dan betina terdapat pada Lampiran 14.
Pembahasan
Komposis jenis kelelawar Megachiroptera
Kelelawar Megachiroptera yang ditemukan di HPGW berjumlah 9 jenis. Survei yang dilakukan Sugiharto et al. (2010) mencatat sebanyak 7 jenis di Kebun
C. bracyotis merupakan jenis yang ditemui hampir pada setiap plot
pengamatan. Menurut Mikleburgh (1992) C. brachyotis merupakan jenis
kelelawar yang dapat bertahan pada berbagai tipe habitat. Berbeda dengan M. sobrinus dan E. spelaea yang cenderung hanya ditemukan pada lahan agroforest. M. sobrinus dan E. spelaea memilih habitat yang memiliki komposisi jenis
tumbuhan berbuah dan berbunga lebih beragam terkait dengan spesialisasinya sebagai pemakan nektar (Prasetya et al. 2011). Hal tersebut sesuai dengan
pernyataan Maryanto dan Yani (2003), Campbell et al. (2007), Maryanto et al.
(2011) yang menyatakan bahwa distribusi kelelawar dipengaruhi oleh keadaan mikro habitat, ketinggian dan kelimpahan tumbuhan pakan.
Lahan agroforest di HPGW memiliki komposisi tumbuhan yang lebih beragam daripada plot pengamatan lainnya. Masyarakat memanfaatkan lahan agroforest yang disediakan oleh pengelola HPGW sebagai lokasi bercocok tanam diantara tegakan pohon. Tanaman yang ditanam oleh masyarakat merupakan tanaman berbuah yang dapat dijual kembali seperti pisang, nangka, rambutan, kopi, singkong, dan lain-lain. Rabenold dan Bromer (1989) menyatakan bahwa perbedaan habitat dapat menimbulkan perbedaan secara fisik, morfometrik, dan perilaku, hal tersebut merupakan hasil dari seleksi alam dimana terdapat hubungan yang kuat antara tumbuhan dan karakter satwa yang berhasil bertahan.
Komposisi pakan kelelawar Megachiroptera
Kelelawar Megachiroptera merupakan satwa yang bergantung dengan keberadaan tumbuhan sebagai pemakan buah, nektar dan polen sehingga distribusi dan kelimpahan serta komposisi dan populasinya dipengaruhi oleh komunitas tumbuhan pada habitatnya (Rabenold dan Bromer 1989). Maryati et al. (2008)
menemukan 21 jenis dari 14 famili tumbuhan pakan di Taman Nasional Gunung Ciremai sedangkan, Sugiharto et. al (2010) menemukan 52 jenis tumbuhan pakan
dari kelelawar yang ditangkap di Kebun Raya Bogor. Hal tersebut menunjukan bahwa penelitian ini memiliki jumlah temuan tumbuhan sumber pakan yang lebih tinggi dari pada dua penelitian sebelumnya. Dalam Maryati (2008) ditemukan bahwa Sapindaceae dan Euphorbiaceae merupakan famili yang paling banyak di manfaatkan oleh kelelawar. Perbedaan temuan dan jumlah pakan kelelawar Megachiroptera ini dipengaruhi oleh beberapa hal. Salah satunya adalah musim buah dan perbedaan tipe habitat pada masing-masing lokasi (Mikleburgh 1992, Tan et al. 2000). Referensi pakan setiap individu kelelawar yang beragam dapat
menjadi faktor pembeda pula. Elangovan et al. 2001 menyatakan selain memakan
buah dan nektar kelelawar juga mengkonsumsi serangga untuk memenuhi kebutuhan nutrisinya dan akan meningkatkan pemangsaan ketika musim kering tiba. C. sphinx selain memanfaatkan nektar dan buah juga memakan daun untuk
memenuhi kebutuhan air, mineral dan karbohidrat (Elangovan et al. 2000)
Fabaceae merupakan jenis yang paling banyak dimanfaatkan oleh kelelawar. Jenis pakan dari famali Fabaceae yang ditemukan adalah Acacia sp., Calliandra
sp., dan Inga sp. Di kawasan HPGW terdapat blok yang didalamnya memiliki
tegakan akasia yang ditanam oleh warga sekitar diduga polen yang ditemukan berasal dari blok tersebut. Tumbuhan kaliandra (nama lokal dari Calliandra sp.)
menyebar secara merata di kawasan HPGW ditandai dengan dijumpainya tumbuhan ini pada setiap plot pengamatan. Calliandra sp. adalah tumbuhan
daun majemuk dan tidak memiliki mahkota bunga (apetalose). Calliandra sp.
merupakan jenis yang paling banyak dimanfaatkan oleh kelelawar.
Tumbuhan dari famili Rubiaceae dan Musaceae merupakan pakan yang paling digemari oleh kelelawar. Enam dari sembilan kelelawar yang ditangkap memanfaatkan pakan dari famili ini. Terdapat empat jenis tumbuhan yang berasal dari famili Rubiaceae yaitu Dioda sp., Coffea sp., Ixora sp., dan Cosmibuena sp.
tumbuhan ini dimanfaatkan oleh C. brachyotis, C. horsfieldii, C. sphinx, C. titthacheilus, M. sobrinus dan R. leschenaulti.
Musaceae dimanfaatkan oleh C. brachyotis, C. sphinx, C. titthacheilus, E.
Maryati et al. 2008, Sugiharto et al. 2010, Edirisinghe dan Kusuminda 2014) Pengelompokan jenis kelelawar berdasarkan jenis pakan
Pengelompokan jenis kelelawar terhadap pakan terjadi dikarenakan referensi pakan yang berbeda-beda. Kesamaan pemanfaatan pakan yang tinggi diantara C. brachyotis dan C. minutus di pengaruhi oleh pemanfatan tershadap
jenis tumbuhan Prestonia sp., Trifolium sp., Utricularia sp., Ficus sp., Heliconia
sp., Calycolpus sp., Cocos sp., dan Adiantum sp. Cynopterus merupakan genus
kelelawar yang lebih adaptif terhadap pakan dan lingkungan. Hal tersebut ditunjukkan dengan jumlah pakan C. brachyotis yang memanfatkan 45 jenis
tumbuhan pakan, C. titthaecheilus memanfaatkan 20 jenis tumbuhan pakan, C. sphinx memanfaatkan 13 jenis pakan. Jumlah ini lebih besar dari pada total
pemanfaatan genus yang lainnya. Diperkuat oleh Fleming et al. (2009) yang
menyatakan bahwa C. brachyotis merupakan jenis kelelawar Megachiroptera yang
komposisi pemangsaan terhadap serangga lebih besar daripada jenis lainnya.
C. horsfieldii dan C. titthacheilus berhubungan atas pemanfaatan jenis Inga
sp. dan Adiantum sp. sedangkan R. amplexicaudatus memiliki kesamaan
pemanfaatan pakan yang lebih tinggi dengan C. titthacheilus atas pemanfaatan Asplenium sp. Pemanfaatan atas jenis Asplenium sp. ini membentuk hubungan
dengan pasangan C. brachyotis dan C. minutus. Bentuk hubungan yang sama
ditunjukkan pula oleh pengelompokan berdasarkan jenis kelamin. C. minutus
jantan, R. amplexicaudatus betina dan C. titthacheilus betina yang membentuk
kelompok atas pemanfaatan Asplenium sp. Asplenium sp. adalah jenis
paku-pakuan yang banyak dijumpai hidup secara epifit pada pohon.
C. brachyotis jantan dan betina, C. minutus betina dan R. leschenaulti jantan
membentuk kelompok atas kesamaan pemanfaatan terhadap jenis Pachystanchys
sp., Prestonia sp., Inga sp., Hisbicus sp., Heliconia sp., dan spesies P. Kelompok
ini memiliki kesamaan pemanfaatan yang tinggi. R. leschenaulti merupakan jenis
kelelawar yang memakan buah namun juga memanfaatkan nektar sebagai pakan dan keberadaannya berhubungan kuat dengan sumber pakan dari famili Musaceae (Edrisinghe dan Kusminanda 2014; Tang et. al 2007; Tang et al. 2010)
Fleming et al. (2009) memasukkan Eonycteris spelaea ke dalam kelompok
kelelawar yang mengunjungi bunga secara oportunis dan Macroglossus sobrinus
bunga yang memiliki kandungan nektar yang tinggi dan mengeluarkan bau khas (Sazima et al. 1999). Pada penelitian ini E. spelaea dan M. sobrinus
memanfaatkan Cyperus sp., Heliconia sp. dan Costus sp. Heliconia sp. dan Costus
sp. memiliki bunga berwarna putih dan mengeluarkan bau yang khas.
Baik dalam pengelompokan jenis maupun menurut jenis kelamin E. spelaea
dan M. sobrinus berada dalam kelompok yang sama. M. sobrinus jantan dan
betina berasosiasi dengan E. spelaea betina, R. leschenaulti betina, dan C. sphinx
jantan serta C. sphinx betina. Survei di Kebun Raya Bogor oleh Kinjo et. al.
(2006) menunjukkan hasil bahwa Eonycteris spelaea dan Macroglossus sobrinus
merupakan jenis yang paling sering mengskses sumber pakan yang sama yaitu jenis dari famili Musaceae.
Pengelompokan jenis kelelawar berdasarkan pemilihan bentuk mahkota bunga
Mahkota bunga merupakan bagian bunga yang paling indah, biasanya tipis dan berwarna-warni yang berfungsi untuk memikat serangga yang membantu dalam penyerbukan (Haryudin dan Rostiana 2008). Sebagian besar pemakan buah dan nektar menggunakan pengelihatan untuk mengidentifikasi keberadaan pakan mereka. Dumont (1997) menyatakan bahwa pemakan nektar memilih pakannya secara spesifik dengan melihat bentuk mahkota bunga. Perilaku kelelawar dalam mencari makan pada malam hari, kebutuhan pakan yang tinggi, mata yang buta warna dan indera penciumannya yang tajam berpengaruh pada bunga yang dipilih (Graham et al. 2003). Hal tersebut menunjukkan adanya hubungan timbal balik
antara bunga dan penyerbuk saling berkaitan.
Kelompok pertama berkorelasi kuat dengan pemanfaatan bentuk mahkota
tubular, salverform, sepaloid, funnelform, cruciform dan caryophyllaceous dan apetalouse. Kelompok pertama ini beranggotakan kelelawar dari genus Cynopterus yang memiliki kesamaan preferensi pakan yang tinggi. Kelompok
kedua berhubungan kuat dengan pemanfaatan bentuk mahkota bunga rosaceouse
dan kelompok ketiga dengan bentuk kelopak bunga rotate. Terdapat enam jenis
pakan yang memiliki tipe bunga rosaceous yaitu Bergenia sp., Schima sp., Schizolobium sp., Hisbiscus sp., Syzygium sp., dan Calyocolpus sp. Bentuk rotate
beranggotakan empat jenis pakan. Touemefortia sp. dari famili Boraginaceae
(rotate) merupakan jenis yang paling banyak dimanfaatkan sebagai pakan oleh
kelelawar.
Betuk apetalouse dan tubuar merupakan bentuk mahkota bunga yang paling
banyak dimanfaatkan oleh kelelawar. Apetalouse merupakan bentuk mahkota
bunga yang tidak memiliki kelopak. Bunga ini biasanya terdiri dari benang sari- benang sari yang bersatu dengan calyx. Tumbuhan pakan dari famili fabaceae
umumnya memiliki bentuk apetalouse. Hal tersebut sesuai dengan Sazima et. al
(1999) yang menyatakan bahwa tipe bunga yang paling digemari oleh kelelawar pemakan nektar dan polen adalah tipe sikat (brush) dan tipe tube (tubular).
Bunga dengan tipe mahkota tubular memproduksi lebih banyak nektar dan
memiliki bau yang atraktif sedangkan tipe mahkota apetalouse banyak nektar dan
memiliki ukuran polen yang relatif besar (Sazima et al. 1999). Sugiharto dan
sporo-pollenin. Lapisan dinding dalamnya (intin) terdiri atas selulosa, pectin, dan nutrisi cytoplasmic.
Hubungan antara karakteristik morfologi sayap kelelawar terhadap habitat
Komponen sayap D3P2, D4P2, D4P1 dan D4P5 merupakan peubah yang dapat dijadikan pembeda jenis ada setiap habitat. Keempat jenis tersebut adalah bagian sayap yang merupakan ruas-ruas jari terakhir. Secara langsung peubah ini mempengaruhi lebar dan panjang sayap total dari kelelawar. Namun demikian bukan berarti peubah lain tidak berpengaruh. Peubah morfologi yang lain tetap berpengaruh namun nilainya sangat kecil.
Model ini hanya mampu menerangkan paling besar 16% dari populasi data dimana 84% dipengaruhi oleh peubah lain diluar data. Dengan kata lain, peubah tersebut hanya mampu mewakili 16% dari model sehingga belum dapat dipastikan bahwa peubah morfometri tersebut dapat dijadikan acuan dalam menilai adaptasi kelelawar Megachiroptera di HPGW.
Komposis tegakan di lokasi tertangkapnya kelelawar diduga dapat menjadi faktor lain yang mempengaruhi keberadaan satwa ini. Apabila dikaitkan dengan kerapatan pohon di lokasi pengamatan, tidak terdapat korelasi yang nyata. Hasil pengamatan menunjukkan terdapat satu peubah adaptif pada plot tutupan lahan pinus yang memiliki kerapatan pohon tertinggi dan dua peubah pada lahan agroforest yang mana menunjukkan nilai kerapatan pohon terendah. Semakin selektif suatu individu ditunjukkan dengan semakin banyaknya peubah morfometri yang adaptif terhadap habitatnya. Hal tersebut tidak sesuai dengan hasil pengamatan apabila dikaitkan dengan nilai kerapatan pohon.
Plot pengamatan pinus terletak diperbatasan kawasan dan plot pengamatan puspa terletak didekat kebun warga. Dari pengamatan langsung, plot yang berbatasan dengan pemukiman warga memiliki komposisi tumbuhan lebih beragam. Sehingga diduga kelelawar yang tertangkap di plot tersebut memanfaatkan habitat sebagai lintasan satwa. Pada tutupan lahan damar tidak ada peubah adaptif yang dihasilkan. Hal tersebut diduga karena letak plot damar yang berada di tengah kawasan dan memiliki potensi tumbuhan pakan yang sedikit.
Namuan demikian, Francis et al. (1990) menyatakan bahwa terdapat
perbedaan ukuran sayap (forearm) pada C. brachyotis yang ditemukan pada hutan
primer dan hutan skunder. Kemudian diperkuat dengan Abdullah (2003) yang menjelaskan perbedaan ukuran dari kelompok C. brachyotis yang ditangkap di
daerah terbuka memiliki ukuran lebih besar dengan C. brachyotis yang ditangkap
dibawah tegakan hutan. Berbeda dengan hasil penelitian terbaru yang menyatakan bahwa ukuran tubuh dan berat total secara negatif berhubungan dengan kemampuan terbang, navigasi dalam menghindari penghalang dan kemapuan manuver terhadap kerapatan area (Rahman dan Abdullah 2011, Jayaraj et al.
Hubungan antara ukuran tengkorak dan gigi kelelawar terhadap tipe bentuk mahkota bunga
Gsl, Pl, Dl dan Gbl adalah ukuran tengkorak yang tidak selektif yang mana pemilihan bentuk mahkota bunga oleh kelelawar tidak dipengaruhi oleh ukuran tersebut walaupun Gsl memiiki hubungan dengan cruciform dan funnelform
namun hubunngannya sangat lemah. Peubah tersebut merupakan peubah morfometri tengkorak yang mempengaruhi lebar dan panjang total tengkorak kelawar.
Ukuran lebar interorbital (Iow) merupakan penciri kuat bagi kelelawar yang memanfaatkan bentuk mahkota bunga rosaceous. Bentuk mahkota bunga inilebih
digemari oleh kelelawar betina antara lain, C. bracyotis (1.48-2.37 mm), C. horsfieldii (2.09 mm), C. minutus (1.38-2.18 mm), C. titthacheilus (2.30-2.58
mm), E. spelaea (1.76-2.31 mm), dan R. leschenaulti (2.44 mm).
Kelelawar yang memanfaatkan tipe caryophllaceous adalah C. bracyotis
jantan (5.13-6.64 mm), C. titthacheilus jantan (6.21-6.95) dan betina (5.91-7.23
mm), C. minutus jantan (5.38-6.16 mm), E. spelaea betina (5.94-7.25 mm), R. amplexicaudatus betina (7.54 mm) dan R. leschenaulti jantan (8.22 mm). Jenis
jenis tersebut memiliki ukuran interorbital yang lebar dan ukuran tengkorak yang relatif besar. Tipe mahkota bunga rosaceous-corona hanya dimanfaatkan oleh C. brachyotis jantan (11.56-13.67 mm) dan betina (11.78-13.98 mm) serta C. minutus jantan 11.96-12.97 mm). Kedua jenis kelelawar ini memiliki ukuran
badan sedang (Suyanto, 2001) hal tersebut ditunjukkan dengan ukuran tengkorak yang lebih kecil dari C. tittacheilus (14.17-16.85 mm) dan lebih besar dari C. horsfieldii (13.25 mm) dan M. sobrinus (11.51 mm).
Ukuran kerakteristik Zw, Ppl dan Mw merupakan karakteristik penduga bagi kelelawar yang memanfaatkan bentuk mahkota bunga rotate. M3m, M2l dan
M2w menjadi karakteristik penduga bagi kelelawar yang memanfaatkan bunga dengan tipe apetalouse. Tipe bunga apetalouse paling banyak dimanfaatkan oleh
kelelawar genus Cynopterus yang memuliki ukuran geraham besar. Hal tersebut
sesuai dengan Dumont (1997) yang menyatakan bahwa ukuran panjang dan lebar geraham mempengaruhi perilaku makan dan mencari pakan. Pada penelitian sebelumnya Mikleburgh (1992), menyatakan bahwa terdapat hubungan yang kuat antara bentuk dan penampilan pakan terhadap pemilihan pakan pada satwa.
Kelelawar yang memanfaatkan bunga bentuk tubular umumnya adalah
kelelawar yang memakan nektar. Namun dalam penelitian ini tipe tubular
dimanfaatkan hampir semua kelelawar yang ditangkap. Jenis pakan yang memiliki bentuk mahkota bunga tubular adalah Quassia sp. Heliconia sp dan Musa sp. Niche overlap
Persaingan dapat didefinisikan sebagai penggunaan sumber daya yang terbatas oleh dua spesies atau lebih (Tarumingkeng 1992). Salah satu cara untuk memahami struktur kominitas adalah dengan mengukur adanya tumpang tindih dalam pemanfaatan sumberdaya diantara spesies yang berbeda pada suatu komunitas guild (Krebs 1989). Guild adalah sebutan untuk suatu komunitas yang
anggotanya memanfaatkan sumberdaya yang sama dengan cara yang sama.
Cynopterus dikategorikan sebagai jenis yang adaptif dalam memanfaatkan pakan
ditandai dengan jumlah pemanfaatan tumbuhan pakan yang tinggi.
Dilihat dari bentuk tengkorak R. leschenaulti memiliki bentuk moncong
(GSL) yang panjang besar (39.78-40.82 mm) dan kuat berbeda dengan M. sobrinus yang memiliki moncong ramping dan kecil (27.54-27.66 mm). Namun
demikian M. sobrinus memiliki nilai niche overlap yang tinggi terhadap R. leschenaulti yang menandakan tingkat pemanfaatan jenis pakan yang tinggi.
Bentuk dan ukuran moncong didukung dengan ukuran lidah yang panjang yang dimiliki oleh M. sobrinus cocok dengan preferensi pakannya sebagai pemakan
nektar (Fleming et al. 2009). Berbeda dengan R. leschenaulti yang termasuk pada
kelelawar pemakan buah namun secara oportunistik memilih bunga sebagai pakan (Fleming dan Muchala 2008). Hasil penelitian menujukkan bahwa M. sobrinus
dan R. leschenaulti menamfaatkan Heliconia sp., Musa sp., Costus sp., dan Cosmibuena sp. sebagai pakan. Adanya tumpang tindih pemanfaatan sumber
pakan pada kelelawar diungkapkan pula oleh Edrisinghe dan Kusminda (2014) yang menyatakan bahwa satu spesies mungkin berbagi pohon pakan yang sama dengan spesies lainnya dalam memanfaatkan sumberdaya pakan.
Tumpang tindih pemanfaatan sumberdaya juga ditunjukkan oleh individu sejenis. Pada individu sejenis persaingan terjadi antara individu jantan dan betina. Persaingan yang paling kuat terjadi antara C. brachyotis jantan dan betiana. C. brachyotis betina bertumpang tindih dalam pemanfaatan sumberdaya pakan
terhadap semua jenis kelelawar yang dimanfaatkan. Hal tersebut ditunjukkan pula oleh jumlah pakan yang ditemukan pada C. brachyotis betina merupakan jumlah
yang paling tinggi. Selain itu pada individu sejenis pesaingan juga terjadi antara M. sobrinus jantan dan betina.
SIMPULAN DAN SARAN
Simpulan
1. Jumlah Jenis pakan kelelawar Megachiroptera yang didapatkan adalah 59 jenis dari 38 famili. Pakan dari famili Fabaceae merupakan jenis pakan yang paling banyak teridentifikasi. Pakan dari famili Musaceae dan Rubiaceae merupakan jenis pakan yang paling digemari.
2. Ukuran morfometri D3P2 dapat dijadikan pembeda jenis kelelawar yang ditemukan pada tutupan lahan pinus; D3P2, D4P2 dan D5P2 dijadikan pembeda jenis kelelawar yang ditemukan pada tutupan lahan puspa; dan D3P1 dan D4P2 dijadikan pembeda jenis kelelawar yang ditemukan pada lahan agroforest. Pada tutupan lahan agathis tidak dapat ditemukan peubah pembeda jenis.
3. Mahkota bunga caryophyllaceous dimanfaatkan oleh kelelawar dengan ukuran
morfometri dental C1M3L (jarak antara gigi taring ke geraham ke-2) pada kelelawar berukuran sedang hingga besar sebagai penciri kuat. Sedangkan bentuk bunga apetallouse lebih fleksibel terhadap karakteristik ukuran gigi
Saran
Perlu adanya penelitian lebih lanjut dalam bentuk explorasi jenis pakan untuk memastikan hasil identifikasi polen yang terdapat di lokasi penelitian. Mengingat peran kelelawar yang sangat penting bagi proses ekologi, maka perlu pembinaan habitat yang lebih intensif sehingga dapat mewujudkan hubungan timbal balik yang menguntungkan dan berkelanjutan antara ekosistem hutan dan keberadaan kelelawar Megachiroptera.
DAFTAR PUSTAKA
Abdullah MT. 2003. Biogeography and variation of Cynopterus brachyotis in
Southeast Asia [PhD thesis]. St. Lucia (AUS) : University of Queensland St Lucia.
Anderson S, Jones JK. 1967. Recent Mammals of The World: A Synopsis of Families. New York (US): Ronald Pr.
Campbell P, Schneider CJ, Zubaid A, Adnan AM, Kunz TH. 2007. Morphological and ecological correlates of coexistence in Malaysian fruit bats (Chiroptera: Pteropodidae). J Mammal. 88: 105–118.
[CITES] Convention on International Trade in Endengered Spesies of Wild Fauna and Flora. 2014. Appendices I, II, and III 14 September 2014 [Internet]. disc-winged bat Thyroptera tricolor (Chiroptera). J Mammal. 87(5):
1013-1019.
Dumont ER, Dávalos LM, Goldberg A, Santana SE, Rex K, Voigt KC. 2011. Morphological innovation, diversification and invasion of a new adaptive zone. Proceedings Of The Royal Society : Biological Science.
Dumont ER. 1997. Cranial shape in fruit, nectar, and exudate feeders:implications for interpreting the fossil record. American J Phys Anthro. 102:187–202.
Edirisinghe WGM, Kusuminda TGT. 2014. On the nectar feeding by the fulvous fruit bat (Rousettus leschenaulti) (Short Communication). Taprobanica: VI
(01) : 66-67.
Elangovan V, Marimuthu G, Kunz TH, 2000. Nectar feeding behavior in the short-nosed fruit bat Cynopterus sphinx (Pteropodidae). Act Chiropterlg. 2:
1–5.
Emmons LH, Feer F. 1997. Neotropical Rainforest Mammals: A Field Guide. 2nd