www.elsevier.com/locate/jinsphys
Time-course haemolymph juvenile hormone titres in solitarious and
gregarious adults of Schistocerca gregaria, and their relation to
pheromone emission, CA volumetric changes and oocyte growth
Amer I. Tawfik
a,1, Karl Treiblmayr
b, Ahmed Hassanali
a,*, E.O. Osir
aaInternational Centre of Insect Physiology and Ecology (ICIPE), P.O. Box 30772, Nairobi, Kenya bInstitute of Zoology, University of Salzburg, Hellbrunnerstrasse 34, A-5020 Salzburg, Austria
Received 28 October 1999; accepted 6 December 1999
Abstract
The haemolymph JH III titres in solitarious and gregarious adult desert locusts, Schistocerca gregaria, were examined in relation to corpus allatum (CA) volumes, aggregation–maturation pheromone production in males and oocyte growth in females. The JH titres of gregarious females were generally higher than those of solitarious females at all ages studied. The titre patterns, however, were similar: relatively high on day 10, dropping to low levels between days 20 and 25, before rising again by day 25. In the solitarious males, the JH titre was very low on day 10 after fledging, but increased gradually and reached a maximal amount on day 30. The JH titre in gregarious males was low on day 10, elevated on day 15 coinciding with the start of the production of the pheromone, and dropped to a relatively low level on day 20 around the time of maximal pheromone production, then rising again by day 25. These results suggest that biosynthesis of the pheromone is associated with a high JH titre peak in the haemolymph. Although a clear relationship was found during the first gonadotropic cycle between JH titres, on one hand, and CA volume and oocyte growth, on the other, in both phases, no such correlation could be discerned in the second cycle.2000 Elsevier Science Ltd. All rights reserved.
Keywords: Phase polymorphism; Corpus allatum; Juvenile hormone; Aggregation pheromone; Desert locust; Schistocerca gregaria
1. Introduction
Two phases of locusts have been distinguished: solita-rious and gregasolita-rious, with a range of intermediate forms between them (Uvarov, 1921; Steedman, 1990). Gre-garisation is a density-driven process associated with improved weather and vegetation, and is accompanied by physiological, behavioural, morphological and pig-mentary changes. The endogenous factors that regulate the transition from a solitary to a gregarious state or vice versa, and then maintain the appropriate phase, have been of considerable interest.
The possible involvement of endocrine factors in phase regulation in locusts has been reviewed exten-sively (Dale and Tobe, 1990; Pener, 1991; Pener and
* Corresponding author. Fax:+254-2-860110/803360.
E-mail address: [email protected] (A. Hassanali).
1 Present address: Zoology/Entomology Department, Faculty of
Science, Assiut University, Assiut 71516, Egypt.
0022-1910/00/$ - see front matter2000 Elsevier Science Ltd. All rights reserved. PII: S 0 0 2 2 - 1 9 1 0 ( 9 9 ) 0 0 2 2 5 - 5
CA into crowded L. migratoria hoppers resulted in a decrease of the E/F (fore-wing to hind-femur length) ratio associated with the solitarious phase. In another study, using radiochemical assay in vitro, Injeyan and Tobe (1981) reported that JH biosynthetic activity of the CA was higher in isolated than in crowded penultimate and last instar female hoppers of S. gregaria. In the same study, the activity of the CA was found to be slightly lower in crowded than isolated adult females, but major differences were temporal. Whereas JH biosynthetic activity of the CA was similar in crowded and isolated
L. migratoria females during the first five or six days
after fledging, by day 8 gland activity was much higher in isolated locusts (Dale and Tobe, 1986). Furthermore, employing the Galleria bioassay, Joly and Joly (1974) and Joly et al. (1977) found that JH titres in the haemo-lymph of fourth and fifth instar hoppers as well as adult females were higher in isolated than in crowded L.
migratoria. On the other hand, allatectomy of gregarious
adult males resulted in complete loss of the yellow col-our associated with gregarious insects, an effect that was reversed by re-implantation of CA or administration of JH (Pener and Lazarovici, 1979). Thus, although age- as well as sex-related differences between phases in CA size and its activity, in haemolymph JH titres, and in effects of external treatment of JH and its analogues, have been implicated in these disparate studies, no coherent picture has emerged (Dale and Tobe, 1990; Pener, 1991; Schneider et al., 1995; Wiesel et al., 1996; Applebaum et al., 1997).
Work at the International Centre of Insect Physiology and Ecology (ICIPE) on the pheromone systems of S.
gregaria has opened up the possibility of comparing
endocrine events with pheromone titres and related phase characters. These studies have demonstrated the existence of distinct sets of releaser pheromones that modulate the aggregation behaviour of different stages of the gregarious desert locust (Obeng-Ofori et al., 1993; Obeng-Ofori et al., 1994a,b; Torto et al. 1994, 1996). The aggregation pheromone produced by older males has also been shown to accelerate maturation of young adults (Mahamat et al., 1993), whereas the nymphal pheromone retards it (Assad et al., 1997). The aggre-gation–maturation pheromone system of adults has been identified as a blend of phenylacetonitrile, benzaldehyde, guaiacol and phenol, with the nitrile present in about 75– 85%. Recently, Deng et al. (1996) showed that locusts are extremely sensitive to crowding. Confining a few or even two solitarious fledglings or mature adults in small cages leads to the production of the adult pheromone within a few days. Conversely, uncrowding gregarious locusts leads to rapid disappearance of the pheromone (Deng et al., 1996). Aggregation–maturation pheromone titres, therefore, constitute a powerful tool for monitor-ing phase and phase shifts.
We undertook the present study with several
objec-tives in mind. First, it was clear that more detailed time-course measurements of haemolymph JH titres of solita-rious and gregasolita-rious S. gregaria adults of both sexes were needed to provide a more rational basis for under-taking experiments involving endocrine interventions and interpreting their results. Second, a major difference between solitarious and gregarious males is that the latter start producing the aggregation–maturation pheromone as they mature (Mahamat et al., 1993); we wondered if this would be reflected in differences in JH titres. Third, we saw the study as an opportune occasion to generate concurrently time-course information on CA volumes in males and females, and on oocyte development in females, to see if these relate to JH titres over the same period on the same group of insects. Herein, we describe our findings.
2. Materials and methods
2.1. Insect cultures
The experiments were conducted using male and female adults of distinctly gregarious and solitary phases of Schistocerca gregaria (Forska˚l) that were cultured at ICIPE in Nairobi (Ochieng-Odero et al., 1994). Insects were reared on a diet of grass and wheat bran at 32±2°C and a 12 h photoperiod as described previously (Tawfik et al., 1996). Adults were staged from the day of final ecdysis and their development was followed for 30 days in rearing rooms that also had other adults of varying levels of maturation. Experimental adults of gregarious phase began mating on day 14, deposited first egg batches on days 15–16 and the second ones on days 26– 28. Corresponding times from the solitary phase were: mating on day 16, first egg-laying on days 18–19, and second egg-laying on days 28–30.
2.2. Identification and quantification of juvenile hormone
the chromatograph by a falling needle injector. For sep-aration of JH derivatives, a fused silica capillary column (J&W DB 1, film thickness 0.25µm, length 30 m, 0.26 mm ID) was used. The injector temperature was set at 300°C and the oven temperature programme moved from 200°C to 290°C (11°C/min). Mass measurement was started 5 min after sample injection, and the differ-ent JHs were eluted within the next 3 min.
2.3. Collection and analyses of volatiles released by gregarious adult males
The collection and analyses of volatiles were carried out as described by Torto et al. (1994). Briefly, air from a compressed air cylinder was passed through a charcoal filter, then over locusts contained in a trapping chamber (10 cm long×3.5 cm ID) and finally through charcoal (120 mg) packed between two glass wool plugs in a glass tube (6 cm long×8 mm ID) at 106 ml/min for 18 h at 32±2°C. Collections were made from sets of nine adult males in groups of three. The adsorbent traps were eluted with dichloromethane and extracts concentrated to 100µl. o-Methyl acetophenone (internal standard, 250 ng) was added. Aliquots (2.5µl) of the extracts were analysed by capillary gas chromatography (GC) on a Hewlett-Packard (HP) 5890 series II gas chromatograph equipped with a flame ionisation detector and an HP capillary column (Carbowax 20 m, 50 m by 0.32 mm ID by 0.3µm film thickness). Nitrogen was used as the car-rier gas at a flow rate of 0.35 ml/min. The oven tempera-ture was programmed for 60°C for 10 min, raised at 5°C/min to 180°C for 5 min and then at a rate of 10°C/min to 220°C, where it was maintained for 15 min. Chromatographic peaks including that of phenylaceton-itrile were integrated using an HP 3396 integrator.
2.4. Morphometric investigations of the corpora allata and ovaries
The volume of the corpora allata was determined immediately after dissection using the formula V (mm3)=4/3
p[(r1+r2)/2]3in which r1and r2are the means (left and right glands) of the larger and the smaller radii, respectively. In each female dissected in Ringer’s sol-ution, 10 terminal oocytes of each ovary were measured using an ocular micrometer at a suitable magnification of a binocular microscope. Thus, 20 terminal oocytes per female were used to calculate the variation in the mean oocyte length during the first and second gonadotropic cycles. Four to five females were used for each age group.
2.5. Statistical analyses
Student’s t-test was used to compare means.
3. Results
3.1. Identification and quantification of juvenile hormone in adult males and females
GC–MS analysis confirmed that only JH III was detectable in the haemolymph of the desert locust S.
gre-garia. JH titres at different ages in adult females and
males for both the solitarious and gregarious phases are shown in Fig. 1.
In females, the pattern of JH titres in the two phases was similar although those in the gregarious insects were mostly higher [Fig. 1(a)]. In both phases, titres were high on day 10 after fledging (80±17 pmol/ml in solitarious insects, 171±34 pmol/ml in gregarious insects). In solita-rious insects, the titres dropped significantly by day 15, fluctuating between 10 and 20 pmol/ml 15–25 days after moult, before rising to about 60±15 pmol/ml by day 30.
Fig. 1. JH titres [±standard deviation (SD)] in the haemolymph of solitarious and gregarious adult females (a) and males (b); and phero-mone titres (±SD) as measured by the amounts of phenylacetonitrile in gregarious (solid circle with continuous line) and solitarious (open circle with broken line) males (*, P,0.05; **, P,0.01; ***,
In gregarious females, JH titres were high on days 10 and 15 after the moult (130–170 pmol/ml), dropped to between 20 and 30 pmol/ml on days 20 and 25, and rose again by day 30 (|85 pmol/ml).
Solitarious and gregarious males had very low haemo-lymph JH titres on day 10 (7.04 and 2.4 pmol/ml, respectively). Thereafter, the pattern of JH titres differed markedly in the two phases. Solitarious adult males showed a gradual increase in JH titres to about 150 pmol/ml by day 30 after fledging [Fig. 1(b)]. How-ever, in gregarious adult males, the titre was conspicu-ously elevated on day 15 (233±29 pmol/ml), coinciding with the start of pheromone emission, and dropped to lower levels on day 20 before rising again by day 25 [Fig. 1(b)].
3.2. Pheromone titres
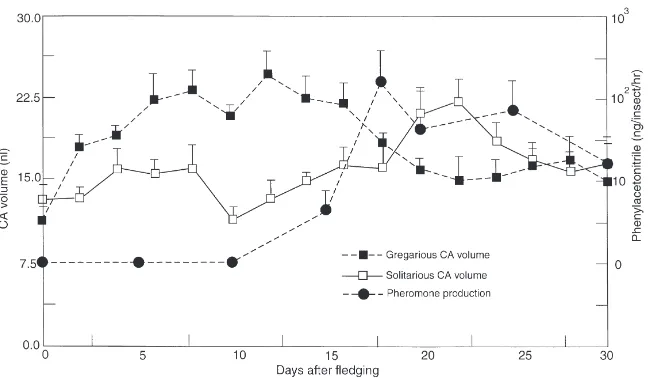
Gas chromatographic analyses of trapped volatiles from different groups of adults confirmed that older males (.10 days after fledging) released a series of aro-matic compounds previously reported, with phenylace-tonitrile comprising 75–85% of the total volatiles (Mahamat et al., 1993; Torto et al., 1994). Maximum titres of this compound occurred between days 18 and 25 after ecdysis. Thus, the start of pheromone production coincided with the first high peak of the haemolymph JH titre [Fig. 1(b)] and reduction in the CA volume (Fig. 3) in gregarious males.
3.3. Changes in the CA volume in adult females during the first and second gonadotropic cycles
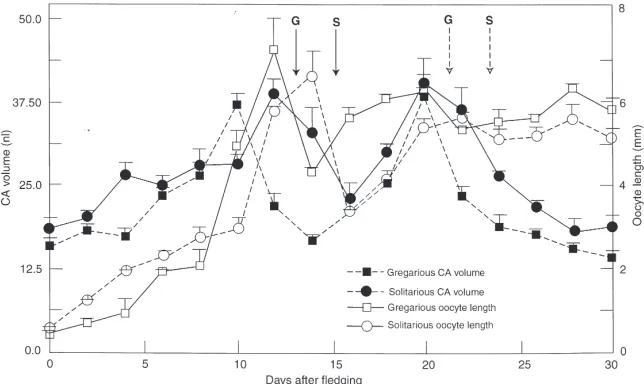
Although the variation in CA volumes was found to be similar in the two phases (Fig. 2), that of gregarious females appeared to peak two days earlier (day 10). This
Fig. 2. Changes in mean volumes (±SD) of the CA of solitarious and gregarious adult females in relation to oocyte growth during the first and second gonadotropic cycles. Arrows indicate time of oviposition in the solitarious (S) and gregarious (G) females.
is in agreement with the JH titre [Fig. 1(a)], which was higher for gregarious females compared with that of soli-tarous. The CA volume of both the solitarious and greg-arious females varied cyclically in relation to growth of the oocytes (Fig. 2). During the first gonadotropic cycle, the maximum length of the basal oocyte peaked just after the first peak of the CA volume (|2 days later) in both phases. This is consistent with a previous observation that it takes 48 h for vitellogenin mRNA to appear fol-lowing administration of precocene-allatectomised L.
migratoria with JH (Wyatt and Davey, 1996).
Interest-ingly, during the second gonadotropic cycle, the CA vol-ume of gregarious females coincided with the maximum length of the basal oocyte, consistent with the previous finding that vitellogenin appears much earlier (2–18 h) on second exposure of JH (Wyatt and Davey, 1996). In contrast, in solitarious females the second peak of the CA volume appeared just before the maximal length of the basal oocyte during the second gonadotropic cycle. Although the volumes of the CA appeared to be larger in the solitarious adult females than in their gregarious counterparts, generally these were either not significant (days 0–8, except day 4) (P.0.05) or, where significant (days 12–20) (P,0.05), the resultant differences were offset by the apparently earlier peak attainment of the CA of gregarious females (day 10 compared with day 12 of solitarious females). However, after day 20, CAs of the solitarious females were somewhat larger.
3.4. Changes in the CA volume in adult males
Fig. 3. Changes in mean volume (±SD) of the CA of solitarious and gregarious adult males, in relation to pheromone production by gregarious adult males.
small peak between day 4 and 8, and then decreased. Thereafter, the CA volume increased, reaching a maximum on day 22 after adult emergence (Fig. 3). The CA volumes were larger in gregarious adult males than in solitarious males from day 2 to 18 (P=0.05–0.0001). In contrast, between days 20 and 24, the CA volumes in the solitarious males were significantly larger than those of their gregarious counterparts (P#0.001).
4. Discussion
Our results from GC–MS analyses of extracts of hae-molymph samples derived from over 150 insects of both phases and sex show that JH III is the only juvenile hor-mone detectable in S. gregaria. This is in agreement with a number of previous findings in Schistocerca (Blight and Wenham, 1976, Trautmann et al., 1976), Locusta (Huibregtse-Minderhoud et al., 1980; Bergot et al., 1981; Pener et al., 1986), and other orthopteroid insects (Loher et al., 1983; Baker et al., 1984; Strambi et al., 1984; Schmidt and Othman, 1993). Studies implicating JH I and JH II in Locusta (Baehr et al., 1979; Fuzeau-Braesch et al., 1982) were based on radioimmunoassays which used reagents that were, in all probability, not suf-ficiently specific (Granger and Goodman, 1983; Tobe and Stay, 1985).
Comparison of the haemolymph JH III titres of adult females of the two S. gregaria phases in the present study [Fig. 1(a)] shows that, in general, those of the gregarious-phase females are higher. This contrasts with previous reports of higher JH titres in solitarious L.
migratoria than in their gregarious counterparts (Joly et
al., 1977; Dale and Tobe, 1986), but confirms obser-vations that differences in CA volumes per se may not accurately reflect differences in JH biosynthetic activity
of the CA (Tobe and Pratt, 1975b; Dale and Tobe, 1986) or in JH haemolymph titres (Dale and Tobe, 1986). The JH titre in the two phases in the first gonadotropic cycle correlates fairly well with the CA volume [Figs. 1(a) and 2] as previously reported for L. migratoria (Ferenz and Kaufner, 1981) and Aiolopus thalassinus (Schmidt and Othman, 1993). Moreover, in the first gonadotropic cycle, the increases in CA volume precede those in oocyte length by about two days in both phases. This is consistent with the previously reported delay between JH application and the onset of vitellogenin synthesis in the first gonadotropic cycle of chemically allatectomised
L. migratoria females (Wyatt and Davey, 1996).
How-ever, during the second cycle from day 15, there was no apparent correlation of JH titres with CA volume or oocyte length in either phase of the insect.
Changes in the CA volume during gonadotropic cycles (Fig. 2) have previously been reported in S.
gre-garia (Highnam, 1962; Highnam et al., 1963; Tobe and
Pratt, 1975a,b; Injeyan and Tobe, 1981), L. migratoria (Highnam and Haskell, 1964) and other acridids (Schmidt and Othman, 1993). The CA volumes of solita-rious females of S. gregaria (Injeyan and Tobe, 1981) and L. migratoria (Dale and Tobe, 1986) were pre-viously reported to be larger than those of gregarious counterparts. Close inspection of our data (Fig. 2) shows that this is probably true only for a brief period (day 4) during the first gonadotropic cycle. At all other times in this cycle, they are similar or just offset because of the apparently faster development of the CA of gregarious females. However, following the peak CA volumes in the second cycle (after day 20), that of solitarious females was indeed larger.
in the present study and in JH biosynthetic activities of the CA of L. migratoria (Tobe and Pratt, 1975b) or in its haemolymph JH titres (Dale and Tobe, 1986)? Second, although JH titres in the first gonadotropic cycles of both phases correlate well with CA volumes and oocyte development, no such correlation is found in the second cycle. In particular, oocyte growth during the second cycle appears to be associated with a low level of JH III. The answer to the first question may lie, in part, in the fact that the earlier work on L. migratoria did not compare JH titres between phases over an extended per-iod. In part, it may be due to phase-related differences in maturation rates in the two species (Pener, 1991) and the possibility that, at specific periods, CA activity levels may be reversed in L. migratoria. A more detailed study of JH titres in this insect may help shed some light on the question. The second question probably reflects our lack of understanding of the way endocrine factors con-trol oocyte development at this reproductive stage of the insect. Is it possible that JH III plays a lesser role in the second gonadotropic cycle and that perhaps another hormone is involved, such as methylated isoprenoids shown to be released by L. migratoria CA (Couillaud et al., 1996)?
In adult male S. gregaria, CA volumes in the two phases show contrasting changes with time, although they start from comparable values (Fig. 3). The increase in CA volume of gregarious males during the first two weeks followed by its decrease was previously reported by Odhiambo (1966). Interestingly, the occurrence of maximal CA size (days 7–16) correlates with the haemo-lymph JH peak and the onset of emission of the aggre-gation–maturation pheromone. This is consistent with previous findings that production of the pheromone is CA-dependent (Loher, 1960; Norris and Pener, 1965; Amerasinghe, 1978). Moreover, in a recent ultra-structural study of the corpora allata of solitarious and gregarious males, Tawfik and Mohammed (1997) found that during the period just preceding the JH peak and the start of pheromone production, CA cells of gregari-ous males were much better developed with numergregari-ous mitochondria and ribosomes compared with those of sol-itarious males. Thus, pheromone biosynthesis and emis-sion in gregarious-phase desert locust adult males appear to be linked to a high concentration of haemolymph JH at an appropriate stage of maturation of the insects.
In an earlier study (Tawfik et al., 1997a), we com-pared the effects of exogeneous JH III on newly emerged adult males and fifth instar nymphs on pheromone pro-duction in maturing adult males. We found that the hor-mone inhibited and/or delayed pherohor-mone production depending upon the method of treatment. Generally, treatments at the nymphal stage resulted in relatively more pronounced effects than at the fledgling stage. These results suggest that the timing and/or duration of the JH peak in the maturing insect is critical in inducing
production of the pheromone. Further studies are needed to throw more light on the question.
In conclusion, the present study and those reported earlier by Tawfik and co-workers (Tawfik et al. 1996, 1999; Tawfik et al., 1997a,b) indicate a need for detailed time-course measurements of endocrine events in locusts and other acridids to help eludicate the physiological mechanisms that underlie phase shifts and phase-related characteristics, including reproductive parameters. Diverse quantifiable phase-related traits (Applebaum et al., 1997; Pener and Yerushalmi, 1998) can now be used accurately to monitor phase changes and the effects of endocrine interventions. Additionally, semiochemical control of maturation rate is now possible (Mahamat et al., 1993; Assad et al., 1997) which would make it poss-ible to differentiate endocrine manifestations of phase from those of maturation. The stage is set for some thoughtful experimentations and, perhaps, fundamental breakthroughs in our understanding of the endocrine control of locust phase dynamics.
Acknowledgements
This work represents part of our work on the desert locust supported by a consortium of donors coordinated by the International Fund for Agricultural Development (IFAD). The funds also supported graduate studies of Amer I. Tawfik. We thank the staff of ICIPE locust col-onies for regular supplies of insects and acknowledge the excellent technical assistance of Bernd Lackner in GC–MS–SIM analyses. We thank two anonymous reviewers for their thorough scrutiny of the earlier ver-sion of the paper, and Professor Lynn Riddiford for reviewing the revised version and insightful suggestions.
References
Amerasinghe, F.B., 1978. Effects of JH.I and JH.III on yellowing, sex-ual activity and pheromone production in allatectomized male
Schi-stocerca gregaria. Journal of Insect Physiology 24, 603–611.
Applebaum, S.W., Avisar, E., Heifetz, Y., 1997. Juvenile hormone and locust phase. Archives of Insect Biochemistry and Physiology 35, 375–391.
Assad, Y.O.H., Hassanali, A., Torto, B., Mahamat, H., Bashir, N.H.H., El Bashir, S., 1997. Effects of fifth-instar volatiles on the sexual maturation of adult desert locust, Schistocerca gregaria. Journal of Chemical Ecology 23, 1373–1388.
Baehr, J.-C., Porcheron, P., Papillon, M., Dray, F., 1979. Haemolymph levels of juvenile hormone, ecdysteroids and protein during the last two larval instars of Locusta migratoria. Journal of Insect Physi-ology 25, 415–421.
Baker, F.C., Lanzrein, B., Miller, C.A., Tsai, L.W., Jamieson, G.C., Schooley, D.A., 1984. Detection of only JH III in several life-stages of Nauphoeta cinerea and Thermobia domestica. Life Sciences 35, 1553–1560.
JH III as the principal juvenile hormone in Locusta migratoria. Experientia 37, 909–910.
Blight, M.M., Wenham, M.J., 1976. Identification of JH III in haemo-lymph from adults and larvae of Schistocerca gregaria. Insect Bio-chemistry 6, 35–38.
Couillaud, F., Mauchamp, B., Girardie, A., 1987. Biological, radioch-emical and physicochradioch-emical evidence for the low activity of dis-connnected corpora allata in locust. Journal of Insect Physiology 33, 223–228.
Couillaud, F., Debernard, S., Darrouzet, E., Rossignol, F., 1996. Hid-den face of juvenile hormone metabolism in the African locust. Archives of Insect Biochemistry and Physiology 32, 387–397. Dale, J.F., Tobe, S.S., 1986. Biosynthesis and titre of juvenile hormone
during the first gonadotropic cycle in isolated and crowded Locusta
migratoria females. Journal of Insect Physiology 32, 763–769.
Dale, J.F., Tobe, S.S., 1990. The endocrine basis of locust phase poly-morphism. In: Chapman, R.F., Joern, A. (Eds.) Biology of Grass-hoppers. John Wiley and Sons, New York, pp. 393–414. Deng, A.L., Torto, B., Hassanali, A., Ali, E.E., 1996. Effects of
shift-ing to crowded or solitary conditions on pheromone release and morphometrics of the desert locust, Schistocerca gregaria (Forska˚l) (Orthoptera: Acrididae). Journal of Insect Physiology 42, 771–776. Ferenz, H.-J., Kaufner, I., 1981. Juvenile hormone synthesis in relation to oogenesis in Locusta migratoria. In: Pratt, G.E., Brooks, G.T. (Eds.) Juvenile Hormone Biochemistry. Elsevier/North-Holland Biochemical Press, Amsterdam, pp. 135–145.
Fuzeau-Braesch, S., Nicolas, G., Baehr, J.-C., Porcheron, P., 1982. A study of hormonal levels of the locust, Locusta migratoria
cineras-cens artificially changed to the solitary state by a chronic CO2
treat-ment of one minute per day. Comparative Biochemistry and Physi-ology 71A, 53–58.
Granger, N.A., Goodman, W.G., 1983. Juvenile hormone radioimmu-noassays — theory and practice. Insect Biochemistry 13, 333–340. Highnam, K.C., 1962. Neurosecretory control of ovarian development in Schistocerca gregaria. Quarterly Journal of Microscopical Science 103, 57–72.
Highnam, K.C., Haskell, P.T., 1964. The endocrine systems of isolated and crowded Locusta and Schistocerca in relation to oocyte growth, and the effects of flying upon maturation. Journal of Insect Physi-ology 10, 849–864.
Highnam, K.C., Lusis, O., Hill, L., 1963. The role of the corpora allata during oocyte growth in the desert locust, Schistocerca gregaria Forsk. Journal of Insect Physiology 9, 587–596.
Huibregtse-Minderhoud, L., Van den Hondel-Franken, M.A.M., Van der Kerk-Van Hoof, A.C., Biessels, H.W.A., Salemink, C.A., Van der Horst, D.J., Beenakkers, A.M.TH., 1980. Quantitative determi-nation of the juvenile hormones in the haemolymph of Locusta
migratoria during normal development and after implantation of
corpora allata. Journal of Insect Physiology 26, 627–631. Injeyan, H.S., Tobe, S.S., 1981. Phase polymorphism in Schistocerca
gregaria: assessment of juvenile hormone synthesis in relation to
vitellogenesis. Journal of Insect Physiology 27, 203–210. Joly, L., 1954. Re´sultats d’implantations syste´matiques de corpora
allata a` de jeunes larves de Locusta migratoria L. Comptes rendus des Sea´nces Societe´ de Biologie 148, 579–583.
Joly, L., Joly, P., 1974. Comparison de la phase gre´gaire et de la phase solitaire de Locusta migratoria migratorioides (Orthopte`re) du point de vue de la teneur de leur he´molymphe en hormone juve´nile. Comptes Rendus Hebdomadaire des Seances de l’Academie des Sciences, Paris 279D, 1007–1009.
Joly, L., Hoffmann, J., Joly, P., 1977. Controˆle humoral de la diffe´ren-ciation phasaire chez Locusta migratoria migratorioides (R. et F.) (Orthopte`res). Acrida 6, 33–42.
Loher, W., 1960. The chemical acceleration of the maturation process and its hormonal control in the male of the desert locust. Proceed-ings of the Royal Society B153, 380–397.
Loher, W., Ruzo, L., Baker, F.C., Miller, C.A., Schooley, D.A., 1983.
Identification of the juvenile hormone from the cricket,
Teleog-ryllus commodus, and juvenile hormone titre changes. Journal of
Insect Physiology 29, 585–589.
Mahamat, H., Hassanali, A., Odongo, H., Torto, B., El Bashir, S., 1993. Studies on the maturation-accelerating pheromone of the desert locust, Schistocerca gregaria (Orthoptera: Acrididae). Chemoecology 4, 159–164.
Norris, M.J., Pener, M.P., 1965. An inhibitory effect of allatectomized males and females on the sexual maturation of young male adults of Schistocerca gregaria (Forsk.) (Orthoptera: Acrididae). Nature 208, 1122.
Nova´k, V.J.A., Ellis, P.E., 1967. The metamorphosis hormones and the phase dimorphism in Schistocerca gregaria. II. Implantation of the glands into hoppers reared in crowded conditions. General and Comparative Endocrinology 9, 477–478.
Obeng-Ofori, D., Torto, B., Hassanali, A., 1993. Evidence for mediation of two releaser pheromones in the aggregation behavior of the gregarious desert locust, Schistocerca gregaria (Forskal) (Orthoptera: Acrididae). Journal of Chemical Ecology 19, 1665– 1676.
Obeng-Ofori, D., Njagi, P.G.N., Torto, B., Hassanali, A., Amiani, H., 1994a. Sex differentiation studies relating to releaser aggregation pheromones of the desert locust, Schistocerca gregaria. Entomolo-gia Experimentalis et Applicata 73, 85–91.
Obeng-Ofori, D., Torto, B., Njagi, P.G.N., Hassanali, A., Amiani, H., 1994b. Fecal volatiles as part of the aggregation pheromone com-plex of the desert locust, Schistocerca gregaria (Forskal) (Orthoptera: Acrididae). Journal of Chemical Ecology 20, 2077– 2087.
Ochieng-Odero, J.P.R., Ndugo, S.M., El Bashir, S., Capstick, P.B., 1994. A method for rearing crowded (gregarious) and isolated (solitary) locusts (Orthoptera: Acrididae) in the laboratory. In: Njagi, P.G.N., Chaudhury, M.F.B. (Eds.) Proceedings of Workshop on Effective Networking of Research and Development on Environmentally Sustainable Locust Control Methods among Locust Affected Countries. ICIPE Science Press, Nairobi, pp. 33–44.
Odhiambo, T.R., 1966. Morphometric changes and the hormonal activity of the corpus allatum in the adult male of the desert locust. Journal of Insect Physiology 12, 655–664.
Pener, M.P., 1991. Locust phase polymorphism and its endocrine relations. Advances in Insect Physiology 23, 1–79.
Pener, M.P., Lazarovici, P., 1979. Effect of exogenous juvenile hor-mones on mating behaviour and yellow colour in allatectomized adult male desert locusts. Physiological Entomology 4, 251–261. Pener, M.P., Yerushalmi, Y., 1998. The physiology of locust phase
polymorphism: an update. Journal of Insect Physiology 44, 365– 377.
Pener, M.P., Dessberg, D., Lazarovici, P., Reuter, C.C., Tsai, L.W., Baker, F.C., 1986. The effect of a synthetic precocene on juvenile hormone III titre in late Locusta eggs. Journal of Insect Physiology 32, 853–857.
Rembold, H., Lackner, B., 1985. Convenient method for the determi-nation of picomole amounts of juvenile hormone. Journal of Chro-matography 323, 355–361.
Rowell, C.H., 1967. Corpus allatum implantation and green/brown polymorphism in three African grasshoppers. Journal of Insect Physiology 13, 1401–1412.
Schmidt, G.H., Othman, K.S.A., 1993. Changes in the size of corpora allata, in the juvenile hormone III titer in the haemolymph, and in the protein content of terminal oocytes throughout the first gonado-tropic cycle in Aiolopus thalassinus (Insecta: Orthoptera: Acrididae). Archives of Insect Biochemistry and Physiology 24, 45–54.
metab-olism in Schistocerca gregaria females. Journal of Insect Physi-ology 41, 23–31.
Steedman, A., 1990. Locust Handbook, 3rd ed. Natural Resources Institute, Chatham.
Strambi, C., Delbecque, J.-P., Connat, J.-L., 1984. Identification by high pressure liquid chromatography and radioimmunoassay of JH-III in Acheta domesticus. Insect Biochemistry 14, 719–723. Tawfik, A.I., Mohammed, M.M., 1997. Ultrastructure of the corpus
allatum in the solitary and gregarious adult male, Schistocerca
gre-garia in relation to pheromone production. Bulletin of the Faculty
of Science, Assiut University 26, 1–18.
Tawfik, A.I., Mat’hova, A., Sehnal, F., Ismail, S.H., 1996. Haemo-lymph ecdysteroids in the solitary and gregarious larvae of
Schisto-cerca gregaria. Archives of Insect Biochemisty and Physiology 31,
427–438.
Tawfik, A.I., Osir, E.O., Hassanali, A., Ismail, S.H., 1997a. Effects of juvenile hormone treatment on phase changes and pheromone production in the desert locust, Schistocerca gregaria (Forskal) (Orthoptera: Acrididae). Journal of Insect Physiology 43, 1177– 1182.
Tawfik, A.I., Vedrova´, A., Li, W., Sehnal, F., Obeng-Ofori, D., 1997b. Haemolymph ecdysteroids and the prothoracic glands in the soli-tary and gregarious adults of Schistocerca gregaria. Journal of Insect Physiology 43, 485–493.
Tawfik, A.I., Vedrova´, A., Sehnal, F., 1999. Ecdysteroids during ovarian development and embryogenesis in solitary and gregarious
Schistocerca gregaria. Archives of Insect Biochemistry and
Physi-ology 41, 134–143.
Tobe, S.S., Pratt, G.E., 1975a. Corpus allatum activity in vitro during ovarian maturation in the desert locust, Schistocerca gregaria. Journal of Experimental Biology 62, 611–627.
Tobe, S.S., Pratt, G.E., 1975b. The synthetic activity and glandular volume of the corpus allatum during ovarian maturation in the desert locust, Schistocerca gregaria. Life Sciences 17, 417–422. Tobe, S.S., Stay, B., 1985. Structure and regulation of the corpus
alla-tum. Advances in Insect Physiology 18, 305–432.
Torto, B., Obeng-Ofori, D., Njagi, P.G.N., Hassanali, A., Amiani, H., 1994. Aggregation pheromone system of adult gregarious desert locust Schistocerca gregaria (Forskal). Journal of Chemical Ecol-ogy 20, 1749–1762.
Torto, B., Njagi, P.G.N., Hassanali, A., Amiani, H., 1996. Aggregation pheromone system of nymphal gregarious desert locust,
Schisto-cerca gregaria (Forska˚l). Journal of Chemical Ecology 22,
2273–2281.
Trautmann, K.H., Suchy´, M., Masner, P., Wipf, H.-K., Schuler, A., 1976. Isolation and identification of juvenile hormones by means of a radioactive isotope dilution method: evidence for JH III in eight species from four orders. In: Gilbert, L.I. (Ed.) The Juvenile Hormones. Plenum, New York, pp. 118–130.
Uvarov, B.P., 1921. A revision of the genus Locusta L. (=Pachytylus
Fieb.), with a new theory as to the periodicity and migration of locusts. Bulletin of Entomological Research 12, 135–163. Wiesel, G., Tappermann, S., Dorn, A., 1996. Effects of juvenile
hor-mone and juvenile horhor-mone analogues on the phase behaviour of
Schistocerca gregaria and Locusta migratoria. Journal of Insect
Physiology 42, 385–395.
![Fig. 1.JH titres [P±standard deviation (SD)] in the haemolymph ofsolitarious and gregarious adult females (a) and males (b); and phero-mone titres (±SD) as measured by the amounts of phenylacetonitrilein gregarious (solid circle with continuous line) and solitarious (opencircle with broken line) males (*, P�0.05; **, P�0.01; ***,�0.001).](https://thumb-ap.123doks.com/thumbv2/123dok/1002041.920773/3.598.310.547.307.673/haemolymph-ofsolitarious-gregarious-phenylacetonitrilein-gregarious-continuous-solitarious-opencircle.webp)