II. TINJAUAN PUSTAKA
2.1 Pengertian Endofit
Mikroba endofit adalah mikroba yang hidup di dalam jaringan tanaman yang tidak terpapar udara dan tidak menginduksi penyakit pada tanaman inang. Mikroba ini tidak menimbulkan penyakit, dan bahkan dapat mensintesis sejumlah alkaloid seperti ergopeptida, loline, lolitrem, dan peramine pada saat terjadi fotosintesis pada tanaman inang. Zat tersebut berfungsi sebagai racun dan atau pertahanan terhadap nematoda, serangga, serta mamalia herbivora. Lolitrem bersifat neurotoksin terhadap mamalia dan dapat mengakibatkan kematian ternak pada padang rumput yang terinfeksi berat oleh endofit (Kemp, 2003 dalam Suada, 2006).
Mikroba endofit hidup di dalam jaringan tanaman pada periode tertentu dan mampu hidup dengan membentuk koloni dalam jaringan tanaman tanpa membahayakan inangnya. Setiap tanaman tingkat tinggi dapat mengandung beberapa mikroba endofit yang mampu menghasilkan senyawa biologi atau metabolit sekunder yang diduga sebagai akibat koevolusi atau transfer genetik (genetic recombination) dari tanaman inangnya ke dalam mikroba endofit (Radji, 2005). Kemampuan mikroba endofit memproduksi senyawa metabolit sekunder sesuai dengan tanaman inangnya merupakan peluang yang sangat besar dan dapat diandalkan untuk memproduksi metabolit sekunder dari mikroba endofit yang diisolasi dari tanaman inangnya tersebut. Dari sekitar 300.000 jenis tanaman yang tersebar di muka bumi ini, masing-masing tanaman mengandung satu atau lebih mikroba endofit yang terdiri dari bakteri dan jamur (Strobel et al., 2003).
Petrini (1992) mendefinisikan endofit yaitu semua organisme yang mengkoloni jaringan tanaman bagian dalam. Sementara Caroll (1988) secara terbatas menggunakan kata endofit untuk merujuk organisme yang menyebabkan infeksi asimptomatik dalam jaringan tanaman, tidak termasuk jamur patogenik seperti mikoriza. Adanya penemuan tentang patogen laten menyebabkan definisi Carroll perlu diperluas yaitu termasuk organisme yang menghuni organ tanaman yang suatu saat dalam hidupnya dapat mengkoloni secara internal jaringan tanaman tanpa menyebabkan kerusakan yang jelas pada inang. Jadi dengan definisi terakhir ini maka endofit mencakup organisme yang sebagian masa hidupnya sebagai epifit dan juga mencakup patogen laten yang tanpa gejala pada inang untuk beberapa lama. Siegel et al. (1990) mengemukakan endofit menunjuk kepada setiap organisme yang hidup dalam tanaman. Contohnya adalah jamur
Acremonium coenophialum pada rumput tall fescue yang menghabiskan siklus
hidupnya dalam jaringan tanaman tetapi meninggalkan inang atau jaringan inang mati untuk bersporulasi.
Jamur endofit adalah jamur yang terdapat di dalam sistem jaringan tumbuhan, seperti daun, bunga, ranting ataupun akar tumbuhan. Jamur ini menginfeksi tumbuhan sehat pada jaringan tertentu dan mampu menghasilkan mikotoksin, enzim, dan antibiotika (Zaifbio, 2009). Petrini et al., (1992) menggolongkan jamur endofit dalam kelompok Ascomycotina dan Deuteromycotina. Keragaman pada jasad ini cukup besar seperti pada Loculoascomycetes, Discomycetes, dan Pyrenomycetes. Strobell et al. (1996), mengemukakan bahwa jamur endofit meliputi genus Pestalotia, Pestalotiopsis,
Monochaetia, dan lain-lain. Sedangkan Clay (1988 dalam Zaifbio, 2009)
melaporkan, bahwa jamur endofit dimasukkan dalam famili Balansiae yang terdiri dari lima genus yaitu Atkinsonella, Balansiae, Balansiopsis, Epichloe, dan
Myriogenospora. Genus Balansiae umumnya dapat menginfeksi tumbuhan
tahunan dan hidup bersimbiosis secara mutualistik dengan tumbuhan inangnya. Dalam simbiosis ini, jamur dapat membantu proses penyerapan unsur hara yang dibutuhkan oleh tumbuhan untuk proses fotosintesis serta melindungi tumbuhan inang dari serangan penyakit, dan hasil dari fotosintesis dapat digunakan oleh jamur untuk mempertahankan kelangsungan hidupnya (Petrini et al., 1992).
2.2 Keberadaan Endofit
Mikroba endofit terdapat pada sebagian besar tanaman, terutama terdiri atas jamur dan bakteri yang hidup interseluler di dalam jaringan tanaman dan merupakan hanya sebagian kecil dari total biomasa tanaman. Jaringan tanaman dapat menjadi inang yang kompleks untuk komunitas mikroba endofit. Umumnya pada jaringan yang sama dapat diisolasi lebih dari satu spesies mikroba endofit. Sampai saat ini belum ada model interaksi yang pasti untuk spesies mikroba endofit. Hubungan antara mikroba dengan tanaman mempunyai kisaran dari simbiosis mutualisme dan komensalisme sampai batas patogen laten (Strobel, 1996).
Arnold (2003) menemukan bahwa makin tua umur daun maka makin banyak jenis endofit yang menghuninya, namun tidak menunjukkan gejala kerusakan pada daun tersebut. Suatu endofit spesies tertentu lebih menyukai
tanaman tertentu pula. Hal ini terbukti suatu spesies endofit dapat tumbuh lebih baik pada media yang mengandung ekstrak tanaman tertentu dibandingkan tanaman lainnya. Kenyataan ini menunjukkan bahwa endofit tidak menginfeksi tanaman secara random saja tetapi juga berperan penting dalam biologi suatu tanaman.
2.3 Kolonisasi Endofit
Beberapa jenis epifit dapat menjadi endofit dalam kondisi ekologi dan iklim yang tepat. Contoh Alternaria alternate, Cladosporium cladosporiodes dan
C. herbarum adalah pengkoloni fitoplane dapat mempenetrasi jaringan daun hidup
pada proses penuaan (senesens). Proses penuaan memodifikasi niche ekologi yang diberi oleh jaringan tanaman kepada true endophyte dengan membiarkan perkembangan organisme yang biasanya lebih baik beradaptasi sebagai saprofit. Perubahan dari epifit ke endofit, memungkinkan jamur epifit melindungi dirinya dari lingkungan tidak menguntungkan (kekeringan, iradiasi) dan sekaligus menghindari aktivitas antagonistik organisme permukaan yang lebih kompetitif.
Kolonisasi endofit pada jaringan tanaman dapat melalui tiga mekanisme yaitu spora airborne yang terbentuk pada inangnya atau sisa-sisa inang. Endofit yang telah diteliti intensif adalah endofit pada rumput-rumputan. Endofit dapat menyebar secara horizontal dan vertikal. Secara horizontal, penyebaran inokulum terjadi melalui udara (airborne) yaitu terbang bersama angin dan jatuh pada permukaan tanaman kemudian tumbuh dan memasuki jaringan tanaman dan berada di antara sel. Penyebaran secara vertikal yaitu endofit yang berada pada
tanaman akan sampai pada biji dan menetap disana, kemudian menyebar dan tumbuh bersama perkecambahan biji dan terus berada pada tanaman keturunannya (Durham, 2003 dalam Suada, 2006). Menurut Arnold (2003), endofit yang menyebar secara horizontal akan menyerupai patogen yaitu menempel dan kemudian tumbuh pada suatu tanaman dan mengambil karbon dari inangnya seperti tidak memberikan pengaruh yang menguntungkan bagi inang tersebut.
2.4 Fisiologi Endofit
Produksi enzim ekstraseluler yang terlibat dalam proses infeksi dan penetrasi diperlukan untuk mendegradasi komponen sel tanaman tertentu adalah sifat umum jamur endofit. Uji melalui penggunaan substrat dan analisis isozim menunjukkan bahwa enzim yang terlibat adalah amilase, selulase, esterase, lipase, ligninase, pektinase dan protease (Sieber et al., 1991 dalam Suada, 2006).
Produksi indole 3-acetic acid (IAA) dan indole-3-acetonitrile telah ditunjukkan in-vitro pada beberapa jamur seperti Aureobasidium pullulans,
Epicoccum purpurascens, Balansia spp. Telah pula dibuktikan bahwa beberapa
endofit yang diuji dapat menghasilkan auksin dan juga sitokinin secara in-vitro. Penelitian menunjukkan bahwa satu strain endofit Hypoxylon serpens yang diisolasi dari tembakau memacu proses pembungaan inang, sementara strain lain menginduksi kelayuan dan menghambat pertumbuhan bibit tembakau (Bergamin-Strotz, 1988 dalam Suada, 2006).
2.5 Beberapa Contoh Jamur Endofit yang Bersifat sebagai Antagonis Mikroba Lain
Bukti pengaruh endofit yang bersifat sebagai antagonis pada mikroba lain telah diketahui pada beberapa tanaman. Beberapa peran jamur endofit seperti pada kakao terhadap serangan Phytophthora sp. menunjukkan bahwa daun kakao yang diinfeksi oleh endofit mengalami kematian 1/3 tanpa endofit oleh Phytophthora. Daun kakao yang diinfeksi endofit menunjukkan gejala serangan Phytophthora
infestans penyebab penyakit hawar daun yang sangat ringan yaitu hanya terjadi
bercak sedikit saja pada tempat infeksi patogen. Sementara daun tanpa endofit menunjukkan serangan patogen yang sangat berat. Antipatogen akibat endofit lebih bersifat lokal dan lebih terekspresi pada daun yang tua (Arnold, 2003).
Isolat dari beberapa spesies jamur endofit yang ditemukan di tanaman padi daerah Wushan, China telah di uji aktivitas antipatogennya secara in-vitro. Beberapa jamur endofit memiliki aktivitas antagonistik terhadap beberapa penyakit pada tanaman padi yaitu Magnaporthe grisea, Xanthomonas oryzae pv.
oryzae. dan Fusarium moniliforme. Jamur endofit pada tanaman padi yang
menunjukkan aktivitas patogennya sekitar 41,2% setidaknya terhadap satu patogen pada tanaman padi. Jamur endofit tersebut antara lain Fusarium sp.,
2.6 Penyakit Blas pada Padi
2.6.1 Gejala dan Akibat Kerusakan Penyakit Blas
Penyakit blas (blast), yang sering disebut ’penyakit pyricularia’, sudah lama dikenal di Indonesia. Tahun 1913 penyakit ini bersama-sama dengan bercak coklat banyak timbul dipesemaian di daerah Surabaya dan Madura, meskipun tidak menimbulkan kerugian yang besar. Blas dikenal di semua negara penanam padi dan dianggap sebagai penyakit padi yang paling penting. Di Cina penyakit ini dikenal sejak abad ke-17, sedang di Jepang pada abad ke-18 (Semangun, 2004).
Gejala penyakit dapat timbul pada daun, batang, bunga, malai, dan biji. Gejala pada daun, yang sering disebut ’blas daun’ (leaf blast), berbentuk bercak-bercak jorong dengan ujung-ujung runcing. Pusat bercak-bercak berwarna kelabu atau keputih-putihan dan biasanya mempunyai tepi coklat atau coklat kemerahan. Bentuk dan warna bervariasi tergantung dari keadaan lingkungan, umur bercak, dan derajat ketahanan jenis padi. Gejala pada daun tua bercak agak kecil dan lebih bulat. Gejala pada serangan Pyricularia bercak-bercak cenderung berkumpul di pangkal helaian daun. Gejala blas yang khas adalah membusuknya ujung tangkai malai yang dikenal dengan nama busuk leher (neck rot). Serangan ini dapat menimbulkan kerugian besar, karena hampir semua biji pada malai itu hampa. Tangkai malai yang busuk mudah patah. Biji yang sakit terdapat bercak-bercak kecil yang bulat (Semangun, 2004).
Penyakit ini menyerang tanaman padi pada beberapa stadia pertumbuhan, mulai dari fase vegetatif sampai stadia pembentukan malai atau generatif.
Serangan yang berat terjadi pada stadia generatif, karena dapat menimbulkan puso dan atau menggagalkan panen (Santika dan Sunaryo, 2008). Penyakit ini telah menurunkan hasil panen padi di Asia Tenggara dan Amerika Selatan sekitar 30-50%, dan mengakibatkan kerugian jutaan dolar Amerika (Shimamoto et al., 2001 dalam Ninasari, 2009). Di Indonesia serangan penyakit blas dapat mencapai luas 1.285 juta ha atau sekitar 12% dari total luas areal pertanaman padi di Indonesia (Balai Besar Penelitian Tanaman Padi, 2010).
2.6.2 Penyebab Penyakit Blas pada Padi
Penyakit blas disebabkan oleh jamur Pyricularia oryzae Cav. Dulu jamur ini juga disebut sebagai P. grisea (Cke) Sacc., namun sekarang keduanya dianggap sebagai jamur yang berbeda (Semangun, 2004). Menurut Dwidjoseputro (1975 dalam Ninasari, 2007) jamur Pyricularia oryzae dapat diklasifikasikan sebagai berikut: Kingdom : Jamur Divisi : Mycota Kelas : Deuteromycetes Ordo : Moniliales Famili : Moniliaciaea Genus : Pyricularia
Spesies : Pyricularia oryzae Cav.
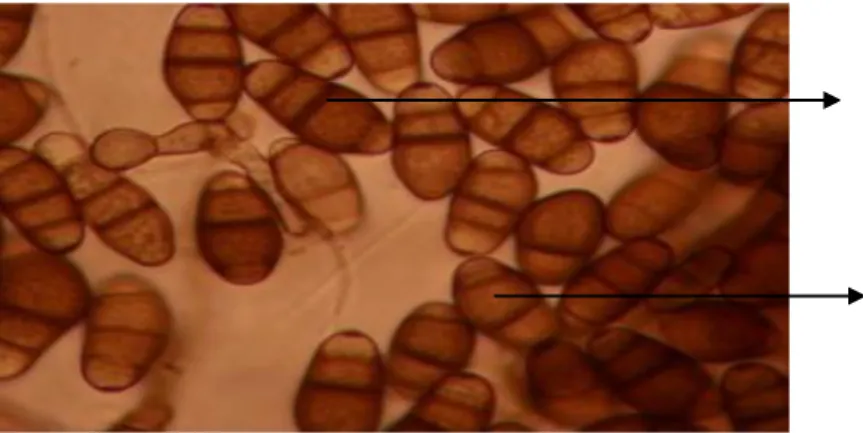
Pyricularia oryzae mempunyai konidiofor bersekat-sekat, jarang
bulat telur dengan ujung runcing, jika masak bersekat dua (Gambar 2.1), dengan ukuran 20-22 x 10-12 μm (Semangun, 2004).
Gambar 2.1. Konidia Pyricularia oryzae Cav. Foto koleksi penulis (2011) pembesaran 1000x
Pyricularia oryzae menghasilkan toxin pyricularian yang memacu
pertumbuhan tanaman tetapi phytotoxic pada konsentrasi yang tinggi. Jamur ini juga menghasilkan enzim-enzim proteolytic yang membantu menembus dinding sel (Singh, 2001 dalam Ninasari, 2007).
Konidianya berbentuk buah alpokat dan bersel tiga, konidia ini dibentuk pada ujung suatu tangkai dan umumnya dilepas pada malam hari saat ada embun atau angin. Jamur ini berkembang biak bila jarak tanam rapat sehingga kelembaban tinggi dan tanaman dipupuk nitrogen secara berlebihan. Penyebaran konidia jamur ini dapat terjadi melalui benih dan angin. Sisa tanaman di lapang dan inang terutama jenis padi-padian yang terinfeksi dapat menjadi sumber penularan bagi pertanaman padi berikutnya.
2.6.3 Daur Hidup Penyakit
Proses infeksi dimulai saat daun dalam keadaan basah dan pada kondisi lingkungan yang mendukung, perkecambahan akan terjadi setelah 3 jam setelah
konidia septum
proses infeksi dimulai. Jika konidia melewati masa kering selama 24 jam maka perkecambahan akan tertunda. Setelah terjadi infeksi hifa akan mempenetrasi melalui epidermis. Kolonisasi tergantung dari salah satu faktor seperti genetik, umur inang, nutrisi, dan faktor lingkungan seperti suhu dan tanah.
Sporulasi terjadi ketika kelembaban di atas 90% konidiofor dibentuk selama 4-6 jam dan satu konidium dibentuk selama 40 menit. Sporulasi maksimum terjadi pada 7-12 hari setelah inokulasi, sporulasi berlanjut sampai 60 hari (Ninasari, 2007).
2.6.4 Faktor-Faktor yang Mempengaruhi Penyakit Blas
Beratnya serangan Pyricularia sangat dipengaruhi oleh faktor luar dan dalam. Kelebihan nitrogen menambah kerentanan tanaman, demikian pula dengan kekurangan air. Diduga bahwa kedua faktor tersebut menyebabkan berkurangnya kadar silisium tanaman. Karena penyakit dibantu oleh kekurangan air, pada umumnya padi tanah kering (gogo) mendapat serangan yang lebih berat daripada padi sawah (Semangun, 2004).
Perkecambahan konidium Pyricularia memerlukan air. Di dalam udara lembab hanya sedikit konidium yang dapat berkecambah. Infeksi blas sangat ditentukan oleh lamanya daun padi basah karena embun. Pada padi gogo daun lebih lama basah karena embun jika dibandingkan dengan padi sawah. Suhu optimum untuk perkecambahan konidium dan pembentukan apresorium adalah 25-30°C. Di negara Jepang, Filipina, dan India telah lama disusun cara peramalan datangnya epidemik blas yang didasarkan atas lamanya daun berembun,
banyaknya konidium di udara, dan dengan memperhatikan keadaan setempat (Semangun, 2004).
Menurut Kuilman (1940 dalam Semangun, 2004) pada masa batang padi tumbuh memanjang (± umur 55 hari) tanaman padi menjadi sangat rentan terhadap infeksi daun oleh Pyricularia. Di antara jenis-jenis padi Indonesia yang diuji ketahanannya di India, Filipina, Jepang, dan Amerika Serikat terbukti bahwa jenis Bengawan mempunyai ketahanan yang cukup baik (Semangun, 2004). Mengingat Pyricularia mudah membentuk ras baru, jenis-jenis padi yang tahan tidak dapat mempertahankan ketahanan dalam jangka waktu yang lama. Bahkan padi yang mempunyai ketahanan horizontal yang dibuat Korea akhirnya menjadi sangat rentan terhadap penyakit ini. Dengan demikian sampai saat ini pembuatan jenis padi yang mempunyai ketahanan yang awet terhadap Pyricularia oryzae masih merupakan tantangan yang berat bagi para ahli penyakit tumbuhan dan ahli pemuliaan (Semangun, 2004).
2.7 Tanaman Padi (Oryza sativa L.)
Padi merupakan tanaman pangan yang berupa rumput-rumputan yang dapat diklasifikasikan sebagai berikut (Anonimus, 2010):
Kingdom : Plantae
Divisi : Spermatophyta Subdivisi : Angiospermae Kelas : Monocotyledoneae
Genus : Oryza
Spesies : Oryza sativa L.
Padi termasuk dalam suku padi-padian atau Poaceae (sinonim: Gramineae atau Glumiflorae). Tanaman semusim, berakar serabut, batang sangat pendek, struktur berupa batang terbentuk dari rangkaian pelepah daun yang saling menopang, daun sempurna dengan pelepah tegak, daun berbentuk lanset. Daun berwarna hijau muda hingga hijau tua, berurat daun sejajar, tertutupi oleh rambut yang pendek dan jarang. Bunga tersusun majemuk, tipe malai bercabang, satuan bunga disebut floret. Setiap bunga padi memiliki enam kepala sari (anther) dan kepala putik (stigma) bercabang dua berbentuk sikat botol. Kedua organ seksual ini umumnya siap reproduksi dalam waktu yang bersamaan. Buah tipe bulir atau kariopsis yang tidak dapat dibedakan mana buah dan bijinya, bentuk hampir bulat hingga lonjong, ukuran 3 mm hingga 15 mm, tertutup oleh palea dan lemma yang dalam bahasa sehari-hari disebut sekam, struktur dominan adalah endospermium yang dimakan orang.
Asal-usul padi budidaya diperkirakan berasal dari daerah lembah Sungai Gangga dan Sungai Brahmaputra dan dari lembah Sungai Yangtse. Di Afrika, padi Oryza glaberrima ditanam di daerah Afrika Barat tropika. Padi pada saat ini tersebar luas di seluruh dunia dan tumbuh di hampir semua bagian dunia yang memiliki cukup air dan suhu udara cukup hangat. Padi menyukai tanah yang lembab dan becek. Sejumlah ahli menduga, padi merupakan hasil evolusi dari tanaman moyang yang hidup di rawa. Pendapat ini berdasar pada adanya tipe padi
yang hidup di rawa-rawa (dapat ditemukan di sejumlah tempat di Pulau Kalimantan).
Dari segi reproduksi, padi merupakan tanaman menyerbuk sendiri, karena 95% atau lebih serbuk sari membuahi sel telur tanaman yang sama. Setelah pembuahan terjadi, zigot dan inti polar yang telah dibuahi segera membelah diri. Zigot berkembang membentuk embrio dan inti polar menjadi endospermia. Pada akhir perkembangan, sebagian besar bulir padi mengadung pati di bagian endospermia. Bagi tanaman muda, pati berfungsi sebagai cadangan makanan. Bagi manusia, pati dimanfaatkan sebagai sumber gizi (Anonimus, 2010).
2.8 Jamur
Dahulu regnum (kingdom) jamur dimasukkan ke dalam regnum plantae, tetapi sekarang jamur berdiri sebagai regnum tersendiri. Jamur adalah mikroorganisme tidak berklorofil, berbentuk hifa atau sel tunggal, eukariotik, berdinding sel dari kitin atau selulosa, bersifat heterotrof, menyerap mutrien melalui dinding selnya dan mengekskresikan enzim-enzim ekstraseluler ke lingkungannya, menghasilkan spora atau konidia, bereproduksi seksual dan aseksual. Sebagian besar tubuh jamur terdiri atas benang-benang yang disebut hifa, yang saling berhubungan menjalin semacam jala yaitu miselium. Miselium dapat dibedakan atas miselium vegetatif yang berfungsi menyerap nutrien dari lingkungan dan miselium fertil yang berfungsi dalam reproduksi (Gandjar, et al., 1999).
Jamur mempunyai empat filum, (Gandjar et al., 2006) yaitu :
1. Filum Chytridiomycota. Filum ini diduga merupakan nenek moyang langsung dari kelompok jamur tingkat tinggi. Diantara angggota jamur, hanya kelompok Chytrid yang memiliki flagella. Chytrid bersifat uniseluler, berkoloni, atau merupakan organisme yang berfilamen yang mengambil nutrient dengan cara absorpsi dan mempunyai sebuah alat gerak yang terletak dibagian posterior, Chytrid demikian disebut zoospore berflagela tunggal. 2. Filum Zygomycota adalah salah satu divisi jamur, terdiri dari dua kelas, yaitu
Trichomycetes dan Zygomycetes. Zygomycota terdiri dari jamur heterotrof yang bereproduksi secara seksual dengan membentuk zigospora. Reproduksi aseksualnya biasanya dengan membentuk aplanospara (sporangiosopora, konidia, sel-sel khamir, artrospora dan klamidiospora), mendapatkan nutrient dengan cara absorpsi. Dinding sel terdiri dari kitosan (pada ordo Mucorales) atau kitin (pada ordo lainnya),
3. Filum Ascomycota mengalami meiosis setelah pembentukan zigot yang berumur pendek dan menghasilkan meiospora dengan pembentukan sel bebas dalam sebuah meiosporangium yang disebut askus.
4. Filum Basidiomycota. Kelompok ini sering disebut jamur oleh orang awam karena banyak jenis-jenis karpusnya (tubuh buahnya) besar dan dapat dilihat dengan kasat mata. Basidiomycota terdiri dari anggota yang makro maupun mikro. Basidiomycota yang mikro adalah basidiokarpnya kecil dan halus, yang kebanyakan patogen pada tanaman dan bersifat saprobik.