1. PENDAHULUAN
LATAR BELAKANG
Ikan patin siam (Pangasionodon hypophthalmus) merupakan salah satu spesies ikan air tawar yang memiliki nilai ekonomis tinggi di Indonesia. Dalam program peningkatan produksi perikanan budidaya tahun 2014, ikan patin menempati urutan ke-3 dengan target peningkatan produksi sebesar 70%/tahun (Kementerian Kelautan dan Perikanan 2010). Berbagai penelitian yang bisa mendukung program Kementerian Kelautan dan Perikanan tersebut telah dilakukan pada bidang nutrisi (Ahmadi 2001; Mokoginta et al. 2000, Suwarsito 2007, Tahapari et al. 2008), reproduksi (Indriastuti 2000) dan lingkungan (Taufik
et al. 2005; Taufik et al. 2007). Sementara itu, penelitian di bidang genetika
dalam rangka memperoleh benih unggul masih dalam tahap permulaan. Program seleksi untuk mendapatkan induk unggul dengan karakter pertumbuhan yang cepat saat ini sedang dilakukan oleh Balai Budidaya Air Tawar Jambi, di bawah Kementerian Kelautan dan Perikanan.
Program seleksi merupakan metode yang efektif untuk meningkatkan produktivitas dan meningkatkan karakter lainnya seperti meningkatkan resistensi penyakit (Gjedrem 1997). Program seleksi secara umum mampu meningkatkan pertumbuhan 6-7% per generasi pada sebagian besar ikan budidaya. Adapun pada beberapa spesies seperti salmon Atlantik, Salmo salar, salmon Coho, channel
catfish, Labeo rohita dan nila, program seleksi dengan menggunakan seleksi
massal maupun famili mampu meningkatkan pertumbuhan 11-14% per generasi (Dunham et al. 2001). Namun demikian metode seleksi memiliki beberapa kelemahan antara lain membutuhkan waktu yang relatif lama dan membutuhkan banyak generasi keturunan. Selain itu seleksi tidak dapat digunakan untuk meningkatkan karakteristik lain yang diinginkan seperti meningkatkan resistensi ikan terhadap suhu dingin (Fletcher et al. 2004).
Perkembangan bioteknologi yang pesat pada saat ini memungkinkan untuk mendapatkan produk perikanan dengan karakteristik yang diinginkan. Transgenesis atau teknologi rekombinan DNA merupakan rekayasa genetik yang
memungkinkan kombinasi ulang (rekombinasi) atau penggabungan ulang gen dari sumber yang berbeda secara in vitro yang kemudian diintroduksikan pada suatu individu. Gen yang telah mengalami rekombinasi ini disebut dengan gen rekombinan atau gen asing. Gen yang diintroduksi atau biasa disebut dengan transgen merupakan pengkode protein tertentu yang mengontrol karakter yang diinginkan dan berguna bagi akuakultur.
Keunggulan teknologi transgenesis dibandingkan teknologi lainnya antara lain yaitu karakter yang diinginkan dapat ditransfer dalam satu generasi dan keunggulan yang didapatkan diturunkan pada generasi selanjutnya (Yaskowiak et
al. 2006). Pelopor transgenesis pada hewan dilakukan pada tikus (dikenal dengan
”super mice”) oleh Palmiter et al. (1982). Penelitian awal pada transgenesis ikan pada saat ini berkembang pada banyak jenis ikan. Pada dua dekade terakhir, paling sedikit 35 jenis ikan diteliti untuk menjadi subjek transgenik terutama melalui perbaikan pertumbuhan melalui transgenesis gen penyandi hormon pertumbuhan (GH, growth hormone).
Devlin et al. (1994) melaporkan bahwa pertumbuhan dapat dipercepat dengan mengintroduksi gen yang mengkodekan GH yang mensintesa peptida GH dalam jumlah yang besar (Over-ekspresi). Transfer gen GH telah diaplikasikan pada beberapa spesies ikan budidaya dan terbukti mampu meningkatkan pertumbuhannya. Over-ekspresi gen GH pada ikan mud loach mampu meningkatkan pertumbuhan lebih dari 32 kali (Nam et al. 2001), pada ikan nila 2-7 kali (Kobayashi et al. 2002-7), salmon Coho 10 kali (Devlin et al. 1995), ikan mas 3 kali (Hinits & Moav 1999) dan salmon Atlantik 2-6 kali (Du et al. 1992) pada generasi kedua.
Secara umum, proses produksi hewan transgenik terdiri dari beberapa tahapan yang dapat diringkas seperti berikut: (1) identifikasi gen yang diinginkan (gen target), (2) isolasi gen target, (3) amplifikasi gen target untuk memproduksi beberapa kopi, (4) penggabungan gen target dengan promoter yang tepat dan sekuens poliadenilasi serta insersi ke dalam plasmid, (5) multiplikasi plasmid dalam bakteri dan purifikasi konstruksi gen untuk injeksi, (6) transfer konstruksi gen ke dalam jaringan resipien, (7) screening keberhasilan integrasi gen eksogen
ke dalam genom resipien, (8) analisis tingkat ekspresi transgen, dan (9) analisis pewarisan transgen pada generasi selanjutnya (Beardmore & Porter 2003).
Pada ikan, pertumbuhan dikontrol antara lain oleh keberadaan hormon pertumbuhan. Hormon pertumbuhan adalah polipeptida yang sangat penting untuk pengaturan pertumbuhan pada vertebrata (Meier et al. 2006). Hormon pertumbuhan pituitari yang juga dikenal sebagai somatotropin pada ikan, merupakan protein kunci yang berperan dalam pengaturan pertumbuhan somatik dan banyak aspek metabolisme lainnya yang terdeteksi pada semua vertebrata (Ryynanen & Primmer 2006). Pada ikan, hormon pertumbuhan terlibat dalam sejumlah proses fisiologi termasuk keseimbangan ionik, metabolisme lipid dan protein, pertumbuhan, reproduksi dan fungsi kekebalan, serta berbagai aspek tingkah laku (Perez-Sanchez 2000).
Secara komersial hormon pertumbuhan berperan penting pada bidang obat-obatan, peternakan, akuakultur dan formulasi pakan hewan, sehingga studi mengenai gen yang mengkodekan hormon tersebut banyak dipelajari pada spesies mamalia dan ikan (Anathy et al. 2001). Dalam akuakultur terapi hormon pertumbuhan dilakukan antara lain melalui penggunaan/pemberian hormon secara langsung pada ikan budidaya atau dapat juga dilakukan dengan produksi ikan transgenik yang telah diintroduksi dengan gen hormon pertumbuhan.
Dalam proses pembuatan ikan transgenik, agar gen yang diintroduksikan dapat aktif maka diperlukan adanya promoter. Promoter adalah sekuens DNA yang memfasilitasi transkripsi gen tertentu. Pemilihan promoter menentukan keberhasilan ekspresi transgen pada ikan transgenik. Promoter dapat bersifat spesifik pada jaringan tertentu. Spesifitas ini ditentukan oleh keberadaan faktor transkripsi yang mempengaruhi ekspresi transgen. Salah satu jenis promoter yang umum digunakan untuk membuat ikan transgenik adalah promoter β-aktin. Menurut Volckaert et al. (1994) promoter β-aktin merupakan promoter yang memiliki keaktifan tanpa adanya rangsangan dari luar sehingga disebut
constitutive promoter. Selain itu promoter β-aktin bersifat ubiquitous artinya dapat
aktif pada semua jaringan otot, serta bersifat house keeping yang dapat aktif kapan saja bila diperlukan.
Promoter β-aktin telah diisolasi dari beberapa jenis ikan dan dilaporkan sebagai regulator dengan aktivitas tinggi dalam mengatur ekpresi transgen pada ikan transgenik. Promoter β-aktin dari ikan medaka mampu mengatur gen penanda LacZ pada embrio ikan medaka (Takagi et al. 1994). Ekspresi gen GFP yang kuat dengan menggunakan promoter β-aktin juga telah ditunjukkan pada ikan medaka (Hamada et al. 1998) dan rainbow trout (Yoshizaki 2001). Selanjutnya promoter ini juga aktif mengatur ekspresi gen penyandi enzim Δ6-desaturase pada ikan zebra (Alimuddin et al. 2005) dan gen penyandi hormon pertumbuhan pada ikan nila (Kobayashi et al. 2007). Promoter β-aktin ikan mas mampu mengatur ekspresi beberapa gen penanda pada beberapa jenis ikan (Liu et
al. 1990). Sementara itu, promoter β-aktin dari ikan zebra dilaporkan aktif
mengatur ekspresi gen GFP pada ikan zebra (Higashijima et al. 1997). Begitu pula dengan promoter β-aktin dari ikan nila mampu mengatur ekspresi gen GFP pada ikan zebra (Alimuddin et al. 2008). Nam et al. (2001) melaporkan bahwa pada ikan mud loach (Misgurnus mizolepis) yang diintroduksi oleh konstruksi gen yang tersusun dari gen GH dan promoter β-aktin yang diisolasi dari spesies yang sama, menunjukkan pertumbuhan yang lebih tinggi dibandingkan non-transgenik.
Sebelum gen target diintroduksikan ke sel inang, gen target dan promoter harus ditempatkan dalam sebuah konstruksi gen. Konstruksi gen berfungsi sebagai kendaraan atau vektor yang membawa gen target ke dalam organisme resipien. Konstruksi gen tersebut memiliki beberapa bagian yang berbeda seperti misalnya yang ditunjukkan pada Gambar 1. Bagian-bagian dalam konstruksi gen antara lain: (1) promoter (P) yang berperan untuk mengontrol aktivitas gen target, (2) gen (cDNA) target, dan (3) sekuens perhentian/terminator (poliadenilasi).
Gambar 1. Peta konstruksi gen dalam bentuk plasmid (P= promoter, RE= enzim restriksi, cDNA= complementary DNA, polyA= poliadenilasi).
Banyak teknik dikembangkan untuk mengintroduksi molekul DNA ke dalam embrio. Pada hewan akuatik, mikroinjeksi merupakan metode yang umum digunakan (Chourrout et al. 1986). Dibandingkan dengan mikroinjeksi, elektroporasi merupakan teknik yang lebih mudah dalam pengerjaannya (Inoue et
al. 1990) karena elektroporasi pada telur hasil fertilisasi dapat memproduksi 10
sampai 100 kali lipat dibandingkan mikroinjeksi (Powers et al. 1992). Elektroporasi adalah proses modifikasi permeabilitas membran sel menggunakan medan listrik. Perubahan permeabilitas bersifat sementara, dengan syarat kejutan listrik tidak melebihi batas kritis bagi sel (Tsong 1983; Serpeusu et al. 1985).
Brackett et al. (1971) pertama kali mendemonstrasikan bahwa sperma kelinci mampu membawa DNA eksogen dalam larutan, dan DNA ini terlokalisasi dalam kepala sperma. Hampir dua dekade berikutnya, Arezzo (1989) dan Lavitrano et al. (1989) membuktikan bahwa sel sperma bulu babi dan tikus dapat digunakan sebagai vektor untuk transfer gen. Sperm-mediated gene transfer (SMGT) merupakan metode yang ideal untuk transfer gen secara massal pada organisme yang fertilisasinya eksternal seperti ikan salmon (Sin et al. 2000).
Keberhasilan transfer gen dengan menggunakan metode elektroporasi melalui sperma telah dibuktikan pada ikan budidaya, antara lain : ikan salmon, zebra, mas, dan nila (diacu dari Spadafora 1998). Penelitian Sin et al. (2000) menunjukkan bahwa kondisi elektroporasi optimal pada sperma salmon adalah pada medan listrik 800 sampai 1000 V/cm, panjang kejutan 27,4 milidetik, dan 2 kejutan. Motilitas sperma pasca elektroporasi bergantung pada medan listrik, panjang kejutan, jumlah kejutan dan kekuatan ionik buffer (Symonds et al. 1994). Berdasarkan penelitian Cheng et al. (2002), motilitas sperma ikan ayu menurun sampai 50% setelah 120 detik ketika dikejutkan dengan voltase 9 kV. Symonds et
al. (1994) juga mendemonstrasikan bahwa aktivitas sperma chinook salmon
menurun dari 82% menjadi 2% pada saat sperma dielektroporasi dengan voltase yang meningkat dari 625 V/cm menjadi 1000 V/cm.
Konsentrasi DNA dalam larutan juga dapat mempengaruhi efisiensi keberhasilan transfer gen (Walker et al. 1995). Konsentrasi DNA bervariasi antara 10 dan 200 µg/ml telah diuji (Sin et al. 1993). Konsentrasi DNA yang tinggi meningkatkan efisiensi pengambilan DNA oleh sperma, dan meningkatkan
efisiensi transfer gen pada telur. Pada kondisi elektroporasi yang optimal, DNA eksogen terdeteksi pada lebih dari 90% embrio ikan salmon yang baru difertilisasi (Walker et al. 1995).
Ekspresi dari gen asing dimulai setelah fase mid-blastula dan levelnya meningkat selama embriogenesis, dan selanjutnya menurun setelah menetas (Gong & Hew 1993; Liu et al. 1990). Kejadian ini disebut sebagai ekspresi sementara (transient expression), yang mungkin disebabkan oleh replikasi ekstrakromosomal DNA asing. Level ekspresi selanjutnya akan menurun yang diikuti dengan degradasi dari ekstrakromosomal DNA. Akibatnya, level ekspresi gen yang terintegrasi ke kromosom resipien tidak setinggi dengan ekspresi sementara. Meskipun hanya beberapa laporan yang menunjukkan integrasi gen dalam genom ikan resipien, teknik ini sangat diperlukan dalam akuakultur (Alimuddin et al. 2003).
Pada beberapa laporan, ekspresi gen dianalisis dengan mengukur level mRNA (messenger RNA) dan protein. mRNA dari gen asing dapat dideteksi dengan menggunakan probe (fragmen DNA yang diberi label radioaktif, biasanya berupa 35P) dan protein dengan cara immunodeteksi dengan menggunakan antibodi. Akan tetapi kedua metode ini membutuhkan banyak waktu dan relatif kompleks. Oleh karena itu, untuk mengembangkan promoter/enhanser yang baik diperlukan suatu metode yang sederhana dan cepat untuk mendeteksi ekspresi gen yang dikendalikannya (Alimuddin et al. 2003).
Pada penelitian ini, peningkatan pertumbuhan ikan patin siam akan dilakukan melalui rekayasa genetika yaitu dengan mengintroduksikan gen GH. Konstruksi gen yang digunakan adalah all fish yang tersusun dari gen GH yang berasal dari ikan patin siam (PhGH) dan promoter β-aktin dari ikan mas (pCcBA). Transfer gen PhGH dilakukan dengan menggunakan metode elektroporasi pada sperma ikan patin siam yang berperan sebagai media pembawa transgen, sebelum digunakan untuk membuahi telur. Diharapkan gen PhGH yang ditransfer mampu terinsersi dan terekspresi pada ikan patin siam.
TUJUAN DAN MANFAAT
Penelitian ini bertujuan untuk menghasilkan ikan patin siam (Pangasionodon hypophthalmus) transgenik F0 melalui over-ekspresi gen penyandi hormon pertumbuhan. Secara khusus penelitian ini bertujuan untuk: 1. Mendapatkan konstruksi gen hormon pertumbuhan all fish yang tersusun dari
promoter β- aktin ikan mas dan gen GH dari ikan patin siam.
2. Mendapatkan kondisi elektroporasi yang optimal pada sperma ikan patin siam.
3. Mendapatkan informasi tingkat ekspresi gen PhGH pada juvenil ikan patin siam.
4. Mendapatkan individu transgenik F0
Manfaat penelitian ini antara lain yaitu untuk menyediakan kandidat ikan patin siam (Pangasionodon hypophthalmus) transgenik F0 sebagai bahan untuk menghasilkan ikan transgenik stabil yang tumbuh cepat.
KEBAHARUAN PENELITIAN
Kebaharuan (novelty) dalam penelitian ini adalah: (1) Konstruksi gen hormon pertumbuhan all fish yang tersusun dari promoter β- aktin ikan mas dan gen GH dari ikan patin siam, (2) Kondisi elektroporasi yang optimal untuk sperma ikan patin siam, (3) Tingkat ekspresi gen PhGH eksogen pada juvenil ikan patin siam dan (4) Individu transgenik F0.
RUANG LINGKUP PENELITIAN
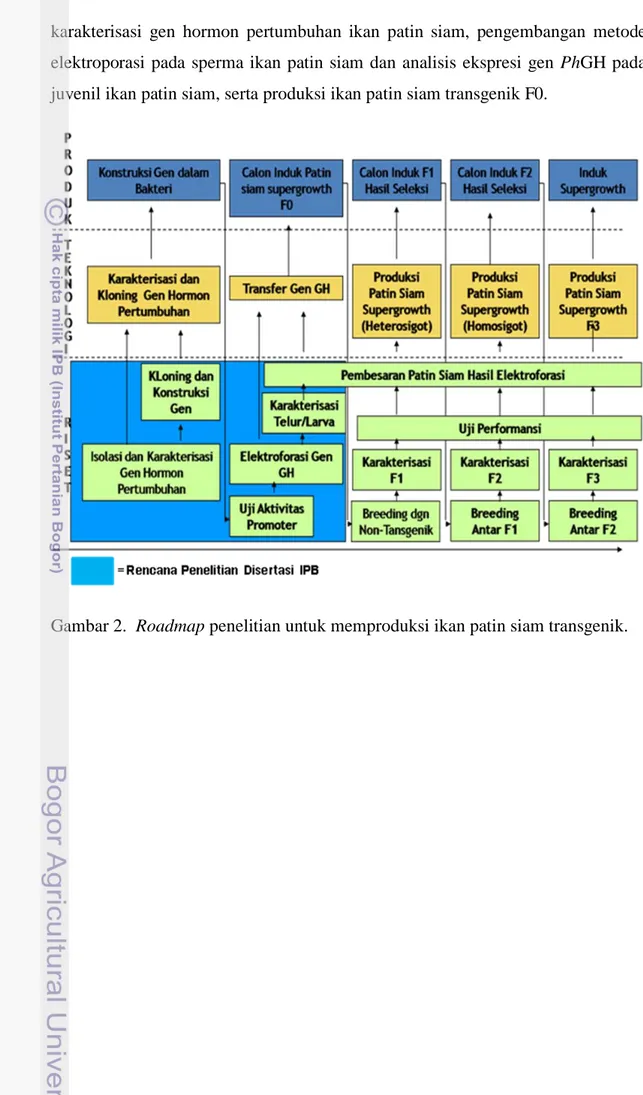
Penelitian ini dilakukan berdasarkan roadmap produksi ikan patin siam transgenik (Gambar 2). Untuk mendapatkan induk transgenik dengan pertumbuhan cepat dan stabil dalam mentransmisikan transgen diperlukan waktu relatif lama sehingga disertasi ini difokuskan pada beberapa tahapan awal penelitian. Penelitian ini dilakukan dalam tiga tahap penelitian yaitu: kloning dan
karakterisasi gen hormon pertumbuhan ikan patin siam, pengembangan metode elektroporasi pada sperma ikan patin siam dan analisis ekspresi gen PhGH pada juvenil ikan patin siam, serta produksi ikan patin siam transgenik F0.