The lectin recognition hypothesis proposes that plant lectins mediate specificity in the Rhizobium–legume symbiosis. Although the hypothesis was developed eight years before nod genes were identified in rhizobia and sixteen years before Nod factor was shown to be a major determinant of host specificity, experiments performed recently using transgenic lectin plants support its main tenets.
Addresses
Department of Molecular, Cell and Developmental Biology, and Molecular Biology Institute, University of California, Los Angeles, CA 90095-1606, USA; e-mail: [email protected]
Current Opinion in Plant Biology1999, 2:320–326 http://biomednet.com/elecref/1369526600200320 © Elsevier Science Ltd ISSN 1369-5266
Abbreviations
Ccd cortical cell divisions CPS capsular polysaccharides EPS exopolysaccharides Had root hair deformation LPS lipopolysaccharides PRA peanut root lectin PSL Pisum sativumlectin SBL soybean lectin
Introduction
The lectin-recognition hypothesis, championed by Hamblin and Kent [1], Bohlool and Schmidt [2], and Dazzo and Hubbell [3], was formulated to explain host specificity between legume and rhizobial symbiotic part-ners. It was based on the strong correlation between the inoculation specificity of bacteria of the family Rhizobiaceae on their legume hosts and the ability of host-produced lectins to bind to Rhizobium cells, and soon became accepted as ‘dogma’. The concept and the dia-grammatic scheme shown in Figure 1 were incorporated into numerous textbooks, and although there were excep-tions to the idea that plant lectins mediated specificity, the hypothesis has held for more than 20 years.
In this review, I will give a brief overview of the lectin recognition hypothesis, the experiments that confirm it, and offer some suggestions as to future research directions.
Why lectins?
Lectins were originally defined as carbohydrate-binding pro-teins of non-immune origin that agglutinate cells and/or precipitate glycoconjugates [4]. This definition has been modified for plant lectins to include those proteins possess-ing at least one noncatalytic domain that binds reversibly to a specific mono- or oligosaccharide [5]. Most plant lectins are multivalent. Some also bind small hydrophobic molecules
such as adenine and derivatives, the cytokinins, which func-tion as plant growth regulators [6,7].
Lectins correlate with the cross-inoculation groups estab-lished by their source legume [8]. Rhizobia that nodulate
Phaseolus vulgariswere found to interact with bean lectin [1], whereas soybean lectin bound and aggregated 22 of 25 strains of Bradyrhizobium (previously known as
Rhizobium) japonicum that nodulate soybean [2]. Twenty-three other bradyrhizobial strains that were not aggregated by SBL (soybean lectin) did not initiate nod-ule formation on soybean.
The lectin recognition hypothesis was formulated many years before the isolation and characterization of nodgenes in rhizobia as well as before Nod factor was recognized as mediating legume-Rhizobiumspecificity. Generic Nod fac-tor is an N-acetyl-glucosamine oligomer (with a backbone of three to five glucosamine units) and an acyl tail of dif-fering lengths and saturation, depending on the rhizobial species. Various other substituents can be present on the reducing end [9] (see WJ Broughton and X Perret, this issue pp 305–311). Specificity resides in both the reducing and non-reducing ends of the molecule. On alfalfa, Nod factor alone, at a very low concentration (10–12M), elicits root hair deformation (Had), whereas a higher concentra-tion (10–7M) is required for extensive cortical cell divisions (Ccd) [10]. Although many legumes undergo the Had
Role of lectins (and rhizobial exopolysaccharides) in
legume nodulation
Ann M Hirsch
Figure 1
The lectin cross-bridge model of Dazzo and Hubbell [3], showing how clover lectin links the cross-reactive antigens of the rhizobia with those of the plant.
Root hair wall
Cross-reactive antigen Clover
lectin
R. trifolii
Root hair cytoplasm
response in response to Nod factor, only a few (alfalfa,
Glycine soja, Phaseolus, Acacia, Lotus corniculatus) undergo Ccd [10–12,13••].
One of the advantages of studying lectins is that the char-acteristics of these proteins, as well as the oligosaccharides that bind to them, are well known. On the basis of its structure, Nod factor is thus an unlikely ligand for a typi-cal legume lectin like SBL or Pisum sativum lectin PSL. In contrast, a novel 46 kDa root lectin isolated from Dolichos bifloros is shown to bind to Nod factors produced by rhi-zobia that nodulate this legume [14•]. It has no similarity in sequence to any other lectins yet described. Moreover, it functions as an apyrase catalysing the hydrolysis of phosphoanhdride bonds of nucleoside di- and triphos-phates [14•]. A Nod factor-binding site, enriched in plasma membrane fractions isolated from Medicago cell suspension cultures has also been reported (see Note added in proof). However, as yet, no correlation has been made with the loss of function of a particular binding pro-tein and a non-nodulation locus, so it is not clear yet whether or not these binding proteins/sites are essential for the symbiotic interaction.
Lectins are found in the right place and at the
right time
Most plant lectins are secreted proteins and are found in vacuoles, cell walls, or intercellular spaces. Although they are found in just about every plant organ, their distribution in root tissue gives the most support to the lectin recogni-tion hypothesis — lectins are localized to root hairs, which are the sites of rhizobial entry for many legumes [15–20]. They are most frequently found at the tips of developing root hairs, although Díaz et al. [18] localized PSL to root hair precursor cells.
Root and seed lectins can be the product of a single gene. This is the case for soybean (Glycine max) [21] and its lectin SBL (or SBA) and pea (Pisum sativum)and its lectin PSL (or PSA) [22]. DNA gel blot hybridization and S1 nuclease analysis and ribonuclease protection experi-ments (used for fine-structure mapping and accurately determining the steady-state level of any mRNA) failed to identify any functional lectin genes in soybean other than Le1, the gene encoding SBL [21]. Although several papers suggested that there is a separate soybean root lectin, we were unable to find one. Our attempts to iso-late a root protein that was reported to cross-react to an antibody made to SBL failed. Although we used the same antibody as Vodkin and Raikhel [23], the protein that we isolated had a high degree of sequence similarity to the acidic ribosomal protein P0 [24]. Thus, if there is a root lectin in soybean, it must be quite different from the pro-tein encoded by Le1. The presence of another lectin may also explain why ‘Sooty’, a soybean cultivar with a mutant lectin gene (le1) nodulates [25], and why root exudate from Sooty reverses the delayed nodulation phenotype of
B. japonicum strain HS111 in the same way as the lectin
derived from Le1soybeans [26]. Another lectin composed of 45 kDa subunits with specificity for 4-O -methylglu-curonic acid [27,28] has been detected in soybean, but the gene encoding it remains unidentified. However, ‘Sooty’ has been tested only with high concentrations of inoculum (S Pueppke, personal communication). Will it nodulate with a below-threshold number of bradyrhizobia or with bradyrhizobia that do not attach properly (see later section)?
What bacterial component binds to
legume lectins?
Rhizobia are typical Gram-negative bacteria with a cyto-plasmic and outer membrane enclosing a pericyto-plasmic space. Much of their outer surface is composed of polysaccharides: lipopolysaccharides (LPS), capsular polysaccharides (CPS or KPS), and acidic exopolysaccharides (EPS). Mutants that are defective in the production of these polysaccharides are often blocked in inducing nodule development, particular-ly in steps involving nodule invasion via infection threads. A considerable amount of data show that rhizobial surface polysaccharides bind to some lectins.
Although not specifically identifying the factor that binds to lectin, Bhuvaneswari and Bauer [29] found that culture conditions for B. japonicum influenced the expression of surface receptors for SBL and that these potential lectin receptors were transiently expressed. Dazzo and Truchet [30] found similar results. The tran-siently expressed lectin receptors were found to be acidic and neutral polysaccharides or lipopolysaccha-rides, depending on the rhizobia.
An alfalfa lectin isolated from seed extract by Kamberger [31] was found in agarose gel diffusion tests to interact with
Rhizobium(Sinorhizobium) meliloti LPS, but not with LPS from heterologous rhizobia. Jayaraman and Das [32] reported that the peanut root lectin (PRA II) bound to LPS of peanut-specific bradyrhizobia, and not to other
Bradyrhizobiumspecies.
New life for the lectin recognition hypothesis
The experiments of Díaz et al. [33] revitalized interest in the lectin recognition hypothesis. Transgenic white clover roots carrying the pea lectin gene were nodulated by the heterologous strain R. leguminosarum bv. viciae, whereas no nodules formed on clover plants that lacked the pea gene. Altering the carbohydrate-binding domain of the introduced lectin by site-directed mutagenesis demonstrated that lectin mediated this heterologous interaction. When transgenic plants carrying the mutated, transgenic lectin were inoculated with the heterologous rhizobia, no nodules formed [34].
We [13••] expanded the applicability of these experiments by investigating soybean and Lotus corniculatus [35], two legumes that are distantly related. We transferred Le1into
Mesorhizobium loti. Experiments on the transgenic L. cor-niculatusshowed the following: first, SBL was expressed in the emerging root hairs in the susceptible zone of the root; second, when the transgenic Lotusroots were inocu-lated with B. japonicum, nodules formed, probably because of the high concentration of heterologous Nod factor brought about by the accumulation of bradyrhizobia at the root hair tips. However, the nodules were devoid of bacteria because infection threads aborted; third, if the sugar-binding site of Le1 was mutated and transgenic plants carrying the mutated lectin gene were inoculated with B. japonicum,no nodules developed even though the defective lectin was properly targeted to the root hair tip. The bradyrhizobia did not attach to these root hairs [13••], and, presumably, a high concentration of heterologous Nod factor was no longer present at the root hair tip.
These experiments indicated that lectins mediate one level of host specificity, and furthermore, that some component
of the bradyrhizobial cell surface must bind to SBL. Although B. japonicumUSDA110 induced nodules on the roots of the transgenic Lotus, neitherB. elkanii USDA31, which does not bind SBL, nor B. japonicum exoB mutants, attached to theLotus roots or induced nodules over the lev-els demonstrated by the control plants [13••]. These results suggested that mediation of host specificity occurs via lectin interacting with an exopolysaccharide component of the rhizobial cell surface.
If exopolysaccharide is a ligand for lectin, one would pre-dict that Exo–rhizobia would be unable to attach to plant root hairs or to elicit infection thread formation. The for-mer is not true, most likely because other rhizobial proteins also function in adhesion (see references in [36]). However, the latter is frequently true. Inoculation with rhizobial exopolysaccharide mutants phenocopies the nodules developed by the transgenic lectin plants. Uninfected nodules develop in response to Exo– mutants because infection threads abort [37,38••]. For Sinorhizobium melilotistrain 1021, succinoglycan appears to be essential for colonization of the root hair. Mutants that either do not produce or overproduce succinoglycan fail to form proper infection threads [38••]. Exo–mutants of R. leguminosarum bv. trifolii also elicit uninfected nodules [39,40•].
How is lectin mediating specificity in the transgenic plants? One possibility is that lectins are a ‘glue’ that attaches the rhizobia to a site on a susceptible root hair, in this way providing a point source of rhizobial-pro-duced signal molecules. First, secreted lectin aggregates the rhizobia and facilitates their attachment to the root hair (Figure 2). Next, the bacteria elicit infection thread formation even though they produce a heterologous Nod factor. Nod–rhizobia do not induce infection threads or induce Ccd on the transgenic roots, arguing for the involvement of Nod factor in these processes [13••]. That infection threads are formed in the root hairs of the transgenic plants suggests that lectins mediate rhizobial attachment even within the thread (Figure 2). Nevertheless, even though the lectin and the surface polysaccharides are complementary, the threads abort and the nodules are uninfected, suggesting that there is some required response that is not duplicated in the transgenic plants. One possibility is that the transgenic lectin may not be properly integrated into a nodulation signal transduction pathway.
Plant receptors for lectins?
Lectins bind glycoconjugates found on the surfaces of for-eign organisms. Indeed, lectins may serve as a sort of ‘cloaking’ device thereby facilitating the entry of a patho-genic microorganism. Brelles-Marino et al. [41] found that infection thread development was enhanced when R. etli
was incubated with bean seed lectin. Does lectin mask rhi-zobial cell surface components such that the root hair cell takes up these foreign cells?
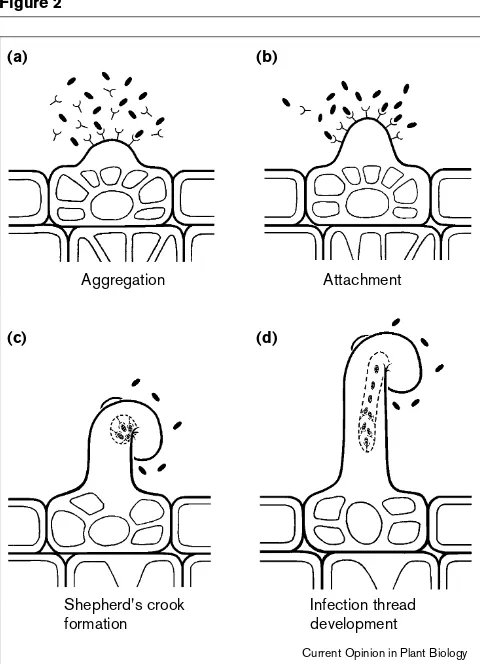
Figure 2
Diagram of how lectin is presumed to function in the transgenic plants carrying a foreign lectin gene. After Kijne [51]. (a)Lectins (Y) are localized to and secreted from the emerging root hair facilitating rhizobial aggregation. (b)The rhizobia are attached to the root hair. (c)The root hair curls around the rhizobia and an infection thread forms at the ‘hyaline spot’. The rhizobia are bound by lectin within the nascent thread. (d)The infection thread extends and the root hair elongates. Rhizobia multiply within the thread.
(a) (b)
(c) (d)
Aggregation
Shepherd's crook formation
Infection thread development
Attachment
A larger question is how the bacteria are engulfed once they make contact with the root hair. The lectins used to formulate the lectin recognition hypothesis have no trans-membrane domains that could connect directly to components of a signal transduction cascade. Legume seed lectins are usually hololectins with two or more carbohy-drate-binding sites, one per subunit; the carbohydrate binding domains are identical or very similar [42]. Proteins that bind to legume lectins (potential lectin receptors) have been detected in seeds (see references in [8]), but minimal information exists on their subcellular location [43]. The significance of these protein associations is also largely unknown.
It is possible that lectins function in signal transduction pathways by binding to transmembrane proteins. Metcalf
et al. [44] found that soybean protoplasts bound several iodinated lectins specifically and saturably. Fluorescence microscopy of the protoplasts showed uniform labeling of the plasma membrane implying that the lectins bound to transmembrane receptors. The lectin–receptor complexes were mobile, as demonstrated by photobleaching studies, and mild trypsinization of the soybean protoplasts removed SBL, but not other lectins [44]. These findings demonstrate that receptors specific for SBL are localized to the plasma membrane.
A gene encoding a new class of receptor-like kinase has been cloned from Arabidopsis [45]. It appears to be a member of a superfamily of genes that define a new class of plant serine/threonine receptor kinases (see Note added in proof). What makes this putative receptor kinase unusual is that, on the basis of sequence informa-tion, the extracellular domain is homologous to a legume lectin, and has binding sites for sugars and for small hydrophobic molecules ([45]; see Note added in proof). However, there are also some significant differences from legume lectins based on molecular modelling, such as the lack of a Ca2+binding site, which may be coupled to a lack of monosaccharide binding activity (see Note added in proof).
Are there proteins in legumes similar to the Arabidopsis
lectin–kinase, or do legume lectins exist as soluble pro-teins that interact with transmembrane kinases once a ligand is bound? Ridge et al. [46••] tested the binding of several lectins with affinities for different sugars on root hairs of uninoculated legumes. They found that five of ten tested lectins bound to the broad-host range legume sira-tro (Macroptilium), whereas only one lectin bound to each of the narrow-host range plants, except for white clover, which bound two. No lectins bound to Arabidopsis root hairs. Sugars at the tips of the root hairs probably exist as glycoconjugates that are transiently expressed. What are these sugar-binding molecules at the root hair tip and are they involved in rhizobial entry? Could they be part of the signal transduction chain?
Figure 3
Two possible models to explain the involvement of lectin in infection thread formation. In the first model, (a–d),lectin acts as a ‘glue’. (a) Lectins (Y) and putative Nod factor receptors (ξ) are randomly distributed on the uninoculated root hair tip. (b) If Nod factor (NF) alone is added, the root hairs deform because Nod factor binds to its receptor, but the receptors do not migrate laterally within the plasma membrane. (c) In the presence of rhizobia, root hairs curl.
(d) Enlargement of boxed area in (c). Lectin molecules bind to rhizobial surface molecules concentrating the source of Nod factor. Nod factor receptors cap after binding to Nod factor due to its localized production. Capping activates the signal transduction pathway. In the second model (e,f), lectins directly interact with another component. (e) Lectins bound to transmembrane proteins are randomly distributed on the uninoculated root hair top. (f) When the lectin binds the rhizobial surface polysaccharides, the attached proteins cap. This event is Nod factor-dependent because infection threads do not form in its absence.
(a) Uninoculated root hair: lectin and Nod Factor receptors.
(c) Rhizobia added to root hair: the hair curls
(e) Uninoculated root hair: lectin associates with a transmembrane receptor
(f) Enlargement of (c). Binding of lectin to rhizobal cell causes lateral migration of receptors. (b) Purified NF added to root hairs: the hair deforms.
(d) Enlargement of (c). Both lectin and NF receptors bind to rhizobial cell.
Conclusions and predictions
As it stands, the lectin recognition hypothesis — with a few modifications — has withstood the test of time. The question remains, however, as to whether lectin is mere-ly a glue that holds the rhizobia and the plant in close contact until Nod factor works its wonders, or whether it plays an active role in specificity by transmitting a signal to one or both symbiotic partners. For example, Kijne and co-workers [47] proposed that lectins may be involved in stabilizing the cell cytoskeleton by trans-membrane interactions. When lectin binds rhizobia, the root hair tip becomes destabilized and an infection thread can form.
Figure 3 illustrates other possibilities. In an uninoculated root hair (Figure 3a), Nod factor receptors and lectins are randomly distributed in and along the membrane of the root hair tip. Adding Nod factor by itself does not cause receptors/binding proteins to migrate even though Nod factor is bound to them; thus, the root hair deforms but does not curl (Figure 3b). When rhizobia are added, the root hair curls around the attached bacteria (Figure 3c). Attachment facilitated by lectins binding to their polysac-charide coats concentrates the rhizobia at a spot on the root hair. Nod factor synthesized by the rhizobia then binds to specific receptor/binding proteins that migrate laterally and form a focused cap (Figure 3d). The size of the cap dictates the strength of the ensuing signal transduction pathway and whether or not the rhizobia will be taken up into the infection thread [48].
In another model, lectins complex with transmembrane proteins, which, in the uninoculated root, are distributed randomly at the root-hair tip (Figure 3e). Upon inocula-tion, the lectin binds to the rhizobial surface and then interacts via a protein-binding domain with the putative transmembrane protein. The complexes cap and, in the process, the transmembrane proteins transmit the signal through an as yet uncharacterized nodulation signal trans-duction pathway (Figure 3f). Nod factor is still required for rhizobial entry because even though nodules develop, bona fide infection threads do not form without rhizobia contacting the root hair cell membrane. However, the effect of Nod factor may be independent of a receptor or binding protein. It is known that Nod factor can modify the biological activity of the root hair cell even in the absence of rhizobia (see references in [9]). Whether or not it does this through an external Nod Factor receptor/bind-ing protein as indicated in Figure 3b or via another mechanism is unknown.
One test of the above possibilities is to determine whether cv. ‘Sooty’ nodulates in response to a low concentration of wild-type bradyrhizobia or to exoB B. japonicum. If there is too little Nod factor brought about by the reduced binding of bradyrhizobia to Sooty root hairs, then the roots will not form nodules.
Many of the components needed for rhizobial endocytosis have been found in legume root hair tips. In addition to lectin, numerous features that are involved in mediating bacterial invasion into host cells — actin, microtubules, clathrin, and various vesicles — have been detected [49,50]. However, we are still missing the critical components of the signal transduction pathway, although attempts to find them are underway. Then, it may be possible to test whether lectins play an active role in propagating a signal that leads to nodule development.
Note added in proof
Two papers have recently been published that are highly relevant to this review [52•,53•]. See main text for details.
Acknowledgements
My thanks to Stefan J Kirchanski, Thomas A LaRue, and Robert W Ridge for reading the manuscript and for their constructive comments. I also want to thank Jan Kijne for insights and the members of my laboratory for their helpful comments. Special thanks go out to those lab members, past and present, who have worked or are working on the lectin project. I am especially grateful to Margaret Kowalczyk for preparing the figures. My apologies to authors whose work I have not cited due to space constraints. Research in the author’s laboratory on lectins is funded by the National Research Initiative Competitive Grant Program, Grants No. 93-37305-9144 and No. 96-35405-353 from the Nitrogen Fixation/Metabolism program.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as:
• of special interest ••of outstanding interest
1. Hamblin J, Kent SP: Possible role of phytohemagglutinin in
Phaseolus vulgaris L. Nat New Biol 1973, 245:28-29.
2. Bohlool BB, Schmidt EL: Lectins: a possible basis for specificity in the Rhizobium-legume root nodule symbiosis. Science1974,
185:269-271.
3. Dazzo FB, Hubbell HD: Cross-reactive antigens and lectins as determinants of symbiotic specificity in the Rhizobium–clover association. Appl Microbiol1975, 30:1017-1033.
4. Etzler ME: Plant lectins: molecular and biological aspects. Annu Rev Plant Physiol1985, 36:209-234.
5. Peumans WJ, Van Damme EJM: Lectins as plant defense proteins.
Plant Physiol1995, 109:347-352.
6. Roberts DD, Goldstein IJ: Binding of hydrophobic ligands to plant lectins: titration with arylaminonapthalene sulfonates. Arch Biochem Biophys1983, 223:479-484.
7. Roberts DD, Goldstein IJ: Adenine binding sites of the lectin from lima beans (Phaseolus lunatus). J Biol Chem1983,
258:13820-13824.
8. Brewin NJ, Kardailsky IV: Legume lectins and nodulation by
Rhizobium. Trends Plant Sci1997, 2:92-98.
9. Schultze M, Kondorosi A: Regulation of symbiotic root nodule development. Annu Rev Genet1998, 32:33-57.
10. Truchet G, Rouche P, Lerouge P, Vasse J, Camut S, de Billy F, Promé J-C, Dénarié J: Sulphated lipooligosaccharide signals of
Rhizobium meliloti elicit root nodule organogenesis in alfalfa.
Nature1991, 351:670-673.
11. Stokkermans TJW, Peters NK: Bradyrhizobium elkanii
lipooligosaccharide signals induce complete nodule structures on
Glycine sojaSiebold et Zucc. Planta1994, 193:413-420. 12. López-Lara IM, van Der Drift KMGM, van Brussel AAN, Haverkamp J,
nodulation signals from broad-host-range Rhizobiumstrain GRH2. Plant Mol Biol 1995, 29:465-477.
13. van Rhijn P, Goldberg RB, Hirsch AM: Lotus nodulation specificity is
•• changed by the presence of a soybean lectin gene.Plant Cell 1998, 10:1233-1249.
Confirmed the validity of the lectin recognition hypothesis, and expanded upon the findings of [33] by showing that the lectin recognition hypothesis is applicable to legumes and rhizobia that are not closely related to each other. It also demonstrated that exopolysaccharide is a likely ligand for soy-bean lectin.
14. Etzler ME, Kalsi G, Ewing NN, Roberts NJ, Days RB, Murphy JB: • A nod factor binding lectin with apyrase activity from legume
roots. Proc Natl Acad Sci USA 1999, 96:5856-5861.
This combination of molecular and biochemical approaches demonstrates that a novel lectin, named LNP, is a Nod factor-binding protein as well as an apyrase. LNP has no similarity in sequence to any lectins that have been described so far. Its putative location on a membrane suggests that it may function in oligosaccharide-signalling events.
15. Dazzo FB, Yanke WE, Brill WJ: Trifoliin A: a Rhizobiumrecognition protein from white clover.Biochim Biophys Actal978, 539:276-286. 16. Law IJ, Strijdom BW: Role of lectins in the specific recognition of
Rhizobiumby Lotononis bainesii.Plant Physiol1984,74:779-785. 17. Díaz, CL, van Spronsen PC, Bakhuizen R, Logman GJJ, Lugtenberg
BJJ, Kijne JW: Correlation between infection by Rhizobium leguminosarumand lectin on the surface ofPisum sativum
L. roots.Planta1986, 168:350-359.
18. Díaz, CL, Logman TJJ, Stam HC, Kijne JW: Sugar-binding activity of pea lectin expressed in white clover hairy roots.Plant Physiol 1995, 109:1167-1177.
19. Kato G, Maruyama Y, Nakamura M: Involvement of lectins in
Rhizobium-pea recognition.Plant Cell Physiol 1981,22:519-771. 20. Gatehouse JA, Boulter D: Isolation and properties of a lectin from
the roots of Pisum sativum(garden pea).Physiol Plant1980,
49:437-442.
21. Okamuro JK, Jofuku KD, Goldberg RB: Soybean seed lectin gene and flanking nonseed protein genes are developmentally regulated in transformed tobacco plants.Proc Natl Acad Sci USA 1986, 83:8240-8244.
22. Kaminski PA, Buffard D, Strosberg AD: The pea lectin gene family contains only one functional lectin gene.Plant Mol Biol1987,
9:497-507.
23. Vodkin LO, Raikhel NV: Soybean lectin and related proteins in seeds and roots of Le+ and Le-soybean varieties. Plant Physiol 1986, 81:558-565.
24. Wycoff K, van Rhijn P, Hirsch AM: The ribosomal protein P0 of soybean (Glycine max L. Merr.) has antigenic cross-reactivity to soybean seed lectin.Plant Mol Biol1997,34:295-306. 25. Pull SP, Pueppke SG, Hymowitz T, Orf JH: Soybean lines lacking
the 120,000-Dalton seed lectin.Science1978, 200:1177-1179. 26. Halverson LJ, Stacey G: Host recognition in the Rhizobium
-soybean symbiosis: evidence for the involvement of lectin in nodulation.Plant Physiol1985, 77:621-625.
27. Dombrink-Kurtzman MA, Dick WE Jr, Burton KA, Cadmus MC, Slodki ME: A soybean lectin having 4-O-methyl-D-glucuronic acid specificity. Biochem Biophys Res Commun1983, 111:798-803. 28. Rutherford WM, Dick WE Jr, Cavins JF, Dombrink-Kurtzman MA,
Slodki ME: Isolation and characterization of a soybean lectin having 4-O-methyglucuronic acid specificity.Biochemistry1986,
25:952-958.
29. Bhuvaneswari TV, Bauer WD: Role of lectins in plant-microorganism interactions. III. Influence of rhizosphere-rhizoplane culture conditions on the soybean lectin-binding properties of rhizobia.Plant Physiol1978, 62:71-74.
30. Dazzo FB, Truchet G: Interactions of lectins and their saccharide receptors in the Rhizobium-legume symbiosis.J Memb Biol1983,
73:1-16.
31. Kamberger W: Binding specificity and purification of Medicago sativa lectin.InAffinity Chromatography, Proc Int Symp Vienna. Edited by Hoffman-Ostenhof O, Breitenbach M, Koller F, Draft D, Scheiner O. Oxford: Pergamon Press; 1978:295-298.
32. Jayaraman V, Das HR: Interaction of peanut root lectin (PRA II) with rhizobial lipopolysaccharides.Biochim Biophys Acta1998, 1381:7-11. 33. Díaz CL, Melchers LS, Hooykaas PJJ, Lugtenberg EJJ, Kijne JW: Root
lectin as a determinant of host–plant specificity in the
Rhizobium-legume symbiosis.Nature1989,338:579-581.
34. van Eijsden R, Díaz C, de Pater BS, Kijne JW: Sugar-binding activity of pea (Pisum sativum) lectin is essential for heterologous infection of transgenic white clover hair roots by Rhizobium leguminosarumbiovar viciae.Plant Mol Biol1995, 29:431-439. 35. Doyle JJ: Phylogenetic perspectives on nodulation: evolving views
of plants and symbiotic bacteria.Trends Plant Sci1998,3:473-478. 36. Niner BM, Hirsch AM: How many Rhizobiumgenes, in addition to
nod, nif/fix, and exo, are needed for nodule development and function?Symbiosis1998, 24:51-102.
37. Finan TM, Hirsch AM, Leigh JA, Johansen E, Kuldau GA, Deegan S, Walker GC, Signer ER: Symbiotic mutants ofRhizobium meliloti
that uncouple plant from bacterial differentiation.Cell1985,
40:869-877.
38. Cheng HP, Walker GC: Succinoglycan is required for initiation and
•• elongation of infection threads during nodulation of alfalfa by
Rhizobium meliloti. J Bacteriol1998, 180:5182-5191.
Green flourescent protein (GFP) was used as reporter for the purpose of tracking the degree of infection thread arrest within alfalfa root hairs caused by different exomutants of Sinorhizobium meliloti. This molecular dissection of the succinoglycan substituents required for proper infection thread devel-opment may give us a better idea of the identity of the receptor within root hairs that allows infection thread development and rhizobial invasion.
39. Rolfe BG, Carlson RW, Ridge RW, Dazzo FB, Mateos PF, Pankhurst CE: Defection infection and nodulation of clovers by exopolysaccharide mutants of Rhizobium leguminosarumbv.
trifolii.Aust J Plant Physiol1996, 23:285-303.
40. Król J, Wielbo J, Mazur A, Kopcinska J, Lotocka, B, Golinowski W, • Skorupska A: Molecular characterization of pssCDEgenes of
Rhizobium leguminosarumbv. trifoliistrain TA1: synthesis and endocytosis of bacteria.Mol Plant–Microbe Inter1998, 11:1142-1148. In contrast to S. meliloti exopolysaccharide mutants [37,38••], infection threads elicited by clover rhizobia mutants penetrate nodule cells, but simi-lar to the S. melilotimutants, endocytosis is defective.
41. Brelles-Marino G, Costa GA, Boiardi JL: Enhancement of infection thread formation by Rhizobium etli incubated with bean seed lectin.Microbiol Res1996, 151:243-246.
42. Sharon N, Lis H: Legume lectins — a large family of homologous proteins.FASEB J 1990, 4:3198-3208.
43. Pusztai A: Plant lectins.Cambridge: Cambridge University Press; 1991.
44. Metcalf TN, Wang L, Schubert KR, Schindler M: Lectin receptors on the plasma membrane of soybean cells. Binding and lateral diffusion.Biochemistry1983, 22:3969-3975.
45. Hervé C, Dabos D, Galaud J-P, Rougé P, Lescure B: Characterization of anArabidopsis thaliana gene that defines a new class of putative plant receptor-kinases with an extracellular lectin-like domain.J Mol Biol1996, 258:778-788.
46. Ridge RW, Kim RA, Yoshida F: The diversity of lectin-detectable
•• sugar residues on root hair tips of selected legumes correlates with the diversity of their host ranges for rhizobia. Protoplasma 1998, 202:84-90.
A promising approach showing that lectins may be binding to glycoconju-gates, which could function in the nodulation signal transduction pathway. These glycoconjugates are likely to be an integral part of the plasma mem-brane at the root hair tip and may represent specific receptors for rhizobia, Nod factor, or other determinants.
47. Kijne JW, Bauchrowitz MA, Díaz CL: Root lectins and rhizobia.Plant Physiol 1997, 115:869-873.
48. Hirsch AM: Tansley Review No. 40. Developmental biology of legume nodulation.New Phytol 1992, 122:211-237.
49. Ridge RW: A model of legume root hair growth and Rhizobium
infection. Symbiosis 1993, 14:359-373.
52. Gressent F, Drouillard S, Mantegazza N, Samain E, Geremia RA, • Canut K, Niebel A, Driguez H, Ranjeva R, Cullimore J, Bono JJ: Ligand
specificty of a high-affinity binding site for lipo-chitooligosaccharide Nod factors in Medicago cell suspension cultures. Proc Natl Acad Sci USA 1999, 96:4704-4709.
Like [14•], this paper uses a biochemical approach to examine a Nod factor-binding site (NFBS2) isolated from Medicago cell cultures and the structural requirements of Nod factor for binding. Although recognition is mediated by both the lipid and oligosaccharide components of the Nod factor molecule, surprisingly, NFBS2 is not selective for the reducing end-sulfate group, which normally is essential for Medicago root hair deformation and nodulation.
53. Hervé C, Serres J, Dabos P, Canut H, Barre A, Rougé P, Lescure B: • Characterization of the Arabidopsis lecRK-a genes: members of
a superfamily encoding putative receptors with an extracellular domain homologous to legume lectins. Plant Mol Biol 1999,
39:671-682.