Summary Pollution often causes visible symptoms of foliar injury. The injury is sometimes associated with an increase in the accessibility of toxic elements to plants as a result of acidification of the soil. We investigated the distribution of elements (N, P, K, Ca, Mg, Mn, S, Fe, B, Cu, Zn, Al, F, Pb, Cd, Cr, Ni and Co) in healthy current-year needles of Scots pine (Pinus sylvestris L.) growing at an unpolluted control site and at a site polluted mainly by SO2, HF and Al3+ from a fertilizer factory established in 1917. Needles from both sites were sampled before the appearance of visible injury and cut into five sections of equal length (tip, base and three middle parts). The mean concentrations of major nutrients were 20--30% lower in needles at the polluted site than in needles at the control site, whereas the concentrations of aluminum and fluo-rine were higher in needles at the polluted site. An increase in concentration from needle base to tip was detected for N, Fe, B, and Al at both sites and for Mn only at the polluted site. Fluoride accumulated in the tips of needles only at the polluted site, which could explain the necroses of needle tips at this site. The distribution of elements along the length of the needles was influenced by pollution, element mobility and the distal accumulation of toxic elements.
Keywords: chemical elements, needle part, nutrients, Pinus sylvestris, pollution.
Introduction
There have been many analyses of the chemical composition of Scots pine needles. As a result, we know the ranges of optimal concentration for particular elements, the thresholds of toxicity for some elements (Fober 1993) and the symptoms of nutrient deficiency as well as the influence of fertilization (Baule and Fricker 1973). The accumulation of elements in plant leaves is a dynamic process that is coupled with other metabolic processes occurring in the plant. Element concentra-tions are also influenced by many factors including foliar age, time of year, and environmental conditions (e.g., edaphic qual-ity and pollution level) at the site (van den Driessche 1974, Ostrowska and Szczubialka 1988, Heliövaara and Väisänen 1989).
In Pinus species, the effects of growing season, origin of seed source, age of needles and position on tree crown on changes in element concentrations are well documented (Will 1957, Giulimondi and Duranti 1975, Fober 1976, Helmisaari 1990, Heisendorf et al. 1993). Because many papers have shown that pollution alters nutrition and the concentration of elements in needles (Skeffington and Roberts 1985, Reich et al. 1988, Heliövaara and Väisänen 1989, Holopainen and Ny-gren 1989, Raitio 1990, Baker et al. 1994, Shaw and McLeod 1995), the chemical composition of needles is frequently used to monitor environmental pollution (Dmuchowski and Byt-nerowicz 1995). In all of the studies of pollution cited above, the mean concentrations of elements in whole needles were usually determined; however, the appearance of the visible injuries suggests that the high concentration of elements asso-ciated with injury or symptoms of deficiency occur only in particular parts of needles. So far, only the work of Jamrich (1972) describes the concentrations of several elements in different parts of needles injured by fluorine emissions.
Therefore, the aim of this study was to determine whether there is an accumulation of toxic elements in the part of healthy needles where necroses often appear. We also assessed the influence of pollution on the distribution of chemical elements along the length of the needle. We analyzed needles without visible injuries, to determine how the elements were distrib-uted before the appearance of necroses. The study was con-ducted with healthy current-year needles of Scots pine (Pinus sylvestris L.) growing at an unpolluted control site and at a polluted site at which typical needle injuries caused by fluoride and sulfur dioxide have been documented (Piskornik and Godzik 1970, Horntvedt and Robak 1975, Malhotra and Blauel 1980, Oleksyn et al. 1988, Karolewski 1990, Giertych and Werner 1996).
Materials and methods
Plant material
Current-year needles were collected after the end of the grow-ing season in September 1994 from 12-year-old Scots pine trees growing at two sites in western Poland. One site is located at Luboñ near Poznañ (52°15′20″ N, 16°50′31″ E), about 2 km
Distribution of elements along the length of Scots pine needles in a
heavily polluted and a control environment
M. J. GIERTYCH,
1L. O. DE TEMMERMAN
2and L. RACHWAL
11
Polish Academy of Sciences, Institute of Dendrology, Parkowa 5, PL-62-035 Kórnik, Poland
2 Institute for Chemical Research, Leuvensesteenweg 17, 3080 Tervuren, Belgium
Received July 11, 1996
from a phosphate fertilizer factory that was established in 1917, and the other site is in the Forest Range Zwierzyniec near Kórnik (52°14′36″ N, 17°05′00″ E), about 15 km from the fertilizer factory and considered to be free of acute pollution (control). Both sites have been intensively studied during the past 12 years (Oleksyn and Bialobok 1986, Oleksyn 1988, Reich et al. 1994, Karolewski and Giertych 1995). The pol-luted site has received long-term pollution in the form of atmospheric deposition of SO2 and HF and it is also charac-terized by an elevated concentration of Al3+ ions in the soil (Reich et al. 1994, Karolewski and Giertych 1995).
Preparation procedure
Healthy-looking, current-year needles were taken from sev-eral branches in different parts of the crown. To minimize possible confounding effect of genetic differences, at each site, samples were taken from three trees (replicates) from one Polish provenance (No. 7 Spala). The needles were transported to the laboratory in plastic bags. The needles were not washed. Each needle was cut with a razor-blade into five equal sections
proportional to the length, defined as a base, tip and three middle parts. Before cutting the needles, the fascicle sheath was removed. The cutting occurred over several days and during this time the needles were stored at 4 °C. The sections from each tree were pooled separately. Needle sections were weighed, oven dried at 65 °C, weighed again to determine the content of water, ground and stored in plastic boxes. Each sample (one needle section per tree) weighed more than 6 g dry weight (gdw). The five sections were analyzed separately for all elements except sulfur, for which three sections (tip, base and one middle) were analyzed.
Chemical analyses
For the analysis of P, K, Ca, Mg, Mn, Fe, B, Cu, Zn, Al, Pb, Cd, Cr, Ni and Co, the modified CII method was followed (CII 1969). Detection limits for heavy metals were 0.35 µg gdw−1 for Cd, Cu, Cr and Co, 0.25 µg gdw−1 for Zn, and 1.25 µg gdw−1 for Pb and Ni. Recoveries of 90--100% were obtained for several reference plant materials: BCR 060, 061, 062 and NIST 1570, 1571, 1572, 1573, 1575. Dried and ground needles were
calci-Table 1. Summary of ANOVA for needle element concentrations, by site and needle section (df = degrees of freedom; MS = mean squares; and
P-values = probability values).
Source of variation Element
N P K Mg
df MS P MS P MS P MS P
Site 1 89399000 0.0097 397060 0.1546 13770000 0.0739 855260 0.0302
Tree (Site) 4 4137000 129300 2380000 78948
Sections 4 14852000 0.0000 24010 0.0002 5680800 0.0000 9785 0.0482
Site × Section 4 1035500 0.2676 5574 0.0863 951300 0.0394 9781 0.0483
Error 16 721100 2254 292800 3216
Ca B S (inorganic) S (organic)
df1 MS P MS P MS P MS P
Site 1 (1) 9785800 0.1465 225.9 0.0052 207100 0.2543 131500 0.7474
Tree (Site) 4 (4) 3025000 7.3 117100 1105000
Sections 4 (2) 1266300 0.0000 108.4 0.0002 97128 0.3710 512200 0.7135
Site × Section 4 (2) 105500 0.1155 9.1 0.4773 35067 0.6762 902600 0.5629
Error 16 (7) 48014 9.9 85472 1445000
Zn Cu Ni Fe
df MS P MS P MS P MS P
Site 1 324.5 0.0244 0.94 0.3738 0.31 0.5749 0.186 0.9335
Tree (Site) 4 26.2 0.94 0.82 23.565
Sections 4 59.3 0.0044 0.77 0.0000 10.46 0.0000 27.265 0.0079
Site × Section 4 5.5 0.7102 0.60 0.0002 0.29 0.6852 5.642 0.4137
Error 16 10.2 0.06 0.51 5.384
Mn Al F Ca/Al (molar)
df MS P MS P MS P MS P
Site 1 1651.1 0.1881 248510 0.0103 1365.5 0.0240 1181.5 0.0358
Tree (Site) 4 657.2 11919 109.0 122.0
Sections 4 2495.1 0.0119 425770 0.0000 1252.8 0.0004 243.5 0.0000
Site × Section 4 2633.0 0.0096 124870 0.0002 1242.2 0.0004 60.7 0.0091
Error 16 546.7 1177 131.5 12.4
nated for 2 h at 450 °C. Silicates were destroyed by adding a mixture of HF and HNO3 to the ash and evaporating (three successive times). The residue was dissolved in HNO3 and the concentration was measured by inductively coupled plasma emission spectrometry (ICP).
Nitrogen was analyzed by the Kjeldahl procedure. Sulfur was determined by ion chromatography (Dionex 2000i, Sun-nyvale, CA) after differentiation of organic and inorganic sul-fur by a slight modification of the procedure described by Jäger and Steubing (1970). Total sulfur was calculated by adding organic and inorganic sulfur.
For the determination of fluoride, dried and ground plant material was extracted with 0.1 N HNO3 followed by the addition of sodium citrate buffer solution. The fluoride con-centration was determined with an ion selective electrode (Roost and Sigg 1978). The detection limit for fluorides is 1 µg gdw−1. To eliminate interactions of other elements present in the samples, the results were calibrated against samples contain-ing standard additions of known fluoride concentration (Jagner
and Pavlova 1972). All chemical analyses were conducted at the laboratory of the Institute for Chemical Research in Tervuren, Belgium.
Statistical analysis
Analysis of variance (ANOVA) was used to assess the influ-ences of site, section and their interaction on chemical element distribution (Statistix 4.0 Analytical Software, St. Paul, MN). Sections were nested within a tree. Site and section effects were considered to be fixed and the tree effect was considered to be random.
Results
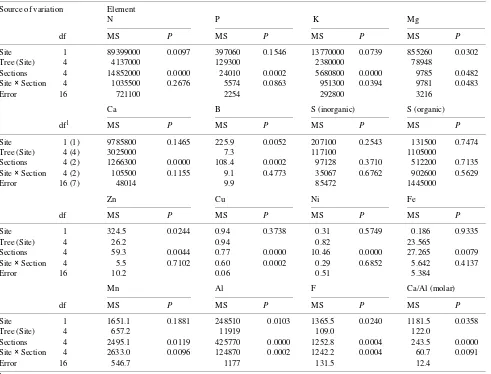
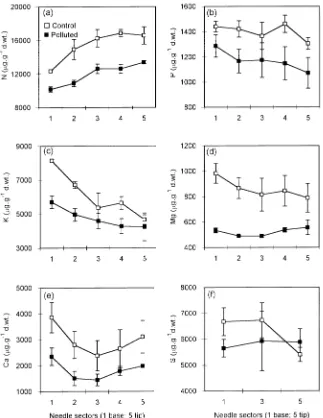
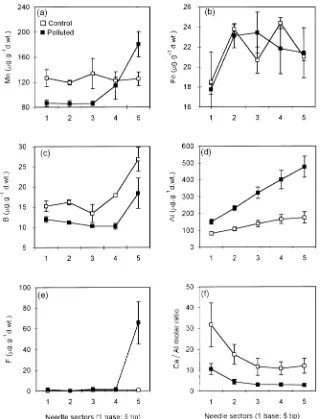
The distribution of most elements along the length of the needle was similar at both sites. Statistically significant in-creases from base to tip sections were noted for N, Fe, B and Al (Table 1, Figures 1a and 2b--d), whereas decreases from
base to tip were noted for P, K, Zn, and Ni and for the Ca/Al molar ratio (Table 1, Figures 1b, 1c, 2f, 3a and 3c). There was a significant interaction between site and needle section for K, Mg, Mn and Cu (Table 1). The concentration of Mn was similar along the length of needles at the control site, but in needles at the polluted site there was an increase in Mn concentration from base to tip (Figure 2a). Magnesium concentration de-creased from base to tip in control needles, whereas it was similar along the length of needles at the polluted site (Fig-ure 1d). A significant interaction was also found for F which accumulated in needle tips only at the polluted site (Table 1, Figure 2e). There were no significant effects of site or needle section on organic, inorganic and total S (Table 1) concentra-tions. The amounts of Pb, Cd, Cr and Co were below the detection limits.
Mean needle concentrations of N, Mg, B and Zn were significantly higher (20--30%) at the control site than at the polluted site (Tables 1 and 2), whereas mean needle concentra-tions of Fe, Mn, inorganic S, organic S and total S were similar
at both sites. Mean needle concentrations of Al and F were 2.3-and 23-fold higher, respectively, at the polluted site than at the control site. The Ca/Al ratio was significantly lower at the polluted site than at the control site (Tables 1 and 2).
Needle water contents were significantly higher at the con-trol site than at the polluted site (53.5 ± 0.6% versus 46.8 ± 0.7%).
Discussion
Significant differences in element concentrations between needle sections were found for several elements. An explana-tion of these differences is difficult because of a lack of infor-mation on potential differences in physiological processes and nutrient requirements in particular parts of the needle. We know that most needle cells are of similar age because most cells are already present in the buds and spring leaf growth occurs by elongation. However, there is little information on the variation in longitudinal morphology of the needle, and so
it is impossible to relate observed differences in the concentra-tion of elements along the needle length with reference to its morphology and development.
For the elements classified as mobile, N, K, B, Zn, Ni and Mn (Helmisaari 1990, Kabata-Pendias and Pendias 1993), differences along the needle length could be the result of accumulation or retranslocation which occurs late in the grow-ing season (Helmisaari 1992). The elements classified as im-mobile Ca, Fe and organic S did not exhibit significant differences in concentration between base and tip parts of the needle except for a lower concentration of Fe in the base section. This could be explained by a lower concentration of chlorophyll in this part of the needle, which is normally cov-ered by the needle sheath.
Fluorides accumulated in the tip of the healthy needles, which is where necroses often occur (Jamrich 1972, Horntvedt and Robak 1975, Malhotra and Blauel 1980). We postulate that the accumulation of highly toxic fluoride in the needle tips and the subsequent development of necroses in this area reduces the influence of this element on the healthy part of needles. Jamrich (1972) found the highest concentration of fluorides in the discolored bands separating the green part of needles from necroses, suggesting that F accumulates in the part of the needle that aborts first.
The concentration of Al exhibited a linear increase from base to tip. This pattern could be associated with the transloca-tion of Al in the transpiratransloca-tion stream. Cronan and Grigal (1995) identified four measurements that indicate threshold condi-tions for Al-induced stress in forest trees. One of these meas-urements is the foliar tissue Ca/Al molar ratio of ≤ 12.5 indicating 50% risk of Al-induced damage (cf. Rengel 1992). The Ca/Al ratio is also positively correlated with the rate of photosynthesis (Reich et al. 1994). In our study, the Ca/Al molar ratio decreased from the base to the tip of needles at both sites. For control needles, the values were 31.7 (base); 17.4; 11.6; 10.8; 12.0 (tip) and the corresponding values for polluted needles were 10.5 (base); 4.4; 3.0; 3.0; 2.8 (tip). Thus, the Ca/Al molar ratio was below the threshold value throughout the length of needles at the polluted site and in the distal half of needles at the control site.
The different distributions of Mn at the two sites----a similar concentration in all parts of control needles and an increase in concentration from base to tip in polluted needles----could be associated with the distribution of Al in the needles. According to Kabata-Pendias and Pendias (1993), an overabundance of Al can increase absorption of other cations, particularly Mn. However, Oleksyn et al. (1995) found that the presence of Figure 3. Mean (± SE) concentrations of heavy metals (Zn, Cu and Ni)
along the length (five sections) of current-year needles of Scots pine trees (n = 3) growing on polluted and control sites.
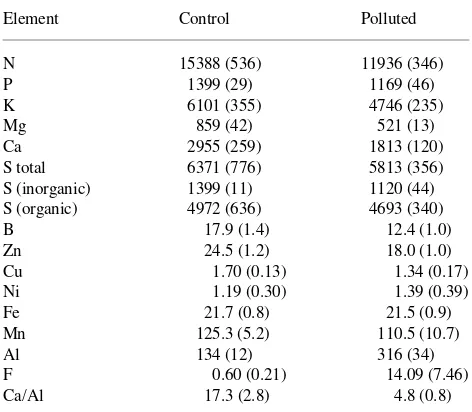
Table 2. Mean (SE) concentrations (µg gdw−1) of elements and molar ratio of Ca/Al in whole needles of Scots pine from a control and polluted site.
Element Control Polluted
N 15388 (536) 11936 (346)
P 1399 (29) 1169 (46)
K 6101 (355) 4746 (235)
Mg 859 (42) 521 (13)
Ca 2955 (259) 1813 (120)
S total 6371 (776) 5813 (356)
S (inorganic) 1399 (11) 1120 (44) S (organic) 4972 (636) 4693 (340)
B 17.9 (1.4) 12.4 (1.0)
Zn 24.5 (1.2) 18.0 (1.0)
Cu 1.70 (0.13) 1.34 (0.17)
Ni 1.19 (0.30) 1.39 (0.39)
Fe 21.7 (0.8) 21.5 (0.9)
Mn 125.3 (5.2) 110.5 (10.7)
Al 134 (12) 316 (34)
F 0.60 (0.21) 14.09 (7.46)
aluminum in solution reduced root and needle concentrations of Mn of hydroponically grown Scots pine seedlings.
At both sites, mean concentrations of the macroelements N, P, K, and Ca were in the usual range for this species in Poland (Fober 1976, 1993, Czarnowska and Stasik 1987, Kucaba 1987). In contrast, the concentration of Mg at the polluted site was very low and may be insufficient for normal growth (Ingestad 1979). According to Fober (1993), changes caused by deficiency of N, Ca, Fe or B are visible in whole needles, whereas changes caused by deficiency of K, P, Mg or Mn are only visible at the needle tip. We observed that concentrations of Ca and Fe were evenly distributed along the length of the needles, whereas concentrations of K and P were lower in needle tips than in other parts of the needle. The concentrations of some microelements (Fe, Zn, B, S, Mn) were very low at both sites and well below toxic concentrations (Kabata-Pen-dias and Pen(Kabata-Pen-dias 1993). Only F at the polluted site occurred at concentrations considered toxic.
Needle concentrations of N, K, Mg, B, and Zn were signifi-cantly higher at the control site that at the polluted site. This difference may be related to the lower water content of pol-luted needles and associated differences in senescence proc-esses between sites. During senescence the concentration of water in needles decreases (Zelawski 1967) as do the needle concentrations of N and K (Helmisaari 1992). An alternative explanation for the significantly higher concentrations of ma-jor elements at the control site than at the polluted site may be associated with the more acidic soil at the polluted site com-pared to the control site, pH 4.2 versus 5.8 (Reich et al. 1994). On acidic soils, accessibility of Al for plants is increased and aluminum limits the uptake of macroelements and some mi-croelements (Kabata-Pendias and Pendias 1993).
We conclude that many elements are unevenly distributed along the length of Scots pine needles. The distribution pat-terns can be influenced by pollution, by differences in the mobility of elements and by the distal accumulation of toxic elements. The distribution of some elements was associated with subsequent injury or deficiency symptoms. Because chemical analyses of whole needles may underestimate ele-ment concentrations in particular needle parts, it is important to identify which part of the needle is used for analysis, particularly when conducting bioindication analyses. Differ-ential distribution of chemical elements along the length of the needle can also influence physiological and biochemical proc-esses and it is necessary to examine this subject.
Acknowledgments
The authors thank A. Niemir and R. Zytkowiak for help in fieldwork and sample preparation, and M. Hoeing, P. Coosemans, R. Van Cauter and H. Baeten for help in conducting laboratory analyses. We are also grateful to Mark G. Tjoelker, Jacek Oleksyn and Piotr Karolewski for helpful comments on earlier versions of this manuscript.
References
Baker, T.R., H.L. Allen, M.M. Schoeneberger and L.W. Kress. 1994. Nutritional response of loblolly pine exposed to ozone and simu-lated acid rain. Can. J. For. Res. 24:453--461.
Baule, H. and C. Fricker. 1973. Die Düngung von Waldbäumen (Pol-ish edition). PWRiL, Warsaw, pp 55--90.
CII. 1969. Méthodes de référence pour la détermination des éléments mineraux dans les végétaux: N, P, K, Na, Ca, Mg. Agron. Trop. 9:827--835.
Cronan, C.S. and D.F. Grigal. 1995. Use of calcium/aluminum ratios as indicators of stress in forest ecosystems. J. Environ. Qual. 24:209--226.
Czarnowska, K. and J. Stasik. 1987. The concentration of mineral components in needles, wood, and bark of Scots pine according to the age of trees. In Biological Reactions of Trees to Industrial Pollution. Materials from the 2nd National Symposium. Ed. R. Si-wecki. Adam Mickiewicz University Press, Poznañ, pp 183--188. In Polish with English summary.
Dmuchowski, W. and A. Bytnerowicz. 1995. Monitoring environ-mental pollution in Poland by chemical analysis of Scots pine (Pinus sylvestris L.) needles. Environ. Pollut. 87:87--104. Fober, H. 1976. Distribution of mineral elements within the crown of
Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies
(L.) Karst.). Arbor. Kórnickie 21:323--331.
Fober, H. 1993. Mineral nutrition. In Biology of Scots Pine. Eds. S. Bialobok, A. Boratyñski and W. Bugala. Sorus, Poznañ, pp 182--193. In Polish with English summary.
Giertych, M.J. and A. Werner. 1996. Phenolic compounds in needles of Scots pine (Pinus sylvestris L.) damaged by industrial pollution. Arbor. Kórnickie 41:165--172.
Giulimondi, G. and G. Duranti. 1975. Dry matter production and nutrient contents of a 4-year-old Pinus radiata plantation. Pubbli-cazioni de Centro di Sperimentazione Agric. For. 13:17--28. In Italian with English summary.
Heisendorf, M., M. Heinze and H.J. Fiedler. 1993. Changes of nutrient element dynamics in the needles of a pine plantation. Forstwiss. Centralbl. 112:179--190. In German with English summary. Heliövaara, K. and R. Väisänen. 1989. Quantitative variation in the
elemental composition of Scots pine needles along a pollutant gradient. Silva Fenn. 23:1--11.
Helmisaari, H.-S. 1990. Temporal variation in nutrient contents of
Pinus sylvestris needles. Scand. J. For. Res. 5:177--193.
Helmisaari, H.-S. 1992. Nutrient retranslocation within the foliage of
Pinus sylvestris. Tree Physiol. 10:45--58.
Holopainen, T. and P. Nygren. 1989. Effects of potassium deficiency and simulated acid rain, alone and in combination, on the ultrastruc-ture of Scots pine needles. Can. J. For. Res. 19:1402--1411. Horntvedt, R. and H. Robak. 1975. Relative susceptibility of eleven
conifer species to fluoride air pollution. Medd. Nor. Inst. Skogforsk. 32:185--206.
Ingestad, T. 1979. Mineral nutrient requirements of Pinus sylvestris
and Picea abies seedlings. Physiol. Plant. 45:373--380.
Jäger, H.J. and L. Steubing. 1970. Fraktionierte Schwefelbestimmung in Pflanzenmaterial zur Beurteilung einer SO2 Einwirkung. Angew. Bot. 44:209--221.
Jagner, D. and V. Pavlova. 1972. A standard addition titration method for the determination of fluorine in silicate rocks. Anal. Chim. Acta 60:153--158.
Kabata-Pendias, A. and H. Pendias. 1993. Trace elements in soil and plants. PWN, Warsaw, pp 89--319. In Polish.
Karolewski, P. 1990. Visible and invisible injury to Scots pine (Pinus sylvestris L.) needles caused by sulphur dioxide. Arbor. Kórnickie 35:127--136.
Karolewski, P. and M.J. Giertych. 1995. Changes in the level of phenols during needle development in Scots pine populations in a control and polluted environment. Eur. J. For. Pathol. 25:297--306. Kucaba, S. 1987. Content of mineral components in yearly organs of Scotch pine (Pinus sylvestris L.) plantation cultures in relation to fertilization and liming. Folia For. Pol. Ser. A. 29:45--55. In Polish with English summary.
Malhotra, S.S. and R.A. Blauel. 1980. Diagnosis of air pollutant and natural stress symptoms on forest vegetation in western Canada. Environ. Can., Can. For. Serv., North. For. Res. Ctr. Edmonton, Alberta. Inf. Rep. NOR-X-228, pp 36--43.
Oleksyn, J. 1988. Report on the IUFRO-1982 provenance experiment on Scots pine (Pinus sylvestris L.). Arbor. Kórnickie 33:211--229. Oleksyn, J. and S. Bialobok. 1986. Net photosynthesis, dark
respira-tion and susceptibility to air pollurespira-tion of 20 European provenances of Scots pine Pinus sylvestris L. Environ. Pollut. 40:287--302. Oleksyn, J., P. Karolewski and L. Rachwal. 1988. Susceptibility of
European Pinus sylvestris L. populations to the action of SO2, NO2, SO2 + NO2 and HF in laboratory and field conditions. Acta Soc. Bot. Pol. 57:107--115.
Oleksyn, J., P. Karolewski, M.J. Giertych, A. Werner, M.G. Tjoelker and P.B. Reich. 1996. Altered root growth and plant chemistry of
Pinus sylvestris seedlings subjected to aluminum in nutrient solu-tion. Trees 10:135--144.
Ostrowska, A. and Z. Szczubialka. 1988. Comparison of the ion system in the needles of pine of various ages and various growth conditions. Ann. Wars. Agric. Univ. SGGW-AR For. Wood Technol. 37:21--30.
Piskornik, Z. and S. Godzik. 1970. Influence of air pollution on plants, Part II. Wiad. Bot. 14:91--102. In Polish.
Raitio, H. 1990. The foliar chemical composition of young Pinus
(Pinus sylvestris L.) with or without decline. In Acidification in Finland. Eds.P. Kauppi, P. Anttila and K. Kenttämies. Springer-Ver-lag, Berlin, Heidelberg, New York, pp 699--714.
Reich, P.B., J. Oleksyn and M.G. Tjoelker. 1994. Relationship of aluminum and calcium to net CO2 exchange among diverse Scots pine provenances under pollution stress in Poland. Oecologia 97:82--92.
Reich, P.B., A.W. Schoettle, H.F. Stroo and R.G. Amundson. 1988. Effects of ozone and acid rain on white pine (Pinus strobus) seed-ling grown in five soils. III. Nutrient relations. Can. J. Bot. 66:1517--1531.
Rengel, Z. 1992. Role of calcium in aluminium toxicity. New Phytol. 121:499--513.
Roost, P. and A. Sigg. 1978. Erfahrungen mit einer potentiometrischen Fluor Bestimmungsmethode für biologische Materialen. Staub-Re-inhalt Luft 38:363--366.
Shaw, P.J.A. and A.R. McLeod. 1995. The effects of SO2 and O3 on the foliar nutrition of Scots pine, Norway spruce and Sitka spruce in the Liphook open-air fumigation experiment. Plant Cell Environ. 18:237--245.
Skeffington, R.A. and T.M. Roberts. 1985. The effects of ozone and acid mist on Scots pine saplings. Oecologia 65:201--206. van den Driessche, R. 1974. Prediction of mineral nutrient status of
trees by foliar analysis. Bot. Rev. 40:347--394.
Will, G.M. 1957. Variation in the mineral content of radiata pine needles with age and position in tree crown. N.Z. For. Serv. Res. Note 11:699--707.
Zelawski, W. 1967. Gaseous exchange and water balance in needles.