L
Journal of Experimental Marine Biology and Ecology 245 (2000) 197–214
www.elsevier.nl / locate / jembe
Energy balance and cold adaptation in the octopus
Pareledone charcoti
a,b ,* a
H.I. Daly , L.S. Peck
a
British Antarctic Survey, High Cross, Madingley Road, Cambridge CB3 0ET, UK
b
Department of Zoology, University of Aberdeen, Tillydrone Avenue, Aberdeen AB24 2TZ, Scotland, UK Received 18 May 1999; received in revised form 23 July 1999; accepted 25 October 1999
Abstract
A complete energy balance equation is calculated for the Antarctic octopus Pareledone charcoti at 08C. Energy used in respiration, growth, and excretion of nitrogenous and faecal waste, was recorded along with the total consumption of energy through food, for three specimens of P.
charcoti (live weights: 73, 51 and 29 g). Growth rates were very slow for cephalopods, with a
mean daily increase in body weight of only 0.11%. Assimilation efficiencies were high, between 95.4 and 97.0%, which is consistent with previous work on octopods. The respiration rate in P.
21
charcoti was low, with a mean of 2.45 mg O h2 for a standard animal of 150 g wet mass at 08C.
21
In the North Sea octopus Eledone cirrhosa, respiration rates of 9.79 mg O h2 at 11.58C and 4.47
21
mg O h2 at 4.58C for a standard animal of 150 g wet mass were recorded. Respiration rates between P. charcoti and E. cirrhosa were compared using a combined Q10 value between P.
charcoti at 08C and E. cirrhosa at 4.58C. This suggests that P. charcoti are respiring at a level predicted by E. cirrhosa rates at 4.5 and 11.58C extrapolated to 08C along the curve Q1053, with no evidence of metabolic compensation for low temperature. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Energy balance; Cold adaptation; Antarctic; Octopus; Pareledone charcoti
1. Introduction
Analyses of energy utilisation in cephalopods are rare. A complete energy balance equation has been calculated for only two species of tropical octopus, Octopus cyanea and Octopus maya (Van Heukelem, 1976), with a partial energy budget constructed for
*Corresponding author. Tel.:144-1224-273-796; fax:144-1224-272-396.
E-mail address: [email protected] (H.I. Daly)
the temperate Octopus vulgaris (O’Dor and Wells, 1987). Recently, energetics in cephalopods from temperate and tropical waters has been reviewed by Wells and Clarke (1996). Energy balance in cold water species has received little attention and the recent successful maintenance of live Antarctic Pareledone charcoti in the BAS aquarium in Cambridge has provided an opportunity for a detailed study of energy balance in a polar species.
The flow of energy through an animal is normally represented by the energy balance equation:
C5P1G1R1U1F
where C is the total consumption of energy in the food, P and G are the values of somatic and gonadal growth, respectively, R is the energy used in respiration, U is the amount utilised in nitrogenous waste excretion and F is the energetic loss through faeces. The component R in the equation above can be divided into several portions which describe the utilisation of energy in more detail (Clarke, 1993; Wells and Clarke, 1996). Each portion of the respiratory costs involved in somatic growth (R ),S
reproductive investment (R ), basal metabolism (R ) or activity (R ) may varyG B A
independently with factors such as food availability or maturity. A substantial portion, termed the specific dynamic action (SDA), the increase in metabolic rate associated with feeding, may comprise the energetic cost of somatic (R ) and gonadal (R ) growthS G
(Clarke, 1993; Peck, 1998). These separate components of respiration highlight the potential for balancing the finite energy resource between each component, for maximum efficiency. In cephalopods, partitioning of resources probably differs between species, with the R component comprising a greater proportion of R in squid, due toA
their active swimming life-style, than in the more sedentary octopuses.
As well as constructing an energy budget for P. charcoti, the present study compares the respiration rates with a related species of the Eledoninae which occurs in temperate regions, Eledone cirrhosa. Also known as the lesser octopus, E. cirrhosa, is found around the British coastline and in the North Sea. The seasonal temperature range in the inshore North Sea is approximately 4–128C (McIntyre, 1958), and specimens have been maintained in the marine aquarium at the University of Aberdeen Zoology Department for a number of months at temperatures of 14–158C (Boyle, 1981). This study compares the respiration rates of P. charcoti at Antarctic temperatures of around 08C, to those of
E. cirrhosa acclimated from 11.58C down to 4.58C. Extrapolation of E. cirrhosa respiration rates to 08C will allow a direct comparison between the species.
One of the most widely used indices to assess the effect of temperature change on metabolism is the Q10 coefficient (Schmidt-Nielsen, 1991), which is essentially a measure of the temperature sensitivity of an organism. The Q10 value is calculated using the equation (Schmidt-Nielsen, 1991):
that metabolic reactions are slowed by approximately half when ambient temperature falls by 108C. A Q10 of less than 2 denotes insensitivity or acclimation to temperature change and a Q10of more than 2 denotes increased sensitivity, with an expected range of 1–3 (Calow, 1981). Q10 calculations for E. cirrhosa acclimated to two temperatures and a combined Q10 including both E. cirrhosa and P. charcoti has allowed the assessment and comparison of the effects of temperature.
2. Materials and methods
During the austral summers of 1993 and 1994, inshore trawls at depths of 50–150 m at Signy Island in the South Orkney Islands (608439S, 458389W) caught a number of specimens of P. charcoti. The specimens were maintained in the aquarium at the BAS station on Signy Island prior to being transferred by ship to the UK. The annual temperature range at this site varies from around 21.88C in winter to 10.38C in summer (Clarke et al., 1988; Clarke and Leakey, 1996).
In May 1993, a single male octopus (specimen 1) initially weighing 64 g live weight, was returned to the UK. It began feeding shortly after arrival and appeared healthy throughout the 18 months that it was maintained in the aquarium. The specimen fed irregularly on frozen / thawed krill Euphausia superba, live amphipods Bovallia gigantea and live mussels Mytilus edulis, and increased in weight to 74 g before dying in November 1994 after releasing a single spermatophore. In May 1994, a further 13 P.
charcoti were returned, from which two males (specimens 2 and 3) initially weighing 47
and 27 g wet weight, successfully established in the aquarium. They fed regularly on live isopods, Antarcturus sp., and mussels M. edulis, and increased in body weight until they both died in November 1994 weighing 51 and 30 g, respectively. The three healthy specimens showed apparently normal behaviour. They were all held in the cold aquarium system in Cambridge which is cooled to 060.58C (Ward and Peck, 1997), and were maintained in a light regime of 12-h dimmed light and 12-h darkness.
For the duration of the trial all the P. charcoti specimens were fed on M. edulis. All food was weighed before addition to each holding tank, and remains removed and weighed after feeding. Frequently, mussels had to be opened using a scalpel before the octopuses would feed. On their arrival in the aquarium live Antarctic isopods,
Antarcturus sp., were successfully used to feed the octopuses. Unfortunately, only a
small stock of live isopods were available for use as food. The frozen / thawed krill, E.
superba, were used as food for specimen 1 on its arrival, and this was only initially
successful when individual defrosted krill were placed under the web on a length of wire and moved gently until the octopus closed its arms and web around it. After feeding successfully for a few weeks during the trial all the specimens rejected any further food offered. Samples of the type of food consumed were dried to constant weight to obtain a conversion factor for wet to dry weights.
mass. The specimens were not anaesthetised, and the period of handling during weighing was kept brief to minimise stress to the animal.
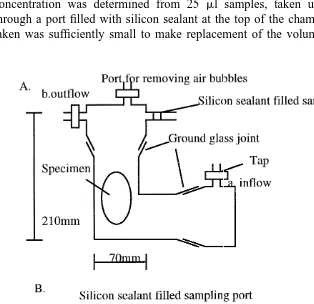
A 1015-ml glass respirometer chamber was used for trials with the 73 g specimen and a cylindrical 237 ml plastic respirometer was used for the two smaller specimens (Fig. 1). Both chambers were supplied with seawater from a raised header tank and had controllable flow-rates and ports filled with silicon sealant for taking samples (Fig. 1). To determine the oxygen consumption rates (V O ), each specimen was placed in the2
respirometer and it was sealed, making sure that there were no air bubbles trapped in the system. A gentle flow of oxygenated water was passed through the respirometer, at a
21
rate of approximately 600 ml min , for 1 h, while observing the behaviour of the specimen and recording the activity and time required for the animal to settle. After this period, the flow was stopped at taps A and B (Fig. 1), to seal the system. The oxygen concentration was determined from 25 ml samples, taken using a gas-tight syringe through a port filled with silicon sealant at the top of the chamber (Fig. 1). The sample taken was sufficiently small to make replacement of the volume removed unnecessary.
To ensure complete mixing of the water, the chamber was gently inverted three times before sampling. Very limited mantle ventilatory movements were observed, and the inversion movement did not appear to unduly disturb the animals. Samples were taken every 10–30 min and oxygen content obtained via injection into a couloximeter (Hersch, 1973; Peck and Uglow, 1990). During trials the respiration chambers were screened from the room to minimise visual disturbance. Oxygen levels in the respirometer were not allowed to fall below 100 mmHg to minimise the possibility of stress caused by low oxygen tension. Trials where air bubbles were observed in the chamber, or when the specimen became highly active were abandoned, so that only resting metabolism with minimum activity was recorded.
Nitrogen excretion trials were carried out alternating with the respiration trials over the same time period and using the same respiration chambers. Oxygenated water was
21
passed through the chamber with the specimen inside at approximately 600 ml min for 1 h before the chamber was sealed. Repeat samples of 20 ml were taken at the beginning and end of an hour-long trial, with the chamber being inverted three times before sampling to ensure complete mixing. Ammonia concentration was measured using the phenolhypochlorite method described by Liddicoat et al. (1975), as modified by Catalano (1987), and scaled to a sample volume of 10 ml (Clarke et al., 1994). Calibration was achieved by spiking seawater samples with a range of concentrations of ammonium sulphate solution.
V O2 and nitrogen excretion rates were obtained from comparisons with parallel controls (no animal in chamber).
All visible faeces were removed from the holding tanks daily and wet weight recorded. Although the faeces were delicate and were easily broken up on contact, care was taken during collection, and an estimated 80% was successfully recovered. As only small amounts of faeces were produced the error introduced into the energy budget during collection was small. Faeces were oven dried on filter paper and the dry weight recorded.
Healthy specimens of E. cirrhosa were obtained by the Department of Zoology, University of Aberdeen from local commercial trawl fishermen. In October 1994 several
E. cirrhosa specimens were transferred by rail from Aberdeen to the cold marine aquarium at BAS, Cambridge. The smallest specimens available were used to allow a more direct comparison of respiration rates, as they were closest in size to the P.
charcoti used earlier. Smaller specimens also experienced less stress when being placed
After a further 48 h acclimation time, respiration trials were carried out every 3 days for 9 days. The specimens were then transferred to 08C, but both died within 24 h of transfer. Body mass and volume were recorded at the beginning and end of the trials. Specimens usually remained inactive during trials but on occasion showed some increased activity in arm movements, which was noted. Trials where considerable activity, such as jetting, was recorded were abandoned.
Throughout the experiment the E. cirrhosa were not fed. The exact time that the specimens last fed before respiration trials began is unknown, but, assuming that they fed regularly in the wild, it would have been approximately 4 days prior to experimenta-tion. Attempts were made to provide live food (Carcinus maenas) at the start of the trial to establish a known feeding time, but this proved unsuccessful due to high prey mortality during transportation.
The joule has been used here as the energy unit, and the conversion factor to calories is 1 calorie54.184 J (Schmidt-Nielsen, 1991).
3. Results
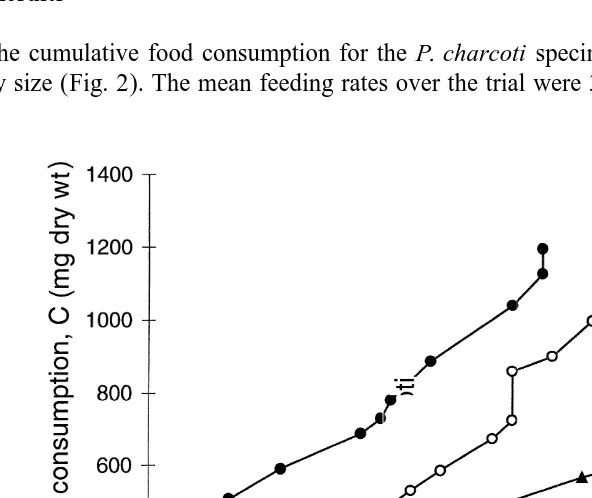
The cumulative food consumption for the P. charcoti specimens varied in relation to
21
body size (Fig. 2). The mean feeding rates over the trial were 31.44 mg day dry mass
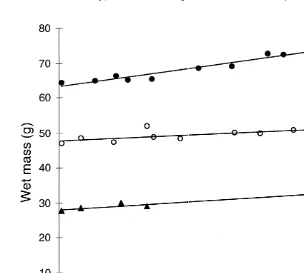
Fig. 3. Growth for Pareledone charcoti, as wet mass (g) increase excluding water released from the mantle cavity or web.
21 21
for specimen 1, 22.78 mg day dry mass for specimen 2 and 14.23 mg day dry mass for specimen 3.
The growth of the three specimens over the trial was very small (Fig. 3). The period over which growth was recorded in the P. charcoti specimens was relatively brief (up to 120 days), and the best fit to the observed growth over this period was linear. The slope of the regression line for each specimen was positive. When each slope was tested for significance of difference from zero, the slope for specimen 1 was significantly different from zero (t510.79, p,0.001), while the slopes for specimens 2 and 3 were not (t52.00, p.0.05 and t51.76, p.0.05, respectively). These results indicate that growth by specimens 2 and 3 over the trial was very slow. Instantaneous growth rates (G), expressed as the percent increase in body mass per day, were calculated from the equation:
ln W22ln W1
]]]]
G5 3100
t22t1
where W and W are body weights at times t and t , respectively (Forsythe and Van1 2 1 2
Heukelem, 1987). Daily growth rates were 0.13% for specimen 1, 0.08% for specimen 2 and 0.12% for specimen 3.
21
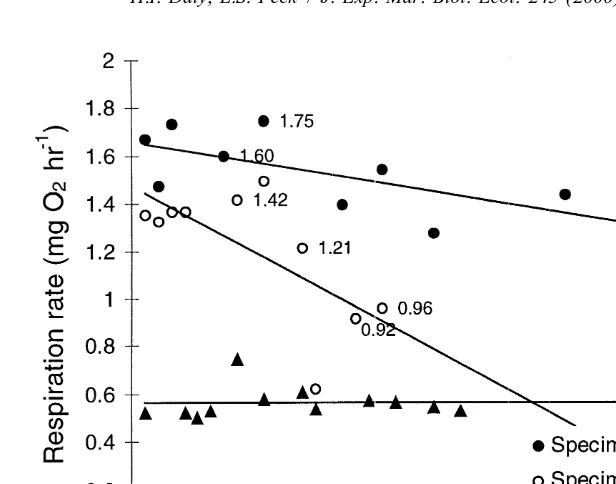
Fig. 4. Respiration rate R (mg O h2 ) for Pareledone charcoti with increasing time after last feeding event.
21
Points labelled with the respiration in mg O h2 indicate trials where increased activity of the specimen was recorded.
remained constant (specimens 3) (Fig. 4). Activity was recorded as low in most trials, and on the occasions where increased activity was noted (Fig. 4), there was not a noticeable elevation in metabolism. The mean respiration rates for P. charcoti are shown in Table 1, which includes comparative values from O’Dor and Wells (1987) for temperate and tropical octopods. All values were calculated for a standard sized animal of 1 kg wet mass using a scaling coefficient 0.75, obtained as a mean of the exponents for octopods listed in O’Dor and Wells (1987). The respiration rates listed are for inactive animals.
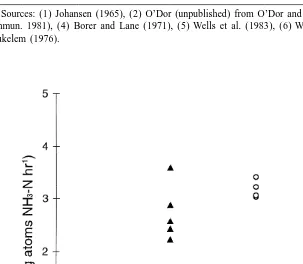
Nitrogen excretion increased with specimen size (Fig. 5). The mean excretion rates
21 21
recorded were 0.170 (S.D.50.007) mg atoms NH -N g3 h for specimen 1, 0.167
21 21
(S.D.50.004) mg atoms NH -N g3 h for specimen 2 and 0.203 (S.D.50.055) mg
21 21
atoms NH -N g3 h for specimen 3.
21
Production of faeces were at the rates of 2.742, 2.700 and 1.950 mg day dry mass for specimens 1, 2 and 3, respectively, over a period of around 80 days. The rates of production of faeces for specimens 1 and 2 were very similar although specimen 1 had a
21
higher mean consumption rate, 31.440 mg day dry mass compared to 22.781 mg
21
day dry mass for specimen 2. Faeces from specimen 1 may have broken up quickly in the tank before it could be collected, possibly by movement of the octopus, and may be underestimated.
Using mean V O2 and ammonia excretion rates, the atomic O:N ratio for each P.
Table 1
Respiration rates calculated for three Pareledone charcoti specimens and comparative mean values from O’Dor and Wells (1987) for a standard sized animal of 1 kg wet mass
a
Species Wet body Temperature Respiration rate Source
21 21
mass (g) (8C) (mg O kg2 h )
Pareledone charcoti 73 0 10.82 Present study
51 0 11.59 Present study
29 0 8.05 Present study
Eledone cirrhosa 312 4.5 18.53 Present study
E. cirrhosa 312 11 40.58 Present study
Octopus dofleini 9600 11 70.41 1
Bathypolypus arcticus 3 13 46.80 2
E. cirrhosa 564 18 61.37 3
Octopus briareus 345 20 45.64 4
Octopus vulgaris 817 21 84.07 5
O. vulgaris 42 22 73.40 6
Octopus cyanea 1000 25 125.71 7
O. briareus 348 30 99.94 4
a
Sources: (1) Johansen (1965), (2) O’Dor (unpublished) from O’Dor and Wells (1987), (3) Milne (pers. commun. 1981), (4) Borer and Lane (1971), (5) Wells et al. (1983), (6) Wells and Wells (1995), (7) Van Heukelem (1976).
21
Table 2
21 a
Components of the energy balance equation for Pareledone charcoti in J day (mW), for mean dry mass
Parameter Specimen 1 Specimen 2 Specimen 3
(9.51 g dry mass) (6.87 g dry mass) (4.01 g dry mass)
C 723.51 (8.37) 524.23 (6.07) 327.59 (3.79)
R 496.08 (5.74) 405.96 (4.70) 184.76 (2.14)
24 24 24
U 0.029 (3.39310 ) 0.022 (2.57310 ) 0.018 (2.04310 )
F 21.39 (0.25) 21.06 (0.24) 15.21 (0.18)
P1G 279.48 (3.23) 101.00 (1.17) 103.43 (1.20)
Scope for growth:
C2(R1U1F ) 206.01 (2.38) 97.19 (1.12) 127.61 (1.48)
Difference between P1G
and scope for growth 73.47 (0.85) 3.81 (0.04) 224.18 (0.28)
a
The mean dry mass of P. charcoti specimens were estimated using data for Pareledone turqueti.
trial and were 24.6 (range 22–27) for specimen 1, 27.2 (range 24–30) for specimen 2 and 13.3 (range 9–19) for specimen 3.
The previous results for C, R, U, F and P1G were used in the construction of an
energy budget for each of the P. charcoti specimens using conversion factors detailed below, and results are listed in Tables 2 and 3.
Energetic input from consumption (C ) was calculated using the conversion 23.01
21
J mg dry mass M. edulis flesh quoted in Thompson and Bayne (1974). The coefficient
21
used to determine the energetic cost of respiration (R), was 13.598 J mg O . This was2
based on fuelling metabolism using the relative percentage of protein, carbohydrate and lipid present in the M. edulis (Giese, 1969), which were fed to the P. charcoti. The
21 21
values 13.347 J mg O for protein respiration, 14.770 J mg2 O for carbohydrate2 21
respiration and 13.723 J mg O2 for lipid respiration from Brafield and Llewellyn (1982) were used for the calculation.
In calculating the amount of energy lost through excretion of nitrogenous waste
21
products (U ), the coefficient 288.278 J mol (Brafield and Soloman, 1972) was used. Only losses through ammonia excretion were considered, which were small in relation to other components in the energy balance equation. The energetic content of octopus
21
faeces (F ), determined by bomb calorimetry in Van Heukelem (1976) as 7.799 J mg dry mass, was used in the calculation.
21
The value 22.259 J g dry mass was used as the energy composition for octopus tissues, combining somatic (P) and gonadal (G) growth. This figure was obtained as a
Table 3
Components of the energy budget for P. charcoti expressed as a percentage of consumption (C) Parameter Specimen 1 Specimen 2 Specimen 3
R 68.57 77.44 56.40
U 0.004 0.004 0.005
F 2.96 4.02 4.64
P1G 38.63 19.27 31.57
mean value from O. vulgaris (Wissing et al., 1973) and O. cyanea (Van Heukelem, 1976), based on bomb calorimetry. As no specimens of P. charcoti were available for estimation of water content, estimates obtained from Pareledone turqueti, a similar Antarctic species, were used to calculate dry mass (Daly, 1996).
Energy budgets for all three P. charcoti specimens were approximately in balance (Table 2) Each energetic component is also shown in milliwatts. The scope for growth
21
calculation underestimated the recorded growth (P1G) by 73.469 J day in specimen
21
1, overestimated it by 24.180 J day in specimen 3, and almost balanced in specimen 2,
21
underestimating by only 3.810 J day . Table 3 illustrates the results expressed as a percentage of C and shows that the proportions calculated for each specimen were broadly consistent. Assimilation efficiency for P. charcoti was calculated as:
C2F
]] Assimilation efficiency (%)5 3100
C
The values 97.04, 95.98 and 95.36% were obtained for specimens 1, 2 and 3, respectively.
3.1. Assessment of cold adaptation in P. charcoti by comparison with E. cirrhosa
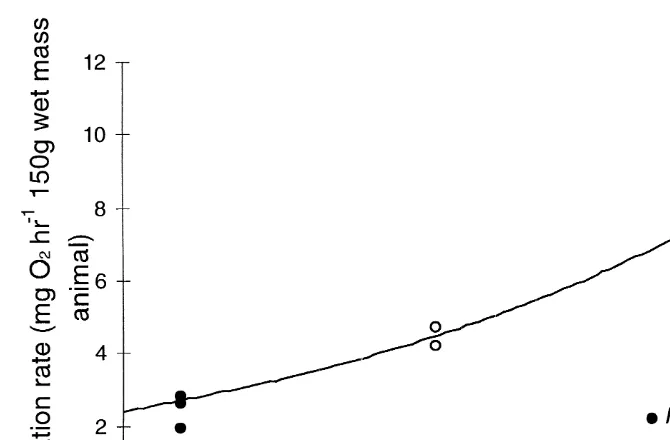
Respiration rate values for both species were corrected to standard wet animal mass of 150 g using the weight exponent 0.75 (O’Dor and Wells, 1987), as this was approximately half way between the masses of the P. charcoti and E. cirrhosa specimens used (Table 4). A Q10 of 3.063 was calculated for E. cirrhosa between 11.5 and 4.58C. As Q10 calculations require respiration rates at two temperatures, a value could not be obtained for P. charcoti alone as respiration was only measured at one temperature (08C). In order to compare results for P. charcoti and E. cirrhosa, a combined Q10 value was calculated including the respiration rates for P. charcoti with the result for E. cirrhosa at 4.58C. This produced a Q10 of 3.807. The combined respiration rate data for the two species fits the curved Q10 relationship of 3 (Fig. 6), suggesting that P. charcoti respire at a rate consistent with extrapolation from E.
cirrhosa rates.
Table 4
Comparative respiration rates for individual Pareledone charcoti specimens at 08C and Eledone cirrhosa at 4.5 and 11.58C for a standard animal of 150 g wet mass
Specimen Wet body Temperature Respiration rate
21
E. cirrhosa 323 11.5 11.05
Fig. 6. Comparative respiration rate for Pareledone charcoti at 08C and Eledone cirrhosa at 4.5 and 11.58C for a standard animal of 150 g wet mass. The curve, Q1053, was calculated using the mean respiration rate of
E. cirrhosa at 4.58C.
4. Discussion
Low temperature is a major factor affecting metabolic and growth rates in polar organisms (Clarke, 1998; Somero, 1998). Clarke (1983, 1991a) also argued that the observed slow growth rates in most marine polar invertebrates studied may be due to a strongly seasonal brief period of growth, which, when averaged over a year appears slow. Thus, the limitation of resources during winter, rather than suppression by low temperature would be responsible for slow growth. However, significant winter growth or year round growth have been found in bryozoans (Barnes, 1995), brachiopods (Peck et al., 1997), and the bivalve Yolida eightsi (Peck, unpubl. data). The impact of reduced resources in the form of low primary production is likely to be greatest on species low in the food chain, such as suspension feeders, with organisms higher in the food chain, such as predatory cephalopods, buffered from the effect to some extent. Temperature has also been shown to be the main factor controlling development in marine invertebrate larvae (Hoegh-Guldberg and Pearse, 1995), where very long development periods have been found in echinoderms (Bosch et al., 1987; Pearse et al., 1991; Stanwell-Smith and Peck, 1998), brachiopods, (Peck and Robinson, 1994), nemerteans (Peck, 1993), isopods
¨
One of the striking biological characteristics in cephalopods is rapid growth, which was shown to vary from 2.00 to 4.65% per day at around 100 g wet body mass in a range of octopod species at 15–258C (Forsythe and Van Heukelem, 1987). The slow growth of 0.13, 0.08 and 0.12% increase in body mass per day recorded for the P.
charcoti specimens (Fig. 3), demonstrate that a potentially different life history strategy
may be adopted by polar cephalopods. Antarctic Pareledone may have a similar strategy to the Arctic octopod Bathypolypus arcticus, which had a growth rate of 0.18% body mass increase per day, estimated using linear regression over an 18-month period (O’Dor and Macalaster, 1983). In B. arcticus a 3.7 g increase over a month was recorded, with a final mass of 70 g, and a proposed life-span of up to 4 years (O’Dor and Macalaster, 1983). The life-span of P. charcoti is not known, but the male specimen, which was held in the aquarium at BAS, Cambridge survived for 18 months, and was collected at least 3 months prior to this.
Respiration rates for a number of octopus species have been summarised in Table 1
21
and mean V O for a 312 g wet mass E. cirrhosa fell from 40.58 to 18.53 mg O kg2 2 21
h at 11.5 and 4.58C, respectively. Respiration in a small Bathypolypus arcticus
21 21
specimen was recorded as 46.80 mg O kg2 h at 138C (Table 1). This temperature is considerably higher than their norm of 4628C (Macalaster, 1976) and is comparable to the rates recorded for E. cirrhosa in this study. In P. charcoti, respiration fell very slowly from routine to a low constant rate, which is close to standard levels (Fig. 4),
21 21
from 10.47 to 9.52 mg O kg2 h in 40 days in specimen 1, and 13.92 to 8.91 mg O2 21 21
kg h in 30 days in specimen 2, compared with the rapid decline recorded by Wells and Wells (1995) for O. vulgaris (approx. 44 g wet body mass), dropping from 162 to 80
21 21
mg O kg2 h after 5 days of fasting at around 228C. Similar slow reductions in metabolism to standard levels were recorded in other Antarctic benthic invertebrates by Peck (1989) where the limpet Nacella concinna and the brachiopod Liothyrella uva took 25–50 days to return to standard or basal levels.
As octopods have well developed nervous and visual systems, they can be easily stressed by handling and external movement. When metabolic rates are raised after disturbance by tank cleaning or handling, they can remain increased for around an hour in temperate species (Wells et al., 1983). It is possible that the time required for the metabolic rate of a polar cephalopod to return to normal after the stress of handling is considerably longer (or shorter) than an hour. However, in the absence of previous data a settling period of one hour between placing the animals in the chamber and taking samples was used to reduce the effect of the initial stress. The P. charcoti usually settled quickly in the chambers and often remained motionless during the trial, suggesting that any stresses were small.
Although ammonia is the dominant excretory product in most marine invertebrates (Bayne et al., 1985; Regnault, 1987), it is not the only nitrogenous compound excreted. Hoeger et al. (1987), reported that urea comprised 30% of the nitrogen excretion in
Octopus rubescens. As we only measured ammonia excretion, the final energetic cost of
excretion may be underestimated. However, nitrogenous waste comprises ,0.01% of the total energy budget, so the error created by recording only ammonia levels is probably small. The excretion rates in the starved P. charcoti at 08C were similar, with a
21 21
21 21
Boucher-Rodoni and Mangold (1985) of 0.560 mg atoms NH -N g3 h in O. vulgaris
21 21
at 15–168C, which had been recently feeding, and 0.307 mg atoms NH -N g3 h in starved animals.
Atomic O:N ratios are linked to the availability of energy stores and the utilisation of body protein. This ratio of oxygen consumption to nitrogen excretion by atomic equivalents produces an index of the relative amounts of protein, compared to carbohydrate and lipid that are being catabolised by the organism. Theoretically, the minimum value for an O:N ratio is around 7, when only protein is being catabolised. Mayzaud and Conover (1988) calculated that organisms catabolising equal proportions of lipid and protein would have an O:N ratio of 24, with values more than this indicating lipid or carbohydrate as the major component in catabolism, and values less indicating increased protein catabolism. The values for specimens 1 and 2 indicate protein accounting for around half of the catabolic substrate, while specimen 3 is catabolising a higher proportion of protein. O:N ratios can be variable, and may fall or rise in response to starvation, depending on the biochemical composition of the organism (Mayzaud and Conover, 1988). In fed O. vulgaris at 15–168C mean O:N ratios of 7.19 and 14.15 were recorded, indicating mainly protein catabolism, while ratios of 9.62 and 24.76 were recorded in starved animals, denoting increased utilisation of lipid or carbohydrate stores (Boucher-Rodoni and Mangold, 1985), which is consistent with the O:N ratios obtained for P. charcoti.
The results in Table 2 represent an approximately balanced energy budget for P.
charcoti, although the scope for growth underestimates the recorded somatic and
gonadal growth (P1G) for specimens 1 and 2, and overestimates growth in specimen 3.
As only resting metabolism was recorded, the activity component of energy utilisation was not estimated. Its inclusion would increase the imbalance in the budgets for specimens 1 and 2, but decrease the underestimation for specimen 3. However, observations indicated that natural long term levels of activity were low. Other possible errors include the loss of tissues in holding systems. Octopods kept in static water holding tanks are often observed to shed skin, mucus and sucker rings, which contribute to energetic loss. In Octopus cyanea, where shed sucker rings have been observed to accumulate in the water, Van Heukelem (1976) recorded the loss as 1.5% of the total energy budget. However, in the static water holding tanks of the P. charcoti specimens in this study, shed skin, mucus and sucker rings were not frequently observed, and the associated energetic loss is likely to be small, and probably less than the 1.5% observed in O. cyanea. Mucus production may also be important in mollusc energy budgets and accounted for nearly 30% of consumed energy in the abalone Haliotis tuberculata (Peck et al., 1987). However, there are no reports of cephalopods producing large amounts of mucus and this error is also likely to be small.
which refused food for part of the trial. This suggests that, although P. charcoti are respiring at a lower level than O. cyanea (Table 1), the proportion of the energy budget required for maintenance is greater, even when periods of reduced food intake are included, when there is a reduced effect of SDA. This increased proportion of energy for basal metabolism in a polar organism could be interpreted as evidence for metabolic cold adaptation. However, numbers involved in these trials were small and such conclusions are difficult to apply with confidence. Growth would also have been slow if the animals were nearing the end of their normal life-span.
Using results from Milne, A.H. (pers. commun.) for E. cirrhosa at 18.58C, a Q10 was also calculated for E. cirrhosa between 18 and 11.58C, producing a value of 1.89. which is well within the normal range. Coefficients for other octopods of 2.14 for Octopus
cyanea around 308C (Van Heukelem, 1976), and 2.18 for Octopus briareus between 20 and 308C (Borer and Lane, 1971) have been recorded, and the Q10 for E. cirrhosa between 11.5 and 4.58C of 3.063 is also within the expected range. The inclusion of P.
charcoti at 08C and E. cirrhosa at 4.58C in a combined Q10 calculation produces the value 3.807. This higher value may indicate that P. charcoti has a metabolic rate even slower than that extrapolated for E. cirrhosa if they could survive at 08C. If P. charcoti was displaying metabolic cold adaptation (MCA) and functioning with an elevated metabolic rate in comparison with E. cirrhosa, the Q10 would be lower than 3.063. However, as the Q10 function describes a curve (Fig. 6), it appears that P. charcoti are respiring at a rate predicted by E. cirrhosa rates extrapolated along this curve to low temperatures. Precht (1958) describes an increase in Q10 values with an increase in the temperature range as ‘inverse’ compensation, i.e. rather than adjusting metabolism to maintain it constant, the changes make it more disparate. O’Dor and Wells (1987) termed this ‘paradoxical’ adaptation, and discuss its benefit to squid making daily vertical migrations from cold deep water to warm surface water for feeding. This is when animals go into torpor at low temperatures or when they increase their activity level to increase the feeding rate to meet increased energy demands at higher temperatures.
The P. charcoti–E. cirrhosa Q10 value is within the range found for other Antarctic invertebrates; Peck (1989) recorded Q10 values of 2.41 in acclimated Nacella concinna from21.5 to14.58C, and Luxmoore (1984) calculated values of 2.00 for Serolis polita and 4.21 for Serolis cornuta between21.5 and 14.58C. The low respiration rates in P.
charcoti support recent conclusions that MCA does not occur in polar marine
invertebrates (Luxmoore, 1984; Clarke, 1991a,b, 1993).
Acknowledgements
References
Barnes, D.K.A., 1995. Seasonal and annual growth in erect species of Antarctic bryozoans. J. Exp. Mar. Biol. Ecol. 188, 181–198.
Bayne, B.L., Brown, D.A., Burns, K., Dixon, D.R., Ivanovici, A., Livingstone, D.R., Lowe, D.M., Moore, M.N., Stebbing, A.R.D., Widdows, J. (Eds.), 1985. The Effects of Stress and Pollution On Marine Animals, Praegar, New York.
Borer, K.T., Lane, C.E., 1971. Oxygen requirements of Octopus briareus Robson at different temperatures and oxygen concentrations. J. Exp. Mar. Biol. Ecol. 7, 263–269.
Bosch, I., Beauchamp, K.A., Steele, M.E., Pearse, J.S., 1987. Development, metamorphosis and seasonal abundance of embryos and larvae of the Antarctic sea urchin Sterechinus neumayeri. Biol. Bull. 173, 126–135.
Boucher-Rodoni, R., Mangold, K., 1985. Ammonia excretion during feeding and starvation in Octopus
vulgaris. Mar. Biol. 86, 193–197.
Boyle, P.R., 1981. Methods for the aquarium maintenance of the common octopus of British waters, Eledone
cirrhosa. Lab. Anim. 15, 327–331.
Brafield, A.E., Soloman, D.J., 1972. Oxycalorific coefficients for animals respiring nitrogenous substrates. Comp. Biochem. Physiol. 43A, 837–841.
Brafield, A.E., Llewellyn, M.H., 1982. Animal Energetics, Blackie, Glasgow. Calow, P., 1981. Invertebrate Biology, Halsted Press, New York.
Catalano, G., 1987. An improved method for the determination of ammonia in seawater. Mar. Chem. 20, 289–295.
Clarke, A., 1983. Life in cold water; the physiological ecology of polar marine invertebrates. Biol. J. Linn. Soc. 14, 77–92.
Clarke, A., 1991a. What is cold adaptation and how should we measure it? Am. Zool. 31, 81–92. Clarke, A., 1991b. Cold adaptation. J. Zool. Lond. 225, 691–699.
Clarke, A., 1993. Seasonal acclimatisation and latitudinal compensation in metabolism, do they exist? Func. Ecol. 7, 139–149.
Clarke, A., 1998. Temperature and energetics, an introduction to cold water physiology. In: Portner, H.O., Playle, R.C. (Eds.), Cold Ocean Physiology, Soc. Exp. Biol. Semin. Ser. 66, pp. 3–30.
Clarke, A., Leakey, R.J.G., 1996. The seasonal cycle of phytoplankton, macronutrients, and the microbial community in a nearshore Antarctic marine ecosystem. Limnol. Oceanogr. 31, 1281–1294.
Clarke, A., Holmes, L.J., White, M.G., 1988. The annual cycle of temperature, chlorophyll and major nutrients at Signy Island, South Orkney Islands, 1969–82. Br. Antarct. Surv. Bull. 80, 65–86.
Clarke, A., Prothero-Thomas, E., Whitehouse, M.J., 1994. Nitrogen excretion in the Antarctic limpet Nacella
concinna (Strebel, 1908). J. Moll. Stud. 60, 141–147.
Cossins, A.R., Bowler, K., 1987. Temperature Biology of Animals, Chapman and Hall, London.
Daly, H.I., 1996. Ecology of the Antarctic octopus Pareledone from the Scotia Sea. PhD dissertation, University of Aberdeen, 162 pp.
Dayton, P.K., 1989. Interdecadal variation in an Antarctic sponge and its predators from oceanographic climate shirts. Science 245, 1484–1486.
Forsythe, J.W., Van Heukelem, W.F., 1987. Growth. In: Boyle, P.R. (Ed.), Cephalopod Life Cycles, Comparative Reviews, Vol. II, Academic Press, London, pp. 135–156.
Giese, A.C., 1969. A new approach to the biochemical composition of the mollusc body. Oceanogr. Mar. Biol. Annu. Rev. 7, 175–229.
Hain, S., 1991. Life maintenance of benthic mollusks from the Eastern Weddell Sea, Antarctica. In: Proc. 10th Int. Malacol. Cong., Tubingen, 1989, pp. 339–341.
Hersch, P.A., 1973. Couloximetry. Am. Lab. 5, 29–36.
Hoeger, U., Mommsen, T.P., O’Dor, R.K., Webber, D., 1987. Oxygen uptake and nitrogen excretion in two cephalopods, octopus and squid. Comp. Biochem. Physiol. 47 (3), 137–152.
Hoegh-Guldberg, O., Pearse, J.S., 1995. Temperature, food availability, and the development of marine invertebrate larvae. Am. Zool. 35, 415–425.
Liddicoat, M.I., Tibbitts, S., Butler, E.I., 1975. The determination of ammonia in seawater. Limnol. Oceanogr. 20, 131–132.
Luxmoore, R.A., 1984. A comparison of the respiration rates of some Antarctic isopods with species from lower latitudes. Br. Antarc. Surv. Bull. 62, 53–65.
Macalaster, E.G., 1976. The natural history and biology of the deep-water octopus Bathypolypus arcticus (Prosch). MSc thesis, Dalhousie University, Nova Scotia.
Mayzaud, P., Conover, R.J., 1988. O:N atomic ratios as a tool to describe zooplankton metabolism. Mar. Ecol. Prog. Ser. 45, 289–302.
McIntyre, A.D. (Ed.), 1958. The ecology of Scottish inshore fishing grounds. I. The bottom fauna of East Coast grounds, Mar. Res. 1, Scottish Home Dept, p. 24.
O’Dor, R.K., Macalaster, E.G., 1983. Bathypolypus arcticus. In: Boyle, P.R. (Ed.), Cephalopod Life Cycles, Species Accounts, Vol. I, Academic Press, London, pp. 401–410.
O’Dor, R.K., Wells, M.J., 1987. Energy and nutrient flow. In: Boyle, P.R. (Ed.), Cephalopod Life Cycles, Comparative Reviews, Vol. II, Academic Press, London, pp. 109–133.
Pearse, J.S., McClintock, J.B., Bosch, I., 1991. Reproduction of Antarctic benthic invertebrates, tempos, modes and timing. Am. Zool. 31, 65–80.
Peck, L.S., 1989. Temperature and basal metabolism in two Antarctic marine herbivores. J. Exp. Mar. Biol. Ecol. 127, 1–12.
Peck, L.S., 1993. Larval development in the Antarctic nemertean Parborlasia corrugatus (Heteronemertea, Lineidae). Mar. Biol. 116, 301–310.
Peck, L.S., 1998. Feeding, metabolism and metabolic scope in Antarctic marine ectotherms. In: Portner, H.O., Playle, R.C. (Eds.), Cold Ocean Physiology, Soc. Exp. Biol. Semin. Ser. 66, pp. 365–390.
Peck, L.S., Uglow, R.F., 1990. Two methods for the assessment of the oxygen content of small volumes of seawater. J. Exp. Mar. Biol. Ecol. 141, 53–62.
Peck, L.S., Robinson, K., 1994. Pelagic larval development in the brooding Antarctic brachiopod Liothyrella
uva. Mar. Biol. 120, 279–286.
Peck, L.S., Culley, M.B., Helm, M.M., 1987. A laboratory energy budget for the ormer Haliotis tuberculata L. J. Exp. Mar. Biol. Ecol. 106, 103–123.
Peck, L.S., Brockington, S., Brey, T., 1997. Growth and metabolism in the Antarctic brachiopod Liothyrella
uva. Philos. Trans. R. Soc. 352, 851–858.
Precht, H., 1958. Concepts of the temperature adaptation of unchanging reaction systems of cold-blooded animals. In: Prosser, C.L. (Ed.), Physiological Adaptation, Lord Baltimore Press, Baltimore, pp. 51–77.
¨
Rauschert, M., 1991. Ergebnisse der faunistischen arbeiten im benthal von King George Island (Sudshetlandin-seln, Antarctis). Ber. Polarforsch. 76, 1–75.
Regnault, M., 1987. Nitrogen excretion in marine and fresh-water crustacea. Biol. Rev. 62, 1–24.
Schmidt-Nielsen, K., 1991. Animal Physiology, Adaptation and Environment, Cambridge University Press, Cambridge.
Somero, G.N., 1998. Adaptation to cold and depth, contrasts between polar and deep-sea animals. In: Portner, H.O., Playle, R.C. (Eds.), Cold Ocean Physiology, Soc. Exp. Biol. Semin. Ser. 66, pp. 33–57.
Stanwell-Smith, D.P., Peck, L.S., 1998. Temperature and embryonic development in relation to spawning and field occurrence of larvae of three Antarctic echinoderms. Biol. Bull. Woods Hole 194, 44–52.
Thompson, R.J., Bayne, B.L., 1974. Some relationships between growth, metabolism and food in the mussel
Mytilus edulis. Mar. Biol. 27, 317–326.
Van Heukelem, W.F., 1976. Growth, bioenergetics and life-span of Octopus cyanea and Octopus maya. PhD dissertation, University of Hawaii, 224 pp.
¨
Wagele, J.W., 1987. On the reproductive biology of Ceratoserolis trilobitoides (Crustacea, Isopoda), latitudinal variation of fecundity and embryonic development. Polar Biol. 7, 11–24.
Ward, J.P., Peck, L.S., 1997. The coldwater marine aquarium at the British Antarctic Survey. Aquar. Sci. Conserv. 1, 56–63.
Wells, M.J., Clarke, A., 1996. Energetics, the cost of living and reproducing for an individual cephalopod. Philos. Trans. R. Soc. Lond. B. 351, 1083–1104.
Wells, M.J., Wells, J., 1995. The control of ventilatory and cardiac responses to changes in ambient oxygen tension and oxygen demand in octopus. J. Exp. Biol. 198, 1717–1727.
White, M.G., 1970. Aspects of the breeding biology of Glyptonotus antarcticus (Eights) (Crustacea, Isopoda) at Signy Island, South Orkney Islands. In: Holgate, M.W. (Ed.), Antarctic Ecology, Vol. 1, Academic Press, London, pp. 279–285.