PERKECAMBAHAN EMBRIO ENDOSPERMIK MANGGA
(
Mangifera indica
L.) VARIETAS ARUMANIS KLON 143
SECARA
IN VITRO
EKA SETIYANI
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Perkecambahan Embrio Endospermik Mangga (Mangifera indica L.) Varietas Arumanis Klon 143 secara In Vitro adalah benar karya saya dengan arahan dari dosen pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Septermber 2013 Eka Setiyani NIM A24080052
ABSTRAK
EKA SETIYANI. Perkecambahan Embrio Endospermik Mangga (Mangifera indica L.) Varietas Arumanis Klon 143 secara In Vitro. Dibimbing oleh NI MADE ARMINI WIENDI.
Penelitian ini bertujuan memperoleh media yang optimal untuk perkecambahan embrio endospermik mangga (Mangifera indica L.) varietas Arumanis klon 143. Perlakuan suhu rendah dan GA3 tidak berpengaruh terhadap perkecambahan embrio endospermik fase kotiledonari mangga (Mangifera indica L.) varietas Arumanis klon 143. Pada percoban I, embrio dengan perlakuan suhu rendah 8 hari menghasilkan proembrio sekunder (83.3%) dan 33 embrio sekunder. Pada percobaan II, sebanyak 83.3% embrio membentuk proembrio sekunder pada media MS dengan 5 µM GA3. Pada percobaan III, media padat yang mengandung media MS dengan 5 mg/l BA menghasilkan embrio sekunder sebanyak 80 embrio, lebih baik dibandingkan media cair. Sebanyak 6.7% embrio berkecambah pada media padat. Pada percobaan IV, media proembrio berpengaruh nyata terhadap embrio yang dihasilkan inokulum, sedangkan media prekultur tidak berpengaruh nyata. Inokulum dari media MS dengan 10 µM GA3 yang dikulturkan pada media prekultur PM menghasilkan rata-rata embrio terbanyak yaitu 25.67 embrio pada 7 MST. Persentase embrio berkecambah terbanyak 32.4% dihasilkan oleh inokulum yang berasal dari media MS dengan setengah konsentrasi hara makro dan mikro ditambah 5% air kelapa yang dikulturkan pada media prekultur PM. Kata kunci: air kelapa, embrio sekunder, GA3, media prekultur, perlakuan suhu
rendah
ABSTRACT
EKA SETIYANI. In Vitro Germination of the Endospermical Embryos in Manggo (Mangifera indica L.) cv. Arumanis Clone 143. Supervised by NI MADE ARMINI WIENDI.
This study aimed to obtain the optimal medium for germination of endospermical embryos in mango (Mangifera indica L.) cv Arumanis clone 143. The effect of cold treatment and GA3 treatment had no significant difference in endospermical embryos germination. In the first experiment, embryos with 8 days cold treatment 83.3% of cotiledonary embryos produced the secondary proembryos with 33 secondary embryos. In the second experiment, the highest percentage of secondary proembryos (83.3%) we obtain in MS medium with 5 µM GA3. In the third experiment, the solid MS medium with 5 mg/l BA produced 80 secondary embryos, highest than in liquid medium with the similar composition, and 6.7% of that embryos germinated on solid medium. In the fourth experiment, the embryo medium induction has significant difference effect in embryo which produced by inoculum, whereas preculture medium had no significant difference effect. Inoculum from MS medium with 10 µM GA3 and cultured on PM preculture medium (MS medium with 5 mg/l 2,4-D, 5 mg/l GA3, 400 mg/l amino acid glutamine, 300 mg/l thrypton and 200 ml/l coconut water)
produced embryo at 25.67 in 7 MST. The highest percentage of germinated embryos (32.4%) produced by inoculum from medium containing MS half-strength major and minor nutrients with 5% coconut water, and then cultured on PM preculture medium.
Keywords: coconut water, cold treatment, GA3, preculture medium, secondary embyo
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Agronomi dan Hortikultura
PERKECAMBAHAN EMBRIO ENDOSPERMIK MANGGA
(
Mangifera indica
L.) VARIETAS ARUMANIS KLON 143
SECARA
IN VITRO
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
2013
Judul Skripsi : Perkecambahan Embrio Endospermik Mangga (Mangifera indica L.) Varietas Arumanis Klon 143 secara In Vitro Nama : Eka Setiyani
NIM : A24080052
Disetujui oleh
Dr Ir Ni Made Armini Wiendi, MS Pembimbing
Diketahui oleh
Dr Ir Agus Purwito, MScAgr Ketua Departemen
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas petunjuk, kekuatan dan kesabaran yang diberikan kepada penulis sehingga dapat melaksanakan penelitian dan penulisan skripsi yang berjudul “Perkecambahan Embrio Endospermik Mangga (Mangifera indica L.) Varietas Arumanis Klon 143 secara In Vitro”.
Pada kesempatan ini penulis ingin menyampaikan terima kasih dan penghargaan kepada:
1. Dr. Ir. Ni Made Armini Wiendi, MS selaku dosen pembimbing yang dengan sabar membimbing penulis selama melaksanakan penelitian dan penulisan skripsi.
2. Dr. Darda Efendi dan Dr. Dewi Sukma sebagai dosen penguji pada ujian skripsi atas masukan yang sangat bermanfaat untuk perbaikan skripsi ini. 3. Dr. Nurul Khumaida selaku dosen pembimbing akademik yang telah
membimbing penulis dalam melaksanakan studi di Departemen Agronomi dan Hortikultura IPB.
4. Orang tua (Bapak Sumardi dan Ibu Ruwatiah), adik tercinta (Dewi Pujiati), dan seluruh keluarga atas do’a, perhatian, dukungan moril dan materil serta kasih sayang yang tiada henti.
5. Rekan-rekan di Lab. Kultur Jaringan 2 atas bantuan teknis dan motivasi dalam melaksanakan penelitian.
6. Rekan-rekan di departemen AGH angkatan 45 atas kebersamaan dan persaudaraan yang telah terjalin selama menjalani studi di Departemen Agronomi dan Hortikultura IPB.
Penulis berharap agar skripsi ini dapat memberikan informasi yang bermanfaat bagi pihak-pihak yang memerlukan.
Bogor, September 2013 Eka Setiyani
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR ix DAFTAR LAMPIRAN x PENDAHULUAN 1 Latar Belakang 1 Tujuan Penelitian 2 TINJAUAN PUSTAKA 4
BAHAN DAN METODE 7
Waktu dan Tempat 7
Bahan dan Alat 7
Rancangan Penelitian 7
HASIL DAN PEMBAHASAN 11
Percobaan I. Perkecambahan Embrio Endospermik Mangga Varietas
Arumanis Klon 143 dengan Perlakuan Suhu Rendah 11 Percobaan II. Perkecambahan Embrio Endospermik Mangga Varietas
Arumanis Klon 143 dengan Perlakuan Giberelin 15 Percobaan III. Perkecambahan Embrio Endospermik Mangga
Varietas Arumanis Klon 143 dengan Perlakuan Media
Prekultur yang Berbeda 18
Percobaan IV. Perkecambahan Embrio Endospermik Mangga Varietas
Arumanis Klon 143 dengan Media Cair dan Padat 20
KESIMPULAN 26
DAFTAR PUSTAKA 26
LAMPIRAN 29
RIWAYAT HIDUP 31
DAFTAR TABEL
1 Komposisi media prekultur dan media perkecambahan embrio endospermik mangga Arumanis klon 143 pada percobaan IV 11 2 Persentase kontaminasi dari embrio endospermik mangga Arumanis
klon 143 setelah perlakuan suhu rendah pada percobaan I 12 3 Persentase embrio endospermik mangga Arumanis klon 143 yang
4 Bobot dan diameter rata-rata proembrio yang dihasilkan embrio endospermik mangga Arumanis klon 143 dengan perlakuan suhu
rendah pada umur 10 MST 14
5 Jumlah embrio endospermik sekunder yang terbentuk dari embrio endospermik mangga Arumanis klon 143 dengan perlakuan suhu
rendah umur 14 MST 14
6 Persentase kontaminasi embrio endospermik mangga Arumanis klon
143 pada perlakuan GA3 15
7 Persentase embrio endospermik mangga Arumanis klon 143 yang
berproliferasi pada perlakuan GA3 16
8 Bobot dan diameter rata-rata proembrio yang dihasilkan embrio endospermik mangga Arumanis klon 143 pada media perlakuan GA3 18 9 Jumlah embrio endospermik sekunder mangga Arumanis klon 143 yang
terbentuk pada media cair dan padat saat berumur 7 MST 19 10 Presentase kontaminasi inokulum pada 4 MST di media perkecambahan
(MP) 20
11 Analisis ragam pengaruh media prekultur dan media asal inokulum terhadap embrio endospermik mangga Arumanis klon 143 fase
kotiledonari yang dihasilkan 23
12 Pengaruh media prekultur dan media asal inokulum terhadap rata-rata embrio endospermik mangga Arumanis klon 143 fase kotiledonari yang
dihasilkan pada 4 MST 23
13 Pengaruh media prekultur dan media asal inokulum terhadap rata-rata embrio endospermik mangga Arumanis klon 143 fase kotiledonari
yang dihasilkan pada 7 MST 24
14 Persentase embrio endospermik mangga Arumanis klon 143 yang berkecambah pada 9 MST di media perkecambahan (MP) 25
DAFTAR GAMBAR
1 Bagan alur penelitian embriogenesis sel endosperma untuk perakitan
tanaman triploid mangga Arumanis klon 143 3
2 Bagan alur percobaan IV 11
3 Kultur embrio endospermik mangga Arumanis klon 143 yang terkontaminasi: (A) inokulum, (B) kontaminan cendawan, (C)
kontaminan bakteri 12
4 A) Embrio endospermik mangga Arumanis klon 143 yang berproliferasi setelah perlakuan suhu rendah pada 10 MST; B) Proembrio yang dihasilkan embrio endospermik dengan perlakuan suhu rendah; C) Embrio endospermik sekunder yang dihasilkan saat 12 MST 13 5 Proliferasi embrio endospermik mangga Arumanis klon 143 umur
10 MST; A) pada media MS0; B) pada media MS1/2; C) pada media
MS-5G; D) pada media MS-10G 16
6 Pembentukan proembrio dan embrio sekunder mangga Arumanis klon 143 pada media cair dan padat: A) kultur pada media cair awal tanam; B) kultur pada media cair 3 MST; C) kultur pada media cair 5 MST; D)
kultur pada media padat awal tanam; E) kultur pada media padat 3 MST; F) kultur pada media padat 5 MST; G) embrio endospermik
sekunder pada 5 MST di media padat 19
7 Perkembangan embrio endospermik mangga Arumanis klon 143 yang bekecambah pada media padat: A) 3 MST; B) 4 MST; C) 5 MST; D)
10 MST 20
8 Proliferasi proembrio mangga Arumanis klon 143 di media prekultur
pada umur 7 MST: A) media PM; B) media M3 21
9 Proembrio endospermik mangga Arumanis klon 143 umur 4 MST pada media induksi embrio (MI) dari media prekultur berbeda: A) inokulum asal media prekultur PM; B) inokulum asal media prekultur M3 22 10 Pembentukan embrio endospermik mangga Arumanis klon 143 umur
4 MST pada media perkecambahan (MP) dengan perlakuan prekultur:
A) media PM; B) media M3 22
11 Berbagai bentuk embrio endospermik mangga Arumanis klon 143 fase kotiledonari pada media perkecambahan (MP) pada umur 7 MST 24 12 Embrio berkecambah pada media perkecambahan (MP): A) inokulum
dengan perlakuan media prekultur PM; B) inokulum dengan perlakuan
media prekultur M3 24
13 Struktur kecambah dari embrio endospermik mangga Arumanis klon 143: A) kotiledonari; B) plumula; C) akar; D) daun 25
DAFTAR LAMPIRAN
1 Komposisi Medium MS (Murashige dan Skoog, 1962) 29 2 Komposisi Medium B5 (Gamborg, Miller, dan Ojima, 1968) 30
PENDAHULUAN
Latar Belakang
Mangga (Mangifera indica L.) merupakan tanaman yang bernilai ekonomi tinggi karena buahnya (Jana et al. 1994). Tanaman mangga berpotensi untuk dikembangkan karena mempunyai tingkat keragaman genetik yang tinggi, sesuai dengan agroklimat Indonesia, buahnya disukai oleh hampir semua lapisan masyarakat dan memiliki pasar yang luas. Tiga varietas mangga unggul Indonesia yang telah dilepas yaitu: Arumanis 143, Golek 31, dan Manalagi 69 (Baswarsiati dan Yuniarti 2007). Mangga Arumanis klon 143 mendominasi bisnis buah mangga di Indonesia dalam lima tahun terakhir (Ihsan dan Sukarmin 2008).
Mangga dianggap sebagai spesies tanaman yang sulit untuk ditangani dalam program pemuliaan karena karakteristik biologi tanaman yaitu: 1. Fase juvenil yang panjang; 2. Tingkat heterozigositas tinggi mengakibatkan hasil hibridisasi yang tak terduga; 3. Hanya ada satu biji per buah; 4. Poliembrioni di beberapa kultivar; dan 5. Luasan tanah yang besar diperlukan untuk penilaian dugaan hibrida (Litz 1997). Karakteristik tersebut menyebabkan pemuliaan konvensional sulit diterapkan pada tanaman mangga. Menurut Ihsan dan Sukarmin (2008), untuk mendapatkan varietas unggul baru mangga diperlukan tahapan yang panjang dan waktu yang cukup lama yaitu 6-20 tahun.
Bioteknologi mengacu pada penerapan biologi molekuler dan genetika sel somatik untuk perbaikan tanaman. Bioteknologi memiliki potensi untuk menyelesaikan beberapa masalah produksi yang paling serius dari kultivar mangga penting dan untuk meningkatkan metodologi pemuliaan. Hal ini akan memungkinkan sasaran dan perubahan sifat agronomi tertentu dalam kultivar yang ada, tanpa mengubah karakter dari klon tersebut. Selain itu, jadwal pemuliaan akan bebas dari batasan masa juvenil dan tahun-tahun tambahan untuk evaluasi pohon. Berbagai pendekatan bioteknologi telah efektif disesuaikan dengan perbaikan genetik tanaman tahunan (Litz 1997). Bioteknologi sebagai pelengkap pemuliaan secara konvensional dan mempercepat program pemuliaan tanaman mangga (Krishna dan Singh 2007).
Salah satu metode bioteknologi tanaman adalah kultur jaringan (kultur in vitro). Teknik in vitro yang digunakan dengan memanfaatkan proses organogenesis maupun embriogenesis. Perbanyakan secara in vitro memanfaatkan kemampuan totipotensi tanaman. Totipotensi merupakan kemampuan sel tanaman secara genetik untuk tumbuh menjadi tanaman yang sempurna bila ditumbuhkan pada kondisi optimum. Menurut Patena et al. (2002), tamanan berkayu seperti mangga sangat sulit untuk dimanipulasi dengan kultur jaringan. Salah satu kesulitannya yaitu mendapatkan bentuk kalus dan menumbuhkan embrio/ planlet atau kalus mudah sekali menjadi coklat.
Embriogenesis adalah proses perkembangan embrio lengkap dari sel somatik, sel endosperma, atau sel zigotik yang diperoleh dari berbagai sumber eksplan. Suatu keuntungan dari embriogenesis somatik adalah embrio-embrio somatik yang dihasilkan bersifat bipolar, yakni memiliki ujung-ujung akar dan pucuk yang diperlukan bagi pertumbuhan tanaman lengkap. Di samping itu,
sel-2
sel yang bersifat embriogenik dapat menghasilkan embrio dalam jumlah besar dalam satu wadah kultur, lebih banyak daripada pucuk-pucuk majemuk yang diregenerasikan secara adventif melalui organogenesis (Zulkarnain 2009). Sari (2012) telah berhasil menginduksi embriogenesis dari sel endosperma mangga Arumanis klon 143. Namun pada penelitian tersebut tidak didapatkan embrio endospermik yang berkembang menjadi planlet.
Perkecambahan embrio merupakan tahap akhir dalam proses embriogenesis. Embrio somatik dewasa yang mengalami perkecambahan diharapkan akan menjadi tanaman lengkap (planlet). Planlet selanjutnya diaklimatisasi sehingga dapat ditanam di lapang dan diketahui hasilnya. Hasil penelitian Jana et al. (1994) sebanyak 55.6% embrio matang dari mangga monoembrionik varietas Alphonso mengalami perkecambahan yang normal, memproduksi tunas dan akar dalam waktu 15 sampai 21 hari pada media yang mengandung N6-benzyladenine (BA) 5 mg/l (22.2 µM). Xiao et al. (2004), melaporkan bahwa persentase embrio somatik mangga monoembrionik varietas Zi-Hua yang menjadi planlet sekitar 30% pada media yang mengandung 5 mg/l (25 µM) kinetin. Penelitian Ali et al. (2010) terhadap perkecambahan embrio somatik kenari menunjukkan, pra-perlakuan suhu rendah (4 °C selama empat minggu) yang dikombinasikan dengan zat pengatur tumbuh tanaman menghasilkan tingkat konversi ke planlet tertinggi (55.0%).
Embriogenesis sangat tergantung pada kultivar mangga (Xiao et al. 2004). Oleh karena itu, perlu dilakukan penelitian untuk mendapatkan media yang sesuai untuk perkecambahan embrio endospermik dari mangga poliembrioni varietas Arumanis klon 143 untuk membentuk planlet.
Tujuan Penelitian
Tujuan dilakukannya penelitian ini adalah:
1. Mempelajari pengaruh perlakuan suhu rendah terhadap kemampuan berkecambah embrio endospermik mangga (M. indica L.) varietas Arumanis klon 143.
2. Mempelajari pengaruh jenis dan konsentrasi media dasar terhadap kemampuan berkecambah embrio endospermik mangga (M. indica L.) varietas Arumanis klon 143.
3. Mempelajari pengaruh konsentrasi giberelin terhadap kemampuan berkecambah embrio endospermik mangga (M. indica L.) varietas Arumanis klon 143.
4. Memperoleh kombinasi media yang optimal untuk perkecambahan embrio endospermik mangga (M. indica L.) varietas Arumanis klon 143.
3
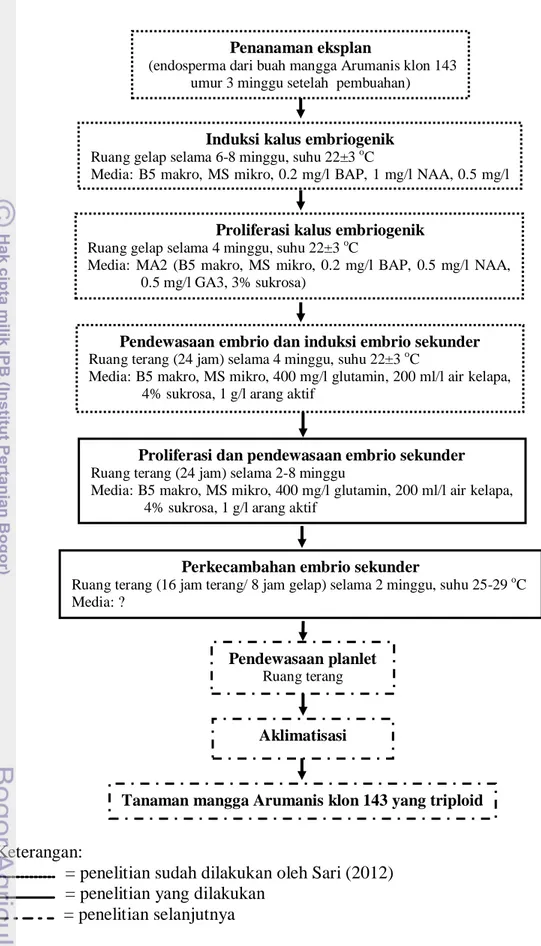
Gambar 1 Bagan alur penelitian embriogenesis sel endosperma untuk perakitan tanaman triploid mangga Arumanis klon 143
Keterangan:
= penelitian sudah dilakukan oleh Sari (2012) = penelitian yang dilakukan
= penelitian selanjutnya
Penanaman eksplan
(endosperma dari buah mangga Arumanis klon 143 umur 3 minggu setelah pembuahan)
Induksi kalus embriogenik
Ruang gelap selama 6-8 minggu, suhu 22±3 oC
Media: B5 makro, MS mikro, 0.2 mg/l BAP, 1 mg/l NAA, 0.5 mg/l GA3, 3% sukrosa
Proliferasi kalus embriogenik
Ruang gelap selama 4 minggu, suhu 22±3 oC
Media: MA2 (B5 makro, MS mikro, 0.2 mg/l BAP, 0.5 mg/l NAA, 0.5 mg/l GA3, 3% sukrosa)
Pendewasaan embrio dan induksi embrio sekunder
Ruang terang (24 jam) selama 4 minggu, suhu 22±3 oC
Media: B5 makro, MS mikro, 400 mg/l glutamin, 200 ml/l air kelapa, 4% sukrosa, 1 g/l arang aktif
Perkecambahan embrio sekunder
Ruang terang (16 jam terang/ 8 jam gelap) selama 2 minggu, suhu 25-29 oC Media: ?
Pendewasaan planlet
Ruang terang
Aklimatisasi
Tanaman mangga Arumanis klon 143 yang triploid Proliferasi dan pendewasaan embrio sekunder
Ruang terang (24 jam) selama 2-8 minggu
Media: B5 makro, MS mikro, 400 mg/l glutamin, 200 ml/l air kelapa, 4% sukrosa, 1 g/l arang aktif
4
TINJAUAN PUSTAKA
Mangga (Mangifera indica L.)
Mangga (Mangifera indica L.) termasuk famili Anacardiaceae, juga dikenal sebagai keluarga mete, dengan sekitar 75 genus dan 700 spesies. Sebagian besar spesies adalah tanaman tropis, dengan beberapa spesies subtropis (Nakasone dan Paull 1998). Mangga merupakan tanaman buah tahunan berupa pohon yang berasal dari negara India. Tanaman ini kemudian menyebar ke wilayah Asia Tenggara termasuk Malaysia dan Indonesia (Iswanto 2007). Jenis yang banyak ditanam di Indonesia yaitu mangga arumanis, golek, gedong, manalagi dan cengkir serta Mangifera foetida yaitu kemang dan kweni (Prihatman2000).
Mangga dapat dikelompokkan menjadi dua berdasarkan kemampuan untuk bereproduksi dengan biji. Kelompok paling besar berbiji monoembrionik. Pembentukan kultivar komersial pada kelompok ini dicapai dengan perbanyakan klonal. Kelompok yang lain terdiri dari poliembrionik. Kelompok ini dapat menghasilkan bibit yang sama dengan induknya dan sejumlah kecil variasi untuk perbanyakan klonal. Sebagian besar kultivar komersial di Indonesia, Filipina, dan Thailand adalah tipe poliembrionik (Berwick 1940). Arumanis sebagai tanaman asli Indonesia memiliki perkecambahan poliembrionik yang memungkinkan mendapatkan bibit mangga dengan sifat sama baiknya dengan induk (Fitmawati et al. 2009).
Arumanis disebut juga sebagai 'Harumanis'. Pohon dari varietas ini kuat dan tinggi dengan kanopi sedikit terbuka. Kulit buah berwarna kuning kehijauan, terdapat titik-titik berwarna kuning terang. Bentuk oval memanjang dengan dasar bulat, panjangnya 11-14 cm dan lebar 4.75-6.5 cm. Bobot buah 200-350 g per buah. Buah varietas ini memiliki kulit tebal, keras dan mudah dipisahkan. Daging buah tebal dan juicy dengan sedikit serat. Warna daging buah kuning lemon, rasa manis agak hambar dengan aroma yang kuat. Benih bersifat poliembrionik. Kultivar ini matang pada musim tertentu dan berulang secara teratur. Perbanyakan secara vegetatif relatif mudah dengan cara grafting. Mangga jenis ini banyak ditanam di daerah dengan kelembaban tinggi di dunia dimana banyak kultivar lain yang gagal untuk berbuah (Knight 1995).
Embriogenesis Sel
Embriogenesis sel somatik adalah proses perkembangan embrio lengkap dari sel-sel somatik yang diperoleh dari berbagai jaringan vegetatif sebagai sumber eksplan. Inisiasi dan diferensiasi embrio somatik tidak melibatkan proses seksual (Zulkarnain 2009). Ada dua macam embrio somatik yang dapat terbentuk yaitu embrio yang terbentuk secara langsung dari sel atau jaringan tanpa melalui pembentukan kalus, dan embrio yang terbentuk secara tidak langsung yaitu melalui tahapan pembentukan kalus (Yuwono 2008). Embriogenesis juga dapat diinduksi dari sel-sel seperti: endosperma, sel zigotik, polen maupun ovul, yang akan menghasilkan embrio dengan ploidi yang berbeda dengan tetuanya seperti
5 embriogenesis sel endosperma yang akan membentuk embrio endospermik dengan ploidi 3n (triploid).
Lingkungan menjadi faktor yang menentukan perkembangan embrio endospermik mangga. Alfaro et al. (1996) menyatakan bahwa suhu antara 22.5 dan 37.5 °C merangsang perkembangan embrio, sedangkan suhu yang lebih rendah (7.5 °C dan 15 °C) menunda perkecambahan embrio mangga. Menurut Litz (1997) embrio endospermik mangga berkembang menjadi dewasa dalam keadaan gelap dan biasanya berkecambah ketika telah sepenuhnya matang. Pemanjangan hipokotil diawali oleh pertumbuhan akar, dan sekitar 2 minggu kemudian muncul tunas.
Keberhasilan perkecambahan menjadi kendala perbanyakan mangga dengan embrio somatik. Menurut Litz (1997), meskipun embrio somatik mangga menunjukkan perubahan tingkat tinggi menjadi planlet, umumnya menurun setelah beberapa kali subkultur. Setelah itu, pemulihan tanaman mengalami masalah, biasanya terjadi dedifferensiasi dari hipokotil dan nekrosis ujung tunas. Berdasarkan hasil penelitian Xiao et al. (2004), persentase konversis embrio somatik mangga monoembrionik varietas Zi-Hua menjadi planlet sekitar 30%. Kultur disimpan di ruang kultur dengan suhu 28±1 oC, fotoperiodisasi 16 jam terang/ 8 jam gelap dan intensitas cahaya 24 µmol m-2 s-1 di bawah lampu flourescen. Planlet tumbuh pesat selama 20 hari pertama pada media konversi, namun lebih lanjut pertumbuhan menjadi lebih lambat pada media yang sama. Fase ini memerlukan optimasi proses konversi lebih lanjut untuk meningkatkan persentase konversi. Waktu yang diperlukan untuk regenerasi planlet sekitar 75-80 hari. Secara spontan pada media konversi terjadi embriogenesis somatik sekunder langsung dari hipokotil embrio primer.
Embrio Endospermik
Embrio endospermik adalah embrio yang dihasilkan dari proses embriogenesis sel endosperma. Sel endosperma memiliki triploid yang terbentuk hasil penggabungan 3 inti sel haploid, satu dari sel gamet jantan dan dua lainnya berasal dari sel gamet betina. Kondisi triploid pada sel endosperma ini merupakan potensi yang dapat digunakan untuk membuat tanaman triploid melalui kultur jaringan (Hanayanti 2011).
Hanayanti (2011) melakukan embriogenesis sel endosperma mangga Gedong Gincu untuk merakit tanaman triploid. Eksplan terbaik adalah endosperma dari buah yang berumur 3 minggu setelah pembentukan buah. Media induksi kalus sel embriosperma adalah media BGC3 (glutamin, BAP dan 2,4-D) dengan waktu pembentukan kalus 2-4 MST. Media perkembangan embriogenesis dan proliferasi terbaik adalah media MGC3 (BAP, NAA dan GA3) dengan perkembangan kalus sel endosperma berhasil mencapai tahap kotiledonari pada 7 MST dan frekuensi proliferasi paling tinggi yaitu 2.75 kali dalam waktu 6 minggu. Perkecambahan embrio berjalan sangat lambat. Pada media P3 dengan
kombinasi glutamin dan air kelapa berhasil membentuk tunas pada 48 MST dengan persentase berkecambah kurang dari 5%. Uji sitologi menunjukkan jumlah embrio zigotik ±40-43/sel dan jumlah kromosom sel endosperma ±62-72/sel.
6
Induksi embriogenesis in vitro sel endosperma mangga Arumanis klon 143 telah dilakukan oleh Sari (2012). Hasil penelitian menunjukkan bahwa induksi kalus dari sel endosperma mangga yang terbaik terjadi pada eksplan dari buah yang berumur 3 minggu. Media MA3 (B5 makro + MS mikro + 0.2 mg/l BAP + 1 mg/l NAA + 0.5 mg/l GA3 + 3% sukrosa) merupakan media yang optimal untuk menginduksi kalus embriogenik dari sel endosperma dan untuk perkecambahan embrio somatik.
Media Kultur dan Zat Pengatur Tumbuh
Medium yang digunakan untuk kultur in vitro tanaman dapat berupa media padat atau cair. Medium padat digunakan untuk menghasilkan kalus yang selanjutnya diinduksi membentuk tanaman yang lengkap (disebut planlet), sedangkan medium cair biasanya digunakan untuk kultur sel. Medium yang digunakan mengandung lima komponen utama, yaitu senyawa anorganik, sumber karbon, vitamin, zat pengatur tumbuh dan suplemen organik (Yuwono 2008).
Menurut Zulkarnain (2008), kebutuhan nutrisi untuk pertumbuhan kultur in vitro yang optimal bervariasi antar spesies tanaman ataupun antar varietas. Bahkan, jaringan yang berasal dari bagian tanaman yang berbeda akan berbeda pula kebutuhan nutrisinya. Oleh karena itu, tidak ada satupun medium dasar yang berlaku universal untuk semua jenis jaringan dan organ.
Pilihan media tertentu terutama tergantung pada spesies tanaman, jaringan atau organ untuk dibudidayakan, dan tujuan percobaan. Jika bahan tanaman telah berhasil dibudidayakan di laboratorium lain, lebih baik untuk memulai dengan metode yang telah diterbitkan. Sebuah titik awal yang cocok untuk inisiasi kalus eksplan dari jaringan dikotil adalah persiapan media dasar MS. Salah satu karakteristik dari media ini adalah konsentrasi ion nitrat, kalium dan amonium yang relatif tinggi dibandingkan media nutrisi lain (Dodds dan Roberts 1985).
Pada penelitian kultur jaringan mangga yang dilakukan oleh Ermayanti et al. (2001) biji yang mengalami kontaminasi dapat berkecambah pada media MS atau B5. Hanayanti (2010) mengecambahkan embrio fase kotiledonari dari eksplan embrio zigotik, nuselus dan endosperma mangga gedong gincu pada 6 jenis media perkecambahan yang terdiri atas media dasar hara makro dari B5 (Gamborg) dan hara mikro juga vitamin dari MS dengan komposisi tambahan (glutamin, BAP, NAA, 2,4-D, air kelapa, dan sukrosa) yang berbeda-beda. Embrio fase kotiledonari berhasil menginisiasi tunas pada media P3 (glutamin + air kelapa + 4% sukrosa), P4 (glutamin + air kelapa + 2% sukrosa), dan P6 (glutamin + 2% sukrosa). Namun pertumbuhan tunas hanya terdapat pada media P3. Pada media P3 embrio fase kotiledonari menunjukkan pertumbuhan tunas dengan bentuk tanaman rosete dan daun yang sangat tebal.
Jana et al. (1994) melakukan perkecambahan embrio somatik dewasa dari mangga monoembrionik varietas Alphonso menggunakan setengah media dasar MS ditambah dengan berbagai tingkat BA (1.0, 3.0, 5.0, dan 10.0 mg/1), sukrosa 20 g/l, 4.2 g/l agar-agar, dan 2.5 g/l arang aktif. Sebanyak 55.6% embrio menjadi planlet diperoleh ketika benzyladenine (BA) sebanyak 5 mg/l ditambahkan ke medium. Menurut Rout et al. (1995), pengenceran media pada tahap perkecambahan dimaksudkan untuk menghindari pengkalusan kembali pada dasar
7 tunas atau struktur embrio somatik. Berdasarkan penelitian Sukmadjaja (2005), perkecambahan embrio somatik cendana pada media MS yang konsentrasi makronya diencerkan sampai setengahnya lebih baik dibandingkan media MS konsentrasi penuh.
Perubahan embrio somatik fase kotiledonari dari mangga monoembrionik varietas Zi-Hua menjadi planlet dilakukan dengan menempatkan embrio endospermik dewasa fase kotiledonari pada media konversi (CM). Media CM memiliki komposisi elemen makro B5 yang dimodifikasi (DeWald et al. 1989), hara mikro dan organik MS (100 mg/l myo-inositol, 2 mg/l glisin, 0.4 mg/l tiamin-HCl, 0.5 mg/l asam nikotinat, 0.5 mg/l pyridoxine-HCl), 5 mg/l (23 µM) kinetin, 500 mg/l asam amino glutamin, 100 mg/l CH, 20% air kelapa, 1 g/l arang aktif, 6.5 g/l agar-agar, dan 40 g/l sukrosa. Perubahan embrio somatik mangga monoembrionik varietas Zi-Hua menjadi planlet sebanyak 30% dicapai setelah 4 minggu kultur pada medium perkecambahan (Xiao et al. 2004).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilakukan pada bulan April 2012 sampai bulan Agustus 2013 di Laboratorium Kultur Jaringan Tanaman II, Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor.
Alat dan Bahan
Bahan yang digunakan sebagai bahan tanaman dalam penelitian ini adalah embrio somatic dari embriogenesis sel endosperma dari mangga (M. indica L.) varietas Arumanis klon 143. Bahan lainnya antara lain media dasar dari MS dan B5, agar-agar sebagai pemadat, sukrosa, zat pengatur tumbuh tanaman (BA, NAA, CAP, IBA, 2,4-D, kinetin, GA3), asam amino glutamin, arang aktif, air kelapa, dan aquades. Bahan sterilan antara lain clorox, alkohol 70% dan air steril.
Alat yang digunakan terdiri dari peralatan gelas (botol kultur ukuran 200 ml, labu ukur, pipet, gelas ukur, corong gelas, dan cawan petri), timbangan digital, pH meter, magnetic stirer, kompor, autoclave, laminar air flow cabinet, peralatan diseksi (pinset, gunting, dan scalpel), bunsen, sprayer, plastik, karet gelang, rak kultur, lemari es, neraca digital, mikroskop, dan kamera.
Rancangan Penelitian
Penelitian ini terdiri dari 4 percobaan terpisah yaitu: I. Inisiasi perkecambahan embrio endospermik mangga varietas Arumanis klon 143 dengan perlakuan suhu rendah, II. Perkecambahan embrio endospermik mangga varietas Arumanis klon 143 dengan perlakuan giberelin, III. Perkecambahan embrio endospermik mangga varietas Arumanis klon 143 dengan media cair dan padat,
8
dan IV. Perkecambahan embrio endospermik mangga varietas Arumanis klon 143 dengan perlakuan media prekultur yang berbeda. Masing-masing percobaan dijelaskan sebagai berikut:
Percobaan I. Perkecambahan Embrio Endospermik Mangga Varietas Arumanis Klon 143 dengan Perlakuan Suhu Rendah
Inokulum yang digunakan adalah embrio endospermik fase kotiledonari (cotiledonary stage) dari mangga Arumanis klon 143 yang sebelumnya dikulturkan pada media P3 (hara makro B5, hara mikro dan vitamin MS, 100 mg/l myo-inositol, 200 ml/l air kelapa, 1 g/l arang aktif, 400 mg/l glutamin, 40 g/l sukrosa, dan 7 g/l agar-agar) selama 6 minggu. Embrio diberi perlakuan suhu rendah pada suhu 8 oC. Perlakuan dilakukan dengan memindahkan embrio endospermik fase kotiledonari dari dalam botol kultur ke cawan petri beralaskan tissu yang telah disterilkan. Cawan petri yang berisi embrio endospermik kemudian disimpan dalam lemari es bersuhu 8 oC dengan waktu penyimpanan selama 4, 6, 8, dan 10 hari. Setelah diberi perlakuan suhu rendah, embrio ditanam pada media 1/2MS-AK. Komposisi media 1/2MS-AK terdiri dari media dasar MS (1/2 konsentrasi hara makro dan mikro, konsentrasi penuh vitamin dan myo-inositol) ditambah dengan 50 ml/l air kelapa, 20 g/l gula, 1 g/l arang aktif dan 7 g/l agar-agar. Derajad keasaman (pH) diatur dengan penambahan KOH 1N atau HCl 1N hingga mencapai 5.7 sebelum diautoklaf. Kultur selanjutnya disimpan pada ruang bersuhu 27-31 oC. Kultur disusun pada rak kultur dan diberi pencahayaan dari lampu fluoresen 18 Watt yang dipasang dengan jarak 33 cm dari rak kultur atau 22.5 cm di atas botol kultur dengan fotoperiodisasi 24 jam terang.
Percobaan ini disusun dengan menggunakan rancangan acak lengkap (RAL). Rancangan perlakuan disusun dengan satu faktor perlakuan yaitu waktu perlakuan suhu rendah. Perlakuan suhu rendah terdiri dari 4 taraf yaitu 4, 6, 8, dan 10 hari. Perlakuan diulang sebanyak 6 kali, sehingga terdapat 24 satuan percobaan. Setiap satuan percobaan terdiri dari 5 embrio endospermik fase kotiledonari. Setiap embrio menjadi satuan amatan.
Pengamatan dilakukan selama 14 minggu terhadap persentase embrio aseptik, persentase embrio yang berproliferasi, bobot dan diameter proembrio yang dihasilkan, jumlah embrio endopsermik sekunder yang dihasilkan, serta persentase embrio yang berkecambah. Data hasil pengamatan dianalisis dengan menggunakan uji F. Apabila terdapat pengaruh yang nyata, maka akan dilanjutkan dengan uji lanjut DMRT (Duncan Multiple Range Test) pada taraf kepercayaan 5%. Pengolahan data dilakukan dengan program Statistical Analysis System (SAS). Percobaan II. Perkecambahan Embrio Endospermik Mangga Varietas
Arumanis Klon 143 dengan Perlakuan Giberelin
Inokulum yang digunakan adalah embrio endospermik fase kotiledonari dari mangga Arumanis klon 143 yang sebelumnya dikulturkan pada media P3 selama 9 minggu. Embrio endospermik fase kotiledonari ditanam pada 4 jenis media perkecambahan yaitu MS0, MS1/2, MS-5G, dan MS-10G. Berikut komposisi masing-masing media:
MS0 = MS konsentrasi penuh + 30 g/l gula + 5 g/l agar-agar + 1 g/l arang aktif MS1/2 = MS (1/2 konsentrasi hara makro dan mikro, konsentrasi penuh bahan
9 MS-5G = MS + 5 µM (1.732 mg/l) GA3 + 30 g/l gula + 5 g/l agar-agar + 1 g/l
arang aktif
MS-10G = MS + 10 µM (3.463 mg/l) GA3 + 30 g/l gula + 5 g/l agar-agar + 1 g/l arang aktif
Derajat keasaman (pH) media diatur hingga mencapai 5.7 sebelum diautoklaf. Media disterilkan dengan autoclave pada suhu 121 oC dan tekanan 17.5 Psi selama 20 menit Embrio endospermik fase kotiledonari yang telah ditanam pada media perlakuan disimpan pada ruang bersuhu 27-31 oC. Kultur disusun pada rak kultur dan diberi pencahayaan dari lampu fluoresen 18 Watt yang dipasang dengan jarak 33 cm dari rak kultur atau 22.5 cm di atas botol kultur dengan fotoperiodisasi 24 jam terang.
Percobaan ini disusun menggunakan rancangan kelompok lengkap teracak (RKLT) dengan satu faktor perlakuan yaitu jenis media. Terdapat 4 taraf media yaitu MS0, MS1/2, MS-5G, dan MS-10G. Perlakuan diulang sebanyak 3 kali, sehingga terdapat 12 satuan percobaan. Setiap satuan percobaan terdiri dari 9 embrio endospermik fase kotiledonari. Setiap embrio menjadi satuan amatan.
Pengamatan dilakukan selama 14 minggu terhadap persentase embrio aseptik, persentase embrio yang berproliferasi, bobot dan diameter proembrio yang dihasilkan, jumlah embrio endopsermik sekunder yang dihasilkan, serta persentase embrio yang berkecambah. Data hasil pengamatan dianalisis dengan menggunakan uji F. Apabila terdapat pengaruh yang nyata, maka akan dilanjutkan dengan uji lanjut DMRT (Duncan Multiple Range Test) pada taraf kepercayaan 5%. Pengolahan data dilakukan dengan program Statistical Analysis System (SAS). Percobaan III. Perkecambahan Embrio Endospermik Mangga Varietas
Arumanis Klon 143 pada Media Cair dan Padat
Inokulum yang digunakan adalah embrio endospermik fase kotiledonari dari mangga Arumanis klon 143 yang sebelumnya dikulturkan pada media P3 selama 8 minggu. Media perlakuan terdiri dari 2 jenis media yaitu media padat dan media cair dengan komposisi yang sama. Komposisi media terdiri dari media dasar MS ditambah 5 mg/l BA, 200 ml/l air kelapa, 2.5 g/l arang aktif, dan 20 g/l gula. Derajat keasamannya diatur hingga mencapai pH 5.7 sebelum diautoklaf. Media padat dibuat dengan menambahkan 7 g/l agar-agar lalu dimasak. Media cair tanpa agar-agar kemudian dituang sebanyak 20 ml/botol ke dalam botol yang ditambahkan kapas pada dasarnya. Penambahan kapas pada dasar media cair dimaksudkan agar embrio yang ditanam nantinya tidak terendam. Kedua jenis media kemudian disterilkan. Inokulum yang telah ditanam pada media perlakuan kemudian disimpan pada ruang bersuhu 25-28 oC. Botol kultur disusun pada rak dan diberi pencahayaan dari lampu fluoresen 18 Watt yang dipasang dengan jarak 33 cm dari rak kultur atau 22.5 cm di atas botol kultur dengan fotoperiodisasi 16 jam terang/18 jam gelap selama 4 minggu.
Percobaan ini disusun dengan menggunakan rancangan acak lengkap (RAL). Rancangan perlakuan disusun dengan satu faktor perlakuan yaitu jenis media yang terdiri dari dua taraf yaitu media padat dan media cair. Perlakuan diulang sebanyak 3 kali sehingga terdapat 6 satuan percobaan. Setiap satuan percobaan terdiri dari 5 embrio endospermik fase kotiledonari. Setiap embrio menjadi satuan amatan.
10
Pengamatan dilakukan selama 10 minggu terhadap inokulum steril, inokulum membentuk kalus proembrio, inokulum berkecambah, dan jumlah embrio endospermik sekunder yang dihasilkan. Data hasil pengamatan dianalisis dengan menggunakan uji F. Apabila terdapat pengaruh yang nyata, maka akan dilanjutkan dengan uji lanjut DMRT (Duncan Multiple Range Test) pada taraf kepercayaan 5%. Pengolahan data dilakukan dengan program Statistical Analysis System (SAS).
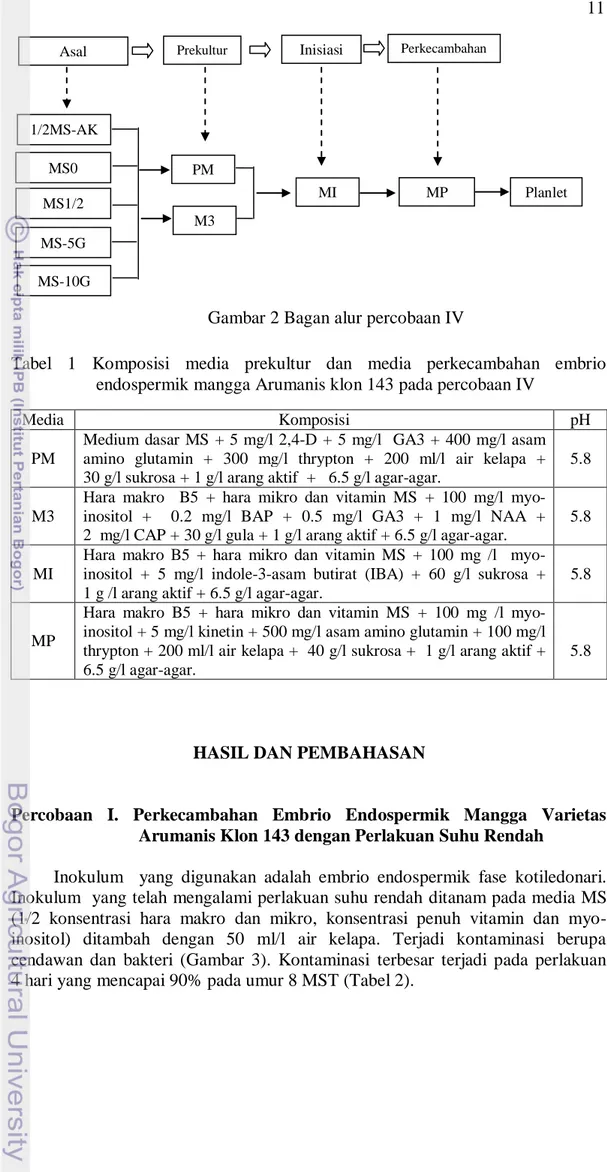
Percobaan IV. Perkecambahan Embrio Endospermik Mangga Varietas Arumanis Klon 143 dengan Perlakuan Prekultur pada Media yang Berbeda
Metode percobaan ini merupakan modifikasi dari penelitian yang dilakukan oleh Xiao et al. (2004) pada mangga monoembrionik varietas Zi-Hua. Percobaan ini terdiri dari 3 tahap yaitu prekultur, inisiasi embrio endospermik, dan perkecambahan embrio endospermik untuk mendapatkan planlet. Setiap tahap percobaan menggunakan media dengan komposisi yang berbeda-beda (Tabel 1).
Inokulum yang digunakan pada percobaan ini berupa poliembrio yang dihasilkan oleh embrio endospermik fase kotiledonari pada percobaan I dan II. Inokulum dibedakan berdasarkan media asalnya yaitu 1/2MS-AK, MS0, MS1/2, MS-5G, dan MS-10G. Inokulum ditanam pada dua jenis media prekultur yang berbeda (PM dan M3) di ruang gelap bersuhu 20 oC selama 1 minggu. Setelah itu inokulum dipindah tanam ke media inisiasi embrio endospermik (MI) di ruang gelap bersuhu 20-27 oC selama 2 minggu. Setelah 2 minggu di media inisiasi, inokulum selanjutnya ditanam pada media perkecambahan (MP) di ruang terang bersuhu 25-29 oC. Kultur disusun pada rak kultur dan diberi pencahayaan dari lampu fluoresen 18 Watt yang dipasang dengan jarak 33 cm dari rak kultur atau 22.5 cm di atas botol kultur dengan fotoperiodisasi 16 jam terang/18 jam gelap selama 9 minggu (Gambar 2).
Percobaan ini disusun dengan menggunakan rancangan acak lengkap (RAL). Rancangan perlakuan disusun secara faktorial dengan 2 faktor perlakuan yaitu media asal inokulum dan media prekultur. Faktor pertama asal media inokulum terdiri dari 5 taraf yaitu media 1/2MS-AK, MS0, MS1/2, MS-5G dan MS-10G. Faktor kedua media prekultur terdiri dari 2 taraf yaitu media PM dan M3. Perlakuan diulang sebanyak 5 kali sehingga terdapat 50 satuan percobaan. Setiap satuan percobaan terdiri dari 5 inokulum proembrio dengan ukuran diameter 0.3-0.5 mm dan bobot ±4 mg. Setiap inokulum menjadi satuan amatan.
Pengamatan dilakukan selama 13 minggu terhadap inokulum steril, jumlah embrio endospermik yang dihasilkan, dan jumlah embrio yang dihasilkan. Data hasil pengamatan dianalisis dengan menggunakan uji F. Apabila terdapat pengaruh yang nyata, maka akan dilanjutkan dengan uji lanjut DMRT (Duncan Multiple Range Test) pada taraf kepercayaan 5%. Pengolahan data dilakukan dengan program Statistical Analysis System (SAS).
11
Gambar 2 Bagan alur percobaan IV
Tabel 1 Komposisi media prekultur dan media perkecambahan embrio endospermik mangga Arumanis klon 143 pada percobaan IV
Media Komposisi pH
PM
Medium dasar MS + 5 mg/l 2,4-D + 5 mg/l GA3 + 400 mg/l asam amino glutamin + 300 mg/l thrypton + 200 ml/l air kelapa + 30 g/l sukrosa + 1 g/l arang aktif + 6.5 g/l agar-agar.
5.8
M3
Hara makro B5 + hara mikro dan vitamin MS + 100 mg/l
myo-inositol + 0.2 mg/l BAP + 0.5 mg/l GA3 + 1 mg/l NAA + 2 mg/l CAP + 30 g/l gula + 1 g/l arang aktif + 6.5 g/l agar-agar.
5.8
MI
Hara makro B5 + hara mikro dan vitamin MS + 100 mg /l myo- inositol + 5 mg/l indole-3-asam butirat (IBA) + 60 g/l sukrosa + 1 g /l arang aktif + 6.5 g/l agar-agar.
5.8
MP
Hara makro B5 + hara mikro dan vitamin MS + 100 mg /l myo-inositol + 5 mg/l kinetin + 500 mg/l asam amino glutamin + 100 mg/l thrypton + 200 ml/l air kelapa + 40 g/l sukrosa + 1 g/l arang aktif + 6.5 g/l agar-agar.
5.8
HASIL DAN PEMBAHASAN
Percobaan I. Perkecambahan Embrio Endospermik Mangga Varietas Arumanis Klon 143 dengan Perlakuan Suhu Rendah
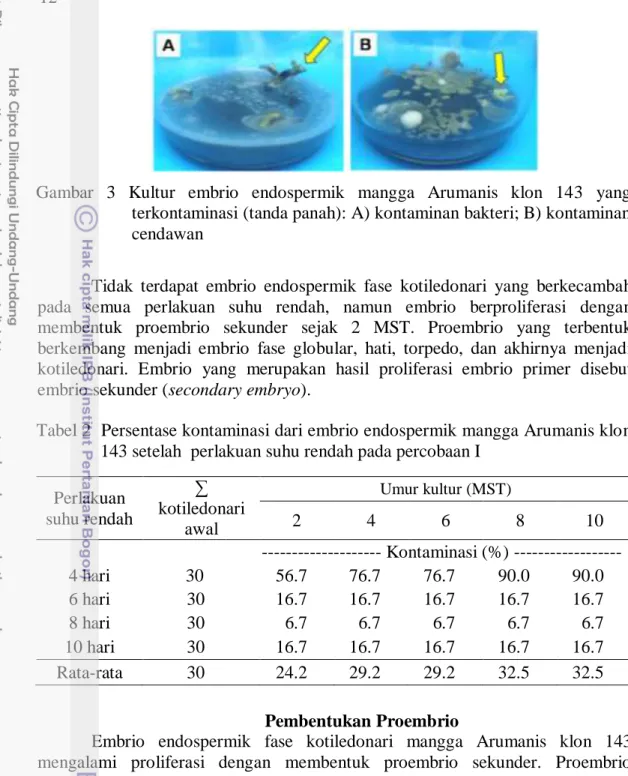
Inokulum yang digunakan adalah embrio endospermik fase kotiledonari. Inokulum yang telah mengalami perlakuan suhu rendah ditanam pada media MS (1/2 konsentrasi hara makro dan mikro, konsentrasi penuh vitamin dan myo-inositol) ditambah dengan 50 ml/l air kelapa. Terjadi kontaminasi berupa cendawan dan bakteri (Gambar 3). Kontaminasi terbesar terjadi pada perlakuan 4 hari yang mencapai 90% pada umur 8 MST (Tabel 2).
Asal inokulum
Prekultur Inisiasi Perkecambahan
nn PM 1/2MS-AK MS0 MS1/2 MS-5G MS-10G M3 MP MI Planlet
12
Gambar 3 Kultur embrio endospermik mangga Arumanis klon 143 yang terkontaminasi (tanda panah): A) kontaminan bakteri; B) kontaminan cendawan
Tabel 2 Persentase kontaminasi dari embrio endospermik mangga Arumanis klon 143 setelah perlakuan suhu rendah pada percobaan I
Perlakuan suhu rendah ∑ kotiledonari awal Umur kultur (MST) 2 4 6 8 10 --- Kontaminasi (%) --- 4 hari 30 56.7 76.7 76.7 90.0 90.0 6 hari 30 16.7 16.7 16.7 16.7 16.7 8 hari 30 6.7 6.7 6.7 6.7 6.7 10 hari 30 16.7 16.7 16.7 16.7 16.7 Rata-rata 30 24.2 29.2 29.2 32.5 32.5
Tidak terdapat embrio endospermik fase kotiledonari yang berkecambah pada semua perlakuan suhu rendah, namun embrio berproliferasi dengan membentuk proembrio sekunder sejak 2 MST. Proembrio yang terbentuk berkembang menjadi embrio fase globular, hati, torpedo, dan akhirnya menjadi kotiledonari. Embrio yang merupakan hasil proliferasi embrio primer disebut embrio sekunder (secondary embryo).
Pembentukan Proembrio
Embrio endospermik fase kotiledonari mangga Arumanis klon 143 mengalami proliferasi dengan membentuk proembrio sekunder. Proembrio sekunder terbentuk pada semua perlakuan suhu rendah. Air kelapa dengan konsentrasi 5% yang ditambahkan ke dalam media diduga menjadi faktor pemicu tumbuhnya proembrio. Air kelapa bermanfaat untuk menginduksi pertumbuhan kalus dan kultur suspensi serta untuk menginduksi morfogenesis (Thorpe et al. 2008). Air kelapa mengandung asam amino, asam organik, asam nukleat, beberapa vitamin, gula dan gula alkohol, hormon tanaman (auksi, sitokinin), mineral, dan zat lain yang tidak teridentifikasi, dimana zat tersebut secara keseluruhan bertanggung jawab terhadap kualitas pertumbuhan (Molnar et al. 2011). Proembrio sekunder terbentuk saat eksplan berumur 2 minggu setelah tanam (MST). Proembrio sekunder mulai terbentuk di bagian hipokotil (Gambar 4A). Proembrio yang terbentuk bersifat remah (friable) dan berwarna kuning (Gambar 4B).
13
Gambar 4 A) Embrio endospermik mangga Arumanis klon 143 yang berproliferasi setelah perlakuan suhu rendah pada 10 MST; B) Proembrio yang dihasilkan embrio endospermik dengan perlakuan suhu rendah;C) Embrio endospermik sekunder pada 12 MST
Tabel 3 Persentase embrio endospermik mangga Arumanis klon 143 yang berproliferasi setelah perlakuan suhu rendahpada percobaan I
Perlakuan suhu rendah Umur kultur (MST) 1 2 3 4 5 6 7 8 9 10 --- Embrio berproliferasi (%)a, b--- 4 hari 0.0 5.0 16.7 58.3ab 58.3 66.7 50.0 75.0 75.0 75.0 6 hari 0.0 4.0 20.0 28.0b 36.0 40.0 44.0 44.0 52.0 52.0 8 hari 0.0 3.3 35.6 64.5a 64.5 67.8 73.3 76.7 76.7 83.3 10 hari 0.0 12.0 16.0 28.0ab 32.0 48.0 56.0 64.0 64.0 64.0 Pengaruh tn tn tn * tn tn tn tn tn tn KK (%) 0.0 97.4 65.5 34.0 37.6 37.0 51.3 28.6 27.8 26.0 Keterangan: aData ditransformasi (x+0.5)1/2 sebelum diolah menggunakan SAS dan data yang
ditampilkan adalah data sebelum transformasi; bAngka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji kepercayaan 5 % (uji selang berganda Duncan); KK: Koefisien Keragaman; tn: tidak nyata; *) berpengaruh nyata p≤0.05
Perlakuan suhu rendah secara umum tidak berpengaruh nyata terhadap persentase embrio berproliferasi. Perlakuan suhu rendah hanya berpengaruh nyata terhadap proliferasi embrio pada 4 MST (Tabel 3). Pada 4 MST, perlakuan suhu rendah 8 hari memiliki persentase embrio berproliferasi tertinggi sebesar 64.5%, namun tidak berbeda nyata dengan perlakuan suhu rendah 4 dan 10 hari yang masing-masing 58.3% dan 28%. Embrio berproliferasi pada perlakuan suhu rendah 4, 6, dan 10 hari tidak berbeda nyata.
Persentase embrio berproliferasi cenderung meningkat setiap minggunya. Penurunaan persentase embrio berproliferasi hanya terjadi pada perlakuan suhu rendah 4 hari dari 6 MST ke 7 MST. Hal ini terjadi karena pada waktu tersebut terjadi kontaminasi pada perlakuan suhu rendah 4 hari. Pada akhir pengamatan (10 MST), persentase embrio berproliferasi tertinggi sebesar 83.3% pada perlakuan suhu rendah 8 hari yang tidak berbeda nyata dengan perlakuan suhu rendah 4 hari (75%), 6 hari (52%), dan 10 hari (64%).
14
Tabel 5 Jumlah embrio endospermik sekunder yang terbentuk dari embrio endospermik mangga Arumanis klon 143 dengan perlakuan suhu rendah umur 14 MST
Perlakuan suhu rendah
Jumlah inokulum Total embrio sekunder Awal Akhir (aseptik) Fase torpedo Fase kotiledonari Jumlah embrio total 4 hari 30 1 0 0 0 6 hari 30 12 24 8 32 8 hari 30 12 18 15 33 10 hari 30 9 16 5 21
Tabel 4 Bobot dan diameter rata-rata proembrio yang dihasilkan embrio endospermik mangga Arumanis klon 143 dengan perlakuan suhu rendah pada umur 10 MST
Perlakuan suhu rendah ∑ Inokulum awal ∑ Inokulum aseptik Bobot proembrio (mg) Diameter proembrio (mm) 4 hari 30 3 666.80 11.75 6 hari 30 25 48.40 3.70 8 hari 30 28 176.70 8.07 10 hari 30 25 109.00 6.68
Proembrio yang terbentuk pada perlakuan suhu rendah 4 hari memiliki bobot rata-rata terbesar yaitu 666.80 mg. Bobot rata-rata proembrio terkecil terdapat pada perlakuan suhu rendah 6 hari yaitu 48.40 mg. Diameter rata-rata proembrio terbesar terdapat pada perlakuan suhu rendah 4 hari yaitu 11.75 mm (Tabel 4).
Pembentukan Embrio Endospermik Sekunder dan Embrio Berkecambah Setelah dilakukan pengukuran bobot dan diameter proembrio, kemudian dilakukan subkultur terhadap embrio beserta proembrio yang terbentuk pada media baru dengan komposisi yang sama seperti media sebelumnya. Pada minggu ke-14 (dihitung sejak embrio ditanam), beberapa proembrio membentuk embrio endospermik sekunder fase torpedo dan kotiledonari (Gambar 4C). Embrio sekunder terbentuk pada proembrio perlakuan suhu rendah 6, 8, dan 10 hari. Tidak ada embrio sekunder yang terbentuk pada perlakuan suhu rendah selama 4 hari. Hal ini diduga bukan karena ketidakmampuan proembrio beregenerasi tetapi karena banyaknya eksplan yang terkena kontaminasi sehingga peluang munculnya embrio sekunder pada perlakuan 4 hari menjadi lebih kecil dibandingkan perlakuan lainnya. Embrio sekunder fase torpedo terbanyak terdapat pada perlakuan suhu rendah selama 6 hari yaitu 24 embrio. Embrio sekunder fase kotiledonari terbanyak terdapat pada perlakuan suhu rendah selama 8 hari yaitu 15 embrio. Jumlah embrio sekunder total terbanyak dihasilkan oleh embrio dengan perlakuan suhu rendah 8 hari yaitu 33 embrio (Tabel 5).
15
Tabel 6 Persentase kontaminasi embrio endospermik mangga Arumanis klon 143 pada perlakuan GA3
Jenis Media ∑ Kotiledonari awal Umur kultur (MST) 2 4 6 8 10 --- Kontaminasi (%) --- MS0 27 22.2 40.7 44.4 51.9 51.9 MS1/2 27 29.6 37.0 37.0 37.0 37.0 MS-5G 27 25.9 33.3 51.9 51.9 51.9 MS-10G 27 29.6 33.3 44.4 51.9 51.9 Rata-rata 27 26.9 36.1 44.4 48.2 48.2
Keterangan: MS0: MS konsentrasi penuh tanpa GA3; MS1/2: MS dengan 1/2 konsentrasi hara makro dan mikro, serta konsentrasi penuh vitamin dan myo-inositol, tanpa GA3; MS-5G: MS konsentrasi penuh ditambah 5 µM GA3; MS-10G: MS konsentrasi penuh ditambah 10 µM GA3
Berdasarkan hasil percobaan I, embrio endospermik yang diberikan perlakuan suhu rendah 8 oC (4, 6, 8, dan 10 hari) tidak mengalami perkecambahan setelah ditanam selama 14 MST pada media setengah MS dengan 5% air kelapa. Hal ini diduga karena suhu perlakuan yang kurang rendah dan waktu perlakuan yang kurang lama sehingga perlakuan tidak mampu mematahkan dormansi embrio. Penelitian Ali et al. (2010) terhadap perkecambahan embrio somatik kenari menunjukkan, pra-perlakuan suhu rendah (4 °C selama empat minggu) yang dikombinasikan dengan zat pengatur tumbuh tanaman menghasilkan tingkat konversi ke planlet tertinggi (55.0%).
Percobaan II. Perkecambahan Embrio Endospermik Mangga Varietas Arumanis Klon 143 dengan Perlakuan Giberelin
Pada percobaan ini, embrio endospermik mangga Arumnis klon 143 dikecambahkan pada beberapa media yang berbeda yaitu media konsentrasi penuh dan setengah konsentrasi (MS0 dan MS1/2), serta media konsentrasi penuh dengan penambahan 5 dan 10 µM GA3 (MS-5G dan MS-10G). Pada 3 MST, inokulum mulai berproliferasi dengan membentuk proembrio di semua media perlakuan. Proembrio yang dihasilkan mengalami proliferasi namun hingga 14 MST tidak ada proembrio yang berkembang menjadi embrio sekunder. Kontaminasi cendawan dan bakteri menyerang inokulum yang ditanam pada media MS0, MS-5G dan MS-10G sebanyak 51.9% dan pada media MS1/2 sebanyak 37.0% (Tabel 6).
Pembentukan Proembrio
Embrio endospermik fase kotiledonari mengalami proliferasi. Proliferasi ditandai dengan terbentuknya proembrio sekunder pada bagian hopokotil embrio (Gambar 5). Proembrio yang dihasilkan berwarna kuning dan bersifat remah (friable)
16
Gambar 5 Proliferasi embrio endospermik mangga Arumanis klon 143 umur 10 MST: A) pada media MS0; B) pada media MS1/2; C) pada media MS-5G; D) pada media MS-10G
Proembrio mulai terbentuk saat 2 MST pada media MS1/2 dan MS-10G. Pada media MS dan MS-5G proembrio terbentuk saat 3 MST. Hasil uji statistik menunjukkan konsentrasi media dasar dan GA3 berpengaruh sangat nyata terhadap persentase embrio berproliferasi pada 5 , 6, dan 7 MST (Tabel 7). Tabel 7 Persentase embrio endospermik mangga Arumanis klon 143 yang
berproliferasi pada perlakuan GA3 Jenis media Umur kultur (MST) 1 2a 3a 4 5 6 7 8 9 10 --- Inokulum berproliferasi (%)b--- MS0 0.0 0.0 23.2 40.7 44.5c 47.2b 47.2b 66.7 66.7 66.7 MS1/2 0.0 5.6 25.0 25.0 36.1c 40.3b 40.3b 40.3 40.3 40.3 MS-5G 0.0 0.0 28.1 33.7 62.2b 83.3a 83.3a 83.3 83.3 83.3 MS-10G 0.0 7.4 29.6 61.1 72.2a 77.8a 77.8a 69.5 69.5 75.0 Pengaruh tn tn tn tn ** ** ** tn tn tn KK (%) 0.0 97.9 42.7 30.1 8.0 13.2 13.2 32.7 32.7 31.0 Keterangan: aData pada minggu ini ditransformasi (x+1)1/2 sebelum diolah menggunakan SAS dan
data yang ditampilkan adalah data sebelum transformasi; bAngka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji kepercayaan 5 % (uji selang berganda Duncan); MS0: MS konsentrasi penuh tanpa GA3; MS1/2: MS dengan 1/2 konsentrasi hara makro dan mikro, serta konsentrasi penuh vitamin dan myo-inositol, tanpa GA3; MS-5G: MS konsentrasi penuh ditambah 5 µM GA3; MS-10G: MS konsentrasi penuh ditambah 10 µM GA3; KK: Koefisien Keragaman: tn: tidak nyata; **) berpengaruh sangat nyata p≤0.01
Media dasar MS yang ditambahkan GA3 dengan konsentrasi 5 dan 10 µM (MS-5G dan MS-10G) cenderung memiliki persentase embrio berproliferasi yang lebih besar dibandingkan media MS tanpa GA3 (MS dan MS1/2). Media dengan 10 µM memiliki persentase embrio berproliferasi yang lebih besar dibandingkan media dengan 5 µM hingga 5 MST. Pada 6 hingga 10 MST, persentase embrio berproliferasi pada media dengan 10 µM menjadi lebih rendah dibandingkan media dengan 5 µM. Pada media tanpa GA3, media dengan konsentrasi setengah hara makro dan mikro (MS1/2) memiliki persentase embrio berproliferasi yang lebih kecil dibandingkan media dengan konsentrasi penuh (MS0). Hal ini sesuai dengan penelitian Rout et al. (1995) yang menyatakan bahwa pengenceran media pada tahap perkecambahan dimaksudkan untuk menghindari pengkalusan kembali
17 pada dasar tunas atau struktur embrio somatik.
Pada 5 MST, persentase embrio berproliferasi terbesar terdapat pada embrio yang ditanam pada media MS dengan 10 µM GA3 yaitu sebesar 72.2% berbeda nyata dengan ketiga media lainnya. Persentase embrio berproliferasi terbesar terdapat pada embrio yang ditanam pada media MS dengan setengah konsentrasi hara makro dan mikro (MS1/2) yaitu sebesar 36.1%, namun tidak berbeda nyata dengan media MS konsentrasi penuh (MS0).
Pada 6 MST, persentase embrio berproliferasi pada media MS tanpa GA3 (MS0 dan MS1/2) berbeda nyata dengan media MS yang ditambahkan GA3 (MS-5G dan MS-10G). Persentase embrio berproliferasi terbesar ada pada media MS-5G yaitu sebesar 83.3%, sedangkan persentase terkecil ada pada media MS1/2. Pada semua media perlakuan, persentase embrio berproliferasi pada 7 MST sama dengan persentase pada 6 MST.
Persentase embrio berproliferasi pada media MS1/2 dan MS-5G mencapai nilai tertinggi pada 6 MST yang tidak bertambah hingga 10 MST yaitu masing-masing sebesar 40.3% dan 83.3%. Pada akhir pengamatan (10 MST), persentase embrio berkalus pada media MS1/2 dan MS-5G merupakan persentase terkecil dan terbesar, namun tidak berbeda nyata dengan media lainnya.
Perlakuan GA3 dan konsentrasi media dasar tidak berpengaruh nyata terhadap bobot dan diameter proembrio yang dihasilkan embrio endospermik fase kotiledonari mangga Arumanis klon 143 pada 10 MST. Bobot proembrio rata-rata terbesar pada media MS1/2 (573.30 mg) dan terkecil pada media MS0 (310.60 g) tidak berbeda nyata dengan media lainnya. Proembrio yang dihasilkan embrio endospermik fase kotiledonari pada media dengan MS0 memiliki bobot proembrio rata-rata yang lebih rendah dibandingkan media MS1/2. Hal ini berbanding terbalik dengan persentase embrio berproliferasi dimana persentase embrio berproliferasi pada media MS0 lebih besar dibandingkan media MS1/2. Pada media tanpa GA3, persentase embrio berkalus berbanding terbalik dengan bobot rata-rata proembrio yang dihasilkan. Bobot rata-rata proembrio yang dihasilkan embrio pada media MS yang ditambahkan GA3 berbanding lurus dengan pesentase embrio berproliferasi dimana media dengan 5 µM GA3 memiliki bobot proembrio rata-rata dan persentase embrio berproliferasi yang lebih besar dibandingkan media dengan 10 µM GA3 (Tabel 8).
Rata-rata diameter proembrio yang dihasilkan embrio endospermik fase kotiledonari terbesar terdapat pada media dengan 5 µM GA3 yaitu sebesar 9.69 mm, namun tidak berbeda nyata dengan media MS0, MS1/2, maupun MS-10G. Sedangkan diameter proembrio rata-rata yang terkecil adalah proembrio yang dihasilkan embrio endospermik fase kotiledonari pada media MS1/2 sebesar 5.52 mm yang juga tidak berbeda nyata dengan ketiga media lainnya. Pada media MS tanpa GA3, diameter rata-rata proembrio berbanding terbalik dengan bobot rata-rata proembrio yang dihasilkan (Tabel 8). Media MS1/2 yang memiliki bobot rata-rata proembrio yang terbesar dibanding media lainnya ternyata memiliki diameter rata-rata proembrio yang terkecil. Pada media MS yang ditambahkan GA3, diameter rata-rata proembrio berbanding lurus dengan bobot rata-rata proembrio yang dihasilkan embrio.
18
Tabel 8 Bobot dan diameter rata-rata proembrio yang dihasilkan embrio endospermik mangga Arumanis klon 143 pada media perlakuan GA3 Jenis Media ∑ Inokulum
awal ∑ Inokulum aseptik Bobot proembrio (mg)a Diameter proembrio (mm)a MS0 27 13 310.60 9.08 MS1/2 27 17 573.30 5.52 MS-5G 27 13 444.20 9.69 MS-10G 27 13 355.50 7.12 Pengaruh tn tn KK (%) 39.73 16.25
Keterangan: aData ditransformasi (x+0.5)1/2 sebelum diolah menggunakan SAS dan data yang ditampilkan adalah data sebelum transformasi; MS0: MS konsentrasi penuh tanpa GA3; MS1/2: MS dengan 1/2 konsentrasi hara makro dan mikro, serta konsentrasi penuh vitamin dan myo-inositol, tanpa GA3; MS-5G: MS konsentrasi penuh ditambah 5 µM GA3; MS-10G: MS konsentrasi penuh ditambah 10 µM GA3; KK: Koefisien Keragaman; tn: tidak nyata
Pembentukan Embrio Endospermik Sekunder dan Embrio Berkecambah Berdasarkan hasil pengamatan hingga 14 MST, tidak ada embrio endospermik fase kotiledonari dari sel endosperma mangga Arumanis klon 143 yang berkecambah pada media tanpa GA3 (MS0 dan MS1/2) maupun media dengan GA3 (MS-5G dan MS-10G). Embrio hanya berproliferasi dengan membentuk proembrio sekunder. Proembrio yang terbentuk mulai 2 MST tidak mengalami perkembangan menjadi embrio endospermik sekunder. Proembrio terus mengalami proliferasi. Diduga media tidak cocok untuk perkembangan proembrio menjadi embrio. Penggunaan media yang tepat dapat menginduksi terjadinya seluruh tahap perkembangan embrio, sebaliknya pada media yang kurang sesuai tidak terlihat perkembangan embrio (Purnamaningsih 2002).
Percobaan III. Perkecambahan Embrio Endospermik Mangga Varietas Arumanis Klon 143 pada Media Cair dan Padat
Pada percobaan ini embrio endospermik fase kotiledonari ditanam pada 2 jenis media yaitu media padat dan media cair dengan komposisi yang sama. Tidak terjadi kontaminasi pada kedua jenis media hingga pengamatan pada 10 MST. Embrio membentuk proembrio dan selanjutnya proembrio berkembang menjadi embrio sekunder. Terdapat embrio yang membentuk tunas, namun embrio tidak berkembang menjadi planlet.
Pembentukan Proembrio
Pada kedua media cair dan padat, embrio mengalami proliferasi dengan membentuk proembrio. Proembrio mulai terbentuk pada embrio yang dikulturkan selama 2 minggu di media perlakuan. Proembrio terbentuk pada bagian hipokotil embrio (Gambar 6). Proembrio yang terbentuk berwarna kuning dan bersifat remah (friable). Presentase embrio yang berproliferasi pada 7 MST sebesar 93.3% pada media cair dan 80% pada media padat. Besarnya presentase embrio yang
19
Tabel 9 Jumlah embrio endospermik sekunder mangga Arumanis klon 143 yang terbentuk pada media cair dan padat saat berumur 7 MST
Jenis media
∑ Embrio primer
Total embrio sekunder Fase torpedo Fase
kotiledonari Total
Cair 15 4 2 6
Padat 15 61 13 74
Gambar 6 Pembentukan proembrio dan embrio endospermik sekunder mangga Arumanis klon 143 pada media cair dan padat: A) kultur pada media cair awal tanam; B) kultur pada media cair 3 MST; C) kultur pada media cair 5 MST; D) kultur pada media padat awal tanam; E) kultur pada media padat 3 MST; F) kultur pada media padat 5 MST; G) embrio endospermik sekunder pada 5 MST di media padat
berproliferasi pada media cair diduga karena lebih banyak permukaan embrio yang mengalami kontak langsung dengan media sehingga menginisiasi sel embrio untuk membentuk proembrio. Selain itu, hara pada media cair juga lebih mudah diserap sel embrio.
Embrio Somatik Sekunder
Proembrio yang terbentuk mengalami regenerasi menjadi embrio sekunder (Gambar 6G). Embrio sekunder dihitung pada 7 MST. Media padat menghasilkan embrio endospermik sekunder lebih banyak dibandingkan media cair. Embrio endospermik sekunder yang terbentuk pada media padat sebanyak 61 embrio fase torpedo dan 13 embrio fase kotiledonari. Pada media cair embrio endospermik sekunder yang sebanyak 4 embrio fase torpedo dan 2 embrio fase kotiledonari (Tabel 9). Pada kedua media masih terdapat embrio fase globular yang berpeluang menjadi embrio dewasa.
Embrio Berkecambah
Tahap perkecambahan adalah fase dimana embrio somatik membentuk tunas dan akar (Purnamaningsih 2002). Pada saat embrio berkecambah, pucuk lembaga (plumula) yang semula berada di dalam kotiledonari mulai tumbuh (Gambar 7A). Menurut Tjitrosoepomo (2009), plumula merupakan bagian embrio
20
Gambar 7 Perkembangan embrio endospermik mangga Arumanis klon 143 yang berkecambah pada media padat: A) 3 MST; B) 4 MST; C) 5 MST; D) 10 MST
Tabel 10 Persentase kontaminasi inokulum pada 4 MST di media perkecambahan (MP)
Media asal inokulum Media prekultur Kontaminasi (%) MS0 PM 60 MS1/2 20 MS-5G 36 MS-10G 0 1/2MS-AK 56 MS0 M3 0 MS1/2 0 MS-5G 56 MS-10G 88 1/2MS-AK 88
yang terdiri atas batang lembaga beserta calon-calon daun. Tidak ada embrio yang berkecambah di media cair, namun 6.7% embrio berkecambah di media padat. Embrio yang ditanam pada media padat berkecambah saat berumur 3 MST.
Pada kecambah normal, plumula akan mengalami morfogenesis menjadi daun yang sempurna. Pengamatan pada embrio yang berkecambah di media padat, plumula tidak mengalami morfogenesis. Ukuran plumula bertambah setiap minggunya, tetapi tetap tidak mengalami morfogenesis (Gambar 7). Embrio yang berkecambah juga tidak membentuk akar. Hingga akhir pengamatan tidak didapatkan planlet hasil perkecambahan embrio somatik sel endosperma mangga Arumanis klon 143 pada media cair maupun padat.
Percobaan IV. Perkecambahan Embrio Endospermik Mangga Varietas Arumanis Klon 143 dengan Perlakuan Media Prekultur yang Berbeda
Media inisiasi embrio (MI) yang digunakan pada percobaan IV tidak mampu menginisiasi embrio mangga Arumanis klon 143 dari inokulum proembrio. Inokulum membentuk embrio fase kotiledonari setelah 3 MST di media perkecambahan (MP) dan berkecambah pada 9 MST. Pada umur 4 minggu di media perkecambahan, inokulum asal media MS-10G dan 1/2MS-AK dengan media prekultur M3 mengalami kontaninasi dengan persentase mencapai 88% (Tabel 10).
21
Gambar 8 Proliferasi proembrio mangga Arumanis klon 143 di media prekultur pada umur 7 MST: A) media PM; B) media M3
G
Prekultur
Terdapat dua jenis media prekultur dengan komposisi yang berbeda yaitu media PM dan media M3 (Tabel 1). Selama satu minggu ditanam pada media prekultur, inokulum berupa proembrio tidak menunjukkan perubahan ukuran. Proembrio juga tidak mengalami morfogenesis pada media prekultur PM dan M3. Hasil pengamatan pada sisa inokulum di media prekultur menunjukkan bahwa media prekultur memiliki potensi untuk menginduksi proliferasi proembrio jika proembrio ditanam dalam waktu yang lebih lama. Hal ini diketahui dari adanya proembrio yang berproliferasi dari sisa inokulum di media prekultur pada saat 7 minggu setelah inokulum ditanam atau 6 minggu setelah inokulum dipindah tanam ke media inisiasi (Gambar 8). Proembrio berwarna kuning dan bersifat remah (friable).
Media PM mampu menginduksi proliferasi proembrio yang lebih baik dibandingkan media M3 pada 7 MST. Beberapa unsur yang terdapat peda media PM diduga menjadi faktor yang menyebabkan kemampuan proliferasi proembrio pada media PM lebih baik daripada media M3. Media PM mengandung 5 mg/l 2,4-D, 5 mg/l GA3, 400 mg/l asam amino glutamin, 300 mg/l thrypton, dan 20 ml/l air kelapa yang tidak terdapat pada media M3. Pada penelitian Jana et al. (1994), embriogenesis somatik pada mangga monoembrionik berhasil diinduksi dengan menggunakan media MS ditambah 5 mg/l 2,4-D dan 5 mg/l GA3. Hasil penelitian Vesco dan Guerra (2001) pada tanaman Feijoa sellowiana menunjukkan bahwa penambahan glutamin 4 mM menghasilkan induksi kalus embriogenik tertinggi. Thorpe et al. (2008) menyataka bahwa air kelapa bermanfaat untuk menginduksi pertumbuhan kalus dan kultur suspensi serta untuk menginduksi morfogenesis.
Media prekultur PM dan M3 tidak mampu menginisiasi morfogenesis pada proembrio. Pengamatan hingga 10 MST, proembrio di media prekultur PM dan M3 tidak mengalami morfogenesis.
Induksi Embrio Somatik
Inokulum yang telah dikulturkan selama 1 minggu di media prekultur, kemudian disubkultur ke media induksi (MI) untuk mendapatkan embrio endospermik fase kotiledonari. Pada 2 MST di media MI, terjadi proliferasi inokulum menjadi 2 kali ukuran semula namun inokulum proembrio belum mengalami morfogenesis. Pengamatan pada 4 MST, inokulum proembrio juga belum mengalami morfogenesis menjadi embrio fase kotiledonari (Gambar 9). Hal ini berbeda dengan hasil penelitian yang dilakukan Xiao et al. (2004), dimana
22
Gambar 9 Proembrio endospermik umur 4 MST pada media induksi embrio (MI) dari media prekultur berbeda: A) inokulum asal media prekultur PM; B) inokulum asal media prekultur M3
Gambar 10 Pembentukan embrio endospermik mangga Arumanis klon 143 umur 4 MST pada media perkecambahan (MP) dengan perlakuan prekultur: A) media PM; B) media M3
embrio somatik dewasa dengan dua kotiledonari dapat dibentuk setelah PEM-like structures dari mangga monoembrionik varietas Zi-Hua dikulturkan selama 3 minggu pada media dengan komposisi hara makro B5, hara mikro MS, 5 mg/l IBA dan 60 g/l sukrosa (komposisi media yang sama dengan media MI). Perbedaan jenis mangga mangga Arumanis yang poliembrionik dan Zi-Hua yang monoembrionik diduga menjadi faktor yang menyebabkan respon kedua mangga berbeda meskipun ditanam pada media yang sama. Menurut Xiao et al. (2004) embriogenesis sangat tergantung pada kultivar mangga.
Perkecambahan Embrio Somatik
Inokulum yang telah dikulturkan selama 2 minggu di media induksi selanjutnya disubkultur ke media perkecambahan. Semua inokulum dari media asal dan media prekultur yang berbeda masih dalam fase proembrio. Inokulum dari kedua media prekultur mulai membentuk embrio somatik fase torpedo dan fase kotiledonari setelah 3 minggu berada pada media perkecambahan yang mengandung hara makro B5, hara mikro dan vitamin MS, ditambah 5 mg/l kinetin, 500 mg/l asam amino glutamin, 100 mg/l thrypton, 200 ml/l air kelapa, dan 40 g/l sukrosa (Gambar 10).
Pada umur 4 dan 7 MST di media perkecambahan (MP), jenis media prekultur tidak berpengaruh nyata terhadap jumlah embrio fase kotiledonari yang dihasilkan. Media asal inokulum berpengaruh nyata terhadap jumlah embrio fase kotiledonari yang dihasilkan pada 4 dan 7 MST. Tidak ada interaksi antara jenis media prekultur dan media asal inokulum yang mempengaruhi jumlah embrio fase kotiledonari yang dihasilkan pada 4 MST, namun terjadi interaksi pada 7 MST (Tabel 11).