Summary Effects of magnesium deficiency and variation in nitrate to ammonium ratio on needle histology and chlorophyll concentration were investigated in current-year and one-year-old needles of clonal Norway spruce trees (Picea abies (L.) Karst.). Six-year-old trees were grown for one year in sand culture with circulating nutrient solutions containing a suffi-cient (0.2 mM) or a limiting (0.04 mM) concentration of Mg. The nitrogen concentration was not varied (5 mM), but the NO3−/NH4+-ratio was adjusted to 0.76 in Mg-sufficient and to 1.86, 0.76 or 0.035 in Mg-limited plants. Visible symptoms of Mg deficiency occurred only in current-year needles, indicat-ing adequate Mg nutrition before the experiment. Under con-ditions of Mg limitation, chlorophyll and Mg concentrations were lowest in needles of trees supplied with NH4+ as the major nitrogen source and highest in needles of trees supplied with NO3− as the major nitrogen source. In current-year and one-year-old needles, starch accumulation induced by Mg defi-ciency was increased when NH4+ was the major nitrogen source. The accumulation of tannin spherules in current-year needles, which occurred in response to Mg deficiency, also increased with decreasing NO3−/NH4+-ratios. Deficient Mg supply caused premature aging in tissues of the vascular bundle, as indicated by modifications of the cambium and increased amounts of collapsed sieve cells. The number of collapsed sieve cells was slightly lower in needles grown in a NH4+-dominated nutrient regime than in needles grown in a NO3−-dominated nutrient regime. We conclude that NH4+ was not directly toxic to Norway spruce trees at the applied concentrations. However, effects of Mg deficiency were considerably greater in an NH4+-dominated nutrient regime than in a NO3−-dominated nutrient regime. Keywords: chlorophyll, magnesium deficiency, needle histol-ogy, Picea abies, starch accumulation.
Introduction
Nitrogen availability has often been thought to limit plant productivity, especially in forest trees. In recent decades, the input of atmospheric nitrogen to forest ecosystems has in-creased dramatically (Skeffington 1990). Nitrogen depositions in the form of NOx, NH3, NO3− and NH4+ reach a total of about 20 kg N ha−1 year−1 in central Europe, and may be as high as 40 to 60 kg N ha−1 year−1 in some areas (van Breemen et al.
1987). Not only the amount but also the nature of the deposited nitrogen may affect forest ecosystems (Nihlgård 1985, van Dijk and Roelofs 1988, Zöttl 1990). In soils with low cation availability, high nitrogen inputs, especially in the form of NH4+, induce ion imbalances by either interfering with the uptake of other ions or causing a relative deficiency in tree tissues as a result of increased growth rates (Huettl 1990, Weissen et al. 1990). This nutritional stress can be further enhanced as a result of high atmospheric acidity input leading to soil acidification and displacement of adsorbed Mg and Ca by Al ions in the cell walls of roots (Ulrich 1987). In addition, direct uptake of gaseous ammonia and nitrogen oxides may cause leaf damage as well as cation losses from the foliage (van der Eerden 1982, Nihlgård 1985).
In acidic forest soils of the temperate zone, NH4+ is the major nitrogen source and is preferentially taken up by some conifer species, including Norway spruce (Picea abies (L.) Karst.) (Marschner et al. 1991, Flaig and Mohr 1992, Gezelius and Näsholm 1993). Uptake of NH4+ by roots leads to acidification of the soil solution causing leaching of cations from the upper soil horizons. Because NH4+ uptake is believed to compete with the uptake of cations, Mg deficiencies may become more pronounced in trees growing on Mg-poor soils; several studies have shown that increased NH4+ nutrition causes a decrease in the uptake of cations and a reduction in foliar concentrations of Mg, K, and P (Ingestad 1979, Boxman and Roelofs 1988, Gijsman 1990), and opposite effects were observed by increas-ing the supply of NO3− (van Dijk and Roelofs 1988, Mohr 1994).
The yellowing symptoms associated with forest decline have often been related to Mg deficiencies (Landmann et al. 1987, Forschner and Wild 1988, Cape et al. 1990, Huettl et al. 1990, Hölldampf et al. 1993, Ke and Skelly 1994). The histo-logical damage in conifer needles caused by Mg deficiencies is well documented. A lack of Mg induces abnormal accumu-lation of starch in the mesophyll cells of chlorotic older nee-dles, possibly because export of carbohydrates is impeded by the collapse of phloem cells (Fink 1989, Forschner et al. 1989, Fink 1991). It is not known if the cellular damage induced by Mg deficiency is modified by the form of nitrogen supplied. Beside indirect effects of the supplied N form on uptake and transport of Mg, and therefore on Mg nutrition, direct impacts of the applied NO3−/NH4+-ratio might also interact with Mg
Histology of magnesium-deficient Norway spruce needles influenced
by nitrogen source
LAURENCE PUECH and BEATE MEHNE-JAKOBS
Institute of Forest Botany and Tree Physiology, Albert-Ludwigs-University, Bertoldstrasse 17, D-79085 Freiburg, Germany
Received July 6, 1995
deficiency. For example, high NH4+ supply has been reported to alter leaf chloroplasts of cultivated plants, inducing a suppres-sion of starch synthesis (Puritch and Barker 1967, Matsumoto et al. 1969, Mehrer and Mohr 1989). The aim of our investiga-tion, therefore, was to examine the influence of different NO3−/NH4+-ratios on the Mg, chlorophyll and starch concentra-tions and the histology of needles of young Mg-deficient Norway spruce trees.
Material and methods
Plant material and study design
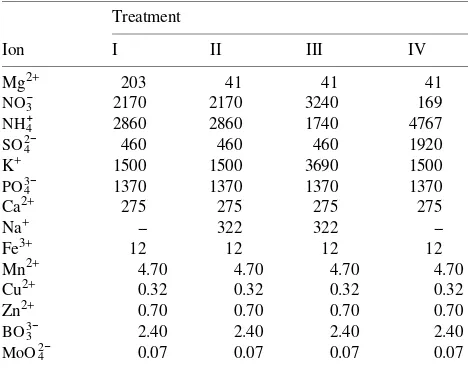
In October 1991, 100 six-year-old, non-mycorrhizal Norway spruce trees (Picea abies) derived from one clone (No. 1213/113, provenance Rothenkirchen 84011, Franken-wald) were each maintained in sand culture in a 70-liter pot in a special outdoor facility. Each pot was individually supplied with nutrients by means of a continuously circulating solution that was refilled continuously and completely replaced every 3 days. The composition of the nutrient solution, which was designed to achieve optimum growth according to Ingestad’s principles (Ingestad 1962, 1979), is presented in Table 1. Initially, the plants were grown with a sufficient (Treatment I) or a limiting Mg supply to induce Mg deficiency (Treat-ment II). In spring 1992, the nitrogen source was varied in a subset of trees from Treatment II to achieve a NO3−- or NH4+ -dominated nutrition (Treatments III and IV) (see Table 1). However, the total nitrogen concentration of 5 mM was not changed. Increases in NO3− and NH4+ concentrations were asso-ciated with increases in the K+ and SO
4
2− concentrations,
re-spectively, of the nutrient solutions (Table 1). During each 3-day period of circulation through the plant pots, the pH of the nutrient solutions decreased from an initial value of 6.0 (5.7 in Treatment IV) to 4.1--3.8 in the NH4+-dominated nutrient solutions (Treatments I, II and IV). In the NO3−-dominated treatment (Treatment III), the pH remained relatively stable at
pH 6.0. The plants were subjected to the experimental regimes for one growing season.
Light microscopy
At the end of September 1992, 10 current-year and 10 one-year-old sun-exposed needles were removed from shoots of the third whorl of trees randomly selected from each treatment. Samples were collected at midday (1100--1300 h) on a sunny day. Three approximately 2-mm-long segments were cut from the middle of each needle and immediately fixed in cold glutaraldehyde in 0.1 M phosphate buffer, pH 7.2, for one day. The samples were then dehydrated in an ascending series of acetone concentrations and embedded in methacryl resin (S. Fink, Albert-Ludwig-Universität, Freiburg, FRG, unpub-lished method). Semithin sections (3 µm) of the embedded segments were cut with a diamond knife on a Leica-2065-mi-crotome (Leica Instruments GmbH, Nuβloch, FRG).
Histological observations and a differential survey of carbo-hydrates, phenolics and lipid-like compounds were performed on the sections. The sections were stained for polysaccharides containing vicinal glycol groups with periodic acid and Schiffs reagent (PAS) as described by Gerlach (1977) and mounted in Eukitt (Hans Thoma, Freiburg, FRG). The autofluorescent properties of lignin, phenolics and lipids under excitation at 395--440 nm were used as the counterstain. For this purpose, the sections were observed on a Zeiss-Axiophot microscope with epi-fluorescence (Carl Zeiss, Oberkochen, FRG) using the filters BP 395--440 (excitation filter), FT 460 (beamsplit-ter) and LP 470 (barrier fil(beamsplit-ter). Photographs were taken with an integrated camera on 100 ASA black-and-white film.
Lignin was stained with phloroglucin in HCl (von Aufseβ 1973). Lipids were stained with Nile blue (Jensen 1962) and suberin and cutin were stained with Sudan IV (Jensen 1962). Tannins were stained with ferric chloride (Jensen 1962) and proteins were stained with Coomassie blue R250 in Clarke’s solution (Cawood et al. 1978).
Determination of starch, chlorophyll and nutrient elements
At the end of September 1992, current-year and one-year-old shoots were harvested around noon from randomly selected trees (n = 6 per treatment). Needles were removed, immedi-ately frozen in liquid nitrogen and stored at −80 °C. The frozen needle tissue was homogenized with a microdismembrator (Braun, Melsungen, FRG). The resulting powder was freeze-dried at --25 °C and stored under vacuum at −20 °C.
For the determination of foliar starch concentrations, 2- to 6-mg aliquots of needle homogenate were extracted for 1 h with 0.5 ml of 0.5 M NaOH in Eppendorf vials (Eppendorf-Netheler-Hinz GmbH, Hamburg, FRG) at ambient tempera-ture (Dekker and Richards 1971). To remove free glucose, the extract was incubated at 95 °C for 3 min, adjusted to pH 4.6 with 0.5 M acetic acid, and then centrifuged at 10,000 g for 5 min (Einig and Hampp 1990). Twenty- to 60-ml aliquots of the supernatant were used for enzymatic determination of starch according to Outlaw and Manchester (1979). The pro-cedure was optimized to obtain a 90% or higher recovery of an
Table 1. Mineral nutrient composition of the four treatment solutions (µM). The balance between anions and cations was made with H+.
added internal standard (maize starch, Sigma Chemical Co., St. Louis, MO).
Chlorophyll concentrations were determined by spectro-photometry after needle tissue homogenization and extraction in 80% acetone (Ziegler and Egle 1965), and calculated ac-cording to Lichtenthaler (1987). Magnesium and K concentra-tions in needles were analyzed after dry ashing and digestion with HNO3/HCl by the AAS/AES flame technique (Perkin
Elmer 4000, Perkin-Elmer & Co GmbH, Überlinger, FRG). The N and S concentrations were determined with a CNS analyzer (Carlo Erba Instruments, Rodano, Italy). To evaluate the data, means and standard errors (SE) were calculated separately for each needle age class and treatment. Analysis of variance (one-way ANOVA) and the Tukey test were used to analyze differences among the treatments within each needle age class.
Results
Needle element and chlorophyll concentrations
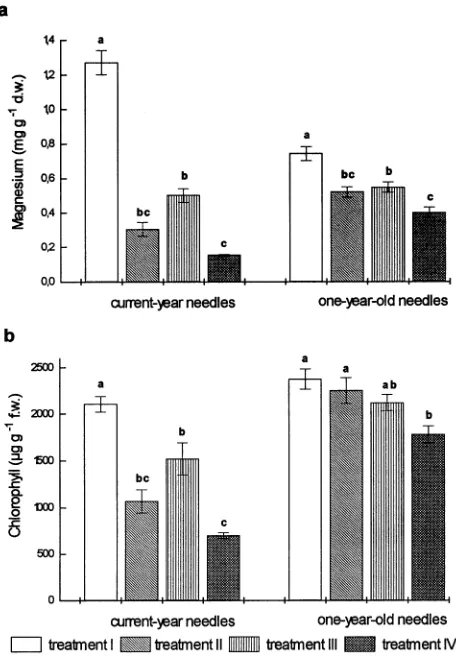
The influence of nitrogen sources on Mg deficiency was ana-lyzed by varying the NO3−/NH4+-ratio of the nutrient solution (0.76 = Treatment II, 1.86 = Treatment III, and 0.035 = Treat-ment IV, Table 1). The nutrient solution used in the Mg-suffi-cient treatment contained a fivefold higher concentration of Mg and a NO3−/NH4+-ratio of 0.76 (Treatment I). After one growing season, Mg concentrations in current-year and one-year-old needles of Mg-limited trees differed significantly depending on whether NO3− or NH4+ was the major nitrogen source (Figure 1a). Current-year needles from trees supplied with NO3− as the major N source (Treatment III) exhibited two-to threefold higher Mg concentrations than current-year need-les of trees supplied with NH4+ as the major N source (Treat-ment IV) (Figure 1a). The differences in foliar Mg concentrations between trees grown in the Mg-limited Treat-ments III and IV cannot be explained by dilution effects caused by different growth rates of the needles, because both needle weight and Mg concentration were higher in needles grown in the NO3−-dominated nutrient treatment (Treatment III) than the NH4+-dominated treatment (Treatment 1V) (Table 2 and Fig-ure 1a). Chlorophyll concentrations of current-year needles were influenced similarly by the treatments (Figure 1b). Com-pared with current-year needles, there was a similar but smaller effect of nitrogen source on Mg concentrations of one-year-old needles (Figure 1a). In contrast, reductions in chlorophyll concentrations of one-year-old needles were only significant in the NH4+-dominated nutrient treatment (Treatment IV, Fig-ure 1b).
Visible symptoms of yellowing occurred only in current-year needles of trees in the Mg-limiting treatments. The symp-toms were most pronounced in trees in the NH4+-dominated treatment (Treatment IV), followed by trees in the treatment with a balanced NO3−/NH4+-ratio (Treatment II). Only slight yellowing was observed in current-year needles of trees in the NO3−-dominated treatment (Treatment III). In accordance with earlier findings, the Mg concentrations, calculated as % of
Figure 1. Magnesium (a) and chlorophyll concentrations (b) in cur-rent-year and one-year-old needles of Norway spruce grown with sufficient (Treatment I) or limiting Mg supply (Treatments II--IV). Ratios of NO3−/NH4+ were: 0.76 (Treatments I and II), 1.86
(Treat-ment III) and 0.035 (Treat(Treat-ment IV). Bars indicate means (± SE, n = 6). Different letters indicate significant differences between treatments.
Table 2. Fresh weight of 100 needles (gfw), N, K and S concentrations (mg g−1 dry weight), and Mg concentrations in % of the N concentra-tion in current-year needles of Norway spruce grown with a sufficient (Treatment I) or a limited Mg supply (Treatments II, III and IV). Ratios of NO3−/NH4+ in the nutrient solutions were: 0.76 (Treatments I
and II), 1.86 (Treatment III) and 0.035 (Treatment IV) (n = 6, SE is given in brackets). Different letters within rows indicate significant differences between treatments.
Treatment
I II III IV
gfw 0.74 (0.05) a 0.75 (0.09) a 0.89 (0.10) a 0.74 (0.07) a
N 32 (0.7) a 28 (0.9) ab 27 (1.1) b 28 (1.6) ab S 1.7 (0.07) a 1.4 (0.12) a 1.3 (0.14) a 1.4 (0.15) a
K 10.3 (0.6) b 11.7 (0.8) b 21.2 (1.1) a 6.9 (0.2) c
Mg in 4.0 (0.21) a 1.1 (0.17) c 1.8 (0.26) b 0.6 (0.03) c
nitrogen concentration, were slightly below the physiological threshold value of 2% for Norway spruce (Mehne-Jakobs 1996) in the NO3−-dominated treatment, and well below the threshold value in the NH4+-dominated treatments (Treatments II and IV; Table 2).
Needle N and S concentrations were slightly lower in all Mg-limited treatments than in the Mg-sufficient treatment (Table 2). Needle S concentrations were not influenced by variation in the NO3−/NH4+-ratio or by a fourfold increase in nutrient solution SO42− concentration in the NH4+-dominated treatment (Treatment IV). Needle K concentrations, however, decreased significantly with decreasing NO3−/NH4+-ratios. In the NO3−-dominated treatment (Treatment III), high foliar K concentrations might reflect the increased K concentration in this treatment (K served as the counterion for NO3−, cf. Ta-ble 1).
Microscopy
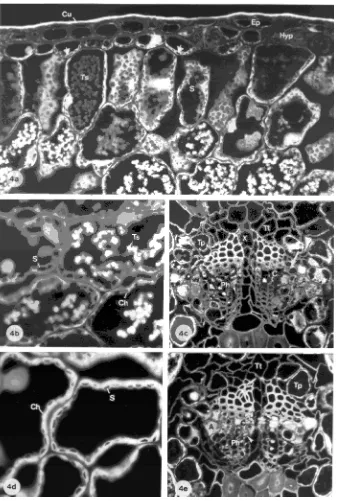
Figures 2--4 are fluorescence micrographs of needle sections stained with PAS. In these micrographs, the red fluorescence of stained polysaccharides and the nuclei appear grey, whereas the blue-green fluorescence of lignin and the yellow fluores-cence of lipids appear white.
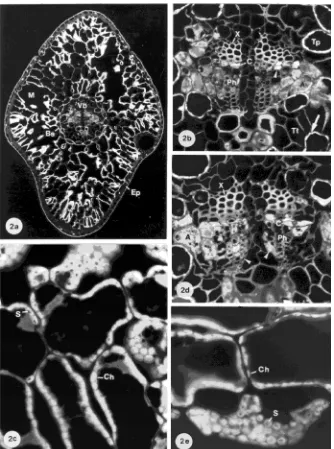
Vascular bundle Healthy current-year needles were charac-terized by an intact epidermis, a turgid mesophyll and an intact vascular bundle (Figure 2a). Between the xylem and phloem, cambial cells (C) are organized in two or three rows (Fig-ure 2b). Their complex content was characterized by a yellow-brown fluorescence (arrowhead) and showed a positive staining with Sudan IV for lipids and with Coomassie blue for proteins. All sieve cells (Ph) had a wide open lumen. Younger sieve cells showed a heterogeneous content whereas the older
sieve cells appeared empty. The albuminous cells (A) bordering the phloem were well developed and filled with light fluoresc-ing material includfluoresc-ing large nuclei. In the one-year-old needles, aging is revealed by the crushed older sieve cells (Figure 2d: arrowhead).
The Mg-deficient treatment caused premature aging in the vascular bundle, characterized by an increased proportion of sieve cell deformations (Figures 3b, 3d, 4c, 4e, Ph) in current-year and one-current-year-old needles, and by modifications of the cambium in one-year-old needles including additional cell rows, interruptions of cell rows and cell distortions (Figure 3d: arrowhead, Figure 4e). Nitrogen source had no influence on the symptoms expressed by the cambium. In the Mg-deficient treatments, the amount of collapsed sieve cells was increased in needles grown in the NO3−-dominated treatment ment III, Figures 3b and 3d) or balanced treatment
(Treat-ment II, sections not shown), but was only slightly increased in the NH4+-dominated treatment (Treatment IV, Figures 4c and 4e) compared with phloem cells in needles from Mg-sufficient control trees. However, a proportion of the sieve cells remained functional in all trees in the Mg-deficient treatments.
Mesophyll Chloroplasts of needles from the Mg-sufficient treatment were almost devoid of starch at the end of the growing season (Figures 2a, 2c, 2e). Only small starch grains (S) were observed in chloroplasts (Ch) of the mesophyll surrounding the vascular bundle and in the endodermis and transfusion paren-chyma cells (Figures 2b: arrow, 2c, 2e). The amount of starch in needles of Mg-deficient trees cultivated with either a bal-anced NO3−/NH4+-ratio (Treatment II) or an NO3−-dominated N form (Treatment III) did not differ markedly from that in needles of Mg-sufficient trees (Figures 3a, 3e; sections of Treatment II not shown). The starch concentration of
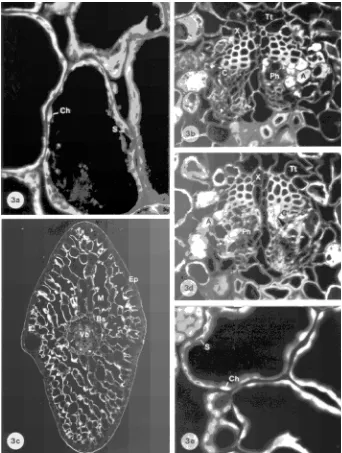
current-Figure 3. Cross sections of need-les from Treatment III (Mg-defi-cient, NO3−-dominated nutrition).
(a) Mesophyll cells of a current-year needle (Magnification = 353×). The number of starch grains in the chloroplasts shown on the photograph is relatively high for the treatment. (b) Vascu-lar bundle of the same needle (195×). Some sieve cells distor-tions were induced by the treat-ment (arrowhead). (c) One-year-old needle (30×). (d) Vascular bundle of a one-year-old needle with a higher propor-tion of distorted sieve cells (218×). Cambium with an in-creased division array and some distorted cells (arrowhead). (e) Small starch grains in chloro-plasts of mesophyll cells of a one-year-old needle (330×).
year needles in Treatments II and III was very heterogeneous. In the NH4+-dominated treatment (Treatment IV), there was a large increase in current-year needles. Many large starch grains were observed in all mesophyll cells (Figures 4a and 4b) and also in guard cells of the stomata and in hypodermal cells (Figures 4a: arrows). Starch accumulation also occurred in the transfusion parenchyma cells (Tp) and in some of the albumi-nous cells (A) bordering the youngest sieve cells (Figure 4c: arrow). Starch grains were smaller in one-year-old needles than in current-year needles of Treatment IV (Figure 4d).
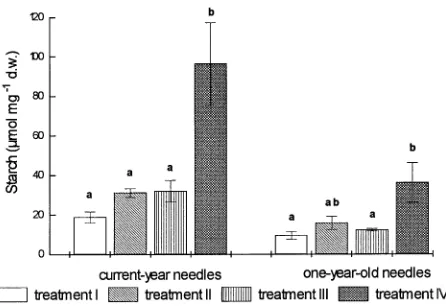
The microscopic observations were confirmed by biochemi-cal analysis. Compared to controls, deficient Mg supply
com-bined with a balanced NO3−/NH4+ supply or NO3−-dominated nutrition caused a twofold increase in starch concentration of the current-year needles, whereas the Mg-deficient plus NH4+ -dominated nutrient regime caused a fivefold increase in starch concentration of current-year needles (Figure 5). In one-year-old needles, treatment-related effects were similar to those of current-year needles but less pronounced.
Few spherules were observed in mesophyll cells of current-year needles in the Mg-sufficient treatment and in the Mg-de-ficient plus NO3−-dominated treatment (Treatments I and III). The number of spherules was greater in Mg-deficient needles when nitrogen sources were balanced (Treatment II, sections
Figure 4. Cross sections of needles from Treatment IV (Mg-deficient, NH4+-dominated nutrition). (a)
not shown), and the increase was greatly enhanced when the nitrogen supply was NH4+-dominated (Treatment IV, see Fig-ures 4a and 4b: Ts). Although all spherules exhibited the same autofluorescence under blue-violet excitation, they stained dif-ferently depending on size. The smaller droplets present in the outer mesophyll cells of the needles grown in the NH4+ -domi-nated nutrient regime (Figure 4a) were positively stained by Nile blue for lipids, Coomassie blue for proteins and ferric chloride for tannins, whereas the larger droplets in the inner part of the mesophyll remained almost unstained and showed autofluorescence. Because the staining intensity increased with decreasing spherule size, it is possible that the stains were physically adsorbed on the surface of the large drops. No spherules were observed in one-year-old needles in any of the treatments.
Discussion
In Norway spruce, Mg concentrations of 0.9 to 1.7 mg g−1 dry
matter are considered optimum, whereas values of less than 0.7 mg g−1 indicate deficiency (Ingestad 1962, 1979, Bergmann 1988). Throughout the literature, Mg deficiency symptoms in Norway spruce are described as tip-yellowing of the older needles, whereas the younger needles remain green (Mies and Zöttl 1985, Bergmann 1988, Kaupenjohann and Zech 1989, Lange et al. 1989). This difference between need-les of different ages has been related to the translocation of Mg from one-year-old needles to the new flush (Weikert et al. 1989). In the present study, however, we observed that Mg deficient treatment caused chlorosis of current-year needles, whereas one-year-old needles remained green, indicating that Mg translocation into the developing shoots did not decrease the Mg concentrations of the one-year-old needles below the deficiency threshold value (Figure 1). Conversely, the amount of Mg supplied by root uptake and by translocation from
one-year-old needles was insufficient to prevent current-year needles from developing chlorosis. Previously, Mehne-Jakobs (1994) observed yellowing of current-year needles of Norway spruce trees grown with a severely restricted Mg supply (5 µM), whereas current-year leaves of trees supplied with a moderately reduced Mg supply (41 µM) did not exhibit pig-ment loss. A difference in the availability of nitrogen likely accounts for the occurrence of current-year needle chlorosis under conditions of moderately reduced Mg supply in the present study and its absence in the previous study (Mehne-Jakobs 1994): a higher nitrogen concentration was supplied in the present experiment than in the earlier study (5 versus 3.5 mM). There is evidence that the development of Mg defi-ciency in conifers depends on the relative availabilities of Mg and nitrogen (Ingestad 1979, Weissen et al. 1990). Previously, the physiological threshold value for Mg in needles was found to be 2% of the nitrogen concentration (Mehne-Jakobs 1996). Magnesium concentrations in current-year needles were below this threshold value in all of the Mg-limited treatments (Treat-ments II, III and IV, Table 2). Concentrations of both Mg and chlorophyll were strongly influenced by nitrogen source (Fig-ures 1a and 1b). Current-year needles of trees grown with NO3− as the predominant form of N exhibited only slight reductions in Mg and chlorophyll concentrations (Treat-ment III, Figure 1b), whereas NH4+-dominated nutrition caused severe pigment loss (Treatment IV, Figure 1b). Because both chlorophyll and Mg concentrations were equally influenced by nitrogen source, the intensity of the chlorosis can be attributed to Mg deficiency rather than to toxic effects of NH4+. The decrease in needle Mg concentration with increasing NH4+ supply cannot be explained by dilution caused by increased growth, because needle weight was highest in the treatment with the lowest NH4+ supply (Treatment III, Table 2). The negative influence of the reduced NO3−/NH4+-ratio on foliar Mg concentration is likely to have been caused by competition between NH4+ uptake and uptake of other cations (Marschner 1995). Decreased foliar Mg concentrations combined with enhanced yellowing have also been observed after ammonium fertilization of Norway spruce stands suffering from Mg defi-ciency (Feger 1990, Hüttl and Fink 1991).
Histological investigation revealed slightly premature aging of the vascular bundle in Mg-deficient needles, which was characterized by an increase in sieve cell distortions, especially when nutrition was NO3−-dominated, and by modifications of the cambium in all Mg-deficiency treatments. Similar symp-toms of precocious aging have been observed in needles col-lected from Mg-deficient Norway spruce (Fink 1989, Forschner et al. 1989, Hannick et al. 1993). Hannick et al. (1993) noted that the abnormalities first occurred in the cam-bium, then in the phloem, and finally in the xylem. In the present study, the Mg-deficient needles generally exhibited alterations of the cambium but few anomalies of the phloem cells. Collapse of sieve cells is a characteristic structural symp-tom in chlorotic older needles of Mg-deficient spruce (Para-meswaran et al. 1985, Fink 1989, Forschner et al. 1989, Fink 1991). In the present experiment, the trees were exposed to a deficient Mg supply for only one growing season, and this did
Figure 5. Starch concentrations in current-year and one-year-old need-les of Norway spruce trees grown with sufficient (Treatment I) or limiting Mg supply (Treatments II--IV) (means ± SE, n = 6). Treat-ments refer to the following NO3−/NH4+-ratios in the nutrient solutions:
not induce true phloem collapse either in the yellowing cur-rent-year or in the green one-year-old needles. However, simi-lar treatments applied after a one-year period of Mg deficiency treatment induced severe phloem collapse and cambial abnor-malities in one-year-old needles (Puech and Mehne-Jakobs, unpublished results). We conclude, therefore, that the minor anomalies in the vascular area of Mg-deficient needles could represent an early stage of pathological alteration that precedes phloem collapse caused by prolonged Mg deficiency.
Because phloem collapse induced by Mg deficiency has only been observed in one-year-old and older needles, it might be associated with the translocation of Mg from one-year-old needles to the developing shoots during spring. Fink (1992) suggested that during flushing in spring, Mg is quickly moblized from the area surrounding the phloem, leading to a sudden and localized severe lack of Mg. Because Mg plays an important role in the phloem loading process (Balke and Hodges 1975, Giaquinta 1983), a severe lack of local Mg might cause a large decrease in osmotic pressure in the sieve cells, which consequently collapse. Drastic reductions in phloem loading have been observed in source leaves of bean as an early effect of Mg deficiency (Cakmak et al. 1994). In the present study, the one-year-old needles probably still con-tained enough Mg to compensate for any localized lack of Mg in the vascular area caused by translocation of Mg into the new flush, and therefore phloem collapse was prevented (cf. Fig-ure 1).
Another symptom apparently related to low Mg concentra-tions, was the occurrence of spherules or droplets in the meso-phyll cells of current-year needles. Similar spherules have been described previously and identified as vacuolar tannin inclusions (Chafe and Durzan 1973, Baur and Walkinshaw 1974, Chabot and Chabot 1975), and were probably condensed tannins or proanthocyanidins (Haslam 1988). Multiple stain-ing of the spherules might be explained by their association with a wide range of cellular compounds. Accumulation of polyphenols and subsequently tannins in plant cells may rep-resent a protective mechanism against herbivores (Claussen et al. 1992). In addition, such compounds have been reported to accumulate in plants exposed to various abiotic stress fac-tors, such as air pollutants (Soikkeli 1981, Jordan et al. 1991) or mineral deficiencies (Fink 1989, Forschner et al. 1989). Based on the findings that condensed tannins could be in-volved in antioxidative processes in plants (Torel et al. 1986, Feucht et al. 1994), and that Mg deficiency may induce oxida-tive stress (Polle et al. 1994), we suggest that the accumulation of spherules in the mesophyll cells represents a defense reac-tion against oxidative stress induced by Mg deficiency.
Starch accumulated in mesophyll cells of Mg-limited trees, especially in current-year needles, even though CO2-fixation
rates were reduced to less than 50% of those of the Mg-suffi-cient needles (Mehne-Jakobs, unpublished results). Increased amounts of starch in Mg-deficient spruce needles have been observed in both field investigations and controlled experi-ments (Forschner et al. 1989, Fink 1991, Mehne-Jakobs 1995). Photosynthate accumulation occurs before chlorophyll con-centrations decrease (Mehne-Jakobs 1995), and is probably a
result of the reduction in phloem export of sucrose caused by Mg deficiency (Cakmak et al. 1994).
Accumulations of starch and tannin spherules in needle mesophyll cells were most pronounced in the NH4+-dominated treatment, which contained the lowest concentrations of Mg and chlorophyll. Because ammonium toxicity reduces starch synthesis (Puritch and Barker 1967, Matsumoto et al. 1969, Mehrer and Mohr 1989), the increased symptoms observed in the NH4+-dominated treatment can be attributed to an increased severity of Mg deficiency, most probably induced by cation antagonism at the root level, rather than to phytotoxic effects of ammonium.
References
Balke, N.E. and T.K. Hodges. 1975. Plasma membrane adenosine triphosphatase of oat roots. Activation and inhibition by Mg2+ and ATP. Plant Physiol. 55:83--86.
Baur, P.S. and C.H. Walkinshaw. 1974. Fine structure of tannin accu-mulations in callus cultures of Pinus elliotti (slash pine). Can. J. Bot. 52:615--619.
Bergmann, W. 1988. Ernährungsstörungen bei Kulturpflanzen. G. Fis-cher Verlag, Stuttgart, New York, pp 762.
Boxman, A.W. and J.G.M. Roelofs. 1988. Some effects of nitrate versus ammonium nutrition on the nutrient fluxes in Pinus sylvestris seedlings. Effect of mycorrhizal infection. Can. J. Bot. 66:1091--1097.
Cakmak, I., C. Hengeler and H. Marschner. 1994. Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants. J. Exp. Bot. 45:1251--1257.
Cape, J.N., P.H. Freer-Smith, I.S. Paterson, J.A. Parkinson and J. Wolfenden. 1990. The nutritional status of Picea abies (L.) Karst. across Europe, and implications for ‘‘forest decline.’’ Trees 4:211--224.
Cawood, A.H., V. Potter, and H.G. Dickinson. 1978. An evaluation of Coomassie blue as a stain for quantitative microdensitometry of proteins in sections. J. Histochem. Cytochem. 26:645--650. Chabot, J.F. and B.F. Chabot. 1975. Developmental and seasonal
patterns of mesophyll ultrastructure in Abies balsamea. Can. J. Bot. 53:295--304.
Chafe, S.C. and D.J. Durzan. 1973. Tannin inclusions in cell suspen-sion cultures of white spruce. Planta 113:251--262.
Claussen, T.P., P.B. Reichardt, J.P. Bryant and F. Provenza. 1992. Condensed tannins in plant defense: a perspective on classical theories. In Plant Polyphenols. Eds. R.W. Hemingway and P.E. Laks. Plenum Press, New York, pp 639--651.
Dekker, R.F.M. and G.N. Richards. 1971. Determination of starch in plant material. J. Sci. Food Agric. 22:441--444.
Einig, W. and R. Hampp. 1990. Carbon partitioning in Norway spruce: amounts of fructose 2,6-biphosphate and of intermediates of starch/sucrose synthesis in relation to needle age and degree of needle loss. Trees 4:9--15.
Feger, K.H. 1990. Bewertung experimentell erhöhter Stickstoff- und Schwefeleinträge in zwei Fichtenökosystemen im Schwarzwald. VDI-Berichte 837:1209--1229.
Feucht, W., D. Treutter and E. Christ. 1994. Accumulation of flavanols in yellowing beech leaves from forest decline sites. Tree Physiol. 14:403--412.
Fink, S. 1991. Structural changes in conifer needles due to Mg and K deficiency. Fert. Res. 27:23--27.
Fink, S. 1992. Histologische und histochemische Untersuchungen zur Nährstoffdynamik in Waldbäumen im Hinblick auf die Neuartigen Waldschäden. Kernforschungszentrum Karlsruhe, PEF-Bericht 98, 88 p.
Flaig, H. and H. Mohr. 1992. Assimilation of nitrate and ammonium by Scots pine (Pinus sylvestris) seedlings under conditions of high nitrogen supply. Physiol. Plant. 84:568--576.
Forschner, W. and A. Wild. 1988. Comparative investigation on the nutrient composition of healthy and injured spruces of different locations. In Air Pollution and Ecosystems. Ed. P. Mathy. D. Reidel Publishing Co., Dordrecht, The Netherlands, pp 604--608. Forschner, W., V. Schmitt and A. Wild. 1989. Investigations on the
starch content and ultrastructure of spruce needles relative to the occurrence of novel forest decline. Bot. Acta 102:208--221. Gerlach, D. 1977. Botanische Mikrotechnik. Georg Thieme Verlag,
Stuttgart, 311 p.
Gezelius, K. and T. Näsholm. 1993. Free amino acids and protein in Scots pine seedlings cultivated at different nutrient availabilities. Tree Physiol. 13:71--86.
Giaquinta, R.T. 1983. Phloem loading of sucrose. Annu. Rev. Plant Physiol. 34:347--387.
Gijsman, A.J. 1990. Nitrogen nutrition of Douglas-fir (Pseudotsuga menziesii) on strongly acid sandy soil. I. Growth, nutrient uptake and ionic balance. Plant Soil 126:53--61.
Hannick, A.F., L. Waterkeyn, F. Weissen and H.J. van Praag. 1993. Vascular tissue anatomy of Norway spruce needles and twigs in relation to magnesium deficiency. Tree Physiol. 13:337--349. Haslam, E. 1988. Plant polyphenols (syn. vegetable tannins) and
chemical defense----a reappraisal. J. Chem. Ecol. 14:1789--1805. Hölldampf, B., A.V. Barker and G. Smith. 1993. Mineral element
composition of declining and healthy stands of red spruce in west-ern Massachusetts. Commun. Soil Sci. Plant Anal. 24:1937--1944. Huettl, R.F. 1990. Nutrient supply and fertilizer experiments in view
of N saturation. Plant Soil 128:45--58.
Huettl, R.F. and S. Fink. 1991. Pollution, nutrition and plant function.
In Plant Growth: Interaction with Nutrition and Environment. Eds. J.R. Porter and D.W. Lawson. Cambridge University Press, Cam-bridge, U.K., pp 207--226.
Huettl, R.F., S. Fink, H.-J. Lutz, M. Poth and J. Wisniewski. 1990. Forest decline, nutrient supply and diagnostic fertilisation in south-western Germany and in southern California. For. Ecol. Manage. 30:341--350.
Ingestad, T. 1962. Macroelement nutrition of pine, spruce and birch seedlings in nutrient solutions. Medd. Statens Skogsforskn. Inst. 51:1--150.
Ingestad, T. 1979. Mineral requirements of Pinus sylvestris and Picea abies seedlings. Physiol. Plant. 45:373--380.
Jensen, W.A. 1962. Botanical histochemistry. W.H. Freeman and Co., San Francisco, London, 408 p.
Jordan, D.N., T.H. Green, A.H. Chappelka, B.J. Lockaby, R.S. Meldahl and D.H. Gjerstad. 1991. Response of total tannins and phenolics in lobolly pine foliage exposed to ozone and acid rain. J. Chem. Ecol. 17:505--513.
Kaupenjohann, M. and W. Zech. 1989. Waldschäden und Düngung. Allg. Forstz. 37:1002--1008.
Ke, J. and J.M. Skelly. 1994. Relationships between symptoms ex-pressed by Norway spruce and foliar and soil elemental status. Water Air Soil Pollut. 74:289--305.
Landmann, G., M. Bonneau and M. Adrian. 1987. Le dépérissement du sapin pectiné et de lépicéa commun est-il en relation avec létat nutritionnel des peuplements? Rev. For. Fr. 1:5--11.
Lange, O.L., R.M. Weikert, M. Wedler, J. Gebel and U. Heber. 1989. Photosynthese und Nährstoffversorgung von Fichten aus einem Waldschadensgebiet auf basenarmem Untergrund. Allg. Forstz. 44:55--65.
Lichtenthaler, H.K. 1987. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Meth. Enzymol. 148:350--382. Marschner, H. 1995. Mineral nutrition of higher plants. Harcourt
Brace Jovanovich Publishers. Academic Press, London, 889 p. Marschner, H., M. Häussling and E. George. 1991. Ammonium and
nitrate uptake rates and rhizosphere pH in non-mycorrhizal roots of Norway spruce (Picea abies (L.) Karst.). Trees 5:14--21.
Matsumoto, H., N. Wakiuchi and E. Takahashi. 1969. The suppression of starch synthesis and the accumulation of uridine diphosphoglu-cose in cucumber leaves due to ammonium toxicity. Physiol. Plant. 22:537--545.
Mehne-Jakobs, B. 1994. Auswirkungen eines experimentell induzier-ten Magnesiummangels auf die Zusammensetzung der Chloroplas-tenpigmente bei Fichte (Picea abies (L.) Karst.). Allg. Forst Jagdztg. 165:221--227.
Mehne-Jakobs, B. 1995. The influence of magnesium deficiency on carbohydrate concentrations in Norway spruce needles (Picea abies
(L.) Karst.). Tree Physiol. 15:577--584.
Mehne-Jakobs, B. 1996. Magnesium deficiency treatment causes re-ductions in photosynthesis of well-nourished Norway spruce. Trees 10:293--300.
Mehrer, I. and H. Mohr. 1989. Ammonium toxicity: description of the syndrome in Sinapis alba and the search of its causation. Physiol. Plant. 77:545--554.
Mies, E. and H.W. Zöttl. 1985. Zeitliche Änderung der Chlorophyll-und Elementgehalte in den Nadeln eines gelb-chlorotischen Ficht-enbestandes. Forstw. Cbl. 104:1--8.
Mohr, H. 1994. Stickstoffeintrag als Ursache neuartiger Waldschäden. Spektr. Wiss. 1:48--53.
Nihlgård, B. 1985. The ammonium hypothesis----an additional expla-nation to the forest dieback in Europe. Ambio 14:2--8.
Outlaw, W.H. and J. Manchester. 1979. Guard cell starch concentration quantitatively related to stomatal aperture. Plant Physiol. 64:79--82. Parameswaran, N., S. Fink and W. Liese. 1985. Feinstrukturelle Un-tersuchungen an Nadeln geschädigter Tannen und Fichten aus Waldschadensgebieten im Schwarzwald. Eur. J. For. Pathol. 15:168--182.
Polle, A., T. Otter and B. Mehne-Jakobs. 1994. Effect of magnesium-deficiency on antioxidative systems in needles of Norway spruce (Picea abies (L.) Karst.) grown with different ratios of nitrate and ammonium as nitrogen sources. New Phytol. 128:621--628. Puritch, G.S. and A.V. Barker. 1967. Structure and function of tomato
leaf chloroplasts during ammonium toxicity. Plant Physiol. 42:1229--1238.
Skeffington, R. A. 1990. Accelerated nitrogen inputs----a new problem or a new perspective? Plant Soil 128:1--11.
Soikkeli, S. 1981. Comparision of cytological injuries in conifer need-les from several polluted industrial environments in Finland. Ann. Bot. Fenn. 18:47--61.
Torel, J., J. Cillard and P. Cillard. 1986. Antioxidant activity of fla-vonoids and reactivity with peroxy radical. Phytochemistry 25:383--385.
Ulrich, G. 1987. Stabilität und Resilienz von Waldökosystemen unter dem Einfluß saurer Deposition. Forstarchiv 58:232--239. van Breemen, N., J. Mulder and J.J.M. Grinsven. 1987. Impacts of
van Dijk, H.F.G. and J.G.M. Roelofs. 1988. Effects of excessive ammonium deposition on the nutritional status and condition of pine needles. Physiol. Plant. 73:494--501.
van der Eerden, L.J.M. 1982. Toxicity of ammonia to plants. Agric. Environ. 7:223--235.
von Aufseß, H. 1973. Mikroskopische Darstellung des Verholzungs-grades durch Färbemethoden. Holz Roh- Werk. 31:24--33. Weikert, R.M., M. Wedler, M. Lippert, P. Schramel and O.L. Lange.
1989. Photosynthetic performance, chloroplast pigments, and min-eral content of various needle age classes of spruce (Picea abies) with and without the needle flush: an experimental approach for analysing forest decline phenomena. Trees 3:161--172.
Weissen, F., A. Hambuckers, H.J. van Praag and J. Remacle. 1990. A decennial control of N-cycle in the Belgian Ardenne forest ecosys-tems. Plant Soil 128:59--66.
Ziegler, R. and K. Egle. 1965. Zur quantitativen Analyse der Chloro-plastenpigmente. Beitr. Biol. Pflanz. 41:11--37.