414
UJI APLIKASI BAKTERI
PSEUDOMONAS ALCALIGENES
TERHADAP KANDUNGAN ASAM SALISILAT DAN TOTAL FENOL
DALAM UPAYA MENEKAN PENYAKIT LAYU
FUSARIUM
PADA
TANAMAN TOMAT
I Ketut Widnyana1, Ni Putu Pandawani1, NI GAG Eka Martiningsih1
Jurusan Agroteknologi Fakultas Pertanian Universitas Mahasaraswati Denpasar Corresponding author email: : widnyanaketut@gmail.com
ABSTRACT
Fusarium wilt disease on tomato plants caused by the fungus Fusarium oxysporum f.sp. lycopersici, is an important disease on tomato plants and have resulted in substantial damage to the tomato-producing countries in the world. This study aims to determine the potential rizobakteri P. alcaligenes in controlling Fusarium wilt on tomato plants through the induction of resistance as indicated by the increased content of total phenols and salicylic acid from the tomato plant tissue. Isolates were tested, namely P. alcaligenes obtained through exploration rizorfere Leguminosae and Solanaceae plants are P. alcaligenes KtS1, P. alcaligenes TrN2, P. alcaligenes TmA1.Three isolates of P. alcaligenes applied through seed soaking, root soaking and watering tomato seedlings.The results showed that the application of the suspension isolates of P. alcaligenes TmA1 through immersion tomato seeds can increase the content of total phenols and salicylic acid in tomato plant tissue reached a value of 606% and 262% compared with control plants.
Keywords: induction of resistance, total phenols, salicylic acid
1.
PENDAHULUAN
Penyakit layu Fusarium pada tanaman tomat disebabkan oleh jamur Fusarium oxysporum f.sp. lycopersici, merupakan penyakit penting pada tanaman tomat dan telah mengakibatkan kerusakan yang besar pada berbagai daerah penghasil tomat di dunia (Wibowo, 2005). Pengendalian penyakit layu Fusarium belum berhasil dengan baik karena patogen bersifat tular tanah dan dapat bertahan lama di dalam tanah tanpa adanya tanaman inang, sehingga rotasi tanaman menjadi tidak efektif. Pengendalian penyakit dengan mengaplikasikan fungisida sintetik ke dalam tanah hanya dapat menekan penyakit layu Fusarium untuk beberapa bulan saja (Alabouvette et al.,
1996), selain itu penggunaan fungisida sintetis secara terus-menerus juga dapat menyebabkan munculnya populasi patogen yang lebih tahan dan juga akan mencemari lingkungan (Freeman et al., 2002).
Berbagai upaya telah dilakukan dalam pengendalian penyakit layu fusarium, diantaranya penggunaan benih sehat, rotasi tanaman, tumpang sari dan dengan pestisida (fungisida), tetapi tidak selalu memberikan hasil yang memuaskan. Alternatif lain untuk mengendalikan penyakit layu Fusarium adalah dengan memanfaatkan mikroba agen pengendali hayati. Pengendalian dengan cara ini dilaporkan cukup efektif dan belum ada yang melaporkan timbulnya ketahanan jamur patogen terhadap agen pengendali hayati (Freeman et al., 2002).
Mikroba yang bersifat menguntungkan bagi tanaman, seperti rizobakteri dari kelompok Pseudomonas spp. dapat berfungsi sebagai penyubur, sebagai sarana pengendali hayati patogen tanaman dan mampu meningkatkan ketahanan tanaman (induced systemic resistance (ISR) (McMilan, 2007). Hasil penelitian sebelumnya menunjukkan bahwa Pseudomonas spp. dapat meningkatkan hasil dan melindungi tanaman gandum dari patogen Phytium spp. melalui perlakuan benih (Weller dan Cook, 1986); melindungi tanaman kacang tanah dari patogen layu dan akar Sclerotium rolfsii (Ganesan dan Gnanamanickam, 1986) dan kacang polong dari patogen
F. oxysporum f.sp. fisi dan Phytium ultimun (Benhamou et al., 1996); dapat menginduksi ketahanan tanaman mentimun terhadap serangan patogen Colletotrichumorbiculare melalui perlakuan pada akar benih (Wei et al.,
415
Pseudomonas spp. yang diisolasi dari rizosphere tanaman kacang-kacangan dan terung-terungan dalam menginduksi ketahanan tanaman tomat terhadap Fusarium oxysporum f.sp. lycopersici yang diindikasikan dengan peningkatan kadar total fenol dan asam salisilat dalam jaringan tanaman tomat.
2.
METODE PENELITIAN
Isolasi jamur F. oxysporum f.sp. lycopersici
Isolat F. oxysporum f.sp. lycopersici diperoleh dari batang tanaman tomat terserang penyakit layu Fusarium. Bahan tanaman tersebut sebelumnya telah didesinfeksi dengan alkohol 70% dan dibilas dengan air steril, kemudian dipotong-potong dengan ukuran + 1 cm dan ditanam pada media PDA yang ditempatkan dalam cawan Petri. Pemurnian dilakukan apabila terdapat mikroba lain yang tumbuh bersama jamur target. Untuk memastikan bahwa jamur tersebut adalah patogen layu Fusarium maka dilakukan pengujian dengan mengikuti prosedur Postulat Koch. Inkubasi dilakukan pada suhu kamar, dalam waktu 3 hari spora jamur patogen sudah tumbuh dan dapat dipergunakan sebagai sumber inokulum.
Jamur Fusarium membentuk koloni berwarna putih pada media PDA dan apabila sudah membentuk spora koloni akan bewarna putih keabu-abuan. Pengamatan di bawah mikroskop dilakukan untuk melihat ciri morfologinya yaitu adanya mikrokonidia berbentuk bulat telor dan makrokonidia berbentuk bulan sabit dan bersepta.
Isolasi Rizobakteri Pseudomonas spp.
Isolat bakteri diperoleh dari koleksi peneliti yang sebelumnya sudah diuji potensinya sebagai perangsang tumbuh tanaman tomat yaitu isolat P.alcaligenes KtS1, TrN2, dan TmA1. Isolat P.alcaligenes KtS1 didapatkan pada rizosfer tanaman kacang tanah dari kabupaten Buleleng, Isolat P.alcaligenes TrN2 didapatkan pada rizosfer tanaman terung dari kabupaten Jembrana, dan isolat P.alcaligenes TmA1 didapatkan dari rizosfer tanaman tomat dari kabupaten Karangasem (widnyana et al., 2013). Isolat Pseudomonas tersebut disimpan pada frezer suhu 5oC dalam media cair dan diremajakan setiap 6 bulan sekali. Satu mililiter suspensi dituangkan ke dalam 10 ml media Kings'B, dan diinkubasi selama 48 jam pada suhu kamar.
Isolasi bakteri antagonis dengan mempergunakan media King’sB dimaksudkan untuk tetap menjaga isolat bakteri yang mampu menghasilkan figment warna fluorescens pada media tersebut yaitu dari kelompok
Pseudomonas pendarflour (Fox, 1993). Spesies bakteri yang menghasilkan pigment warna flouerescens di dalam media King’s B agar diantaranya adalah P. fluorescens, P. putida , dan P. aeruginosa.
Persiapan media dan aplikasi isolat P.alcaligenes pada tomat
Pengujian potensi rizobakteri Pseudomonas spp. sebagai agens pengendali hayatidilakukan di rumah kaca, diawali dengan menyiapkan media tanam untuk tanaman tomat. Media tanam dibuat dari campuran tanah dan kompos (5:2) yang sudah dioven untuk meminimalkan kontaminasi. Pengovenan media tanam dilakukan selama 2 jam pada suhu minimal 600C (Widnyana, 2011). Media tanam yang sudah dioven dimasukkan ke dalam polybag, masing-masing sebanyak 4 kg.
Perbanyakan patogen secara massal dilakukan pada media PDA dalam cawan Petri. Biakan murni jamur
P.oxysporum f.sp lycopersici ditumbuhkan dalam cawan Petri (sesuai kebutuhan), dan setelah pertumbuhan jamur memenuhi cawan petri (8 hari) kemudian dilarutkan dalam air steril untuk memisahkan konidianya.
416
Suspensi konidia diaplikasikan dengan menyiramkan pada pot perlakuan sebanyak 20 ml/tanaman dan mencapai kepadatan 104 konidia/ml (Mathiyazhagana, 2004). Konsentrasi dari bakteri Pseudomonas spp. yang digunakan adalah 10 8 cfu/ml, sebelumnya bakteri ditumbuhkan selama 48 jam pada media King’s B agar dalam suhu kamar (Widnyana, 2011).
Pengujian potensi isolate P.alcaligenes sebagai agens pengendali hayati dilakukan dengan cara mengaplikasikanya pada tanaman tomat melalui perendaman benih, pencelupan akar, dan penyiraman bibit, yaitu masing-masing benih direndam selama 30 menit, akar direndam selama 10 menit, dan bibit umur 2 minggu pada pesemaian disiram merata dengan isolat P.alcaligenes umur 48 jam pada media PD broth.
Uji potensi penginduksi ketahanan tanaman melalui analisis kandungan asam salisilat dan total fenol
Analisis jaringan tanaman dilakukan di laboratorium Balai Besar Pasca Panen Bogor, yang bertujuan untuk mengetahui adanya perubahan terhadap kandungan substansi kimia tertentu yang berkaitan dengan fungsi ketahanan tanaman terhadap patogen setelah dilakukan perlakuan induksi ketahanan sistemik dengan suspensi isolat P.alcaligenes Analisis dilakukan terhadap kandungan dua substansi kimia yaitu Asam Salisilat dan Total Fenol. Analisis terhadap kandungan ketiga substansi kimia tersebut dilakukan dengan metode HPLC (High Performance Liquid Chromatografi)
Tanaman tomat yang akan dianalisis jaringannya adalah tanaman yang sudah diberikan perlakuan isolat bakteri P.alcaligenes yaitu pada pengujian potensi sebagai pengendali hayati dan PGPR. Pada setiap uji jaringan digunakan 3 unit tanaman tomat yang diambil bagian batang dan daunnya masing-masing 10g, kemudian bagian tersebut digabung menjadi satu unit sampel yang disiapkan untuk analisis jaringan dengan berat 50 g bagian tanaman per sampel.
Hasil analisis terhadap kandungan ketiga substansi kimia tersebut juga dibandingkan secara kuantitatif antara kontrol (tanpa perlakuan induksi) dengan perlakuan induksi dengan isolat bakteri P.alcaligenes
3.
HASIL PENELITIAN
Analisa total fenol dan asam salisilat terhadap jaringan tanaman tomat (bagian akar, batang, dan daun) dilakukan di Laboratorium Balai Besar Pasca Panen di Bogor. Hasil analisa kandungan total fenol dan kandungan asam salisilat disajikan pada Tabel 1.
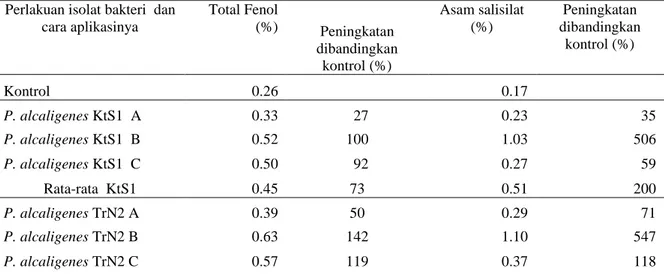
Tabel 1. Kandungan total fenol dan asam salisilat jaringan tanaman tomat (%) setelah diaplikasikan dengan isolat bakteri P. alcaligenes
Perlakuan isolat bakteri dan cara aplikasinya Total Fenol (%) Peningkatan dibandingkan kontrol (%) Asam salisilat (%) Peningkatan dibandingkan kontrol (%) Kontrol 0.26 0.17 P. alcaligenes KtS1 A 0.33 27 0.23 35 P. alcaligenes KtS1 B 0.52 100 1.03 506 P. alcaligenes KtS1 C 0.50 92 0.27 59 Rata-rata KtS1 0.45 73 0.51 200 P. alcaligenes TrN2 A 0.39 50 0.29 71 P. alcaligenes TrN2 B 0.63 142 1.10 547 P. alcaligenes TrN2 C 0.57 119 0.37 118
417
Keterangan : A = perendaman akar bibit, B = perendaman benih, C = penyiraman pesemaian bibit
Tabel 1, menunjukkan bahwa peningkatan kandungan total fenol yang paling tinggi pada ketiga jenis isolat
P.alcaligenes (TmA1, TrN2, da KtS1) terjadi pada perlakuan aplikasi bakteri melalui perendaman benih (B), kemudian diikuti dengan perlakuan penyiraman pesemaian bibit (C) dan pencelupan akar (A). Kandungan total fenol pada perlakuan TmA1B meningkat 262% dibandingkan dengan kontrol, pada TrN2B meningkat 142%, dan pada KtS1B meningkat 100% disbanding kontrol. Peningkatan kandungan tertinggi ke dua terjadi pada perlakuan isolat bakteri P.alcaligenes melalui penyiraman pesemaian bibit tomat dan peningkatan yang paling rendah terjadi pada pencelupan akar. Kandungan total fenol tertinggi sebagai akibat dari perlakuan isolat
P.alcaligenes terjadi pada isolat P.alcaligenes TmA1, diikuti dengan P.alcaligenes TrN2, dan P.alcaligenes
KtS1.
Kandungan asam salisilat yang paling tinggi terdapat pada perlakuan P.alcaligenes TmA1 dengan rata-rata 0.65%, diikuti dengan perlakuan TrN2 (0.59 %) dan perlakuan KtS1 (0.51%). Kandungan asam salisilat tertinggi dari semua kombinasi perlakuan yang dilakukan terdapat pada perlakuan TmA1B (1.20%), diikuti dengan TrN2A (1.10% ) dan KtS1A (1.03%)
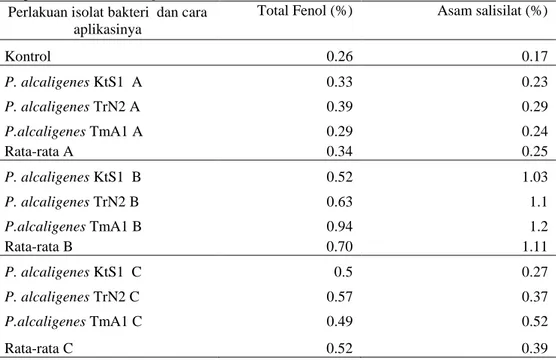
Tabel 2. Kandungan total fenol dan asam salisilat jaringan tanaman tomat (%) dikelompokkan berdasarkan cara aplikasi isolat P.alcaligenes
Perlakuan isolat bakteri dan cara aplikasinya
Total Fenol (%) Asam salisilat (%)
Kontrol 0.26 0.17 P. alcaligenes KtS1 A 0.33 0.23 P. alcaligenes TrN2 A 0.39 0.29 P.alcaligenes TmA1 A 0.29 0.24 Rata-rata A 0.34 0.25 P. alcaligenes KtS1 B 0.52 1.03 P. alcaligenes TrN2 B 0.63 1.1 P.alcaligenes TmA1 B 0.94 1.2 Rata-rata B 0.70 1.11 P. alcaligenes KtS1 C 0.5 0.27 P. alcaligenes TrN2 C 0.57 0.37 P.alcaligenes TmA1 C 0.49 0.52 Rata-rata C 0.52 0.39
Keterangan : A = perendaman akar bibit, B = perendaman benih, C = penyiraman pesemaian bibit
Tabel 2, menunjukkan bahwa kandungan total fenol tertinggi terdapat pada perlakuan aplikasi isolat
P.alcaligenes melalui perendaman benih (B) yaitu rata-rata 0.70%, diikuti dengan perlakuan penyiraman pesemaian (C) dengan rata-rata 0.52% dan perlakuan pencelupan akar bibit (A) yaitu rata-rata 0.34%. hal yang
Rata-rata TrN2 0.53 103.67 0.59 245.33
P. alcaligenes TmA1 A 0.29 12 0.24 41
P. alcaligenes TmA1 B 0.94 262 1.20 606
P. alcaligenes TmA1 C 0.49 88 0.52 206
418
sama juga terjadi pada kandungan asam salisilat yaitu rata-rata kandungan asam salisilat tertinggi terdapat pada perlakuan B (1.11%), diikuti dengan perlakuan C (0.39%), dan perlakuan A (0.25%).
Ketahanan tanaman dapat diinduksi dengan inokulasi mikroba non patogenik, dan perlakuan kimia tertentu. Patogen menstimulasi pembentukan fitoaleksin dalam jumlah relatif sedikit dibandingkan dengan induksi yang dilakukan oleh non-patogenik. Ketahanan systemik terinduksi pada berbagai tanaman yang disebabkan oleh agens penginduksi mikroba maupun senyawa kimia ditunjukkan dengan peningkatan kandungan senyawa-senyawa tertentu, seperti enzim -1.3. glukanase, kitinase, -1.4 glukosidase, citonase, peroksidase, fenol dan asam salisilat (Maurhofer et al., 1994; Wei et al., 1996)
Analisis regresi antara kandungan total fenol pada jaringan tanaman tomat dengan persentase serangan penyakit layu menunjukkan nilai R2 = 0.558** (sangat nyata) dengan persamaan regresi Y = 104.81-135.51X, dimana X adalah kandungan total fenol jaringan tanaman. Hal ini menunjukkan bahwa adanya kandungan total fenol sangat mempengaruhi ketahanan tanaman tomat terhadap penyakit layu Fusarium dengan asumsi bahwa 55.8% ketahanan tanaman tomat disebabkan oleh adanya kandungan fenolik dalam jaringan tanaman. Apabila kandungan total fenol mencapai 0,8% saja maka ada kemungkinan dapat menekan penyakit layu Fusarium sampai 100%.
Analisa regresi antara kandungan asam salisilat dengan persentase serangan penyakit layu Fusarium
menunjukkan nilai R2 = 0.47* (nyata), dengan persamaan regresi Y = 74.024 – 63.82X, dimana X adalah kandungan asam salisilat pada jaringan tanaman tomat. Hal ini berarti dapat diasumsikan bahwa sebanyak 47.2% ketahanan tanaman tomat disebabkan oleh adanya kandungan asam salisilat dalam jaringan tanaman tomat, dimana apabila kandungan asam salisilat mencapai 1% maka peluang persentase penyakit yang terjadi adalah sebesar 10.2% pada minggu IX. Hal ini sesuai dengan pendapat De Meyer dan Hofte (1997) dan Maurhofer et al., (1998) yang menyatakan bahwa asam salisilat adalah metabolit Pseudomonas yang terindikasi memacu peningkatan ketahanan. Telah diketahui secara luas bahwa pelakuan asam salisilat pada tanaman menyebabkan stimulasi ketahanan (Sticher et al., 1997). Namun hampir semua penelitian mengenai peran asam salisilat yang diproduksi bakteri dalam stimulasi ketahanan menyimpulkan bahwa bukan asam salisilat saja yang dimiliki sinyal mikroba (Audenaert et al., 2002; Ran et al., 2005b). Biosintesis asam salisilat sering dihubungkan dengan produksi asam salisilat yang mengandung siderophore, seperti pyochelin pada P.aeruginosa (Audenaert et al.,
2002) atau pseudomonine pada P. fluorescens (Mercado-Blanco et al., 2001).
4.
KESIMPULAN
1. Aplikasi isolat P.alcaligenes terbukti mampu meningkatkan ketahanan tanaman tomat terhadap penyakit layu Fusarium yang diindikasikan dengan meningkatnya kandungan asam salisilat dan total fenol dalam jaringan tanaman tomat yang sebelumnya diaplikasikan dengan isolat P.alcaligenes KtS1, TrN2, dan TmA1.
2. Peningkatan kandungan total fenol yang paling tinggi pada ketiga jenis isolat P.alcaligenes terjadi pada perlakuan aplikasi bakteri melalui perendaman benih (B), kemudian diikuti dengan perlakuan penyiraman pesemaian bibit (C) dan pencelupan akar (A).
3. Kandungan asam salisilat yang paling tinggi terdapat pada perlakuan P.alcaligenes TmA1 dengan rata-rata 0.65%, diikuti dengan perlakuan TrN2 (0.59 %) dan perlakuan KtS1 (0.51%).
419
DAFTAR PUSTAKA
Alabouvette, R., P. Lemanceau, and C. Steinberg. (1996). Biological Control of Fusarium Wilts: Opportunities for Developing A Comercial Product.
Audenaert, K.T. Pattery, P. Cornelis, and M. Hofte. (2002). Induction of systemic resistance to Botrytis cinerea
in tomato by Pseudomonas aeruginosa 7NSK2: role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant-Microbe Interact. 15:1147–1156.
Benhamou, N., R.R. Belanger, and T.C. Paulitz. (1996). Induction of differential host responses by
Pseudomonas fluorescens in Ri T-DNA-transformed pea roots after challenge with Fusarium oxysporum
f.sp. pisi and Phytium ultimum. Phytopathology 86:1174-1185.
De Meyer, G. and M. Hofte. (1997). Salicylic acid produced by the rhizobacterium P. aer,uginosa 7NSK2 induces resistance to leaf infection by Botrytis cinerea on bean. Phytopathology87:588-593.
Fox, R.T.V. (1993). Principles of diagnostic techniques in plant pathology. International Mycological Institute. An institute of Cab International. Cambridge University Press. UK.
Freeman S., A. Zveibel, H. Vintal and M. Maymon. (2002). Isolation of nonpatogenic mutants of Fusarium oxysporum f.sp. melonis for biological kontrol of Fusarium wilts in cucurbits. Phytopathology 92:164-168.
Ganesan, P., and S.S. Gnanamanickam. (1986). Biological kontrol of Sclerotium rolfsii Sacc. In peanut by inoculation with Pseudomonas fluorescens. Centre of advanced study in Botany, University of Madras, India.
Maurhofer, M., C.Hase, P.Meuwly, J.P. Metraux, and G.Defago. (1994). Induction of systemic resistance of tobacco to tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens strain CHA0: influence of the gacA gene and of pyoverdine production. Phytopathology 84:139-146.
McMilan, S. (2007). Promoting Growth with PGPR. The Canadian Organic Grower. Soil Foodweb Canada Ltd. Soil Biology Lab. & Learning Centre
Mercado-Blanco, J., K.M.G.M. van der Drift, P.E. Olsson, J.E. Thomas-Oates, L.C. van Loon, and P.A.H.M. Bakker. (2001). Analysis of the pmsCEAB gene cluster involved in biosynthesis of salicylic acid and the siderophore pseudomonine in the biocontrol strain Pseudomonas fluorescens WCS374. J.Bacteriol. 183:1909–1920.
Ran, L.X., L.C. Van Loon, and P.A.H.M. Bakker. (2005b). No role for bacterially produced salicylic acid in rhizobacterial induction of systemic resistance in Arabidopsis. Phytopathology 95:1349–1355.
Sticher, L., B. Mauch-Mani, and J.P. Me´traux. (1997). Systemic acquired resistance. Annu. Rev. Phytopathol. 35:235–270.
Weller, D.M., and R.J.Cook. (1986). Increased growth of wheat by seed treatments with fluorescens pseudomonads, and implications of Phytium kontrol. Can.J. Plant. Pathol. 8:328-334.
Wibowo, A. (2005). Colonization of tomato root by antagonistik bacterial strains to fusarium wilt of tomato. Department of entomology and phytopathology, Faculty of agriculture, Gajah Mada. Jurnal Perlindungan Tanaman Indonesia, 11(2) : 66-76.
Widnyana, I.K. (2011). Upaya mendapatkan agens biokontrol penyakit layu tomat. Fusarium oxysporum f.sp
lycopersici melalui eksplorasi dan uji potensi PGPR isolat bakteri Pseudomonas spp. Jurnal Lingkungan Hidup Bumi Lestari 11 (2) : 266 – 275.
Widnyana, I.K., Dewa Ngurah Suprapta, I Made Sudana, I Gede Rai Maya Temaja. (2013). Pseudomonas alcaligenes, Potential Antagonist Against Fusarium oxysporum f.sp.lycopersicum the Cause of Fusarium